
sesso
Complesso dei caratteri anatomici, morfologici, fisiologici (e nell’uomo anche psicologici) che determinano e distinguono, tra gli individui di una stessa specie animale o vegetale, i maschi dalle femmine. Il s. è un attributo di molti organismi viventi ed è in relazione con un sistema di riproduzione, definita sessuale o gamica in contrapposizione alla riproduzione agamica o vegetativa.
Caratteri sessuali
Il s. maschile è caratterizzato dalla produzione di cellule germinali (gameti) piccole e generalmente mobili (microgameti nelle piante; spermatozoi o spermi negli animali), quello femminile dalla produzione di gameti di dimensioni maggiori (macrogameti o ovuli nelle piante; uova negli animali), talora molto cospicue per l’accumulo di sostanze di riserva. La riproduzione sessuale consiste, tipicamente, nella unione di un gamete maschile con uno femminile (fecondazione). Negli organismi unicellulari l’unica cellula che costituisce il corpo si trasforma completamente in uno o più gameti. Negli organismi pluricellulari i gameti sono formati, attraverso la gametogenesi, all’interno di particolari organi. Questi organi sono chiamati gonadi negli animali, gametangi nei vegetali (e precisamente microgametangi o spermatangi le gonadi maschili, macrogametangi o oangi quelle femminili).
La presenza di gonadi dell’uno o dell’altro s. costituisce il carattere sessuale primario; esso può essere associato ad altri caratteri, quali la presenza di organi accessori delle gonadi, destinati a facilitarne la funzione, di organi copulatori, che provvedono all’incontro degli elementi germinali dei due s., di organi destinati alle cure prenatali e postnatali della prole, o infine di altri caratteri morfologici, fisiologici, psicologici, le cui relazioni con la funzione riproduttiva non sono sempre, a prima vista, altrettanto evidenti. A tutti questi caratteri si dà il nome di caratteri sessuali secondari. Quando i gameti maschili e femminili sono prodotti da individui diversi, si ha la condizione dei s. separati (gonocorismo o dioicismo); quando invece i gameti dei due s. sono prodotti da un solo individuo, contemporaneamente o in tempi successivi, si ha l’ermafroditismo o monoicismo. Nei vari gruppi di animali e di piante vigono condizioni diverse; in alcuni prevale l’ermafroditismo, in altri il gonocorismo; l’una e l’altra condizione possono passare all’opposta per processi evolutivi.
Sviluppo degli studi
In passato, l’identità sessuale era conosciuta soltanto in alcune forme animali, poiché non si avevano strumenti idonei (come il microscopio) per osservare i gameti. Si riteneva che gli animali inferiori e anche alcuni Vertebrati potessero nascere per generazione spontanea o per generazione senza accoppiamento e si ignorava il s. delle piante (benché Teofrasto ne avesse intuito l’esistenza). Nel 1696 R.J. Camerarius identificò il s. delle piante dioiche; gli spermatozoi furono descritti nel 1677, da A. van Leeuwenhoek, ma la loro funzione fu riconosciuta solo nel 19° secolo. Le uova di molti animali erano note ab antiquo. Nel 1672 R. de Graf scoprì il follicolo ovarico dei Mammiferi, ma la cellula-uovo di questi animali fu scoperta solo nel 1827 da K.E. von Baer. Nel 19° sec. furono studiati i particolari istologici della fecondazione: G.B. Amici (1823, 1830) osservò la formazione del budello pollinico nelle piante e la sua penetrazione nell’ovulo; N. Prungsheim descrisse, nell’alga Vaucheria, la penetrazione del gamete maschile in quello femminile (1855); nel 1875 O. Hertwig comprese, studiando l’uovo di riccio di mare, che il fenomeno essenziale della fecondazione è la fusione dei nuclei dei gameti (cariogamia). Quattro anni dopo H. Fol osservò direttamente la penetrazione dello spermatozoo nell’uovo. Soltanto quando sono stati chiariti fenomeni citologici inerenti al s. e alla fecondazione, e il s. è stato considerato alla stregua degli altri caratteri ereditari, si è potuto chiarire il suo determinismo.
Determinazione del sesso
Il s. si trasmette come un carattere ereditario. In molti organismi dioici l’eterozigosi di uno dei due s. è visibile al microscopio ottico, in quanto esiste una coppia di cromosomi (chiamati eterocromosomi o cromosomi sessuali) che è distribuita diversamente nei due s.: in sostanza, il s. è determinato da differenze cromosomiche, anche se il meccanismo di determinazione può essere sorprendentemente diverso da specie a specie. In alcune piante dioiche, e in molti Mammiferi, uomo compreso, il s. maschile è correlato alla presenza di una coppia di cromosomi XY e quello femminile a una coppia XX. Il s. maschile è definito eterogametico, perché nella gametogenesi maschile, in conseguenza della meiosi, si formano due categorie di spermatozoi, il 50% provvisti di un cromosoma X e il 50% di un cromosoma Y. Le uova invece sono tutte provviste di un cromosoma X. A seconda che l’uovo sia fecondato da uno spermatozoo dell’uno o dell’altro tipo, si forma uno zigote col corredo cromosomico maschile o femminile. In altri casi (per es., uccelli e farfalle) il s. eterozigote (ed eterogametico) è quello femminile (ZW), mentre quello maschile ha una coppia di cromosomi uguali (ZZ). Ancora, in alcuni Insetti, come gli Ortotteri, le femmine possiedono una coppia di cromosomi X, i maschi soltanto un cromosoma X (il s. maschile è il sesso eterogametico).
Il fatto che il s. sia un carattere genetico è dimostrato, oltre che dall’esistenza dei cromosomi sessuali, anche da vari altri fenomeni, che si verificano sia in organismi provvisti di cromosomi sessuali sia in quelli in cui la coppia di eterocromosomi non è riconoscibile. In alcuni animali esistono sistemi particolari di determinazione del s.; per es., nelle api, in altri Imenotteri e nei Rotiferi il s. dipende dal grado di ploidia delle uova: i maschi nascono (per partenogenesi) da uova non fecondate, aploidi, le femmine da uova fecondate, diploidi.
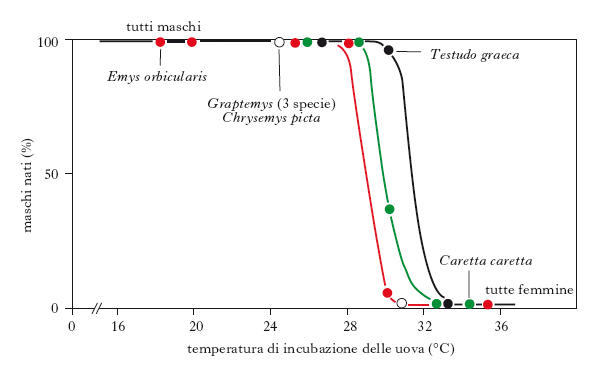
La determinazione del s. è un processo complesso, dovuto sia a cause genetiche sia ambientali, come è anche dimostrato dai fenomeni di intersessualità (➔). Variabili ambientali come temperatura, fotoperiodo, pH, disponibilità trofica e competizione locale per l’accoppiamento, ma anche batteri o fattori citoplasmatici, possono influenzare la determinazione del s. in molti Vertebrati e taluni invertebrati. La determinazione ambientale del s. (ESD, environmental sex determination) è quindi il meccanismo con cui particolari valori e/o caratteristiche di un fattore ambientale, indipendentemente dal genotipo dell’individuo, sono in grado di influenzare irreversibilmente la determinazione primaria del s. durante lo sviluppo embrionale o postembrionale. La temperatura, per es., è uno dei fattori che determinano il s. nei Rettili (lucertole, coccodrilli, tartarughe; fig. 1) e in alcuni Teleostei. Nei pesci concorrono il fotoperiodo e il pH del mezzo ambientale, mentre in alcuni Mammiferi è determinante il nutrimento. Il fotoperiodo sembra tra i fattori maggiormente coinvolti anche nella ESD dei Crostacei, anche se in alcuni di questi organismi è stata dimostrata la presenza di fattori epigenetici che possono alterare la determinazione genetica del s.: negli oociti dell’isopode Armadillidium vulgare, per es., è stato identificato un batterio femminilizzante termosensibile a volte presente come fattore citoplasmatico, cioè come DNA batterico integrato non stabilmente nel genoma dell’ospite. Negli oociti dell’anfipode Gammarus duebeni, invece, il fotoperiodo, insieme alla presenza di microsporidi nel citoplasma, induce la determinazione del s. femminile. La ESD è favorita dalla selezione naturale laddove la fitness dell’individuo, in particolari condizioni ambientali, dipende dal sesso. Inoltre, poiché la ESD produce un’alterazione del rapporto numerico tra i s., il principale vantaggio delle specie a ESD è la stretta relazione causale tra i valori dei parametri ambientali e il rapporto numerico tra i s. della popolazione.
Il rischio per le specie a ESD è che modificazioni estreme delle variabili ambientali interessate, sia naturali sia causate dall’uomo, possano indurre uno scostamento eccessivo del rapporto numerico tra i s. da quello di 1:1.
Determinazione del s. nell’Uomo
Affinché un individuo della specie umana si sviluppi in un maschio o in una femmina fenotipicamente normale, durante l’ontogenesi deve verificarsi una serie di complessi eventi molecolari e morfologici, nei tempi appropriati e nella sequenza corretta. Il s. genetico dell’embrione è comunque determinato dal suo assetto cromosomico, in primo luogo dalla presenza/assenza del cromosoma Y. È stato osservato che la maggior parte dei soggetti affetti da sindrome di Turner (fenotipo femminile, con ovari ridotti, amenorrea, caratteri sessuali secondari poco sviluppati) hanno un solo cromosoma X (e non due, come nel corredo femminile normale); invece gli individui affetti da sindrome di Klinefelter (fenotipo maschile, alta statura, ipoplasia testicolare, azoospermia) hanno un cromosoma Y e due cromosomi X, invece di uno come nel corredo maschile normale. Il fatto che individui che possiedono un cromosoma Y, indipendentemente dal numero di cromosomi X presenti, abbiano comunque uno sviluppo in senso maschile, dimostra che nell’uomo (così come nel topo, e probabilmente in tutti i Mammiferi) il cromosoma Y porta i geni per la mascolinità. Poiché nei Mammiferi il differenziamento sessuale coincide con il differenziamento delle gonadi in testicoli od ovaie, si deduce che uno o più geni del cromosoma Y siano necessari per determinare lo sviluppo della gonade embrionale indifferenziata in testicolo. La gonade maschile a sua volta produce il testosterone e il fattore antimulleriano, ormoni responsabili della regolazione del differenziamento dei genitali interni ed esterni, determinando così il s. fenotipico. L’assenza del cromosoma Y permette invece il differenziamento della gonade embrionale in ovaio e quindi lo sviluppo dell’individuo in senso femminile.
Un gene candidato a codificare il fattore responsabile del differenziamento testicolare è il gene SRY (sex region of Y), identificato da P. Goodfellow (1990) sul braccio corto del cromosoma Y umano. Una sequenza nucleotidica omologa a SRY è stata riscontrata sul cromosoma Y di tutti i Mammiferi finora analizzati. SRY codifica una proteina nucleare capace di riconoscere specifiche sequenze di DNA e di legarvisi, regolando così l’attività di altri geni. Questo gene da solo non è in grado di assicurare un corretto differenziamento testicolare, ma funziona come un ‘interruttore’ genetico che inizia la serie di eventi molecolari (attivazione o repressione di un certo numero di geni) che porterà alla formazione della gonade maschile. Questa idea è confermata dal fatto che nell’Uomo sono noti casi di anomalo differenziamento sessuale di individui con cariotipo 46, XY (disgenesia gonadica XY), che non sono correlati a mutazioni del gene SRY, e dal fatto che esistono patologie ereditarie associate a inversione del s. causate da mutazioni autosomiche e non dei cromosomi sessuali: per es., la sindrome di Denys-Drash (delezione parziale del cromosoma 11); la sindrome di Smith-Leimli-Opitz (delezione parziale del braccio lungo del cromosoma 7); il nanismo campomelico (riarrangiamenti cromosomici che coinvolgono una regione del braccio lungo del cromosoma 17).
Organi della riproduzione sessuale
Gli organi primari della riproduzione sessuale sono le ghiandole sessuali, cioè gli ovari, che producono le cellule-uovo, e i testicoli, produttori delle cellule spermatiche o spermatozoi, gli uni e gli altri insieme denominati gonadi o ghiandole germinali. Nei Vertebrati le ghiandole sessuali, oltre che produrre i gameti, funzionano da ghiandole a secrezione interna secernendo gli ormoni sessuali; sempre nei Vertebrati, le ghiandole sessuali, di origine mesodermica, sono in genere organi pari, ma in alcuni gruppi, come nei Ciclostomi, in certi Pesci, nella femmina di molte specie di Uccelli, sono impari e tali risultano o per fusione dei loro abbozzi pari, o per rudimentazione della gonade di un lato. Si formano molto precocemente nell’embrione, da due ispessimenti appaiati dell’epitelio celomatico, ai lati del mesentere dorsale, detti creste genitali, che successivamente si evolvono in ovari o in testicoli.
Oltre che dalle ghiandole sessuali, gli organi della riproduzione sono costituiti dai dotti per l’emissione dei prodotti sessuali (gonodotti), gli organi sessuali accessori (come le ghiandole destinate a secernere gli involucri dell’uovo, l’albume, il guscio ecc.) che favoriscono e assicurano l’incontro delle cellule riproduttive o gameti; in complesso tutte le strutture facenti parte dell’apparato genitale. I gonodotti della gonade femminile si dicono ovidutti, quelli della gonade maschile vasi deferenti o semplicemente deferenti, e in ambedue i casi possono funzionare come tali i nefridi (per es., negli Anellidi) o nefridi modificati in celomodotti. Nei Vertebrati, eccettuati i Teleostei e alcuni altri Pesci, funzionano da ovidutti, nella femmina, i dotti di Müller modificati; nel maschio invece funzionano da deferenti, negli Anamni i dotti del pronefro, negli Amnioti i dotti di Wolff, che si originano in rapporto al pronefro e al mesonefro. In certe specie di Pesci e Anfibi, i duttuli efferenti del testicolo rappresentano tubuli renali modificati per convogliare gli spermatozoi nel dotto del pronefro che funziona da deferente; negli Amnioti, con la scomparsa del mesonefro, il dotto di Wolff persiste come deferente e come dotto dell’epididimo che si connette ai duttuli efferenti del testicolo, cioè tubuli del mesonefro modificati e persistenti come tali. Fra gli Invertebrati, per es. gli Artropodi, le gonadi, tipicamente due, di origine celoteliale, sono provviste di gonodotti e relativi annessi che, nel maschio, sono: una o due vescicole seminali, in cui gli spermatozoi si concentrano prima della eiaculazione, da uno a tre paia di ghiandole annesse, il canale eiaculatore, l’organo copulatore (pene o cirro); nella femmina: i ricettacoli seminali, in cui si raccolgono i gameti maschili, le spermateche, in cui si conservano gli spermi destinati alla fecondazione, la vagina, atta ad accogliere l’organo copulatore, e gli ovidepositori per la deposizione delle uova. La presenza di organi copulatori caratterizza gli animali che hanno fecondazione interna, cosa che particolarmente si verifica negli Artropodi e nei Vertebrati; nelle femmine degli organismi vivipari è spesso presente l’utero, rappresentato da una parte dilatata dei gonodotti, destinata a ospitare l’embrione o gli embrioni durante lo sviluppo e la gestazione; ma anche negli animali ovipari, per es. nei Platelminti, si trovano uteri in cui si raccolgono le uova dopo la fecondazione. Nella condizione sessuale ermafrodita di questi Invertebrati, nell’apparato genitale femminile, oltre l’ovario o germigeno (germario), si ha un vitellario o vitellogeno che provvede a fornire le uova di cellule vitelline; e in quello maschile, oltre le vescicole seminali, si hanno speciali ghiandole prostatiche (per gli organi sessuali dell’Uomo (➔ genitale, apparato).
Gli ormoni sessuali sono rappresentati dal testosterone e dagli altri androgeni, dall’estradiolo e dagli altri estrogeni, dal progesterone e suoi derivati: pregnenolone, pregnandiolo ecc. Gli androgeni promuovono lo sviluppo dei caratteri sessuali secondari maschili; gli estrogeni e il progesterone, oltre a controllare la comparsa dei caratteri sessuali secondari femminili, inducono le periodiche modificazioni dell’endometrio che caratterizzano i cicli sessuali. Gli androgeni sono prodotti, oltre che dal testicolo, anche dal surrene e forse, in scarsa misura, dall’ovario. Gli estrogeni sono elaborati dall’ovario, dalla placenta e anche dal testicolo (specialmente nello stallone). Sostanze ad azione estrogena (fitoestrogeni, xenoestrogeni) sono isolabili anche da alcune piante (finocchio, anice, mandragora ecc.) e da alcuni materiali (bitume, nafta, fango, acqua del Mar Morto). La produzione del progesterone e derivati avviene da parte del corpo luteo, dei surreni e anche della placenta. Nella struttura chimica gli ormoni sessuali presentano stretta parentela, perché si possono considerare tutti come derivati di un medesimo idrocarburo tetraciclico saturo, il ciclopentanoperidrofrenantene (che costituisce, tra l’altro, anche il nucleo degli steroli e degli acidi biliari). Ciò spiega anche una certa ambivalenza dei loro effetti biologici (singoli ormoni posseggono alcune proprietà in comune con quelle del gruppo confinante) e tende ad avvalorare l’ipotesi che i due tipici ormoni sessuali, testosterone ed estradiolo, siano elaborati a partire da una medesima sostanza.