
Evoluzione. Plasticita del fenotipo
Evoluzione. Plasticità del fenotipo
Lo studio della proprietà di un genotipo di produrre differenti fenotipi quando esso viene esposto ad ambienti diversi (plasticità fenotipica) rappresenta un tema centrale della moderna ecologia evolutiva, o ecologia darwiniana. Tale disciplina indaga i meccanismi con cui l'ambiente fisico e quello biologico determinano l'evoluzione di caratteri semplici o complessi che influiscono sulla fitness. Gli aspetti evolutivi del rapporto organismo/ambiente possono oggi essere indagati in profondità solo dopo che modelli teorici e studi sperimentali, oltre a confermare l'idea (risalente a Richard Woltereck e Ivan Schmal'gauzen) che la funzione di mappa genotipo-fenotipo non è lineare, hanno anche sottolineato l'importanza del fenomeno della plasticità fenotipica nell'evoluzione. La plasticità può manifestarsi o come 'polifenismo' (in tal caso, come nel polimorfismo classico, i fenotipi sono discontinui) o come 'norma di reazione' (e allora si ottiene una gamma continua di fenotipi espressi dal medesimo genotipo). Qui tratteremo la plasticità a carico di caratteri con variazione continua.
Se la norma di reazione può essere vista come quella funzione ‒ definita nello spazio fenotipo/ambiente ‒ che connette l'input ambientale all'output fenotipico, la plasticità fenotipica rappresenta allora una misura della norma di reazione di un organismo. La plasticità determina il possibile insieme di habitat idonei per un genotipo. Stante la loro immobilità, le piante sono in media molto più capaci di plasticità degli animali; tra questi sembrano più dotate di plasticità le specie sessili come le spugne e i coralli, rispetto a quelle tipicamente mobili. La plasticità ha una base genetica e dunque può evolvere come qualunque altro carattere; inoltre è stato dimostrato che lo stesso genotipo è più o meno plastico a seconda dell'ambiente in cui si trova e dei caratteri (biochimici, fisiologici, anatomici, morfologici) considerati. Studi risalenti agli anni Settanta del Novecento hanno chiarito il rapporto tra analisi statistica della varianza e norma di reazione, suggerendo che il grado di ereditarietà di un carattere può cambiare con l'ambiente. Negli ultimi decenni sono stati compiuti notevoli progressi nello studio della plasticità fenotipica.
Mentre in passato la plasticità era vista come un elemento di disturbo nello studio dei fenomeni evolutivi, oggi essa rappresenta il principale interesse di ricerche che si servono di una varietà di metodiche, includenti sia la genetica quantitativa e quella molecolare sia i numerosi approcci metodologici mirati alla costruzione di modelli evolutivi delle risposte plastiche. Qui saranno considerati solo alcuni tra gli aspetti più rilevanti della ricerca sulla plasticità fenotipica e verranno discusse le aree di ricerca in cui sono stati ottenuti progressi sostanziali e quelle ancora da approfondire. Alcune questioni, come lo studio della genetica quantitativa su cui si basa la plasticità fenotipica, possono dirsi risolte, almeno a grandi linee, oppure sono superate da domande e da approcci nuovi. Altre questioni, come i costi biologici della plasticità, rappresentano le frontiere della ricerca in questo campo e probabilmente diventeranno le aree di maggiore sviluppo futuro.
Da problema a paradigma
Il concetto di plasticità del fenotipo (la capacità di un genotipo di produrre distinti fenotipi quando è esposto a differenti ambienti durante la sua crescita) è al centro della vecchia questione riguardante l'opposizione nature/nurture (ove nurture va qui intesa come somma delle influenze che modificano l'espressione delle potenzialità genetiche di un organismo), che ha interessato filosofi da Platone a Locke. Nella letteratura biologica moderna, la plasticità del fenotipo è stata considerata un problema dai tempi della riscoperta delle leggi di Mendel fino ai primi anni Ottanta del Novecento, quando divenne parte integrante della comprensione di come gli organismi si sviluppano e interagiscono con il loro ambiente. Molti biologi avevano ancora una visione erronea circa la natura della plasticità, specialmente in rapporto alla genetica degli organismi. Nel 1952, infatti, Douglas S. Falconer intitolò appropriatamente un suo lavoro, oramai classico, The problem of environment and selection; in esso si sosteneva che gli effetti ambientali rappresentavano un problema poiché interferivano con la selezione naturale e artificiale dei caratteri. Ancora oggi la contrapposizione fra genetica e plasticità ha i suoi sostenitori, come se la plasticità non fosse di fatto una proprietà esclusiva del genotipo.
Nonostante questa confusione, la plasticità fenotipica è ora riconosciuta come un importante concetto nel pensiero evolutivo moderno, particolarmente dopo la pubblicazione di diverse importanti rassegne da parte di Amy D. Bradshaw, Carl D. Schlichting, Sonia E. Sultan, Mary Jane West-Eberhard e Samuel M. Scheiner. In questa sede saranno discussi quelli che sembrano essere i temi principali che orientano gli studi sulla plasticità. Non considereremo, invece, il campo della biologia molecolare delle risposte plastiche, un settore in rapida espansione ma concettualmente distinto dal tema qui affrontato e che richiederebbe un'approfondita, autonoma trattazione.
Variazione genetica ed ereditabilitàdella plasticità
Anche soltanto uno sguardo superficiale alla letteratura settoriale rivelerebbe centinaia di studi sulle interazioni gene-ambiente (G×E, cioè il prodotto della variazione genetica per plasticità), in molti organismi diversi (v. Tav. I). In effetti alla domanda sull'eventuale esistenza di variabilità genetica della risposta plastica si deve rispondere affermativamente: in natura esiste una variazione genetica per le risposte plastiche. Si tratta ora di documentare specifici casi oggetto studio, quando questa documentazione diventa necessaria per poter passare alla formulazione di domande più complesse.
Come per l'ereditabilità della plasticità, Samuel M. Scheiner e Richard F. Lyman nel 1989 hanno introdotto una procedura formale per calcolarla, ma per ragioni logistiche e concettuali tale misura è stata raramente usata in letteratura. Da un punto di vista logistico è oneroso eseguire una serie di prove sperimentali con un numero sufficientemente alto di famiglie geneticamente collegate (cresciute in un cospicuo numero di ambienti diversi) per ottenere stime di ereditabilità; inoltre, l'intervallo di confidenza di tali stime è talmente ampio da rendere non particolarmente ricco di informazioni lo sforzo sperimentale. Questo è un problema ben noto nella stima quantitativa dell'ereditabilità in generale, che però si amplifica quando bisogna moltiplicare il numero degli esperimenti per il numero delle diverse condizioni ambientali in cui sono eseguiti.
Concettualmente il problema è ancora più pesante a causa delle numerose critiche che fin dagli anni Settanta del XIX sec. ha ricevuto il concetto stesso di ereditabilità, a partire dal lavoro di Richard C. Lewontin del 1974. È lo studio della plasticità in sé, infatti, che ci ha permesso di comprendere i limiti intrinseci alla misurazione dell'ereditabilità: le stime di ereditabilità di un qualsiasi carattere possono variare fortemente in rapporto all'ambiente nel quale essa è stata misurata, e non esiste una regola generale che permetta di estrapolare i risultati da alcune condizioni ambientali ad altre, né dalle condizioni di laboratorio a quelle naturali. Di conseguenza, stimare le varie ereditabilità della variabilità fenotipica di un carattere in singoli ambienti rischia di essere una perdita di tempo e di risorse.
Selezione della plasticità fenotipica
L'osservazione comune dell'esistenza di variazione genetica intrapopolazionale per la plasticità implica che la plasticità possa evolvere in risposta alla selezione naturale; la qual cosa, a sua volta, fa pensare che le popolazioni naturali possiedano una plasticità fenotipica di natura adattativa. Sorprendentemente questa area di ricerca ha ricevuto poca attenzione. Ancora una volta ci sono valide ragioni di ordine logistico per evitare di studiare gli effetti della selezione sulla plasticità, date le grandi dimensioni di tali esperimenti e anche la difficoltà di effettuare repliche in condizioni ambientali che siano ecologicamente plausibili e tenute sotto controllo. Ancora, domande correlate a come (e quanto frequentemente) la selezione naturale agisca sulla plasticità sono concettualmente cruciali non solo per la nostra comprensione della relazione G×E ma anche per capire, più in generale, l'evoluzione fenotipica.
La letteratura recente sul problema della selezione e della plasticità raccoglie articoli che esemplificano differenti approcci al problema. Heidrun Huber e collaboratori, per esempio, hanno eseguito nel 2004 una classica stima dei coefficienti di selezione, focalizzandosi sul carattere evitamento dell'ombra nella pianta Impatiens capensis. Gli autori hanno roscontrato un alto grado di variazione microambientale nei coefficienti di selezione, con il fenotipo 'evitamento dell'ombra' che si presentava come vantaggioso solo in alcuni micrositi. Benché questa distribuzione potrebbe portare al mantenimento della plasticità fenotipica in risposta a cambiamenti nella qualità della luce (il segnale che scatena l'evitamento dell'ombra), gli autori hanno anche osservato che la densità di piantine era un cattivo predittore della variazione microambientale nella selezione direzionale. Invece, la disponibilità d'acqua appariva avere un ruolo insospettato, probabilmente poiché il fenotipo evitamento dell'ombra è costoso nei micrositi secchi. Questo è il tipo di studio che dovrebbe essere più frequente in letteratura, dato il grado di approfondimento che esso può fornire nello studio della genetica ecologica delle risposte plastiche. Esso evidenzia, peraltro, come il mondo reale sia molto più complesso quando viene confrontato con le nostre semplici aspettative basate su esperimenti eseguiti in condizioni controllate.
Un secondo approccio allo studio della selezione e della plasticità è stato intrapreso da Markus Fischer e coautori nel 2004 attraverso la selezione artificiale della plasticità delle forme di crescita di Ranunculus reptans. Questo ranuncolo adotta due forme di crescita: le piante del tipo guerrilla si diffondono velocemente mandando propaggini in tutte le direzioni, dopodiché aumenta la crescita di quelle propaggini che si vengono a trovare in microambienti favorevoli; gli individui di tipo phalanx crescono più lentamente manifestando una distribuzione più regolare delle propaggini (in realtà esiste un continuum di forme tra queste due strategie). Fischer e i suoi collaboratori selezionavano le piante in base a una diminuita o aumentata guerrillaness e per la maggiore o minore plasticità del carattere. Nonostante abbiano ottenuto risposte significative alla selezione sulla guerrillaness, non hanno riscontrato alcun cambiamento della loro plasticità, concludendo che l'evoluzione delle strategie di crescita in questa specie è possibile, alterando l'altezza piuttosto che la forma della norma di reazione.
Un altro approccio sfrutta la manipolazione sperimentale (in quanto distinta dalla selezione naturale o artificiale). Anne Lyytinen e collaboratori nel 2004 hanno studiato la farfalla Bicyclus anynana, una specie che presenta due forme stagionali: una forma della stagione umida con macchie ocellari sulle ali, e una forma della stagione secca senza macchie. Utilizzando Uccelli insettivori, gli sperimentatori hanno esposto le farfalle ai predatori disponendole su uno sfondo di foglie marroni oppure verdi, colori che rappresentano rispettivamente i substrati tipici delle stagioni secca e umida. I risultati di questi esperimenti corroborano l'idea che esiste una controselezione delle macchie ocellari nella stagione secca (l'assenza di macchie ocellari favorisce di molto la mimetizzazione criptica), alternata con una selezione a favore delle macchie ocellari durante la stagione umida (deviando gli attacchi verso parti meno vitali del corpo, le macchie ocellari possono infatti conferire una buona protezione contro gli uccelli predatori). La combinazione dei due regimi selettivi potrebbe essere sufficiente a mantenere la plasticità fenotipica adattativa nella formazione delle macchie ocellari.
Queste ricerche evidenziano la capacità risolutiva delle strategie sperimentali multiple, non necessariamente tutte impiegate nello stesso laboratorio, che combinano sia gli studi sul campo e quelli in condizioni controllate, sia le manipolazioni sperimentali e la raccolta accurata dei dati osservativi. Un peculiare problema di questo tipo di ricerche può essere rappresentato dalla circostanza che spesso esse si basano su studi a bassa tecnologia, che presentano un'esecuzione tediosa e che richiedono alti costi di personale e tempi lunghi; si tratta di una combinazione di fattori che, se confrontati con gli standard operativi della ricerca nelle scienze ad alta tecnologia, diventano piuttosto difficili da giustificare presso i finanziatori della ricerca.
Costi e limiti alla plasticità fenotipica
Benché in natura esista una variazione genetica per la plasticità e sia dimostrato che la selezione naturale favorisce alcuni tipi di risposte plastiche, possono esservi anche limiti all'evoluzione della plasticità adattativa. Oltre all'esistenza di possibili vincoli genetici e/o dello sviluppo, Thomas J. DeWitt e collaboratori nel 1998 hanno avanzato l'ipotesi che vi possano essere diversi tipi di costi misurabili necessari al mantenimento della plasticità, o limiti alla stessa capacità di un organismo di essere adattivamente plastico. Benché importante, la differenza concettuale tra costi e limiti di un adattamento è spesso poco considerata: la conseguenza dei costi è la diminuzione della fitness persino quando viene espresso un fenotipo ottimale, mentre un primo limite consiste nell'incapacità di esprimere il fenotipo ottimale. Inizialmente, è stato difficile rilevare i costi della plasticità, benché lavori recenti siano poi riusciti a determinarli in molti sistemi diversi. In alcuni casi, i costi sono stati rilevati nella stessa specie solo in alcuni distretti dell'area di distribuzione geografica, forse in relazione all'esistenza di pressioni selettive circoscritte, locali; e forse potrebbe essere più o meno difficile dimostrare l'esistenza di costi di tipo diverso a seconda della specifica ecologia evolutiva della specie in esame.
Secondo la modellistica applicabile a popolazioni strutturate spazialmente, la selezione naturale favorisce le norme di reazione che bilanciano l'evitamento del costo con l'acquisizione di risorse. Ci si può aspettare, per esempio, che il costo di mantenimento di una risposta plastica possa provocare l'evoluzione di norme di reazione che aumentano l'adattamento agli ambienti che l'organismo incontra più spesso. I costi generati dalla produzione di una risposta plastica sono pagati soltanto quando uno specifico fenotipo è generato in un dato ambiente, il che rende questi costi proporzionali alla frequenza con cui si presenta l'ambiente (o gli ambienti) ove il fenotipo considerato è favorito. La ricerca sui costi della plasticità è ancora agli esordi, ma si tratta di un campo importante da un punto di vista teorico, anche se impegnativo dal punto di vista empirico; in futuro potrebbe diventare un'area di grande interesse.
Correlazioni genetiche come limitazioni?
L'argomento delle correlazioni genetiche come limitazioni spesso viene avvicinato alla plasticità fenotipica, benché in due contesti separati. Vogliamo ricordare che la correlazione genetica è una correlazione tra due caratteri fenotipici, calcolata utilizzando la frazione di varianza fenotipica statisticamente associata alle differenze genetiche tra gli individui.
Il primo tipo di contesto nel quale le correlazioni genetiche e la plasticità vengono discusse è stato preso in esame da Sara Via nel 1987 e riguarda il modo alternativo di visualizzare le norme di reazione attraverso descrizioni grafiche delle medie genetiche dell'espressione di un carattere in un primo ambiente contro l'espressione dello stesso carattere in un secondo ambiente, generando così una correlazione genetica per quel carattere attraverso gli ambienti. Benché questo sia un buon modo per visualizzare la plasticità in termini di genetica quantitativa, esso presenta alcuni limiti dovuti al fatto che si possono visualizzare solo due ambienti per volta.
Il secondo tipo di contesto è stato analizzato da Stephen Stearns e collaboratori nel 1991 e si riferisce al fatto che G×E può alterare la grandezza, e alcune volte il segno, della correlazione genetica tra i due caratteri misurata in un certo numero di ambienti. Le correlazioni genetiche sono misurate in un dato contesto ambientale e se il contesto ambientale viene cambiato esse stesse possono essere plastiche. Ci sono due motivi per considerare importante tutto questo: il primo è che lo studio delle modificazioni indotte dall'ambiente sulle correlazioni genetiche tra caratteri è un buon modo per avvicinare il problema più generale della dipendenza dei vincoli e del trade-off tra caratteri dal contesto. Il secondo motivo è che, poiché spesso si assume che sui tempi evolutivi le correlazioni genetiche si mantengono costanti, la dimostrazione che esse possono essere modificate in una sola generazione da cambiamenti ambientali ha ovvie conseguenze di portata teorica generale per la genetica evolutiva dei caratteri quantitativi.
C'è anche un motivo teorico sempre più stringente per dubitare che le correlazioni genetiche sui vincoli contengano informazioni significative, e quindi utili al di là della pura descrizione statistica della variazione. Gli studi compiuti da David Houle nel 1991 e da Mark H. Gromko nel 1995 hanno dimostrato che non si possono ricavare informazioni sulla sottostante architettura genetica di un carattere a partire da osservazioni sulla correlazione genetica, poiché cause diverse possono generare lo stesso modello di correlazione; sicché, si possono utilizzare le ipotesi sulle cause per giungere a predizioni sulle correlazioni osservate, ma non si possono individuare le cause a partire dalla conoscenza delle correlazioni. Questo non significa che lo studio delle correlazioni genetiche sia inutile: le distribuzioni osservate possono suggerire ipotesi circa le cause, le quali possono essere convalidate ricorrendo al controllo sperimentale. Comunque, il dibattito in generale sul migliore uso possibile delle correlazioni genetiche, e in particolare sul loro impiego nelle ricerche sulla plasticità, è destinato a continuare ancora per qualche tempo.
Basi genetiche della plasticità fenotipica
I differenti modelli proposti per spiegare le basi genetiche della risposta plastica sono un argomento importante nella letteratura sull'evoluzione della plasticità. Nella rassegna del 1993 Samuel M. Scheiner riassume i tre principali modelli: sovradominanza, pleiotropia, epistasi. Il modello della sovradominanza stabilisce che la plasticità è una funzione inversa dell'eterozigosi: più un genotipo è eterozigotico, meno sarà plastico, poiché l'eterozigosi aiuta a tamponare le influenze ambientali. Il modello della pleiotropia si fonda sul fatto che la plasticità origina da alcuni geni con effetti pleiotropici su un determinato carattere espresso in differenti ambienti. Il modello epistatico, infine, prevede l'esistenza di due distinti insiemi di geni; uno determina l'altezza della norma di reazione (cioè il valore della media attraverso gli ambienti), l'altro determina la sua forma (ovvero la sua pendenza, nel caso di una norma di reazione di tipo lineare), e questi due insiemi interagiscono tra loro epistaticamente.
Tali questioni sono state affrontate dalla ricerca empirica dell'ultimo decennio. Da complessi studi nel campo della biologia molecolare della plasticità, come quelli sulla risposta di evitamento dell'ombra nelle piante, o quelli sulla risposta allo shock termico, sappiamo che l'eterozigosi (nel modello della sovradominanza) ha poco a che fare con le basi genetiche della plasticità, e che sia l'effetto pleiotropico sia quello epistatico sono caratteristici di tutti i casi di risposta plastica fino a oggi indagati. Si potrebbe concludere che il problema delle basi genetiche della plasticità, che sono sempre state concettualmente difficili da definire, si è dissolto nei molti problemi relativi alle specifiche interazioni molecolari sottostanti ai particolari tipi di plasticità.
È importante allora riflettere su una domanda di base: perché consideriamo la plasticità come un qualcosa di unitario, che possiede una specifica base genetica? Sarebbe come domandarsi se il carattere X (per es., la taglia) dipenda dalla pleiotropia, dall'epistasi, o se sia influenzato dal grado di eterozigosi. La risposta potrebbe essere che tale carattere dipende da tutti e tre i fattori; nessuno dei tre 'meccanismi' è da solo particolarmente informativo, poiché è verosimile che essi concorrano tutti a costituire il fondamento dei caratteri fenotipici più complessi.
Modelli matematici dell'evoluzione della plasticità
Tra gli addetti ai lavori, l'espressione 'modellare la plasticità' si riferisce anche alla costruzione di modelli teorici relativi alle modalità di evoluzione della plasticità fenotipica. Benché una parte di questa modellistica si fondi sull'esame di modelli della genetica della risposta plastica discussi in precedenza, i due significati di modellamento della plasticità sono di fatto abbastanza indipendenti uno dall'altro e possono essere discussi separatamente. Sempre Scheiner, nel 1993, propone un utile schema che riassume i tipi di modelli usati per studiare l'evoluzione della plasticità. Le categorie di modelli sono essenzialmente tre: di ottimalità, di genetica quantitativa, e gametici. I modelli di ottimalità sono indipendenti da qualsiasi considerazione di natura genetica, poiché essi si interrogano (date certe limitazioni e in presenza della necessaria variazione genetica) su quale tipo di strategia ottimale possa evolvere in determinate condizioni. La mancanza di genetica potrebbe essere vista come una limitazione del modello di ottimalità, ma potrebbe anche essere interpretata come un vantaggio, dato che lo scopo del modello è quello di esplorare una serie di ipotetici scenari selettivi del tipo 'se-allora', piuttosto che quello di fare predizioni quantitative su traiettorie evolutive effettive.
I modelli di genetica quantitativa, nonostante il loro nome, sono anche largamente indipendenti dalla genetica vera e propria. Essi considerano la genetica come una scatola nera, da cui emerge la necessaria varianza genetica 'additiva' (cioè la variazione genetica che permette a un carattere di rispondere alla selezione). Questi sono modelli statistici e incorrono nelle stesse limitazioni delle correlazioni genetiche. I modelli gametici sono gli unici che incorporano la genetica reale, in quanto descrivono ciò che accade a caratteri che non solo sono sotto l'influenza di processi evolutivi come la selezione e la migrazione, ma che sono anche influenzati da un certo numero di loci, interagenti tra loro secondo specifiche modalità. Mentre il pregio dei modelli gametici risiede nella loro capacità di esprimere direttamente i fenomeni genetici, come la pleiotropia e l'epistasi, il loro problema è che sono limitati a scenari semplici (per necessità di trattabilità matematica), e diventano imprevedibili quando si cerca di renderli più realistici. Eventuali modelli di simulazione riescono solo parzialmente a ovviare a questo problema, poiché sono computazionalmente ingestibili oppure a causa del fatto che non sono in grado di garantire soluzioni generali.
Un'analisi della letteratura recente sulla produzione di modelli dell'evoluzione delle norme di reazione mostra che la maggioranza dei ricercatori è interessata ai modelli di ottimalità, spesso legati a particolari situazioni ecologico-evolutive. Per esempio, Torbjørn Ergon e collaboratori nel 2004 hanno impiegato un approccio di ottimalità per studiare le relazioni tra taglia del corpo e consumo di energia durante l'inverno nell'arvicola Microtus agrestis. Il modello implicava un compromesso tra i benefici per la sopravvivenza dell'essere grande e il costo che questo comporta in termini di approvvigionamento del cibo. Le previsioni del modello sono che le arvicole dovrebbero essere più piccole in ambienti che sono più impegnativi, con una risultante correlazione negativa tra consumo energetico giornaliero e massa corporea. Tuttavia, se nel modulare il fabbisogno energetico come funzione dei costi di approvvigionamento gli animali mostrano una plasticità fenotipica, allora il modello predice l'esistenza di correlazione positiva tra massa corporea e consumo energetico. Gli autori hanno sottoposto il loro modello a convalida sperimentale, trovando che il consumo energetico era maggiore nei siti dove le arvicole erano più piccole, nonostante l'esistenza di correlazione positiva tra le due misure all'interno dei siti. La loro conclusione è che la variazione di taglia è attribuibile a eterogeneità nella qualità del cibo o nella sua disponibilità, piuttosto che nella plasticità adattativa nell'attività di approvvigionamento.
Una combinazione tra modelli di ottimalità e lavoro sperimentale nella convalida di previsioni teoriche è stata utilizzata per studiare, per esempio, la distribuzione delle risorse basata su caratteri della storia vitale nell'ostrica del Pacifico (Crassostrea gigas), l'architettura radicale e l'acquisizione delle risorse nel fagiolo (Phaseolus vulgaris), e il controllo materno del sesso nella prole in Agaonidi del genere Otitesella in risposta alla densità locale di popolazione. Questo particolare aspetto della modellistica della plasticità fenotipica è maturo per una rassegna generale che potrebbe essere d'aiuto sia ai biologi teorici sia a quelli sperimentali.
Macroevoluzione e plasticità fenotipica
La plasticità è stata anche proposta come un meccanismo potenzialmente importante per facilitare la macroevoluzione. Ciò può avvenire attraverso almeno due modalità: da una parte la plasticità può portare all'assimilazione genetica di un carattere quando una popolazione occupa un nuovo ambiente. In sostanza, una variazione già esistente per la plasticità consentirebbe a una popolazione di permanere in nuove condizioni ambientali, anche se in condizioni adattative molto lontane da quelle ottimali. Tale persistenza darebbe il tempo necessario alla comparsa di variazione genetica nuova (per mutazione e/o ricombinazione), potendo quindi permettere alla selezione naturale di aumentare l'adattamento alle nuove condizioni. Se queste condizioni ambientali persistono, la selezione favorirebbe una diminuzione della plasticità attraverso l'assimilazione genetica del carattere (o dei caratteri). Un meccanismo di questo tipo potrebbe essere all'opera, per esempio, durante la ben nota fase di latenza che accompagna la colonizzazione di molte specie invasive, prima che esse si diffondano nel nuovo ambiente.
Dall'altra parte, i cambiamenti fenotipici indotti geneticamente sono armonizzati dalla plasticità naturale del sistema di sviluppo ('aggiustamento fenotipico'). Un esempio ci viene dall'abilità di alcuni quadrupedi di sviluppare posture quasi bipedi attraverso un complesso cambiamento del loro sistema muscolo-scheletrico in risposta a una mutazione che rende non funzionali le zampe anteriori. La selezione naturale può quindi assimilare il nuovo fenotipo prendendo l'aspetto di un'evoluzione a mosaico.
La ricerca futura sulle conseguenze macroevolutive della plasticità dovrà documentare l'esistenza sia di casi di assimilazione genetica che di aggiustamento fenotipico, posizionarli nel loro contesto filogenetico, e sviluppare approcci empirici per poterli studiare. Si tratta di un compito oneroso, sia concettualmente sia operativamente, ma che potrebbe anche rappresentare una delle prossime rivoluzioni nel nostro modo di pensare l'origine delle novità evolutive.
Lo studio della plasticità fenotipica: il futuro
È sempre rischioso fare previsioni sulla direzione della scienza, persino sul breve termine; nondimeno, le speculazioni sulle prospettive della ricerca nello studio della plasticità fenotipica potrebbero rappresentare una spinta intellettuale per i ricercatori interessati a questo settore della ricerca biologica. Vorrei sottolineare come alcune domande scientifiche siano state mal poste o non siano più scientificamente rilevanti. Tra queste è forse paradigmatico il tema delle basi genetiche delle risposte plastiche. Mai come in questo caso, una domanda concettualmente corretta formulata all'inizio di una ricerca è stata completamente superata dagli studi sulle basi molecolari della plasticità di specifici caratteri. Anche le domande sulle correlazioni genetiche e la plasticità sono ormai diventate del tutto desuete. Benché abbiano generato ricadute benefiche per gli studi di genetica quantitativa e per la caratterizzazione delle matrici di varianza e co-varianza genetica, dobbiamo però accettare il fatto che si tratti niente altro che studi di correlazioni. Un'altra area di studio della plasticità che non sembra in grado di schiudere nuovi orizzonti è la quantificazione dei modelli di variazione genetica quantitativa e l'ereditabilità della plasticità. Questo non per la presenza di un problema concettuale intrinseco, ma perché siamo effettivamente riusciti a dare risposte a domande di ampio respiro: adesso sappiamo che c'è sufficiente variazione genetica a disposizione delle risposte plastiche.
Per quanto riguarda la quantificazione e la comprensione dei modelli di selezione per le risposte plastiche resta ancora molto lavoro da fare. Siamo a conoscenza che ci può essere selezione a carico della plasticità, e, d'altro canto, che la plasticità può rispondere alla selezione. Comunque sappiamo ancora poco delle condizioni ecologiche che favoriscono la selezione stabilizzante e quella direzionale sulle norme di reazione, e poco conosciamo dei tipi di life-history, dei sistemi di accoppiamento e persino della storia filogenetica che sono più o meno riconducibili alla variazione genetica che può rispondere a tali pressioni selettive. Le ricerche di questo tipo, benché vitali per la nostra comprensione dell'ecologia evolutiva della plasticità, sono operativamente pesanti e noiose. Nonostante ciò, tali ricerche dovrebbero essere condotte nella maniera più rigorosa possibile.
Il giudizio circa i modelli teorici sulla plasticità è di tipo misto. Benché la genetica quantitativa e i modelli gametici abbiano prodotto conoscenze approfondite e interessanti sull'evoluzione delle norme di reazione, sembra che l'accoppiamento dei modelli di ottimizzazione con i dati empirici costituisca l'approccio scientifico più fruttuoso. Questa proposta metodologica, comunque, si basa sulla valutazione del compromesso tra realismo, precisione e generalità dei modelli e la loro utilità nella biologia quantitativa; argomenti questi che sono al di fuori dello scopo di questo articolo. Lo studio della relazione G×E, in senso lato, si è modificato negli ultimi decenni passando da tema di interesse marginale e per pochi ricercatori a qualcosa che non può più essere ignorato da nessun ecologo evolutivo. Le risposte plastiche a condizioni ambientali eterogenee, ben lontane dall'essere considerate un grattacapo, rappresentano uno dei fenomeni più comuni che caratterizzano il mondo vivente.
Bibliografia
Agrawal 2002: Agrawal, Anurag A. e altri, Ecological genetics of an induced plant defense against herbivores: additive genetic variance and costs of phenotypic plasticity, "Evolution", 56, 2002, pp. 2206-2213.
Bettencourt 2002: Bettencourt, Brian R. e altri, Response to natural and laboratory selection at the Drosophila HSP70 genes, "Evolution", 56, 2002, pp. 1796-1801.
Bradshaw 1965: Bradshaw, Amy D., Evolutionary significance of phenotypic plasticity in plants, "Advances in genetics", 13, 1965, pp. 115-155.
DeWitt 1998: DeWitt, Thomas J., Costs and limits of phenotypic plasticity: tests with predator-induced morphology and life history in a freshwater snail, "Journal of evolutionary biology", 11, 1998, pp. 465-480.
DeWitt 1998: DeWitt, Thomas J. - Sih, Andrew - Wilson, David S., Costs and limits of phenotypic plasticity, "Trends in ecology and evolution", 13, 1998, pp. 77-81.
Ergon 2004: Ergon, Torbjørn e altri, Optimal body size and energy expenditure during winter: why are voles smaller in declining populations?, "The American naturalist", 163, 2004, E442-E457.
Ernande, Dieckmann 2004: Ernande, Bruno - Dieckmann, Ulf, The evolution of phenotypic plasticity in spatially structured environments: implications of intraspecific competition, plasticity costs and environmental characteristics, "Journal of evolutionary biology", 17, 2004, pp. 613-628.
Falconer 1952: Falconer, Douglas S., The problem of environment and selection, "The American naturalist", 86, 1952, pp. 293-298.
Fischer 2004: Fischer, Markus e altri, Experimental life-history evolution: selection on growth form and its plasticity in a clonal plant, "Journal of evolutionary biology", 17, 2004, pp. 331-341.
Gromko 1995: Gromko, Mark H., Unpredictability of correlated response to selection: pleiotropy and sampling interact, "Evolution", 49, 1995, pp. 685-693.
Ho 2004: Ho, Melissa D. - Cannon, Bryan C. - Lynch, Jonathan, Optimization modeling of plant root architecture for water and phosphorous acquisition, "Journal of theoretical biology", 226, 2004, pp. 331-340.
Houle 1998: Houle, David, Genetic covariance of fitness correlates: what genetic correlations are made of and why it matters, "Evolution", 45, 1991, pp. 630-648.
Huber 2004: Huber, Heidrun e altri, Frequency and microenvironmental pattern of selection on plastic shade-avoidance traits in a natural population of Impatiens capensis, "The American naturalist", 163, 2004, pp. 548-563.
Köhler 2000: Köhler, Heinz-R. e altri, Selection favours low hsp70 levels in chronically metal-stressed soil arthropods, "Journal of evolutionary biology", 13, 2000, pp. 569-582.
Lewontin 1974: Lewontin, Richard C., The analysis of variance and the analysis of causes, "American journal of human genetics", 26, 1974, pp. 400-411.
Lyytinen 2004: Lyytinen, Anne e altri, Does predation maintain eyespot plasticity in Bicyclus anynana?, "Proceedings of the Royal Society B", 271, 2004, pp. 279-283.
Merila 2004: Merila, Juha - Laurila, Anssi - Lindgren, Beatrice, Variation in the degree and costs of adaptive phenotypic plasticity among Rana temporaria populations, "Journal of evolutionary biology", 17, 2004, pp. 1132-1140.
Pienaar, Greef 2003: Pienaar, Jason - Greef, Jaco M., Maternal control of offspring sex and male morphology in the Otitesella fig wasps, "Journal of evolutionary biology", 16, 2003, pp. 244-253.
Pigliucci 2001: Pigliucci, Massimo, Phenotypic plasticity: beyond nature and nurture, Baltimore-London, Johns Hopkins University Press, 2001.
Relyea 2002: Relyea, Rick A., Costs of phenotypic plasticity, "The American naturalist", 159, 2002, pp. 272-282.
Scheiner 1993: Scheiner, Samuel M., Genetics and evolution of phenotypic plasticity, "Annual review of ecology and systematics", 24, 1993, pp. 35-68.
Scheiner, Berrigan 1998: Scheiner, Samuel M. - Berrigan, David, The genetics of phenotypic plasticity VIII. The cost of plasticity in Daphnia pulex, "Evolution", 52, 1998, pp. 368-378.
Scheiner, Lyman 1989: Scheiner, Samuel M. - Lyman, Richard F., The genetics of phenotypic plasticity I. Heritability, "Journal of evolutionary biology", 2, 1989, pp. 95-107.
Schlichting 1986: Schlichting, Carl D., The evolution of phenotypic plasticity in plants, "Annual review of ecology and systematics", 17, 1986, pp. 667-693.
Schlichting, Smith 2002: Schlichting, Carl D. - Smith, Harry, Phenotypic plasticity: linking molecular mechanisms with evolutionary outcomes, "Evolutionary ecology", 16, 2002, pp. 189-211.
Shipley 2000: Shipley, Bill, Cause and correlation in biology: a user's guide to path analysis, structural equations and causal inference, Cambridge-New York, Cambridge University Press, 2000.
Stearns 1991: Stearns, Stephen - de Jong, Gerhardus J. - Newman, Robert A., The effects of phenotypic plasticity on genetic correlations, "Trends in ecology and evolution", 6, 1991, pp. 122-126.
Sultan 1987: Sultan, Sonia E., Evolutionary implications of phenotypic plasticity in plants, "Evolutionary biology", 21, 1987, pp. 127-178.
Via 1987: Via, Sara, Genetic constraints on the evolution of phenotypic plasticity, in: Genetic constraints on adaptive evolution, edited by Volker Loeschcke, Berlin-London, Springer, 1987, pp. 47-71.
West-Eberhard 1989: West-Eberhard, Mary Jane, Phenotypic plasticity and the origins of diversity, "Annual review of ecology and systematics", 20, 1989, pp. 249-278.
West-Eberhard 2003: West-Eberhard, Mary Jane, Developmental plasticity and evolution, New York-Oxford, Oxford University Press, 2003.
Tavola I
Plasticità gxe e altre fonti di confusione terminologica
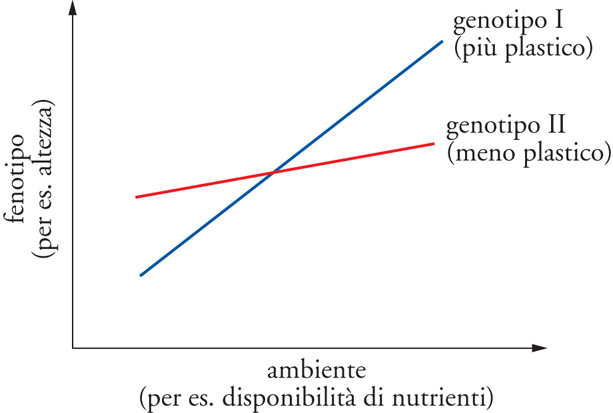
In questo articolo, ho provato a mantenere distinte le espressioni plasticità fenotipica e GxE, ma esiste una notevole confusione su questa distinzione, in particolare perché molti autori impiegano entrambe per significare due cose distinte, a seconda che si riferiscano a proprietà di genotipi individuali o di popolazioni di genotipi (tab. 1).
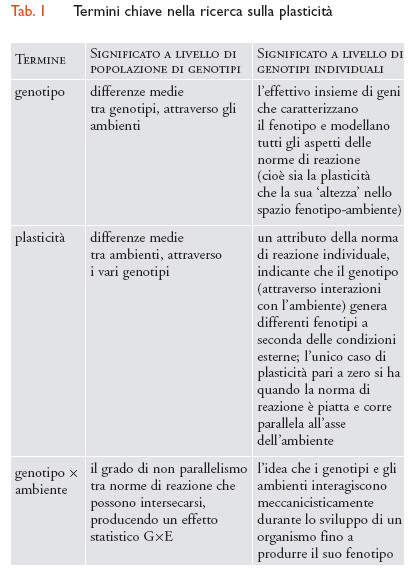
La fig. 1 mostra il semplice caso di una popolazione che comprende solo due genotipi. Le linee rappresentano le norme di reazione di ciascun genotipo (cioè le funzioni fenotipo-ambiente specifiche di un genotipo). Entrambe le linee presentano una pendenza nello spazio fenotipo-ambiente; il che significa che entrambi i genotipi sono plastici. La popolazione mostra GxE nel senso che esiste variazione genetica per la pendenza della norma di reazione, la quale può essere rilevata da una tipica analisi della varianza.
Fin qui ho utilizzato il termine plasticità per indicare le proprietà di norme individuali di reazione e GxE con riferimento a un attributo statistico dell’intera popolazione. Comunque si può anche parlare di plasticità come attributo non di un individuo ma di una popolazione, nel qual caso essa rappresenta una misura statistica di come la media fenotipica
attraverso i genotipi di un carattere cambia con l’ambiente (in fig. 1 questa media aumenta da sinistra verso destra, poiché entrambe le norme di reazione hanno una pendenza positiva). Inoltre, GxE può anche essere usato a livello del
singolo genotipo, nel qual caso rappresenta l’idea che i fenotipi sono il risultato di interazioni (meccanicistiche) in corso tra geni e ambienti durante lo sviluppo di un organismo. Una fonte di confusione deriva dall’assenza di una connessione necessaria tra questi termini quando ci si sposta dall’analisi di livello individuale a quella di livello popolazionale.
Per esempio, le relazioni GxE sono caratteristiche di qualsiasi genotipo individuale durante la sua ontogenesi; ma una popolazione di genotipi potrebbe avere norme di reazione dello sviluppo individuali somiglianti, che potrebbero non generare alcuna GxE statisticamente significativa a livello di popolazione. Questo è un esempio particolare del problema più generale posto dal fatto che il livello popolazionale (statistico) di descrizione potrebbe mancare del corrispettivo a livello individuale (puramente meccanicistico).