
Insetti
Classe del phylum Artropodi, collocata, in seguito alle revisioni sistematiche degli ultimi decenni del 20° sec., nella superclasse degli Esapodi. Sono considerati il gruppo più ricco di forme esistente sulla Terra: le specie descritte sono oltre un milione e il loro numero aumenta con la continua scoperta di nuove specie.
Diffusione
Gli I. compaiono nell’Era paleozoica e le prime testimonianze della loro presenza si trovano nel Devoniano medio con frammenti della specie, probabilmente alata, Rhyniognatha hirsti, inizialmente classificata come collembolo. Alla fine del Carbonifero sono già presenti i Blattoidei, gli Efemerotteri, gli Odonati, gli Ortotteri; nel Permiano compaiono molti degli ordini odierni di I. Neotteri: Coleotteri, Neurotteri, Mecotteri, Psocotteri, mentre molti dei gruppi più antichi scompaiono nel corso della grande estinzione del Permiano-Triassico. Nell’Era mesozoica si diffondono i Ditteri, i Lepidotteri e i Tricotteri, comparsi con forme primitive alla fine del Paleozoico. A partire dal Cretaceo inferiore, un gran numero dei moderni gruppi di successo si è evoluto parallelamente (➔ coevoluzione) alle Angiosperme, le piante da fiore, che a loro volta hanno subito un incremento esponenziale di numero. Nell’Era cenozoica si diffondono i primi gruppi sociali di Imenotteri (comparsi già nel Cretaceo).
Gli Insetti, dotati di una corazza (esoscheletro) che ha loro permesso di evolversi in infinite varietà di forme, e forniti di istinti affinati, sensibili organi di senso, capacità di adattamento a ogni tipo di ambiente, occupano sulla Terra una posizione ‘dominante’ rispetto agli altri gruppi animali, e condizionano notevolmente l’equilibrio generale della natura.
La durata della vita degli I., se si fa eccezione per alcuni taxa sociali e pochi altri, è generalmente breve: la gran parte vive pochi mesi, poche settimane o pochi giorni. Invece la durata del periodo preimaginale può prolungarsi anche per parecchi anni. Si diffondono (attivamente o passivamente) in modo straordinario, compiendo talora, anche in masse di milioni di individui, migrazioni che sorvolano continenti e oceani. Vivono solitari, in gruppo, o in società (omogenee o eterogenee; stagionali o persistenti; ecc.) di notevole complessità, e spesso in simbiosi (con piante e animali) a 2, 3 o più simbionti.
Morfologia esterna e anatomia
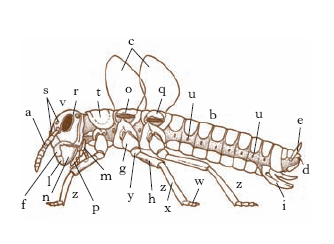
Regioni morfologiche. - Il corpo degli I. ha forma e dimensioni varie, ma è sempre diviso in tre regioni morfologiche o tagmi: testa, torace e addome, ognuna costituita da un certo numero di segmenti (metameri o somiti) più o meno modificati (fig. 1).

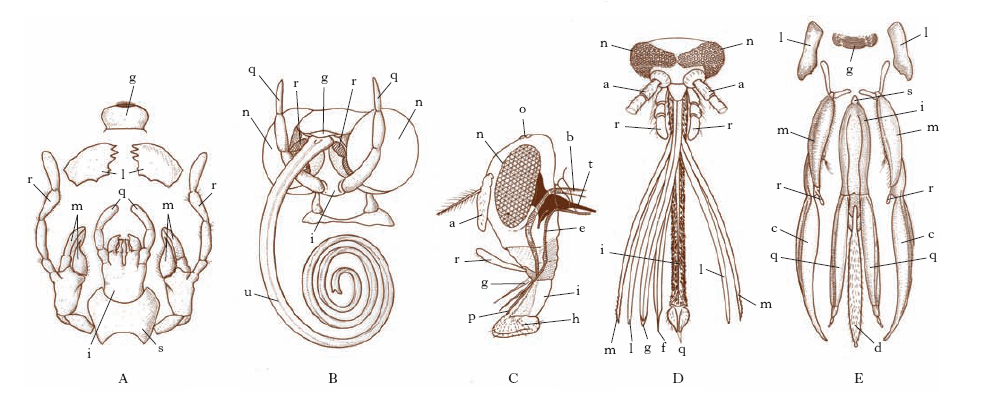
La testa è composta da 6 metameri fusi insieme a formare una capsula rigida, il cranio. I primi 3 segmenti compongono il procefalo, fornito tipicamente di 2 antenne (generalmente pluriarticolate e più o meno ricche di organi di senso; fig. 2), 2 occhi composti e 3 o 2 ocelli; dal primo somite deriva anche il labbro superiore; i successivi 3 somiti del capo, costituenti lo gnatocefalo, sono modificati nelle appendici boccali. L’apparato boccale è tipicamente composto dal labbro superiore, dalle appendici boccali (2 mandibole, il primo paio di mascelle e il secondo paio di mascelle riunite insieme a formare il labbro inferiore) e dalla prefaringe; esso però subisce, in correlazione con particolari regimi alimentari propri delle varie specie e dei vari stadi di vita, profonde modificazioni: si hanno così vari tipi di apparati boccali (fig. 3), come il masticatore tipico, il masticatore lambente, il succhiante, il pungente succhiante ecc.
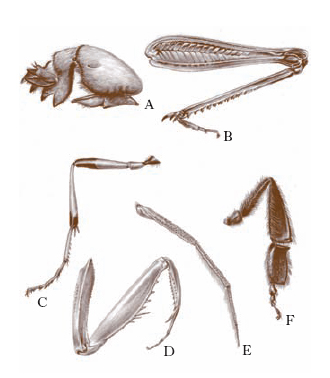
Il torace è composto da tre metameri: protorace, mesotorace e metatorace, ognuno dei quali presenta una regione dorsale (tergite o noto), due laterali (pleure) e una ventrale (sterno), suscettibili di ulteriori suddivisioni in scleriti. È provvisto di tre paia di appendici metameriche, le zampe (fig. 4), costituite da sei segmenti (anca o coxa, trocantere, femore, tibia, tarso e pretarso, sul quale si articolano le unghie) e, a eccezione di Archeognati e Tisanuri, di 1 o 2 paia di espansioni laterali (su mesotorace e metatorace), le ali, variamente sviluppate e conformate, talora fornite di apparati di collegamento, percorse da un complesso di tubuli sclerificati, le venature o nervature, che hanno grande importanza per il volo e che contengono trachee, nervi ed emolinfa.
L’addome è composto da un numero variabile di metameri (comunemente 11) detti uriti; essi sono assai meno differenziati di quelli toracici e provvisti, a seconda dei gruppi e degli stadi, di processi e di appendici varie (stili, cerci, pseudozampe, tracheobranchie ecc.).
Sistema tegumentale. - È costituito, procedendo dall’esterno all’interno, dalla cuticola o esoscheletro, dall’epidermide e dalla membrana basale. L’esoscheletro, secreto dall’epidermide, è pluristratificato, inerte e rigido, riveste interamente il corpo degli I. e si prolunga all’interno con dei processi, detti apodemi, di varia forma ed estensione, che costituiscono l’endoscheletro, apparato di rinforzo, di attacco per i muscoli e di sostegno per i visceri. Nelle giunture tra i somiti e le articolazioni delle appendici la cuticola è più sottile e flessibile, rendendo possibili i movimenti. Per permettere l’accrescimento, la vecchia cuticola viene abbandonata e ne viene elaborata una nuova nel processo della muta. I colori, anche molto vivaci, che ornano il corpo degli I. possono essere strutturali o pigmentali. I primi sono il risultato dell’azione della luce su microstrutture cellulari; i secondi della presenza (nella cuticola, nell’epidermide, nel sangue, nel tessuto adiposo ecc.) di pigmenti vari.
Sistema muscolare. - I muscoli degli I. sono quasi tutti formati da fibre striate. La loro disposizione generale, il loro numero e il loro comportamento variano da gruppo a gruppo.
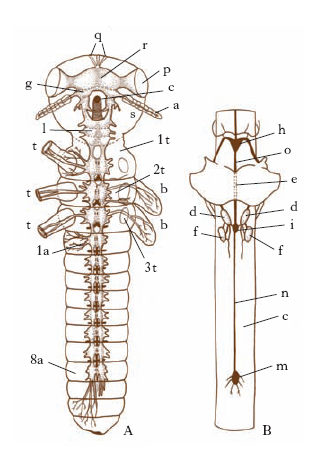
Sistema nervoso. - Comprende un sistema centrale, uno viscerale e uno periferico (fig. 5). Il sistema centrale è costituito da una massa gangliare sopraesofagea (cervello o cerebro), contenuta di regola nel cranio, da una sottoesofagea (gnatocerebro o gnatencefalo), collegata alla precedente da due connettivi paraesofagei, e da una catena ganglionare ventrale, formata da due serie adiacenti di gangli segmentali, riuniti tra loro longitudinalmente da connettivi e trasversalmente da commessure.
Il sistema viscerale ha funzioni neurovegetative, e comprende un simpatico dorsale (stomatogastrico), che innerva la porzione anteriore del sistema digerente e il cuore, e un simpatico ventrale che innerva trachee, stigmi, proctodeo e organi genitali.
Il sistema periferico è costituito da un plesso di neuroni bipolari e multipolari collegati con il sistema centrale, alcuni dei quali mandano i loro processi distali a innervare il tegumento, i muscoli somatici ecc., mentre altri li inviano agli organi di senso.
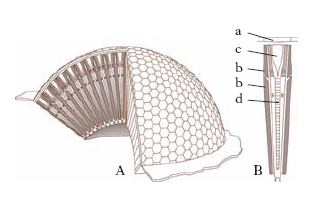
Organi di senso o sensilli. - Esclusi gli occhi e gli ocelli, dalla struttura particolare, i sensilli sono costituiti tipicamente da un apparato esterno (in genere un’appendice tegumentale cava), da un numero vario di neuroni, da due o tre cellule epidermiche specializzate (cellule accessorie) e da un corpo sensoriale. A seconda della loro funzione, gli organi di senso si distinguono in chemiorecettori olfattivi, chemiorecettori gustativi, termorecettori, igrorecettori, esterorecettori (tattili), propriorecettori, fonorecettori e fotorecettori. Questi ultimi (occhi e ocelli) differiscono dagli altri per la mancanza di apparato esterno e perché presentano esternamente una cuticola trasparente. Gli occhi composti (fig. 6), che percepiscono le immagini, sono costituiti da un numero variabile di gruppi di cellule fotorecettrici (dette ommatidi), posseggono un apparato diottrico e un apparato catottrico per ogni ommatidio; gli ocelli, che percepiscono variazioni di intensità della luce, hanno un solo apparato diottrico per tutte le cellule sensoriali, e possono avere posizione dorsale o laterale.
Organi e apparati di produzione di suoni. - I suoni emessi dagli I. possono essere determinati da colpi o strofinii contro superfici circostanti, dallo sfregamento reciproco di parti del corpo, dalla vibrazione di membrane speciali.
Organi emananti luce. - Alcuni I. debbono la loro luminescenza a batteri fosforescenti, altri a organi luminosi propriamente detti, variamente distribuiti e nei quali la luminescenza è il risultato dell’azione di due sostanze: la luciferina e la luciferasi, di cui la seconda agisce come un enzima del gruppo delle ossidasi (➔ bioluminescenza).
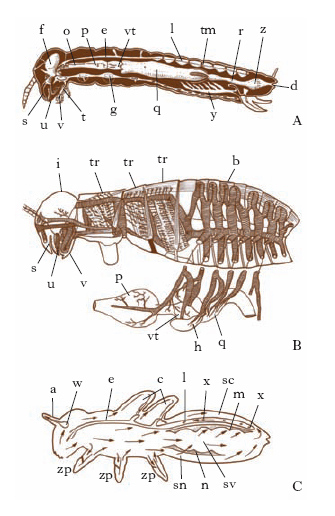
Sistema digerente. - Comprende gli organi di presa e ingestione (labbro superiore, appendici boccali, prefaringe ecc.) e il tubo digerente (fig. 7A). Questo è costituito da: a) una porzione anteriore, lo stomodeo, invaginazione ectodermica cefalica, che può essere differenziato in faringe, ingluvie, ventriglio, valvo;la cardiaca e che ha funzione di ingestione e prima elaborazione dell’alimento; b) una porzione mediana, il mesentero (o mesentere), di origine probabilmente endodermica, che è la sede delle principali funzioni della digestione e dell’assorbimento, e il cui epitelio produce una membrana (membrana peritrofica) che avvolge il cibo e funziona come un diaframma dializzante; c) una porzione posteriore, il proctodeo, invaginazione ectodermica caudale, che può essere differenziato in valvola pilorica, ileo, colon, retto, ampolle e papille rettali ecc., ed è deputato al riassorbimento dell’acqua e all’eliminazione dei cataboliti e delle parti non digerite. I regimi alimentari degli I. sono straordinariamente vari e passano dalla pantofagia alle specializzazioni più estreme.
Sistema respiratorio. - La respirazione si effettua, nella grande maggioranza dei casi, mediante un apparato tracheale (fig. 7B) più o meno differenziato. In quasi tutte le specie terrestri e alcune acquatiche l’aria penetra nelle trachee, a loro volta anastomizzate in tracheole, per un numero vario di aperture pari (stigmi), mentre l’anidride carbonica viene eliminata lungo le medesime vie attraverso il tegumento. In altre specie e stadi larvali acquatici (Efemerotteri, Odonati, Plecotteri), invece, la respirazione avviene mediante tracheobranchie (estroflessioni filamentose o foliari del tegumento, a pareti sottilissime, contenenti ramificazioni tracheali). In alcune specie infine (e particolarmente tra gli endoparassiti) manca completamente l’apparato tracheale (I. apneustici) e la respirazione avviene attraverso il tegumento. A parte quest’ultimo caso, nella grande maggioranza degli I. l’emolinfa non interviene nel trasporto dell’ossigeno e l’apparato tracheale porta direttamente l’ossigeno alle cellule.
Sistema circolatorio. - Negli I. la circolazione è in parte vascolare (vaso dorsale e centri contrattili accessori, con funzione cardiaca) e in gran parte lacunare (fig. 7C), poiché il sangue o emolinfa compie il più del suo percorso fuori dei vasi, liberamente fra organi e tessuti, nella cavità del corpo (lacunoma o emocele) che è divisa più o meno completamente in seni superficiali e profondi da diaframmi.
Sistema escretore. - I più importanti organi escretori degli I. sono i tubi malpighiani in numero vario (da 2 a più di 200) con l’estremità distale chiusa e la prossimale aperta nella regione mesentero-proctodeale, che assorbono dall’emolinfa i prodotti catabolici e li eliminano attraverso il tubo digerente. Oltre ai tubi malpighiani, presentano attività escretrici anche altri organi e tessuti: nefrociti, corpo adiposo, tegumento ecc.
Sistema secretore. - a) Nel corpo degli I. si trovano numerose ghiandole esocrine che secernono sostanze varie, utilizzate in diversi modi (cera, lacca, seta, muco, sostanze repellenti, urticanti, velenose, adesive ecc.) e anche feromoni (ghiandole a feromoni). b) L’apparato endocrino è rappresentato dai corpora allata, dai corpora cardiaca (o pharyngea), dalle ghiandole protoraciche e da vari territori neurocrini (del cerebro, dello gnatocerebro, dei gangli della catena nervosa ventrale ecc.). Gli ormoni elaborati da tali strutture (per es. l’ecdisone, la neotenina o ormone giovanile ecc.) hanno un’enorme importanza, in quanto regolano lo sviluppo postembrionale, le mute, le metamorfosi, le diapause, la riproduzione ecc.
Sistema riproduttore. - Gli I. sono animali tipicamente gonocorici (a sessi distinti). Possono tuttavia presentarsi casi di ginandromorfismo, di intersessualità e di ermafroditismo. Il sistema riproduttore deriva in parte dall’ectoderma e in parte dal mesoderma e comprende vari organi: le gonadi (testicoli e ovari), i gonodotti (deferente e ovidutto), i genitali esterni (organo di copulazione o edeago e di ovideposizione o ovopositore). A queste parti principali se ne aggiungono altre accessorie: vescicole seminali e ghiandole accessorie per il maschio; calici ovarici, ghiandole colleteriche, borsa copulatrice, spermateca ecc. per la femmina. L’apparato riproduttore sbocca ventralmente (o anteriormente) rispetto all’intestino (generalmente nell’8° e nel 9° urosternite). Lo sbocco è unico, raramente doppio. Le femmine dei Lepidotteri hanno una seconda apertura genitale, anteriore a quella tipica, che funziona come ostio di copulazione. In alcuni gruppi l’ovoposizione è svolta da un organo di origine estranea all’apparato riproduttore, detto ovopositore di sostituzione.
Riproduzione e sviluppo
La forma di riproduzione più comune fra gli Insetti è quella anfigonica. Abbastanza frequente è però la partenogenesi. Più rare sono la pedogenesi e la poliembrionia. La fecondità degli I. varia ampiamente: vi sono specie che depongono poche uova, altre capaci di emetterne centinaia, migliaia o milioni. Gli I. possono essere ovipari, ovovivipari e vivipari. Le cure parentali e le attività di protezione delle uova sono molto frequenti e, spesso, complesse.
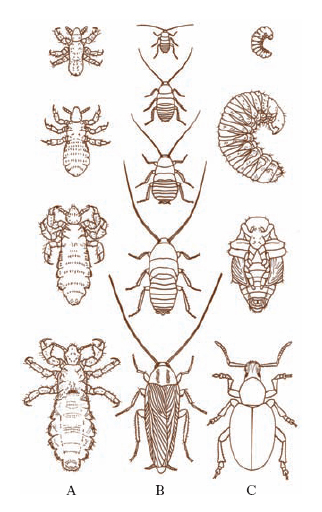
Lo sviluppo comprende quello embrionale e quello postembrionale (fig. 8). Quest’ultimo è sempre discontinuo e si effettua attraverso ripetuti rinnovamenti della cuticola (mute), cui si aggiunge un complesso più o meno notevole di trasformazioni, che prende il nome di metamorfosi. L’ultimo stadio della vita postembrionale degli I. è detto i. perfetto o imago, oltre che immagine o imagine, e corrisponde allo stato adulto caratterizzato dalla maturità sessuale e dall’attività riproduttiva. La forma più tipica di sviluppo è l’olometabolia (metamorfosi completa), in cui l’insetto passa per gli stadi di larva (bruco nelle farfalle), pupa o ninfa (crisalide nelle farfalle) e imagine. Vi sono I. ametaboli (a metamorfosi appena accennate), eterometaboli (a metamorfosi radicali), catametaboli (a metamorfosi regressive).
Classificazione
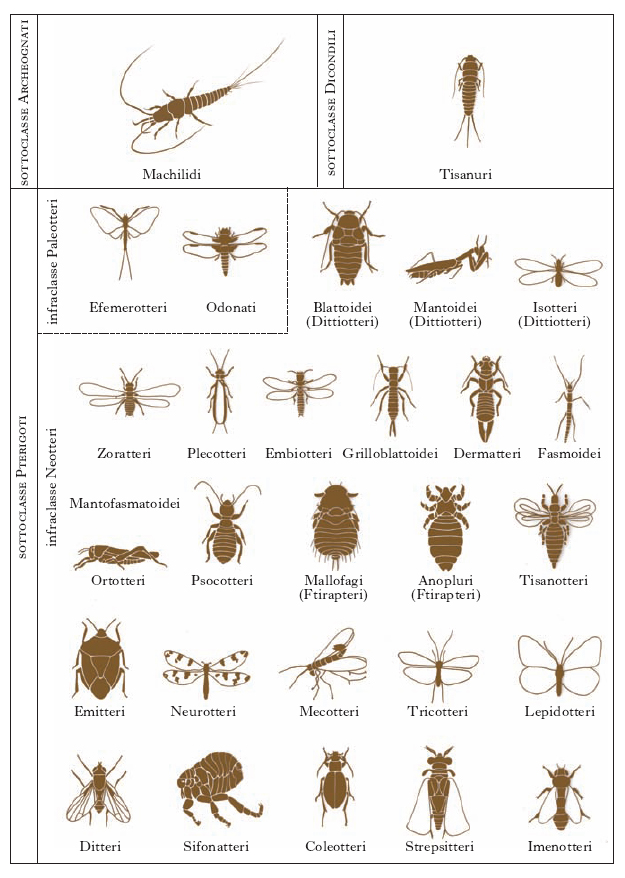
La classificazione degli I. è oggetto di analisi e discussione e subisce frequenti rimaneggiamenti. Nella classificazione tradizionale, gli I. erano divisi in due sottoclassi (Apterigoti e Pterigoti) e in 30 ordini. In gran parte delle classificazioni moderne il gruppo degli Apterigoti non ha valore di sottoclasse: Collemboli, Dipluri e Proturi non sono considerati Insetti, e la loro posizione è discussa; alcuni li raggruppano in una classe separata di Esapodi, gli Entognati. La classe I. raccoglie invece 3 sottoclassi (fig. 9): due di I. primitivamente atteri, precedentemente riunite nei Tisanuri, gli Archeognati e i Dicondili (o Tisanuri); una di I. alati, gli Pterigoti, a loro volta divisi nelle infraclassi Paleotteri (ordini: Efemerotteri e Odonati) e Neotteri (25 ordini).
Gli I. e l’uomo
Considerando i rapporti con l’uomo, alcuni I. non recano né danni né vantaggi (ma, in realtà, facendo parte di biocenosi, contribuiscono al mantenimento degli equilibri biologici), altri si possono dividere in I. utili e I. dannosi.
Insetti utili. - Alcuni I. hanno utilità diretta fornendo materie pregiate: alcune cocciniglie, un tempo usate dai tintori come colorante, le api per la produzione di miele, cera, pappa reale ecc., gli I. produttori di seta (per es. il baco da seta e altri Lepidotteri Bombicidi). Inoltre molte specie di I. vengono impiegate nei paesi tropicali come cibo dalle popolazioni umane (cavallette, formiche, bruchi o crisalidi di farfalle, e perfino alcune blatte); altri I. sono stati usati o sono ancora usati in farmacopea. Gli I. hanno assunto enorme importanza come materiale biologico nella ricerca scientifica, e in particolare negli studi di genetica (i moscerini del genere Drosophila).
Numerose sono le funzioni che gli I. esercitano nelle attività umane. Alcuni rimuovono il terreno e lo concimano con i loro escrementi, altri attaccano piante spontanee parassite, velenose o infestanti, altri ancora sono pronubi, svolgono cioè l’importantissima funzione dell’impollinazione delle piante Angiosperme con la conseguente formazione dei frutti. Gli I. predatori e parassiti degli Artropodi fitofagi o comunque nocivi contribuiscono a rendere stabili gli equilibri biologici e sono talvolta utilizzati dall’uomo nella lotta biologica: Coleotteri Coccinellidi; Imenotteri Calcidoidei, Braconidi ed Evanidi; ecc.
Insetti dannosi. - Gli I. possono danneggiare le colture agricole, talvolta in modo disastroso, divorare le derrate alimentari, i manufatti, fungere da vettori di virus, batteri, funghi delle piante coltivate ecc. Infine numerosi sono gli I. parassiti dell’uomo e degli animali domestici o vettori degli agenti patogeni di gravi malattie.
Il danno provocato all’agricoltura è andato aumentando con il passare degli anni, grazie anche in parte alla pratica agricola della monocoltura e all’uso incontrollato degli insetticidi che hanno eliminato oltre ai fitofagi anche i loro predatori e parassiti, vale a dire i fattori che limitavano la crescita numerica delle popolazioni dannose, e hanno determinato l’insorgere dei fenomeni di resistenza genetica. Gli I. possono provocare danni alle piante coltivate o utili all’uomo in vari modi: divorando le foglie, i fusti, le radici, i frutti (I. fillofagi, carpofagi ecc.), scavando gallerie nel legno degli alberi d’alto fusto, oppure nelle foglie (minatori), producendo galle ecc. Il danno è particolarmente ingente quando la specie dannosa viene introdotta in un ambiente nuovo; è questo il caso della fillossera della vite, e della dorifora della patata, importate dall’America in Europa e divenute rapidamente infestanti, con conseguenze disastrose. I più noti I. nocivi all’agricoltura sono: a) fra gli Emitteri, i Psillidi (le larve di Psylla piri attaccano il pero), gli Afididi (Aphis pomi arreca danni al melo e al pero), i Fillosseridi (con la già citata fillossera), i Coccoidei (tra le cocciniglie Pseudaulacaspis pentagona attacca alberi da frutto, specie forestali, piante ornamentali e ortaggi); b) fra i Lepidotteri, i Tineidi (cui appartengono le tarme e le tignole, tra cui Sitotroga cerealella o tignola del grano, Tineola pellionella e T. bisselliela, le comuni tarme, dannose a pellicce, indumenti di lana ecc.), gli Iponomeutidi (i bruchi di Yponomeuta malinellus, o tignola del melo, si nutrono delle foglie di meli, peri e susini; Prays oleellus, tignola dell’olivo, provoca danni ingenti ad agrumeti e oliveti), i Tortricidi (le larve della tignola del pesco, Cydia molesta, attaccano i getti apicali e, in seconda generazione, i frutti del pesco, dell’albicocco, del mandorlo, del susino; Cydia, o Carpocapsa, pomonella è detta verme della mela), i Piralidi (le larve di Galleria mellonella divorano la cera degli alveari; Ephestia kühniella da larva si nutre di farine e le trasforma in ammassi di detriti; le larve di Ostrinia nubilalis, scavano gallerie nei fusti del mais, passando poi alle pannocchie, e attaccano anche canapa, fagioli, tabacco, crisantemi, dalie ecc.), i Cossidi (le larve del perdilegno rosso, Cossus cossus, e del perdilegno bianco, Zeuzera pyrina, scavano gallerie sotto la corteccia di alberi adulti di molte specie da frutto, ornamentali o forestali), i Nottuidi (le larve di Agrotis segetum si nutrono di specie selvatiche ma anche di cereali, patata, barbabietola, tabacco, vite ecc.), i Limantridi (le larve polifaghe di Lymantria dispar attaccano alberi da frutto e molte latifoglie a portamento arboreo o arbustivo e sono considerate la principale specie defoliante dei boschi di latifoglie in Europa e America Settentrionale), i Lasiocampidi (con Lasiocampa quercus o bombice della quercia), i Taumetopeidi (con la processionaria del pino, Thaumetopoea pityocampa), i Pieridi (con il genere Pieris, le cavolaie, come per es. P. brassicae, le cui larve si nutrono delle foglie di Crucifere, per es. cavoli, colza, rapa ecc.); c) fra i Ditteri, i Bibionidi (Bibio hortolanus ha larve che rodono la porzione sotterranea di piante erbacee, talora distrugge i raccolti di ortaggi), i Cecidomidi (con Mayetiola destructor, dannosa ai cereali), i Tefritidi (comprendenti specie parassite endofaghe di frutti, fusti e radici, fra cui Ceratitis capitata, la mosca mediterranea della frutta, e Bactrocera oleae, la mosca dell’olivo, un vero flagello per gli oliveti), gli Agromizidi (le larve del genere Agromyza scavano gallerie nelle foglie del carciofo); d) fra i Coleotteri, i Dermestidi (tra cui il genere Anthrenus, che arreca danni a pelli, tappeti ecc. ), gli Elateridi (Agriotes lineatus o elaterio dei cereali ha larve che si nutrono delle radichette di varie piante, come vite, tabacco, ortaggi, cereali ecc.), i Tenebrionidi (con Tenebrio molitor o larva della farina e Tribolium castaneum o tribolio, le cui larve deteriorano farine); i Cerambicidi (le larve scavano gallerie nel legno di piante sia vive sia morte, spesso marcescenti; Cerambyx cerdo attacca in genere le querce), i Crisomelidi (le larve sono fitofaghe, raramente radicivore: Lilioceris lilii, delle Gigliacee, Chrysomela populi, dei pioppi, Galerucella luteola, dell’olmo; le larve dei tonchi, genere Bruchus, si nutrono dei semi di Leguminose), i Curculionidi (con Otiorhynchus sulcatus o oziorrinco, il genere Calandra, Curculio nucum che attacca le nocciole, Anthonomus pomorum le cui larve si nutrono delle gemme fiorifere di melo e pero, impedendone la fruttificazione), gli Scolitidi (scavano gallerie, da larve e da adulti, nei tronchi degli alberi da frutto o spontanei, e possono arrecare danni ingenti); e) fra gli Imenotteri: i Tentredinidi (con specie fitofaghe ma non molto dannose: le larve del genere Hoplocampa attaccano i frutti di peri, meli, susini ecc. ), i Cinipidi (per la maggior parte cecidiogeni, cioè produttori di galle sui vegetali, ma non molto dannosi: Cynips quercus, delle querce, Diplolepis rosae, delle rose), i Formicidi, (con poche specie moderatamente dannose: Crematogaster scutellaris scava gallerie nei tronchi degli alberi, Messor barbarus asporta semi di graminacee per il nido, le formiche argentine, Linepithema humile, penetrano nelle abitazioni dove ricercano sostanze alimentari).
Gli I. dannosi all’uomo e agli animali domestici possono risultare nocivi parassitando direttamente l’uomo e gli animali domestici, ma molto maggiore è la loro importanza come vettori di agenti patogeni. Il numero dei vettori è andato aumentando per gli stessi motivi già considerati per gli I. dannosi all’agricoltura. I principali parassiti e vettori sono: a) fra i Blattoidei, i Blattidi (in Italia sono diffuse Blatta orientalis e Periplaneta americana, cosmopolite, numerose sono le specie tropicali. Invadono le abitazioni e si cibano di detriti organici. Provocano reazioni allergiche con i prodotti del loro metabolismo e sono vettori di agenti patogeni, responsabili per es. di dissenterie batteriche, salmonellosi, poliomieliti, toxoplasmosi, colera ecc., per quanto questo ruolo non sia stato ancora ben chiarito); b) fra i Mallofagi (o Ftirapteri), ectoparassiti dei peli e delle penne di Uccelli e Mammiferi, i Tricodectidi (tra cui Trichodectes canis, del cane); c) fra gli Anopluri, i pidocchi, ectoparassiti ematofagi, vettori di agenti patogeni; d) fra gli Emitteri, i Cimicidi (con Cimex lectularius, la cimice dell’uomo), i Reduvidi (il genere Triatoma e Rhodnius prolixus trasmettono il protozoo Trypanosoma cruzi, responsabile del morbo di Chagas); e) fra i Ditteri, gli Psicodidi (cui appartiene il genere Phlebotomus, i pappataci), i Culicidi (che comprendono le zanzare), i Ceratopogonidi (il genere Leptoconops attacca in massa l’uomo provocando reazioni allergiche, edemi, febbri e talora stato di shock), i Simulidi (le specie del gruppo Simulium reptans possono causare la morte dei bovini con punture velenose), i Tabanidi (Tabanus bovinus, dei bovini, ematofago, trasmette tripanosomi, il genere Haematopota attacca anche l’uomo), gli Estridi (depongono sui Ruminanti le uova o le larve, che divengono parassite sottocutanee o delle cavità dell’ospite; le larve di Dermatobia hominis, Cuterebrini, producono miasi cutanee nell’uomo e in altri mammiferi, le larve dei Gastrofilini provocano miasi gastriche su cavalli e asini), i Muscidi (Musca domestica può essere vettrice di numerose malattie batteriche e virali, Stomoxys calcitrans, pungitrice ematofaga, trasmette tripanosomi), i Glossinidi (le mosche tse-tse, gen. Glossina, trasmettono il nagana, del bestiame domestico, e la malattia del sonno, dell’uomo), gli Ippoboscidi (Hippobosca equina, dei cavalli, ematofaga, trasmette tripanosomi), i Sifonatteri (con le pulci); f) fra gli Imenotteri, i Betilidi (Scleroderma domesticum provoca con la puntura forti reazioni allergiche); g) fra i Coleotteri, gli Stafinilidi (Paederus fuscipes provoca dermatiti e lesioni oculari nelle persone che lavorano nelle risaie).
Lotta contro gli I. nocivi
I tipi di lotta contro gli I. nocivi in uso o in fase di sperimentazione sono: la lotta chimica, la lotta biologica, la lotta genetica, la lotta integrata e la lotta guidata.
Lotta chimica. - È quella più comune, che si conduce mediante l’uso di insetticidi di vario genere.
Gli insetticidi organici di sintesi (idrocarburi clorurati ecc.) sono ancora usati, ma hanno risolto solo parzialmente il problema del controllo degli I. nocivi, registrando in alcuni casi anche gravi insuccessi. Inoltre l’uso massiccio di tali sostanze ha creato una nuova serie di problemi quali lo squilibrio dei sistemi biologici, la rarefazione degli I. utili (pronubi ed entomofagi), e quindi l’abnorme moltiplicazione di quelli dannosi, l’aumento delle concentrazioni nell’ambiente di sostanze tossiche che, oltre a essere pericolose per l’uomo stesso, provocano l’avvelenamento di molte catene alimentari, e infine l’insorgere e il diffondersi della resistenza genetica negli I. che si vogliono combattere. Quest’ultimo fenomeno è dovuto, almeno per quanto riguarda alcune specie (mosche e zanzare), al fatto che il gene che conferisce la resistenza era già presente nelle popolazioni, sia pure con frequenza molto bassa, ancor prima dell’inizio dei trattamenti con insetticidi; individui portatori di geni della resistenza sono quindi sopravvissuti e hanno dato origine a popolazioni resistenti. In alcuni casi un unico gene controlla la resistenza a più categorie di fosfati organici, in altri casi geni distinti determinano la resistenza a insetticidi distinti. Per quanto riguarda il meccanismo d’azione della resistenza, è ormai noto che molte specie parassite riescono a rompere e a rendere inattive le molecole di DDT e di altri insetticidi clorurati e in alcuni casi hanno acquisito la capacità di utilizzare tali sostanze a livello metabolico. Sembra che a volte la resistenza ai fosfati organici sia il risultato di modificazioni di vari enzimi (esterasi, transferasi, ossidasi a funzioni multiple). Nell’applicazione della lotta chimica risultano quindi indispensabili, oltre a un’identificazione precisa della specie da combattere, una perfetta conoscenza del suo ciclo di sviluppo, della sua costituzione genetica e della sua ecologia, un’accurata scelta del prodotto chimico, del suo dosaggio, delle modalità di impiego, della delimitazione degli ambienti da trattare, del periodo e della frequenza dei trattamenti.
Lotta biologica. - I principali metodi di lotta biologica, che esclude l’uso di insetticidi, sono: a) la lotta biologica classica, introdotta fin dalla fine del 19° sec. e che utilizza agenti patogeni, parassiti o predatori della specie da combattere; b) la lotta mediante feromoni, in particolari attrattivi sessuali, che attirano gli I. in trappola oppure ostacolano l’incontro dei due sessi (tecnica della confusione); c) la lotta mediante ormoni (o di loro analoghi ad azione mimetica) e in particolare quelli che controllano la muta, la metamorfosi, la diapausa ecc.; d) la lotta mediante fattori anticopula, cioè sostanze che inibiscono l’oogenesi, la percezione degli attrattivi sessuali ecc., oppure fattori di monogamia.
Lotta genetica. - Combatte le specie dannose, impedendone la riproduzione o facendo in modo che la progenie della specie da combattere sia sterile o inadatta all’ambiente. Essa si attua mediante: a) il lancio di maschi sterilizzati fisicamente con raggi X o chimicamente con chemosterilizzanti; b) il lancio di individui portatori di geni letali o di aberrazioni cromosomiche (traslocazioni) che causano la produzione di gameti sterili; c) il lancio di maschi non conspecifici appartenenti a specie gemelle o di maschi della medesima specie, ma portatori di incompatibilità citoplasmatica con le popolazioni da combattere; d) il lancio di ceppi selezionati in modo da ridurre l’idoneità della specie da combattere o da modificarne il comportamento.
Lotta integrata. - Utilizza i vantaggi offerti dalla lotta biologica e dalla lotta chimica, integrandoli a vicenda. È stata finora sperimentata principalmente in agricoltura, con risultati soddisfacenti, e può essere definita come un sistema di regolazione dei fitofagi che utilizza tutte le tecniche appropriate, tenendo conto dell’ambiente in cui si opera e della dinamica di popolazioni degli I. da combattere. Il suo scopo è mantenere la densità di popolazione dei fitofagi a un livello a cui essa non causi danni economici. In primo luogo quindi viene calcolata la cosiddetta ‘soglia economica’, cioè il livello di densità relativa del fitofago al quale devono essere applicate le misure di lotta per impedire un incremento della popolazione tale da raggiungere il danno economico; è necessario, quindi, in un intervento di lotta integrata, un preliminare campionamento dell’i. dannoso per valutare la densità di popolazione. In tal modo si può pervenire a una graduale riduzione dei trattamenti con insetticidi; tale riduzione porta alla ricostruzione della cosiddetta ‘resistenza biotica’ dell’ambiente e cioè alla possibilità di azione di fattori biotici di mortalità degli I. nocivi, rappresentati in genere da altri Artropodi loro predatori. I trattamenti chimici debbono essere eseguiti con prodotti scelti tra i meno polivalenti e meno persistenti, applicati nel periodo del ciclo di sviluppo del parassita in cui esso è più suscettibile a questi interventi e devono, infine, essere sostituiti nei limiti del possibile con metodi di lotta biologica o genetica. La lotta integrata va quindi studiata e applicata caso per caso, essendo necessaria la conoscenza della biologia, del ciclo di sviluppo e della densità di popolazione del fitofago, dei suoi predatori e parassiti e lo studio dei prodotti chimici più idonei, dei periodi e delle modalità dei trattamenti.
Lotta guidata. - Consiste nell’uso deliberato dei soli mezzi chimici, che viene però calibrato in maniera tale da interferire il meno possibile con i fattori naturali di controllo delle popolazioni. Anche in questo caso vanno calcolati quindi la soglia economica, la densità di popolazione e i momenti del ciclo biologico del parassita nei quali è più opportuno effettuare il trattamento chimico.