
cromosoma
Nome dato da W. Waldeyer nel 1888 ai piccoli corpi intensamente colorabili, in genere di forma bastoncellare, visibili nel nucleo della cellula durante la mitosi. Secondo la teoria cromosomica, dimostrata da T.H. Morgan con ricerche su Drosophila melanogaster, i c. sono le strutture essenziali dell’eredità.
Caratteri generali
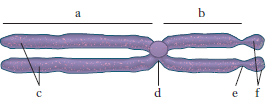
La struttura base è costituita da un filamento a doppia elica di DNA avvolto intorno agli istoni (proteine basiche). Nella sequenza di basi del DNA è scritta tutta l’informazione genetica di una cellula per la formazione di proteine; una sequenza di basi, che codifica una proteina o meglio una catena polipetidica, prende il nome di gene. Quando il DNA si duplica, durante l’interfase del ciclo cellulare, l’informazione genetica viene riprodotta fedelmente ed è trasmessa alle cellule figlie. Al momento della mitosi il DNA è quindi già duplicato e il c. appare diviso in due elementi, i cromatidi, uniti a livello del centromero (fig. 1). Quando si completa la divisione cellulare ognuna delle cellule figlie riceve uno dei due cromatidi. I cromatidi sono costituiti da un’unica molecola di DNA a doppia elica molto spiralizzata. La spiralizzazione di questa molecola si realizza mediante stretti avvolgimenti intorno alle proteine istoniche in strutture fondamentali chiamate nucleosomi.
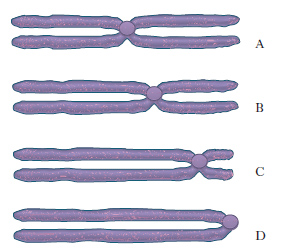
Durante la metafase mitotica, i c. appaiono al microscopio ottico ben divisi nei due cromatidi ed è possibile studiare e individuare ogni singolo c. in base ad alcuni parametri: a) lunghezza relativa dei c.; b) presenza di una costrizione, situata sempre nella stessa posizione (costrizione primaria o centromero); a seconda della posizione del centromero, che è considerato il centro cinetico del c. e corrisponde al punto in cui il c. si attacca alle fibre del fuso durante l’anafase, si distinguono: c. metacentrici o submetacentrici, con centromero in posizione mediana e submediana; c. acrocentrici, con centromero subterminale; c. telocentrici, in cui il centromero è terminale (fig. 2). Il rapporto fra le lunghezze dei due bracci di un c., misurate dal centromero, è detto indice centromerico; nel metacentrico l’indice centromerico è pari a 1; c) presenza di costrizioni secondarie (studi citologici e biochimici hanno rivelato che la costrizione secondaria dei c. è il sito dove sono localizzati i geni che codificano per l’RNA ribosomale, cioè l’RNA che costituisce i ribosomi; questo RNA, appena prodotto, si addensa a formare, insieme a una componente proteica, i nucleoli che rimangono vicini alla costrizione secondaria e che per tale ragione viene chiamata regione dell’organizzatore nucleolare); d) presenza di satelliti, formazioni sferiche terminali unite al c. con un sottile filamento. Ulteriori dati sull’individuazione dei c. si ottengono con particolari tecniche di colorazione (➔ bandeggio).
Il numero e la forma dei c. sono costanti per ogni specie di animali e di piante; negli organismi diploidi il corredo cromosomico completo diploide (indicato con il simbolo 2 n) è rappresentato da due serie di c. identici tra loro due a due, detti omologhi; per es. l’uomo ha 2 n = 46 c. (23 coppie); il moscerino Drosophila melanogaster 2 n = 8 c. (4 coppie); il granturco Zea mais 2 n = 20 c. (10 coppie); il topo 2 n = 40 c. (20 coppie) e così via (per le eccezioni a questa regola ➔ poliploidia). L’informazione genetica dei c. omologhi deriva per metà dal gamete maschile e per metà dal gamete femminile. Durante il processo di formazione dei gameti (negli organismi a riproduzione sessuale) il numero diploide 2 n dei c. viene, attraverso il processo della meiosi (o gametogenesi), dimezzato a n cromosomi (numero aploide). All’atto della fecondazione tra i due gameti si ricostituisce nello zigote il numero diploide.
Tipi di cromosomi
C. sessuali In molte specie di animali e in alcuni vegetali a sessi separati si osserva una coppia di c. che differiscono nel maschio e nella femmina, e che sono appunto chiamati c. sessuali (o eterocromosomi), mentre gli altri c. del corredo prendono il nome di autosomi. Si possono avere diversi casi: la femmina ha due cromosomi uguali (XX) e il maschio due diversi (XY) oppure uno solo (XO); in questo caso il sesso femminile è omozigote o omogametico, il maschile eterozigote o digametico. Alcuni geni sono localizzati sui c. sessuali: nell’uomo appartengono a questa categoria, detta dei caratteri ‘legati al sesso’, il daltonismo, l’emofilia e altri geni. C. giganti Detti anche salivari, sono presenti nel nucleo delle grosse cellule delle ghiandole salivari delle larve di alcuni Ditteri (in particolare Chironomus e Drosophila), si caratterizzano per: a) l’appaiamento dei c. omologhi (sinapsi somatica), per cui il numero di c. risulta aploide; b) la notevole lunghezza, dovuta al fatto che essi si trovano in uno stato di forte despiralizzazione; c) l’ammasso eterocromatico ( cromocentro) nel quale sono riuniti tutti i centromeri e buona parte del c. Y; d) la presenza di un’alternanza di bande chiare e scure che sono state interpretate come formate dalla giustapposizione dei cromomeri allineati sui numerosi cromatidi appaiati (e per questa ragione i c. salivari vengono detti anche politenici); si ritiene infatti che i c. salivari originino da un processo di endomitosi, cioè da una serie di duplicazioni successive dei c. non seguiti da divisioni cellulari. In particolari stadi della vita larvale di Drosophila è possibile individuare lungo i bracci di questi c. dei rigonfiamenti detti anelli di Balbiani (o puffs) al cui interno si verifica una sintesi molto attiva di RNA. C. a spazzola Nome dato da J. Rückert a particolari c., detti anche piumosi, presenti negli oociti di alcuni vertebrati (specialmente anfibi), durante lo stadio meiotico del diplotene; vanno perciò considerati bivalenti. Lungo i cromatidi, molto despiralizzati, si evidenziano i cromomeri, in corrispondenza dei quali si formano due anse; poiché le anse sono molto numerose, i c. assumono un aspetto piumoso. È stato dimostrato che lo stadio piumoso corrisponde a un periodo di attiva sintesi di RNA. C. soprannumerari In numerose specie animali e vegetali alcuni individui presentano un numero di c. diverso da quello normale e costante della specie a causa dell’esistenza dei c. soprannumerari eterocromatici (detti anche c. accessori o B-cromosomi), i quali non sono omologhi a nessun membro del corredo cromosomico della specie; tale fenomeno pertanto non va confuso con i casi di ploidia (➔ poliploidia).
C. dei Procarioti
Nei virus il c. è rappresentato da una singola molecola di DNA o di RNA strettamente arrotolata. Il DNA può essere a singolo filamento o a doppio filamento, può avere forma ad anello o essere lineare; in alcuni batteriofagi (fago lambda) il c. ha doppia elica, forma bastoncellare e le estremità sono ‘adesive’, per cui dopo che il virus è penetrato nella cellula ospite, il c. prende forma di anello. Nei batteri la regione che contiene il DNA viene detta nucleoide, per analogia con il nucleo degli Eucarioti. Nel nucleoide il DNA è strettamente attorcigliato, tanto da risultare escluso dalle particelle citoplasmatiche per quanto non sia circondato da una membrana come nel nucleo degli Eucarioti. Il c. batterico è stato studiato in Escherichia coli, che ha un unico c. circolare a doppio filamento di DNA.
Malattie cromosomiche
Sindromi associate ad alterazioni della struttura o del numero dei cromosomi. Le più note nell’uomo sono: la trisomia 13; la trisomia 18; la trisomia 21 (➔ Down, John Langdon Haydon).