
midollo
Anatomia e medicina
Formazione molle di vario tipo racchiusa, circoscritta o protetta dal tessuto osseo.
M. allungato
La regione di transizione fra encefalo e m. spinale.
Anatomia comparata
Deriva embriologicamente dal mielencefalo le cui pareti, nelle porzioni dorsali e ventrali ispessite, sono divaricate, specie in avanti. Ne consegue la grande espansione della volta della vescicola, la quale conserva il suo carattere non nervoso, epiteliale, e viene a costituire il tetto del IV ventricolo. Il m. allungato è sede di importanti centri che controllano attività fisiologiche indispensabili, come la regolazione della contrazione cardiaca, la respirazione, il metabolismo.
Nei Missini e Petromizontiformi la tela coroidea (➔ mielencefalo) si estende nel IV ventricolo a costituire un plesso coroideo, riccamente vascolarizzato. Simile plesso coroideo si trova nel m. allungato di tutti i Vertebrati. Nelle pareti laterali hanno le loro radici e i rispettivi centri motori i nervi cranici dal V al X paio. Sempre nei Missini e Petromizontiformi, da un certo numero di cellule gangliari giganti (cellule di Müller) si dipartono in lunghi neuriti, che connettono il m. allungato con i muscoli della coda. Un solco mediano ventrale percorre il m. allungato, in continuazione con la fessura ventrale del m. spinale. Nelle pareti laterali e ventrali decorrono tratti longitudinali di fibre ascendenti e discendenti che connettono l’encefalo al m. spinale. Due prominenze irregolari, i corpi restiformi (peduncoli cerebellari inferiori), centri di equilibrio, sporgono negli angoli laterali, anteriori, del m. allungato di Elasmobranchi, Condrostei, Olostei e Dipnoi e stabiliscono la correlazione tra i nervi sensitivi dei canali semicircolari e i muscoli del tronco.
Con lo sviluppo della linea laterale, nei Pesci, si differenziano importanti centri nervosi che, come in alcuni Teleostei, sporgono esternamente sui lati del m. allungato come due cospicui lobi del vago o internamente come lobi acustico-laterali che riducono la cavità del ventricolo. Tali lobi mancano negli Anfibi adulti e nei Rettili, nei quali non vi è linea laterale. Un paio di altri tratti di fibre, le piramidi, corrono ventralmente nel m. allungato e si continuano in avanti nel mesencefalo come peduncoli cerebrali (crura cerebri), molto evidenti nei bassi Vertebrati, meno nei Mammiferi, causa lo sviluppo del ponte di Varolio. In posizione latero-dorsale rispetto alle piramidi si trovano le olive, importanti centri di associazione, già presenti nei Pesci, particolarmente sviluppate nei Mammiferi, e i nuclei relativi che con tratti di fibre si collegano al cervelletto.
Alle 10 paia di nervi encefalici che, con le loro radici, emergono negli Anamni dal m. allungato, si aggiungono negli Amnioti l’XI paio, o spinale-accessorio, e il XII, l’ipoglosso.
Anatomia umana
Il m. allungato, o bulbo, è la porzione del tronco dell’encefalo in rapporto di continuità con il m. spinale; la porzione superiore è situata nella fossa cranica posteriore, la porzione inferiore occupa invece la porzione superiore dello speco vertebrale. Le sue caratteristiche morfologiche e strutturali rappresentano una transizione tra organi tronco-encefalici e m. spinale con il quale condivide molte formazioni.
La superficie esterna è segnata da solchi, proseguimento degli analoghi solchi del m. spinale, e da caratteristici rilievi. Procedendo dalla superficie anteriore verso la laterale, si notano: la fessura mediana anteriore, che termina in alto in una fossetta, detta foro cieco, e nella porzione inferiore è quasi cancellata dai fasci piramidali, che l’attraversano per incrociarsi con quelli del lato opposto (decussazione dei fasci piramidali); il solco laterale anteriore, dal cui fondo emergono le radicole del nervo ipoglosso; il solco dei nervi misti, che contiene le radici dei nervi vago, spinale e glossofaringeo. I rilievi sono rappresentati dalle piramidi e dalle olive.
La faccia posteriore del bulbo nella porzione inferiore ha conformazione analoga al m. spinale, con la fessura mediana posteriore e i cordoni posteriori; l’unica particolarità è rappresentata da due tubercoli grigiastri che si notano nei cordoni posteriori: la clava, che corrisponde al nucleo gracile, e il tubercolo cuneato, che corrisponde all’omonimo nucleo. Superiormente a queste formazioni i cordoni posteriori si divaricano, si portano obliquamente in alto e come peduncoli cerebellari inferiori, o corpi restiformi, raggiungono l’ilo del cervelletto. L’area triangolare delimitata dai due peduncoli è occupata da una sottile lamina, la tela coroidea del IV ventricolo, che cela la faccia ventricolare del bulbo; questa costituisce il triangolo inferiore del pavimento del IV ventricolo, o fossa romboidale.
Le formazioni grigie, che corrispondono alla sostanza grigia del m. spinale, costituiscono i nuclei della maggior parte dei nervi cranici: i nuclei dell’ipoglosso, dell’abducente, del patetico, dell’oculomotore comune; il nucleo dello spinale e quelli delle porzioni motorie del vago e del glosso faringeo che costituiscono assieme il nucleo ambiguo; il nucleo del facciale che è posto al limite con la protuberanza; i nuclei sensitivi del vago, del glosso faringeo e dell’intermediario, quelli dell’acustico (cocleare e vestibolare) e il nucleo della radice discendente del trigemino.
La sostanza bianca comprende numerosi sistemi di fibre a decorso discendente o ascendente. Delle fibre discendenti una parte si arresta nel bulbo, a livello dei nuclei motori somatici, dal III al XII paio, una parte raggiunge il m. spinale, dopo aver costituito la formazione delle piramidi. Le vie ascendenti, analogamente, non hanno comportamento uniforme: alcune attraversano il bulbo senza arrestarsi, altre al suo livello hanno una stazione sinaptica. Si limitano ad attraversarlo le fibre della via spinocerebellare e, infine, l’esiguo contingente mononeuronico di fibre spino-talamiche e spino-tettali della sensibilità protopatica.
M. spinale
La parte del sistema nervoso centrale che occupa il canale vertebrale.
Anatomia comparata
Insieme con l’encefalo, con cui costituisce il neurasse o sistema nervoso centrale dei Vertebrati, si sviluppa dal tubo neurale che, separatosi dal resto dell’ectoblasto, viene accrescendosi e differenziandosi, per moltiplicazione attiva delle cellule che ne costituiscono le pareti, assumendo la forma di un asse cilindrico schiacciato in senso dorso-ventrale. L’accrescimento in spessore del tubo è limitato alle pareti laterali, soprattutto in senso ventrale, mentre la piastra basale e la piastra tegmentale del tubo restano sottili. La prima costituisce il fondo del solco anteriore o ventrale, che corre per tutta la lunghezza del midollo spinale; la piastra tegmentale, in un primo tempo spostata dorsalmente per la crescita delle pareti del tubo e per l’aumento del diametro verticale del canale centrale (canale neurale o ependimale), viene a trovarsi in corrispondenza del setto dorsale che, nei Vertebrati più elevati, si costituisce per invasione di connettivo e di vasi nel solco dorsale, e delimita le due metà del m. spinale.
Nel tubo neurale si differenziano: le cellule dell’ependima, che conservano il loro carattere epiteliale e tappezzano il canale centrale; le cellule di sostegno della glia o neuroglia e i neuroblasti, che daranno il vero tessuto nervoso emettendo prolungamenti protoplasmatici i quali realizzano, come fibre afferenti ed efferenti, prima nude, quindi provviste di guaina, le varie connessioni a distanza. Si costituisce così una zona di rivestimento di fibre, verso la superficie esterna del m. spinale, lo strato marginale, conosciuto come sostanza bianca (fibre mieliniche o midollate), la quale circonda la parte più interna in prevalenza occupata dai corpi cellulari delle cellule nervose nonché da fibre amieliniche (neuroni motori e di associazione), la sostanza grigia. In sezione trasversa del m. spinale nell’adulto dei Vertebrati più elevati (dagli Anfibi in poi), la sostanza grigia si distribuisce in modo da assumere una forma di lettera H, nella cui barra trasversale corre, al centro, il canale ependimale. Le branche superiori dell’H corrispondono alle colonne (o corna) posteriori o dorsali della sostanza grigia, che decorrono per tutta la lunghezza del m. spinale; quelle inferiori alle colonne (o corna) anteriori o ventrali: le une e le altre sono in connessione rispettivamente con le radici dorsali e ventrali dei nervi spinali.
Si riscontrano variazioni nella distribuzione della sostanza grigia nel m. spinale delle varie classi di Vertebrati: la sostanza grigia e bianca nei Missini e Petromizontiformi non sono distintamente differenziate; nei Pesci la sostanza grigia occupa in sezione una zona a triangolo isoscele con il vertice in alto, e sono presenti solo le colonne ventrali. Nello strato marginale o periferico la sostanza bianca è distribuita in colonne longitudinali, i funicoli, composti di fascicoli o tratti di fibre (midollate) lungo le quali corrono gli impulsi, in direzione sia ascendente sia discendente, dall’encefalo al m. spinale e viceversa. Funicoli laterali si trovano sui due lati del m. spinale fra le colonne dorsali e ventrali di sostanza grigia; sia il funicolo dorsale sia quello ventrale sono divisi, rispettivamente dal setto dorsale e dalla fessura ventrale, in due metà; il funicolo dorsale comprende tratti ascendenti di fibre che convogliano impulsi prevalentemente sensori verso l’encefalo; il funicolo ventrale riunisce tratti discendenti per gli impulsi prevalentemente motori in direzione caudale o provenienti dall’encefalo. Simile organizzazione e distribuzione delle colonne e dei fascicoli caratterizzano il m. spinale delle classi più elevate di Vertebrati.
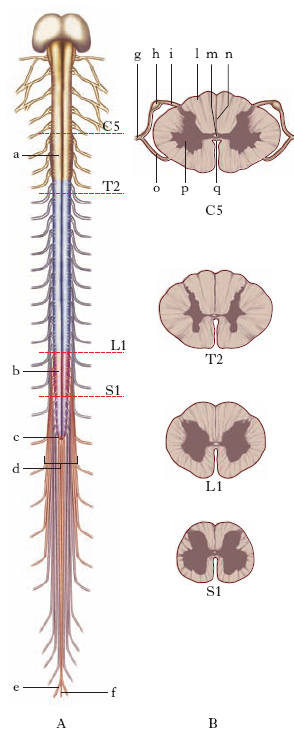
Nei Missini, Petromizontiformi e Pesci il m. spinale, più o meno tubolare o appiattito, ha un diametro quasi uniforme per tutta la sua lunghezza; nei Tetrapodi, invece, per lo sviluppo degli arti e dei nervi a questi destinati, il maggior numero di corpi cellulari determina due ingrossamenti del diametro del m. spinale a questi livelli: l’intumescenza cervicale, in relazione con gli arti anteriori, e l’intumescenza lombare, specialmente sviluppata negli Uccelli, in relazione con gli arti posteriori. Con la riduzione degli arti (in certi Sauri e Ofidi) tali intumescenze non appaiono.
In molti Vertebrati lo sviluppo e l’accrescimento in lunghezza del m. spinale e della colonna vertebrale non vanno di pari passo: può accadere che il m. spinale sia più corto della colonna vertebrale. Di conseguenza, i nervi spinali, che originariamente emergono ad angolo retto dal m. spinale, costretti a un decorso obliquo nello speco vertebrale, vengono a formare un fascio di nervi paralleli al m., la cosiddetta coda equina, con riduzione della parte terminale del m. a un esile cordone, non nervoso, il filum terminale. Nei Vertebrati diversi dall’Uomo, salvo eccezioni, si prolunga caudalmente più che nell’Uomo; sempre si manifesta una tendenza (specialmente nei Teleostei e negli Anuri) al suo accorciamento rispetto alla colonna vertebrale.
Anatomia umana
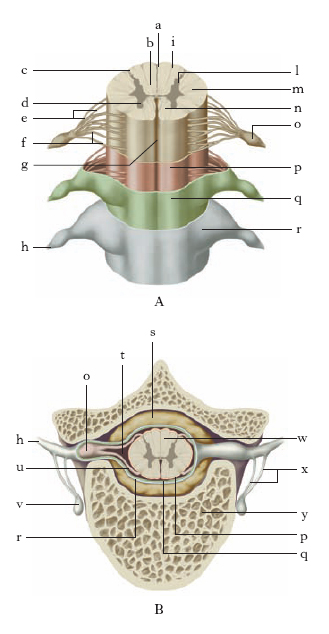
Circondato, come l’encefalo, dalle meningi, il m. spinale occupa la parte centrale del canale vertebrale (fig. 1 e 2), ha la forma di un lungo stelo grossolanamente cilindrico, della lunghezza di 41-45 cm, modicamente appiattito dall’avanti all’indietro, con due rigonfiamenti: il rigonfiamento cervicale e il rigonfiamento lombare, dove ha origine il plesso lombare. Superiormente il m. spinale si continua con il m. allungato e corrisponde al bordo superiore della prima vertebra; il suo limite inferiore, nell’adulto, corrisponde alla seconda vertebra dorsale, nel feto arriva più in basso. È però collegato al coccige dal filum terminale.
La superficie esterna del m. è divisa in due metà da due solchi: la fessura mediana anteriore e il solco mediano posteriore; sui lati esistono altri due solchi, assai meno pronunciati: il solco laterale ventrale e il solco laterale dorsale: dal primo fuoriescono le radicole delle radici anteriori, nel secondo penetrano, invece, i filamenti radicolari delle radici posteriori. Questi solchi permettono di distinguere nella superficie esterna tre cordoni: il cordone anteriore, il cordone laterale e il cordone posteriore. Dalla superficie del m., in ciascuna metà laterale e con disposizione seriata, si distaccano esili gruppi di fibre nervose (radicole) che si raccolgono in formazioni cordoniformi più voluminose, dette radici (radici spinali anteriori, o di moto, e posteriori, o di senso): dalla confluenza di ciascuna radice anteriore con la corrispondente radice posteriore prendono origine i nervi spinali e, da questi, i nervi periferici. Ciascuna coppia di radici permette di dividere il m. spinale in altrettanti segmenti, o neuromeri, diversi fra loro per grandezza e spessore.
Generalmente si considerano nel m. 4 porzioni: il m. cervicale va dal limite superiore del m. fino a buona parte del rigonfiamento cervicale e comprende i segmenti relativi alle radici degli 8 nervi cervicali; il m. toracico, o dorsale, si estende fino al rigonfiamento lombare e comprende i segmenti relativi alle radici dei 12 nervi toracici; il m. lombare corrisponde all’omonimo rigonfiamento e che è connesso con le radici dei 5 nervi lombari; il tratto terminale, di forma conoide e per questo detto cono terminale, è connesso con le radici dei 5 nervi sacrali e del nervo coccigeo.
Caratteristiche fondamentali della conformazione generale del m. spinale, quali risultano in una sezione condotta perpendicolarmente al suo maggior asse, sono la presenza in posizione centrale di un canale (canale centrale dell’ependima), residuo, in parte obliterato, della cavità primitiva, e la netta separazione delle due sostanze nervose fondamentali, con la collocazione della sostanza grigia al centro e della bianca alla periferia.
La sostanza grigia è formata in grande prevalenza da cellule nervose del tipo multipolare e da uno stroma gliovascolare. Essa, in sezione trasversale e in ciascuna metà laterale del m., presenta una forma a semiluna: sono dette corna, rispettivamente corno anteriore e corno posteriore, le porzioni estreme, massa intermedia il segmento interposto tra le due corna e commessura grigia un ponte che unisce le due semilune e fa assumere al loro insieme una forma di H. Corno laterale è denominata una piccola protuberanza che si osserva alla base del corno anteriore, nel segmento inferiore della parte cervicale e in quello superiore del tratto toracico. Va aggiunto che il corno posteriore è in rapporto con una cappa di tessuto gelatinoso, detta sostanza gelatinosa di Rolando, e per suo tramite con altre due formazioni, la zona spongiosa e la zona marginale; quest’ultima, assieme allo strato gliare sottopiale forma la zona di Lissauer.
La collezione dei neuroni è in rapporto alla funzione svolta che può essere di moto, di senso o di collegamento. Le cellule della motilità somatica, o motoneuroni, occupano il corno anteriore e, almeno nel tratto cervicale e toracico, sono ordinate in 5 colonne, una centrale, due mediali e due laterali; i loro neuriti emergono dal m., assieme a quelli degli effettori viscerali a formare le radicole, e con queste le radici spinali anteriori. I motoneuroni sono distinti in due tipi, alfa e gamma; gli alfa, che prevalgono sugli altri per numero e per dimensioni del pirenoforo e della fibra che ne prende origine, inviano il loro neurite alle placche motrici delle fibre muscolari striate, innervandone ciascuno un numero variabile, di solito un centinaio; i motoneuroni gamma provvedono invece all’innervazione delle fibre muscolari dei fusi neuromuscolari.
I neuroni sensitivi sono dislocati nel corno posteriore: la colonna più evidente è situata in prossimità della commessura grigia, prende il nome di colonna di Clarke e Stilling e corrisponde al nucleo dorsale di altre nomenclature; i suoi neuriti costituiscono il contingente più cospicuo delle fibre che conducono all’encefalo gli impulsi della sensibilità propriocettiva incosciente: in massima parte raggiungono il cervelletto come fascio spino-cerebellare. Gli altri raggruppamenti sono rappresentati dalle colonne delle cellule marginali, dalla colonna della sostanza gelatinosa e dal nucleo proprio del corno posteriore; le cellule di questi raggruppamenti danno origine alle fibre che conducono all’encefalo gli impulsi della sensibilità protopatica: la colonna delle cellule marginali e quella delle cellule gelatinose la sensibilità termica e quella dolorifica, le cellule del nucleo proprio del corno posteriore la componente tattile della sensibilità protopatica.
I neuroni di collegamento sono situati nella base del corno posteriore e nella zona confinante della commessura grigia: i raggruppamenti più noti sono rappresentati dai due nuclei commessurali, l’anteriore e il posteriore, e dal nucleo reticolare, che è situato in prossimità della sostanza bianca del cordone laterale.
Gli effettori viscerali sono raggruppati nella massa intermedia: in prossimità del cordone laterale, e limitatamente al midollo toracico e alla porzione superiore di quello lombare, è situata la colonna di Coppo e Terni, le cui cellule, tramite la catena simpatica, provvedono all’innervazione orto-simpatica dei visceri della testa, del collo e del tronco. L’innervazione parasimpatica è rappresentata da un’esile colonna, estesa a tutto il m., ad azione vasodilatatrice, denominata colonna intermedia laterale, e da due colonne (anteriore l’una, posteriore l’altra) poste in prossimità della commessura grigia ma presenti nel solo tratto lombo-sacrale, i cui neuriti provvedono alla innervazione dei visceri del piccolo bacino (intestino retto, vescica, organi genitali).
La sostanza bianca, che, come si è detto, in ciascun antimero è divisa in tre cordoni (anteriore, laterale e posteriore) dalle fessure mediane e dalle linee di impianto delle radici, comprende tre sistemi fondamentali di fibre: il sistema discendente, o di moto, il sistema ascendente, o sensitivo, e quello di collegamento tra i differenti livelli dell’asse midollare o con il tronco dell’encefalo. Comune a tutti e tre i cordoni è uno strato di fibre di collegamento intramidollare (sistema fondamentale di collegamento o circumgriseo) disposto profondamente a ridosso della sostanza grigia, che circonda in tutta la sua circonferenza, fuorché a livello delle teste del corno posteriore. Il cordone anteriore contiene in prevalenza fibre discendenti, alcune sparse (fibre tetto-, reticolo-, vestibolo-spinali), altre riunite in fasci sufficientemente individualizzati, il fascicolo longitudinale mediale e il fascio piramidale diretto: il primo è costituito di fibre vestibolari, il secondo è così denominato perché, a differenza del fascio omonimo del cordone laterale, nel m. decorre nello stesso lato di quello da cui prende origine, incrociandosi le sue fibre solo in prossimità dei vari livelli di terminazione. Le fibre ascendenti sono situate in posizione laterale e rappresentano uno sconfinamento nel cordone anteriore del lemnisco spinale del cordone laterale, di cui hanno lo stesso significato funzionale. Il cordone laterale comprende i principali sistemi di moto e di senso.
Le fibre di moto sono rappresentate principalmente dal fascio piramidale crociato, che racchiude il maggior contingente delle fibre che prendono origine dalla corteccia pre-rolandica del lobo frontale: caratteristica di questo fascio è l’incrocio compatto di tutte le sue fibre in corrispondenza della porzione inferiore del m. allungato. Le rimanenti fibre effettrici costituiscono i fasci rubro-spinale, olivo-spinale, vestibolo-spinale, reticolo-spinale e la via centrale della calotta.
Le vie sensitive costituiscono, prevalentemente, i sistemi di conduzione della sensibilità propriocettiva incosciente e della sensibilità protopatica. I primi sono rappresentati da due fasci che derivano dalla colonna di Clarke e Stilling e da altre formazioni colonnari del corno posteriore e che occupano nel cordone laterale una posizione affatto superficiale: di questi, uno è il fascio spino-cerebellare dorsale, o diretto o di Flechsig, che raggiunge il cervelletto con il corpo restiforme, l’altro è il fascio spino-cerebellare ventrale o crociato o di Gowers, che penetra nel cervelletto attraverso il peduncolo cerebellare superiore. Le vie per la sensibilità protopatica sono situate profondamente agli anzidetti fasci e solo in prossimità delle radici anteriori hanno una posizione superficiale: in massima parte derivano dal nucleo proprio del corno posteriore, dalla colonna delle cellule marginali e da quelle delle cellule gelatinose: in prossimità della loro origine incrociano la linea mediana e ascendono nella metà controlaterale del midollo in direzione dei nuclei posteriori del talamo e della lamina quadrigemina, che raggiungono in piccola parte direttamente, in una proporzione maggiore previa interruzione nei nuclei bulbari laterali o in quelli successivi del tronco dell’encefalo, di modo che le vie spino-talamica e spino-tettale sono in prevalenza polineuroniche. Altri sistemi ascendenti del cordone laterale sono rappresentati dalle vie per la sensibilità enterocettiva, che costituiscono il cosiddetto lemnisco viscerale, e da quelle per le formazioni nucleari del tronco cerebrale, come il fascio spino-olivare, così denominato per la sua terminazione nel nucleo olivare inferiore. Il cordone posteriore contiene le fibre per la sensibilità epicritica, e per origine, terminazione e significato funzionale delle sue fibre costituisce un sistema omogeneo, nonostante una solcatura longitudinale (solco intermedio) vi faccia distinguere due fascicoli: il fascicolo gracile o di Goll e il fascicolo cuneato o di Burdach. Le fibre del cordone posteriore hanno origine extramidollare, nei gangli intervertebrali e fatta eccezione per alcune, che stabiliscono un collegamento intramidollare, raggiungono il bulbo, dove si interrompono in due nuclei che hanno la stessa denominazione dei fascicoli che vi pervengono.
Patologia
Le sindromi midollari sono dovute a una lesione circoscritta e unica che interessa trasversalmente il m. spinale, in tutta la sua estensione (sindromi di sezione trasversa) o solo in parte (sindrome di emisezione) a un qualsiasi livello.
La sindrome midollare di sezione trasversa è caratterizzata da: a) paralisi di tutti e quattro gli arti (tetraplegia) nelle lesioni del m. cervicale al di sopra dei centri spinali per gli arti superiori, o solo degli arti inferiori (paraplegia) per lesione del m. dorsale; b) anestesia completa (perdita della sensibilità) nelle parti del corpo sottostanti all’interruzione; c) incontinenza urinaria e fecale, disturbi della funzione sessuale (segmenti sacrali del midollo spinale); d) abolizione della sudorazione ecc.
La sindrome midollare di emisezione trasversa, o sindrome di Brown-Séquard, è dovuta a lesioni che determinano la sezione trasversa solo di una metà del m. spinale; è caratterizzata da paralisi di uno o di entrambi gli arti d’uno stesso lato, perdita delle sensibilità spaziali (senso di posizione, sensibilità vibratoria ecc.) dallo stesso lato della lesione, con anestesia termodolorifica dal lato opposto e una zona di grave e globale ipoestesia nella superficie cutanea innervata dal segmento midollare colpito. Responsabili di sindromi midollari possono essere emorragie, processi infiammatori (➔ mielite), traumi e compressioni; queste ultime possono essere provocate da affezioni delle vertebre (tumori, fratture, morbo di Pott), delle meningi spinali (cisti, ispessimenti di natura infiammatoria) o del tessuto nervoso (tumori).
M. osseo
Tessuto molle che occupa il canale delle ossa lunghe, i canali di Havers e gli spazi intertrabecolari delle epifisi e delle ossa piatte e che costituisce il più importante organo emopoietico dell’adulto. È costituito da una sostanza amorfa, molle, trasparente, da elementi cellulari (parenchima mieloide propriamente detto) riuniti in nidi o focolai, e da uno stroma di sostegno. Quest’ultimo a sua volta è formato da esili fibre reticolari, da una rete vascolare particolarmente abbondante, da istiociti (o cellule reticolari) e da cellule adipose. Nei costituenti cellulari del parenchima mieloide sono rappresentati tutti gli elementi maturi e immaturi della serie eritrocitica, granulocitica e megacariocitica.
La diversa proporzione tra cellule dello stroma e parenchima mieloide fa assumere al m. osseo due aspetti diversi, che permettono la distinzione in m. rosso e m. giallo. Il primo, particolarmente ricco di elementi ematopoietici, è proprio del periodo fetale (m. rosso fetale); nell’adulto permane nei corpi vertebrali, nello sterno e nelle coste. Con il progredire dell’età si verifica la graduale sostituzione del tessuto emopoietico in tessuto adiposo, dando origine al m. giallo. Il prevalere della sostanza amorfa dà invece al m. un aspetto gelatinoso (m. gelatinoso, evidente nelle fasi dello sviluppo nelle ossa larghe del cranio e della faccia).
La funzione emopoietica del m. osseo si esplica attraverso tre meccanismi fondamentali: proliferazione cellulare, differenziazione cellulare e passaggio delle cellule mature dal m. al sangue. Lo studio del m. osseo per scopi clinici mira a confermare o chiarire diagnosi per i quali lo studio del sangue periferico è di scarso o di nessun ausilio. Esso si attua prelevando con appositi aghi cavi m. dallo sterno o da altre ossa che ne sono particolarmente ricche (coste, cresta iliaca, apofisi spinose vertebrali, epifisi superiori tibiali). In condizioni normali, la popolazione cellulare nucleata del m. osseo è costituita da: cellule della normopoiesi; cellule della granulocitopoiesi neutrofila, eosinofila e basofila; cellule della piastrinopoiesi; cellule indifferenziate; linfociti, plasmociti, monociti; eccezionalmente, cellule adipose, cellule dell’endotelio dei seni e, soprattutto nei bambini, osteoblasti e osteoclasti.
Botanica
Complesso di cellule parenchimatiche, spesso più o meno sclerificate, che occupa il centro del fusto e della radice nella struttura primaria; manca nell’atactostele.
Macchia midollare
Gruppo di cellule parenchimatiche, presente nel legno secondario di alcuni alberi (betulla, salice ecc.), che nella sezione trasversale appare come una macchia oblunga, bruna o rossastra, per lo più nella zona primaticcia degli anelli legnosi. Le macchie midollari sono considerate come tessuto di riserva (le loro cellule contengono molto amido), ma l’origine è dovuta alla presenza delle larve di Ditteri del genere Agromyza, le quali scavano piccole gallerie nel legno appena formatosi dal cambio. Tali gallerie vengono poi riempite da parenchima che si forma dai raggi midollari (liste di parenchima disposte in senso radiale nei fusti e nelle radici; la loro origine, costituzione e funzione sono varie (➔ radice).
Nei Licheni a tallo eteromero il m. o zona midollare è un complesso di ife lasse, che costituiscono una sorta di tessuto spugnoso, privo o quasi di gonidi, posto al disotto dello strato gonidiale.