
filamento
Astronomia
Struttura oscura, dall’aspetto appunto filamentoso, che si osserva nelle regioni attive del Sole. I f. sono dovuti a protuberanze della superficie solare.
Biologia
In genetica, ciascuna delle due catene di una molecola di DNA, normalmente costituita da una coppia di f. fra loro complementari. Nel processo di sintesi del DNA si chiama f. guida (leading strand) quello che è sintetizzato in maniera continua, in direzione 5′3′, e f. tardivo (lagging strand) quello sintetizzato in direzione opposta 3′5′, in maniera discontinua sotto forma di brevi frammenti successivamente legati da legami covalenti.
In biologia cellulare, f. intermedio, una delle componenti del citoscheletro, di 8-12 nm di diametro, che aiutano a mantenere la forma e la mobilità cellulare e che costituiscono punti di ancoraggio per altre strutture cellulari. Le proteine dei f. intermedi sono le più abbondanti fra le proteine specifiche di vari tipi di cellule differenziate; si ritiene che siano i principali elementi nella determinazione della struttura di molte cellule e tessuti dei Metazoi e che svolgano un ruolo più specifico e complesso nella regolazione dei processi cellulari rispetto alle altre due componenti del citoscheletro, i microtubuli e i microfilamenti, costituiti dalle proteine ubiquitarie, actina e tubulina. Durante la lisi della cellula, queste vengono solubilizzate, mentre la maggior parte dei f. intermedi è insolubile. Le proteine dei f. intermedi sono state purificate e identificate mediante la lisi delle cellule, sfruttando la loro insolubilità e la capacità delle loro subunità proteiche di riformare f. dopo la denaturazione.
Gli studi condotti mediante l’impiego di anticorpi specifici, e soprattutto mediante il sequenziamento degli mRNA, hanno permesso di identificare nei Vertebrati almeno 5 tipi di proteine costituenti i f. intermedi, espressi in cellule differenti: la vimentina è espressa principalmente in cellule di origine mesenchimale, quali i fibroblasti, e nelle cellule endoteliali dei vasi sanguigni; i f. di desmina si trovano prevalentemente nelle cellule muscolari di qualsiasi tipo e in particolare nel muscolo striato; i neurofilamenti, costituiti da 3 tipi diversi di polipeptidi, sono presenti negli assoni dei neuroni sia centrali sia periferici; la proteina acida fibrillare della glia forma i f. intermedi delle cellule gliali circondanti i neuroni e viene espressa esclusivamente in alcuni tipi di cellule gliali; le citocheratine costituiscono i f. intermedi delle cellule epiteliali.
La filamina è una proteina dimerica (peso molecolare 240.000-280.000), diffusa in numerosi tipi cellulari, che lega tra loro i microfilamenti di actina mediante legami trasversali. La filamina appartiene alla famiglia delle cross-linking proteins, proteine che legano altre proteine. Per es., piccolissime quantità di filamina possono trasformare i f. di actina in soluzione in un gel solido. La sua funzione è quindi quella di stabilizzare tra loro i vari componenti del citoscheletro (➔ citoplasma), dato che la filamina, oltre che all’actina, si può associare anche alle altre proteine che lo costituiscono.
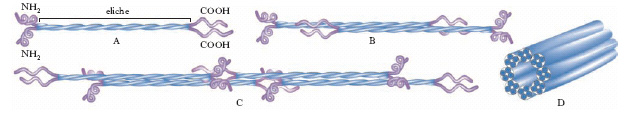
Tutte le subunità proteiche dei f. intermedi, anche se formate da proteine diverse, hanno una struttura comune (v. fig.): presentano un dominio centrale a bastoncino lungo circa 40 nm, costituito da due eliche superavvolte (dimeri); all’estremità carbossilica e amminica del bastoncino sono presenti tratti, non elicoidali, di lunghezza e sequenza variabili. Il dominio centrale contiene inoltre 3 elementi spaziatori a struttura non elicoidale. Le subunità dimeriche polimerizzano spontaneamente formando i protofilamenti. Otto protofilamenti formano poi il f. cilindrico, di circa 10 nm, che costituisce il f. intermedio. Molte prove sperimentali suggeriscono che i geni che codificano le proteine dei f. intermedi appartengano alla stessa famiglia, cioè si siano evoluti da un unico gene primordiale. Poiché le lamine nucleari si trovano nelle cellule di tutti i Metazoi e hanno una struttura molto simile a quella dei f. intermedi, è possibile che le proteine dei f. intermedi si siano evolute da un precursore simile alle lamine.
I f. intermedi sono presenti anche nelle cellule tumorali; poiché queste ultime mantengono molte delle proprietà differenziate delle cellule dalle quali derivano, il tessuto di provenienza può essere identificato mediante anticorpi fluorescenti diretti contro i f. e questa tipizzazione è spesso importante per la terapia. Per es., i più comuni carcinomi mammari e del tratto gastrointestinale contengono citocheratine e sono privi di vimentina; è quindi evidente che derivano dalle cellule epiteliali e non dalle sottostanti cellule dello stroma di tipo mesenchimale, che tipicamente contengono invece f. di vimentina.
Botanica
Parte sterile, di norma assottigliata, dello stame, che sorregge la parte fertile o antera, ove si sviluppano i granuli pollinici. Varia molto per lunghezza (a volte è così breve che l’antera appare sessile); può essere filiforme o dilatato alla base o appiattito; talora è ramificato e può anche avere appendici stipoliformi, o essere rivestito di peli staminali (Commelinacee). Il f. s’inserisce in un punto del connettivo dell’antera. Ha struttura anatomica molto semplice: l’epidermide, con pochi stomi, ricopre un parenchima normale; al centro si ha un esile fascio vascolare.
Tecnica
In elettrotecnica, nelle lampade elettriche a incandescenza, conduttore filiforme racchiuso all’interno del bulbo e reso luminoso, per incandescenza, dal passaggio della corrente elettrica.