
fotosintesi clorofilliana
Il più importante processo di riduzione biologica che si compie sulla superficie terrestre, grazie al quale le piante fotosintetiche assorbono energia luminosa, che convertono in energia chimica potenziale e, partendo da composti inorganici semplici (anidride carbonica e acqua), formano sostanze organiche (glucidi ecc.) le quali costituiscono i composti della materia vivente, sia vegetale sia animale.
Reazione fotosintetica
Lo svolgimento della f. si suole rappresentare con la reazione globale
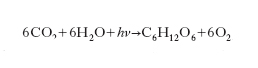
dove C6H12O6 rappresenta uno zucchero e hν un fotone. Tale reazione è fortemente endotermica.
L’energia luminosa utilizzata allo scopo dalle piante fotosintetiche (per semplicità, ‘piante verdi’) costituisce la fonte esterna di energia che, al livello del cloroplasto, entra nel mondo biologico e gli assicura il rifornimento energetico necessario a mantenere lo stato costante e il funzionamento degli organismi. La reazione comporta diminuzione di entropia e aumento di energia libera. La variazione di energia libera è ΔF=+2870 kJ/mol; la variazione di entropia è ΔS=−182 J/mol•K (cioè, −55 kJ/mol a 25°C circa). L’ossigeno che si sviluppa nella f. proviene dalla scissione fotolitica di molecole di acqua e non, come si supponeva un tempo, dalla CO2, la quale viene incorporata come tale in un composto fosforilato a 5 atomi di carbonio, ribulosio-1,5-difosfato (Ru-d-P), che è presente nel cloroplasto ed è continuamente rigenerato.
Cloroplasti e fotosistemi
Sede della f. sono le strutture laminari dei cloroplasti (alcuni organismi primitivi, come le Cianofite e i Batteri fotosintetici, sono privi di plastidi e contengono lamelle isolate). Nelle piante superiori i cloroplasti sono numerosi, in media da 30 a 40 per cellula nel parenchima clorofilliano (tessuto a palizzata) situato tipicamente sotto l’epidermide superiore della foglia, l’organo fotosintetico per eccellenza ancorché la f. si svolga in tutte le parti verdi della pianta. Nel cloroplasto delle alghe verdi (Clorofite) e delle piante superiori, l’interno del plastidio, delimitato da una doppia membrana propria, è caratterizzato dalla presenza di due tipi di lamelle: le lamelle dello stroma, che decorrono secondo l’asse maggiore del plastidio, e le lamelle dei grani, che si presentano al microscopio elettronico come pile di dischi, a doppia membrana su entrambe le facce, a guisa di monete una sull’altra. I pigmenti sono associati alle lamelle, e mancano nello stroma del cloroplasto. Oltre alla clorofilla a, le piante verdi contengono anche clorofilla b e pigmenti carotenoidi. Le piante con cloroplasti granulari e provviste di clorofilla b producono amido di assimilazione.
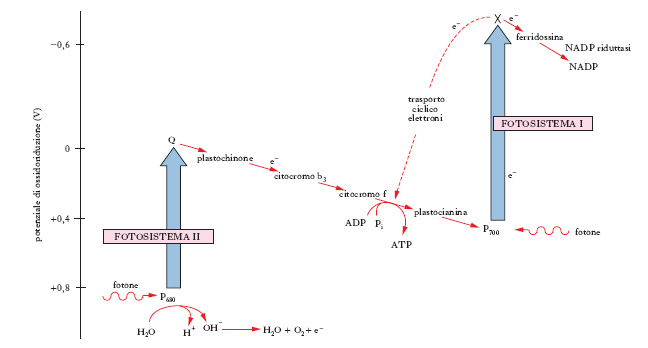
Nel cloroplasto si trovano sistemi multienzimatici, che presiedono ai seguenti processi: assorbimento di energia luminosa e sua conversione in energia chimica con variazione del potenziale di ossidoriduzione; fosforilazione fotosintetica; riduzione della CO2 e sintesi di glucidi. I primi due processi caratterizzano la fase luminosa della f., il terzo la fase oscura o enzimatica. Nella fase luminosa si ha una serie di reazioni che si svolgono, all’interno dei grani, in sistemi di pigmenti detti fotosistemi (I e II), due speciali siti attivi delle unità fotosintetiche. Queste sono complessi di 250-400 molecole di clorofilla, che fungono da antenne, assorbendo fotoni e convogliandoli a due centri di reazione (fig. 1), denominati P700 (fotosistema I) e P680 (fotosistema II) perché presentano un massimo di assorbimento rispettivamente a 700 nm e 680 nm. I due centri di reazione sono direttamente collegati a una serie di accettori di elettroni. I fotosistemi I e II contengono sia clorofilla a che b, ma in proporzioni diverse. Il centro di reazione P700 assorbe energia. La molecola P700, attivata, cede elettroni all’accettore X, che può cederli alla ferridossina. Essi sono impiegati, in definitiva, per ridurre NADP a NADPH2; ovvero sono trasportati da una catena di trasportatori e impiegati per sintesi di ATP, prima di ritornare, dopo aver ceduto l’energia di eccitazione, a P700 e riportarlo allo stato fondamentale. Il deficit di elettroni trasferiti al NADP viene colmato mediante trasferimento di altri elettroni dal centro di reazione P680 attraverso coenzima Q→plastochinone→citocromi b3 e f→plastocianina, al fotosistema I, in P700. In corrispondenza di P680 avviene la reazione di fotolisi di molecole di acqua. Mediante questa reazione sono prodotti elettroni, che riportano allo stato neutro P680, nonché ioni H+, che sono impiegati per la riduzione di NADP (attivato a NADP–) a NADPH2 e radicale OH•, dai quali si formano acqua e l’ossigeno che viene svolto nella fotosintesi.

In fig. 1 è riassunto il percorso del trasporto di elettroni nella fotosintesi. L’ATP e il NADPH2 prodotti nella fase luminosa sono impiegati per l’organicazione dell’anidride carbonica nella fase oscura. Il primo prodotto stabile che si forma nella reazione oscura è l’acido 3-fosfoglicerico, a tre atomi di carbonio; l’accettore della CO2 è il ribulosio-1,5-difosfato, zucchero a 5 atomi di carbonio. Si forma un β-chetoacido, molto labile, che si scinde dando due molecole di acido fosfoglicerico; questo, con l’intervento di ATP e NADPH2 originati nella fase luminosa, è ridotto ad aldeide fosfoglicerica, che è capace d’isomerizzarsi dando fosfodiossiacetone. L’aldeide fosfoglicerica e il fosfodiossiacetone in parte si condensano dando fruttosio-1,6-difosfato, che è il punto di partenza per la formazione del saccarosio e dell’amido; parte delle molecole di fruttosio-1,6-difosfato e di fosfodiossiacetone rigenerano l’accettore iniziale di anidride carbonica, cioè il ribulosio-1,5-difosfato. Questo ciclo (ciclo di Calvin) è schematizzato in fig. 2.
Fotofosforilazione
La sintesi di ATP, oltre che per liberazione dell’energia di ossidazione dei substrati respiratori (fosforilazione ossidativa), può avvenire anche utilizzando energia luminosa (fosforilazione fotosintetica o fotofosforilazione). A partire da ADP si forma ATP enzimaticamente in accoppiamento al trasporto di elettroni dalla clorofilla eccitata dalla luce, e così si conserva, come fosfato ad alta energia, una parte dell’energia luminosa assorbita. La fotofosforilazione può avvenire con un trasporto non ciclico di elettroni dal fotosistema II al I; oppure, con un trasporto ciclico, e in questo caso con produzione di ATP ma non di NADPH. La fotofosforilazione ciclica ha lo scopo di regolare il rapporto tra le due sostanze, per es., se nella luce manca la lunghezza d’onda di 680 nm, propria del fotosistema II, o se vi è eccesso di NADPH.
Fotorespirazione
Nelle cellule clorofilliane delle piante C3 ben illuminate viene consumato ossigeno e liberata CO2 di recente organicazione a causa del processo di fotorespirazione, che è diverso dalla respirazione mitocondriale e ha luogo specialmente nei cloroplasti e nei perossisomi. Il processo è dovuto al fatto che la ribulosio-difosfato-carbossilasi funziona anche come una ossigenasi. In questo caso, in presenza di luce e O2, il ribulosio-1,5-difosfato viene scisso in 3-fosfoglicerato e 2-fosfoglicolato; quest’ultimo può essere metabolizzato a: glicolato-gliossilato-glicina-serina-idrossipiruvato-glicerato-3-fosfoglicerato (ciclo del glicolato). La conversione glicolato-gliossilato avviene nei perossisomi. Nel passaggio glicina-serina si libera CO2 senza produzione di ATP o di NADPH, e senza altro vantaggio. Le normali piante C3, nelle quali vige il ciclo di Calvin, perdono a causa della fotorespirazione dal 25 al 50% del carbonio organicato.
Fotosintesi delle piante C4 e CAM
Molte piante originarie delle regioni arido-calde e a elevata luminosità, come il mais, la canna da zucchero, il sorgo ecc., hanno sviluppato una via fotosintetica collaterale. Esse hanno cellule verdi oltre che nel mesofillo anche a ridosso dei fasci vascolari, intorno ai quali formano quasi una guaina. Le cellule verdi del mesofillo fissano la CO2 sul fosfoenolpiruvato (PEP) formando acido ossalacetico (OAA) e fosfato secondo la reazione, catalizzata dalla PEP-carbossilasi: PEP+CO2→ OAA+fosfato. Successivamente interviene la malico-deidrogenasi che permette la riduzione dell’OAA ad acido malico con l’intervento di NADPH fornito dall’apparato fotosintetico dei cloroplasti: OAA+NADPH→acido malico+NADP. Poiché con la fissazione della CO2 si formano acidi a 4 atomi di carbonio (OAA, malato) si è parlato di f. C4 o di piante C4. L’acido malico viene poi traslocato alle cellule verdi della guaina vascolare, e qui viene decarbossilato, con l’intervento dell’enzima malico: acido malico+NADP→ CO2+NADPH+acido piruvico. La CO2 liberata è utilizzata per il ciclo di Calvin, mentre il piruvato viene traslocato al mesofillo e qui riconvertito in PEP. La collocazione fra cellule del mesofillo e cellule della guaina vascolare si basa sul fatto che le piante C4 hanno un’elevata efficienza fotosintetica e sono in grado di mantenere la concentrazione di CO2 nell’atmosfera a valori pressoché nulli, purché l’intensità luminosa sia elevata. Poiché le piante C4 utilizzano la Ru-d-P-carbossilasi solo nelle cellule della guaina vascolare, dove la concentrazione della CO2 è sempre notevole, esse non presentano il fenomeno della fotorespirazione. Ciò compensa il dispendio energetico che la via C4 comporta, perché quando essa opera contemporaneamente al ciclo di Calvin sono necessari 2 ATP in più per ogni mole di CO2 organicata.
Un comportamento simile a quello delle piante C4 si riscontra nelle Crassulacee e in parecchie piante succulente, dette piante CAM dall’inglese crassulacean acid metabolism. Queste piante fissano la CO2 nelle ore notturne quando hanno gli stomi aperti e la incorporano nel PEP, formando acido malico e altri acidi organici intermedi del ciclo di Krebs. Durante il giorno, a stomi chiusi, il malato viene decarbossilato e la CO2 liberata viene fissata secondo il ciclo di Calvin. La chiusura degli stomi alla luce, durante il giorno, si interpreta come un adattamento a economizzare l’acqua.