
Emisferi cerebrali
Emisferi cerebrali
di Henry Hecaen e Giovanni Berlucchi
EMISFERI CEREBRALI
Dominanza cerebrale
di Henry Hecaen
sommario: 1. Introduzione. 2. Emisfero sinistro. a) Dominanza dell'emisfero. b) Aprassia ideativa, aprassia ideomotrice, aprassia costruttiva. c) Dsturbi della percezione visiva. 3. Emisfero destro. a) Somatoagnosia e nosoagnosia. b) Agnosia spaziale. c) Aprassia costruttiva e altre aprassie. d) Disorientamenti spaziali e metamorfopsie. e) Dsturbi della percezione visiva. f) Disturbi della sfera uditiva. 4. Considerazioni generali. a) Natura della specializzazione emisferica. b) Localizzazione della specializzazione emisferica. c) Pevalenza manuale e specializzazione emisferica funzionale. d) Origine della specializzazione emisferica. □ Bibliografia.
1. Introduzione
Sembra che si debba far risalire a M. Dax, e quindi al 1836, la prima indicazione di una possibile specializzazione di uno dei due emisferi cerebrali per una funzione mentale superiore. Ma la nozione di dominanza cerebrale è stata stabilita con certezza solo trenta anni più tardi, appena qualche anno dopo che era stato ammesso il principio della localizzazione funzionale corticale. In effetti, anche se nella sua pubblicazione del 1861 P. Broca localizzò la facoltà del linguaggio articolato a livello del piede della terza circonvoluzione frontale, dovettero passare quattro anni prima che egli si ritenesse in diritto di affermare la prevalenza dell'emisfero sinistro per questa facoltà (v. Broca, 1861 e 1865). Egli infatti si rifiutò sempre di conferire attribuzioni diverse alle due metà dell'encefalo. In effetti, l'ammettere una differenza funzionale tra due organi pari e simmetrici, quali i due emisferi, avrebbe violato, a suo giudizio, una legge fisiologica sino ad allora senza eccezione. Broca metteva in relazione questa lateralizzazione sinistra con la predominanza manuale destra, caratteristica comune alla maggior parte degli uomini. E in funzione di questa predisposizione organica, che implica che un destrimane abbia un sinistrismo cerebrale, che l'emisfero sinistro si sviluppa più precocemente e diviene quindi più adatto a dirigere l'esecuzione e la coordinazione di quegli atti sia intellettuali sia muscolari che costituiscono il linguaggio articolato: nasce così l'abitudine di parlare con l'emisfero sinistro. Ma non si tratta che di linguaggio articolato, poiché, per Broca, la funzione del linguaggio in generale appartiene a entrambi gli emisferi. Inoltre, secondo Broca, in un piccolo numero di individui la predominanza innata delle forze motrici dell'emisfero destro, dando una prevalenza alle funzioni della mano sinistra, inverte l'ordine dei fenomeni: questi individui avranno, al contrario della maggioranza, un destrismo cerebrale. A partire da queste constatazioni, si finì più o meno implicitamente per considerare il cervello sinistro del destrimane come l'emisfero dominante per tutte le funzioni simboliche. In seguito, i lavori di Liepmann (v., 1900) sulla aprassia confermarono questo modo di vedere mettendo in luce l'importanza dell'emisfero sinistro nella realizzazione della funzione gestuale. Per quel che concerne i disturbi della gnosia visiva, si inferiva da questa ipotesi, anziché da documenti anatomoclinici, la lateralizzazione della lesione a sinistra. Si riconosceva così a uno dei due emisferi - detto dominante - una funzione esclusiva in ragione dei centri specializzati che esso possiede.
Durante questo periodo del tutto iniziale, solo due autori, il famosissimo H. Jackson (v., 1932) e il quasi sconosciuto Th. D. Dunn (v., 1895), riconoscevano una funzione speciale all'emisfero destro. Le prime descrizioni di disturbi nel riconoscimento del corpo (v. Babinski, 1914; v. Schilder, 1935; v. Pötzl, 1928; ecc.) indussero a considerare la possibilità per questo emisfero di assumere più specificamente alcune funzioni.
Poco dopo, durante il periodo tra le due guerre mondiali, alcuni clinici come Reichardt, Lange, Pötzl, Dide furono colpiti dalla particolare importanza dell'emisfero destro per le funzioni spaziali. Le loro intuizioni resteranno di fatto isolate; ricerche più precise non sono state effettuate che molto più tardi con i lavori anatomoclinici di R. Brain (v., 1941), di McFie e altri (v., 1950), di Hecaen e altri (v., 1951 e 1956).
D'altronde questi primi lavori incontrarono agli inizi delle forti resistenze, ma numerose ricerche successive confermarono i primi risultati. Naturalmente i nuovi dati così raccolti sollevavano altri problemi che a loro volta richiedevano nuovi orientamenti della ricerca, facilitati d'altronde dagli sviluppi tecnici.
I dati attualmente raccolti provengono anche dallo studio clinico o sperimentale di grandi serie di malati con lesioni unilaterali, da osservazioni compiute nei soggetti che avevano subito la sezione delle commissure neocorticali e anche da ricerche sperimentali realizzate nei soggetti normali (ascolto dicotico, presentazione tachistoscopica e in seguito registrazione dei potenziali evocati). I dati anatomici, per molto tempo inesistenti, cominciano ad accordarsi con i fatti anatornoclinici e sperimentali con il lavoro di Geschwind e Levitsky (v., 1968).
Tenendo conto dei vari risultati presenteremo, in modo necessariamente schematico, le funzioni proprie di ciascun emisfero.
2. Emisfero sinistro
a) Dominanza dell'emisfero
La dominanza dell'emisfero sinistro per la funzione del linguaggio, una volta affermata da Broca, non fu più messa in discussione, a maggior ragione in quanto i lavori di Wernicke (v., 1874) dimostrarono ben presto che la lesione di una zona più posteriore dello stesso emisfero era in grado di impedire la comprensione del linguaggio. L'integrità della comprensione verbale anche in presenza di lesioni anteriori sinistre era parsa a Broca uno degli argomenti essenziali in favore della partecipazione dei due emisferi alla funzione generale del linguaggio. Le scoperte successive consentirono di supporre una rappresentazione relativamente precisa delle funzioni del linguaggio sull'emisfero sinistro. La sistematizzazione dell'‛area del linguaggio', operata da Dejerine antecedentemente alla prima guerra mondiale, non è stata più fatta oggetto di modificazioni di rilievo. Solamente alla nozione di ‛centro' si è sostituita quella più ampia di ‛zona funzionale'. I fatti di recupero, spesso notevoli, si comprendono meglio se si ammette un'organizzazione funzionale secondo zone esclusivamente preferenziali, in luogo di una rappresentazione di meccanismi molto limitati e rigidi. Bisogna tuttavia segnalare che la partecipazione, d'altronde assai limitata, dell'emisfero di destra alle funzioni del linguaggio è stata dimostrata sia dai risultati di alcune emisferectomie sinistre in destrimani (v. Smith, 1966) sia, soprattutto, con lo studio dei soggetti con sezione delle commissure cerebrali (split brain). In questi soggetti, limitando le afferenze a un solo emisfero, Sperry e Gazzaniga (v., 1967) hanno potuto dimostrare, utilizzando risposte non verbali, che l'emisfero di destra, malgrado un'impossibilità di espressione, era capace di riconoscere materiale verbale presentato oralmente o per scritto o offerto alla discriminazione tattile. Per contro, l'emisfero di destra non ha che capacità molto limitate per comprendere le dimensioni sintattiche (v. Gazzaniga e Hillyard, 1971).
Bisogna anche sottolineare che lo studio delle asimmetrie sensoriali nei soggetti normali ha recato importanti precisazioni. Infatti D. Kimura (v., 1961) utilizzando il metodo di ascolto biaurale di Broadbent ha potuto dimostrare nei soggetti normali la superiorità dell'emisfero dominante nel trattare il materiale verbale, sia esso significativo o meno. Con lo stesso metodo Shankweiler e Studdert-Kennedy (v., 1967) hanno dimostrato che questa superiorità si manifesta a partire dai livelli fonemici o subfonemici.
b) Aprassia ideativa, aprassia ideomotrice, aprassia costruttiva
Dalle lesioni di questo emisfero dipendono anche tre tipi di disturbi del gesto, indipendententi dalle disfunzioni della motricità elementare: l'aprassia ideativa, l'aprassia ideomotrice e l'aprassia costruttiva. L'aprassia ideativa riguarda i disturbi nella manipolazione degli oggetti, mentre nell'aprassia ideomotrice sono alterati i gesti del simbolo convenzionale, i gesti espressivi o descrittivi. La relazione di queste aprassie con i disturbi del linguaggio è ancora mal definita, ma sembra che esse possano manifestarsi, sebbene assai raramente, indipendentemente dai disturbi del linguaggio rivelando alterazioni di un codice particolare di comunicazione o di una funzione di programmazione delle attività.
L'aprassia costruttiva, anche se inizialmente descritta nel caso di lesioni dell'emisfero sinistro, è ugualmente prodotta da lesioni dell'emisfero destro. Il soggetto è incapace di disegnare, di costruire, di copiare semplici modelli, geometrici o figurativi. Questo disturbo può essere totalmente indipendente dai disordini di linguaggio. Benché la questione non sia definitivamente risolta, l'aprassia costruttiva dovuta a lesioni dell'emisfero sinistro sembrerebbe avere caratteristiche diverse da quelle causate da lesioni a destra. Per Warrington e altri (v., 1966) e Hecaen e Assai (v., 1970) essa riflette un disturbo delle attività di programmazione che si esprime nell'ambito della costruzione, come quello che si manifesta nelle sequenze gestuali richieste per la manipolazione degli oggetti (aprassia ideativa). In effetti, quando un malato affetto da una lesione a sinistra presenta dei deficit nel copiare un modello, si constata un miglioramento delle sue prestazioni allorché disponga di elementi di riferimento previamente tracciati.
c) Disturbi della percezione visiva
I disturbi della percezione visiva susseguenti a lesioni di questo emisfero riguardano essenzialmente il riconoscimento del significato degli oggetti, dei colori e dei segni grafici, mentre i dati visivi elementari sono ben rilevati. Per alcuni, tuttavia, l'importanza dei disturbi del linguaggio sarebbe essenziale. La mancanza del riconoscimento del significato dell'oggetto, benché limitata alla presentazione ottica, dimostrerebbe sia un'impossibilità di denominazione (afasia ottica: v. Freund, 1888; v. Geschwind, 1965), sia una impossibilità di associare le qualità visive dell'oggetto (v. De Renzi e altri, 1969).
Per la modalità visiva, il metodo tachistoscopico con presentazione, simultanea o successiva, negli emicampi ha permesso di rilevare nei soggetti normali la superiorità dell'emicampo destro, cioè dell'emisfero sinistro, nell'elaborazione del materiale verbale.
3. Emisfero destro
a) Somatoagnosia e nosoagnosia
Lo studio dei disturbi del riconoscimento del corpo, designati in generale come somatoagnosia, consentì di mettere in evidenza la specificità funzionale dell'emisfero destro, precedentemente considerato muto. Nel caso di lesioni a destra, si constatano in effetti comportamenti particolari dei malati: il soggetto nega il deficit motorio che ha colpito il suo lato sinistro (nosoagnosia), si comporta come se ignorasse l'esistenza di questo lato (emisomatoagnosia), o può addirittura attribuirne l'appartenenza ad altri. Benché tali sintomi non sopravvengano in genere che nei soggetti con disturbi della coscienza e che presentano disturbi di senso in metà del corpo, il fatto che non siano che eccezionalmente constatati in caso di lesioni a sinistra obbliga a considerare che una regione dell'emisfero destro, il complesso parieto-temporo-occipitale, intervenga nell'integrazione dei dati sensoriali provenienti da una metà del corpo. A seguito della distruzione di questa zona, l'emicorpo è ignorato dal malato o considerato addirittura estraneo a lui stesso.
b) Agnosia spaziale
L'importanza predominante dell'emisfero destro per le funzioni spaziali, suggerita dai cimici nel periodo tra le due guerre mondiali, è stata ora confermata da numerose ricerche condotte sia su gruppi di malati con lesioni emisferiche unilaterali o su soggetti con sezione delle commissure cerebrali (split brain), sia su soggetti normali sottoposti a una presentazione tachistoscopica (v. Durnford e Kimura, 1971), misurando i tempi di reazione secondo il campo e secondo il materiale (v. Rizzolatti e altri, 1971); per quel che concerne la modalità tattile, è stata anche dimostrata l'importanza dell'emisfero destro quando la prova ha un carattere spaziale (v. Benton e altri, 1973; v. De Renzi e altri, 1969).
È noto, in patologia, in seguito alla pubblicazione di R. Brain (v., 1941), che l'agnosia della metà dello spazio controlaterale alla lesione (agnosia spaziale unilaterale) dipende soprattutto dalle lesioni di questo emisfero. Altri lavori, in particolare quelli di De Renzi e altri (v., 1970), hanno dimostrato che questo disturbo non è limitato alla modalità visiva ma riguarda una rappresentazione dello spazio indipendente dalle modalità sensoriali.
c) Aprassia costruttiva e altre aprassie
L'aprassia costruttiva determinata dalle lesioni dell'emisfero destro appare non solamente più frequente e in generale più grave di quella dipendente dalle lesioni dell'emisfero dominante, ma presenta caratteristiche che rivelano l'importanza dei fattori visuo-spaziali. A differenza di quello che avviene nel caso dei disturbi prassici determinati dalle lesioni dell'emisfero sinistro, la presenza del modello e dei riferimenti non migliora la prestazione costruttiva del malato con lesione a destra.
Gazzaniga, Bogen e Sperry (v., 1965) hanno anche constatato che i soggetti con sezione delle commissure emisferiche (split brain), per quanto destrimani, copiano correttamente un cubo di Necker e mettono insieme dei cubi o i pezzi di un rompicapo in modo più esatto con la mano sinistra che con la mano destra. Poiché questi soggetti accoppiano correttamente con la mano destra il disegno che funge da stimolo con il modello che gli corrisponde e non presentano difficoltà motorie della mano destra, si deve ammettere che il deficit nel copiare sia situato fra il sistema visivo e il sistema motorio.
Oltre all'aprassia costruttiva, le lesioni dell'emisfero destro possono provocare un altro tipo di aprassia, l'aprassia del vestirsi, nella quale l'alterazione sembra tradurre un'impossibilità di articolare lo spazio corporeo e quello extracorporeo.
d) Disorientamenti spaziali e metamorfopsie
Allo stesso modo i diversi disorientamenti spaziali come il disturbo delle nozioni topografiche (difficoltà d'orientamento su una mappa o su una carta geografica) e la perdita della memoria topografica perdita dell'orientamento in luoghi ben noti al soggetto) sembrerebbero rivelare principalmente lesioni dell'emisfero destro, sebbene sia difficile escludere formalmente una partecipazione, anche limitata, di quello sinistro.
La disorganizzazione spaziale si rivela in modo netto nella scrittura, nella lettura e nel calcolo. I disturbi grafici, lessicali e di calcolo determinati da lesioni dell'emisfero destro influiscono sull'organizzazione spaziale degli elementi grafici (iterazione di lettere o di trattini, trascuratezza della parte sinistra del testo da ricopiare o da leggere, errata disposizione delle cifre nelle operazioni aritmetiche) e non sul riconoscimento e il mantenimento dei codici (v. Hecaen e Marcie, 1974).
Anche manifestazioni parossistiche come le illusioni ottiche di deformazione degli oggetti e dello spazio (metamorfopsie) dipendono con frequenza significativa da lesioni di destra (v. Teuber e altri, 1960; v. Mullan e Penfield, 1959; v. Hecaen e Angelergues, 1965).
Queste diverse ricerche confermano pertanto la predominanza dell'emisfero destro nelle funzioni spaziali e allo stesso tempo fanno pensare che i disturbi provocati dalle lesioni di questo emisfero non siano limitati alla modalità visiva, ma riflettano una disorganizzazione sopramodale.
Non si deve nascondere, tuttavia, l'esistenza di alcune discordanze nei dati raccolti. Così Semmes e altri (v., 1960) nella serie di cerebrolesi da loro studiata hanno constatato che i deficit in una prova di ricerca di itinerari con l'aiuto di riferimenti visivi o tattili erano significativamente maggiori soltanto nel caso di lesioni parietali, ma indipendentemente dal lato emisferico. Al contrario, B. Milner (v., 1965) e S. Corkin (v., 1965), utilizzando una prova di apprendimento di labirinto (da seguire con uno stiletto con punti di riferimento visivi o tattili), non hanno trovato deficit se non nel caso di lesioni dell'emisfero destro (lobectomia temporale). F. Newcombe (v., 1969) ha ottenuto in feriti di guerra i medesimi risultati con una prova molto simile. La differenza tra i due compiti, locomotorio in un caso, apprendimento del labirinto nell'altro, e la differenza tra le eziologie delle lesioni nelle due serie potrebbero essere responsabili di questa diversità dei risultati. Tuttavia, utilizzando un compito locomotorio di ricerca di itinerari, Hecaen e altri (v., 1972) hanno constatato che in una serie di soggetti con lesioni emisferiche unilaterali, il gruppo con lesioni a destra e, nell'ambito di questo, il gruppo con lesioni parietali, presentava un deficit significativamente superiore a quello dei soggetti con lesioni a sinistra. Ma bisogna anche sottolineare che i punteggi dei soggetti con lesioni a sinistra erano significativamente inferiori a quelli dei soggetti di controllo. Infine, nella serie di G. Ratcliff e F. Newcombe (v., 1973), solo le lesioni bilaterali posteriori provocavano dei deficit in una prova locomotoria di ricerca di itinerari, anche se in alcuni di questi casi si potevano constatare dei buoni risultati nell'apprendimento del labirinto.
De Renzi e altri (v., 1969, 1970 e 1971) hanno sottoposto gruppi di malati con lesioni emisferiche unilaterali a una serie di prove spaziali di complessità crescente. Nei compiti elementari, quali percezione della direzione spaziale (v. De Renzi e altri, 1971) e visione della profondità con le sole indicazioni della disparità (v. Carmon e Bechtoldt, 1969; v. Benton e Hecaen, 1970), si constatavano deficit solo nei casi di lesioni posteriori a destra. Al contrario, in compiti alquanto più complessi i deficit si ritrovano in tutti i gruppi, con lesioni sia a destra sia a sinistra; essi sono solamente più marcati nel caso di lesioni a destra, ma non in modo significativo. Infine, nei compiti ancora più complessi (riconoscimento di figure presentate in posizione rovesciata, ricordo della localizzazione spaziale di un oggetto), i deficit sono simili nei due gruppi con lesioni posteriori a destra e a sinistra.
De Renzi (v., 1974) riconosce una serie di livelli nelle disorganizzazioni spaziali: solamente nei compiti più elementari si trova una dominanza manifesta dell'emisfero destro, che può anche essere assoluta. Nelle prove meno elementari il contributo di ogni emisfero potrebbe essere diverso: spaziale per l'emisfero destro, verbale per l'emisfero sinistro.
L'ipotesi di De Renzi; sebbene molto seducente, urta contro due ordini di difficoltà. Da una parte è stato constatato, sia pure invero con tecniche diverse, che le deviazioni delle coordinate visive, significativamente più marcate nel caso di lesioni posteriori a destra, sono altresì significativamente più importanti negli altri gruppi con lesioni a destra o a sinistra che nei soggetti di controllo (v. Tzavaras e Hecaen, 1972). D'altra parte, si è visto che l'orientamento spaziale, studiato con prove così complesse come la ricerca di itinerari (v. Hecaen e altri, 1972) o l'apprendimento del labirinto (v. Milner, 1965; v. Newcombe, 1969), è alterato in modo predominante dalle lesioni a destra.
Va rilevato, tuttavia, che questa ipotesi è in accordo con il fatto che i soggetti con lesioni a sinistra si comportano come i soggetti normali quando si tratta di valutare la profondità con i soli indici di disparità, e con la presenza, nel caso di lesioni a sinistra, di deficit dell'orientamento topografico. È dunque importante determinare quali siano i fattori ‛intellettuali' e ‛verbali' suscettibili di intervenire nel caso di lesioni a sinistra per provocare i deficit spaziali di questo tipo.
Non bisogna inoltre dimenticare il possibile intervento di un fattore da lesioni frontali dell'uno o dell'altro emisfero in certe disorganizzazioni spaziali (v. Cohen, 1959; v. Albert e Hecaen, 1971).
e) Disturbi della percezione visiva
Sembra ugualmente evidente il ruolo dell'emisfero destro in certi disturbi della percezione visiva: che si tratti di riconoscimento, di apprendimento o di memoria concernente le forme complesse, sono stati constatati deficit nei malati con lesioni di questo emisfero (v. Kimura, 1963; v. Milner, 1971; v. De Renzi e altri, 1966, 1968 e 1969).
Le ricerche di Levy, Trevarthen e Sperry (v., 1972) nei soggetti con sezione delle commissure emisferiche (split brain) hanno confermato la capacità percettiva superiore dell'emisfero destro, quali che siano i caratteri del materiale presentato: elementi non significativi, oggetti familiari o visi umani.
I dati anatomoclinici avevano fatto pensare che un tipo particolare di agnosia visiva, la prosopagnosia o agnosia per le fisionomie individuata da Bodamer (v., 1947), di- pendesse principalmente da lesioni dell'emisfero destro (v. Hecaen e Angelergues, 1962). Tuttavia non esiste ancora alcun caso assolutamente decisivo per escludere formalmente l'importanza della bilateralità delle lesioni. Numerose ricerche sperimentali condotte su serie di malati con lesioni unilaterali hanno dimostrato l'importanza predominante dell'emisfero destro nel riconoscimento, nella memorizzazione o nell'appaiamento dei visi umani. Nei soggetti normali ricerche come quelle di Rizzolatti, Umiltà e Berlucchi (v., 1971) hanno dimostrato una maggiore rapidità di discriminazione per le facce presentate nell'emicampo visivo sinistro rispetto all'emicampo destro. Sebbene per De Renzi e altri (v., 1968) il disturbo nel riconoscimento dei visi umani sia un aspetto del deficit della percezione delle forme complesse, alcuni hanno sostenuto l'ipotesi di un disordine legato a quel materiale squisitamente specifico che è il viso umano (v. Yin, 1970; v. Tzavaras e altri, 1970).
f) Disturbi della sfera uditiva
Per quanto riguarda la sfera uditiva vi sono relativamente pochi elementi anatomoclinici precisi sull'agnosia dei suoni o sull'amusia. Le ricerche di B. Milner (v., 1962) per mezzo del test di Seashore hanno dimostrato che le lobectomie temporali eseguite sull'emisfero destro provocano una diminuzione dei punteggi relativi alla percezione dell'altezza del suono e alla memoria melodica. È anche stato dimostrato da Shankweiler (v., 1966) un deficit nel riconoscimento delle melodie familiari nel caso di lobectomie temporali a destra. Vignolo (v., 1969), basandosi sui risultati ottenuti sottoponendo gruppi di malati con lesioni emisferiche unilaterali, a destra o a sinistra, a due tipi di prove uditive, è pervenuto alla seguente sistematizzazione: nel caso di lesioni a destra è alterata la discriminazione degli insiemi sonori complessi, mentre nel caso di lesioni a sinistra è alterato il riconoscimento dei suoni significativi.
Nei soggetti normali, l'utilizzazione del metodo dicotico ha rivelato una superiorità dell'orecchio sinistro, cioè dell'emisfero destro, nel riconoscimento di melodie (v. Kimura, 1964), dei suoni sonar (v. Chaney e Webster, 1966), degli accordi (v. Gordon, 1970), dei rumori familiari (v. Curry 1967), dei suoni vocali umani non verbali (v. Kimura, 1973).
4. Considerazioni generali
Questa presentazione necessariamente sommaria delle nostre conoscenze sulla specializzazione funzionale emisferica potrebbe essere riassunta nella seguente formula: l'emisfero sinistro presiede essenzialmente alle funzioni verbali e di astrazione, l'emisfero destro alle funzioni averbali percettivo-spaziali. Appare ora necessario ricordare i problemi generali che pone l'interpretazione di questi dati e le vie verso le quali sembrerebbero indirizzarsi le ricerche.
a) Natura della specializzazione emisferica
Il primo di questi problemi è quello della natura di questa specializzazione e della complementarietà che esiste tra i due emisferi nelle prestazioni dei soggetti normali.
Per alcuni autori, come Milner, la differenza funzionale tra i due emisferi sarebbe connessa alla natura verbale o non verbale del materiale che ciascuno di essi vaglia, qualunque sia la modalità di presentazione e qualunque sia il compito richiesto (riconoscimento, apprendimento, memoria). Recenti lavori di P. M. Corsi (v., 1972) dimostrano così una dissociazione nelle prove di memoria a breve termine con materiale verbale o non verbale (posizione) a seconda che il danno all'ippocampo sia sul lato destro o sul sinistro. Del pari, solamente nelle lesioni frontali localizzate a un lato esiste un deficit nell'apprendimento del labirinto in conseguenza del mancato rispetto delle regole che presiedono a questo apprendimento (v. Milner, 1965). Vi sono ugualmente dei deficit nella prova di giudizio del tempo di presentazione di un oggetto; in effetti il deficit non si constata nel caso di lesioni frontali a sinistra se la serie di oggetti ha carattere verbale e nel caso di lesioni frontali a destra se la serie di oggetti ha caratteri non verbali (v. Milner, 1971).
Al contrario, per altri autori, questa differenza funzionale interemisferica risiederebbe nel modo con cui ogni emisfero elabora l'informazione. Partendo da uno studio sulle agnosie visive, già nel 1963, con Angelergues avevamo prospettato l'ipotesi che i deficit propri delle lesioni dell'emisfero destro dipendano da un deficit di individuazione di oggetti simili in una data categoria, mentre quelli delle lesioni a sinistra riflettano un deficit della categorizzazione (v. Hecaen e Angelergues, 1963). De Renzi e altri (v., 1969) hanno attribuito all'emisfero destro un ruolo di discriminazione e di organizzazione dei dati sensoriali, mentre all'emisfero sinistro sarebbe devoluto il compito di denominare e associare questi dati per permettere la comprensione del significato. De Renzi e altri (v., 1969) e Vignolo (v., 1969) sostengono così l'esistenza di due livelli di agnosia, indipendentemente dalla modalità sensoriale, riprendendo pertanto la suddivisione tra agnosia percettiva e agnosia associativa proposta da Lissàuert (v., 1890). Le lesioni emisferiche a destra provocherebbero il primo tipo di agnosia; il disturbo si manifesterebbe allorché il compito percettivo comporta una certa complessità. Il deficit da lesioni a sinistra - agnosia associativa - consisterebbe in una impossibilità di associare gli attributi al loro specifico oggetto (colori, rumori, forma).
Con lo studio dei soggetti con sezione delle commissure emisferiche (split brain) si è potuta riprendere la questione del modo particolare di elaborazione dei dati sensoriali proprio di ogni emisfero. In effetti, l'esame di questi soggetti permette di giudicare le prestazioni fornite da ognuno degli emisferi non lesi, costituendo inoltre ciascuno dei due un controllo nel confronto con l'altro.
Per R. D. Nebes (v., 1971) se all'emisfero di destra spetta il compito di organizzare l'informazione sotto forma di totalità, esso deve per questo rivelarsi superiore in tutti i compiti che richiedono una concettualizzazione della configurazione totale dello stimolo a partire da informazioni frammentarie non facilmente verbalizzabili. Due esperimenti dimostrano in effetti, secondo questo autore, che la capacità dell'emisfero destro di associare una parte al tutto si rivela superiore a quella dell'emisfero sinistro qualunque sia la modalità di presentazione e la natura intermodale o intramodale del compito.
J. Levy, C. Trevarthen e R. W. Sperry (v., 1972) hanno comparato la capacità di percepire e di rispondere dell'uno e dell'altro emisfero utilizzando la tendenza di ciascuno di essi a effettuare un completamento percettivo di modelli attraverso la linea mediana. Gli stimoli consistevano in figure ‛chimeriche' formate unendo la parte destra di una figura alla parte sinistra di un'altra, presentate tachistoscopicamente: visi umani, immagini nere non denominabili con tipi di ramificazioni, figure familiari, figure disposte in catena composte di croci e quadrati. In ragione del completamento che ciascun emisfero fa di quel che vede, esiste nel caso delle presentazioni di queste figure ‛chimeriche' un conflitto tra gli emisferi. La risposta alle presentazioni si dava indicando, sia con la mano destra, sia con quella sinistra, la figura completa e infine con la denominazione senza la presenza di immagini. La superiorità dell'emisfero destro si affermava in tutte le prove comportanti una risposta mediante designazione manuale, anche se le figure chimeriche consistevano in fotografie di visi, in immagini non denominabili a organizzazione complessa, in rappresentazioni di oggetti familiari o in figure disposte in catena. Soltanto quando veniva richiesta una risposta verbale si manifestava una superiorità dell'emisfero sinistro.
Dunque queste constatazioni sono in favore di una superiorità dell'emisfero destro nella capacità percettiva primitiva che costituisce la percezione delle forme, mentre l'emisfero sinistro sembrerebbe mal organizzato per compiere questo apprendimento percettivo.
Allorché è richiesta una risposta verbale, l'emisfero sinistro diventa dominante: l'informazione è allora elaborata differentemente, in modo più analitico, con la ricerca di caratteristiche distintive alle quali si possa dare un'etichetta verbale. Per Levy, Trevarthen e Sperry (v., 1972) non si tratta di una semplice predominanza nella competizione tra i due emisferi ma di una differenza fondamentale nel loro modo di elaborare l'informazione.
J. Levy (v., 1974) proseguendo questo tipo di ricerca con la stessa tecnica ha osservato, in presenza ditali figure, che allorché veniva realizzata una categorizzazione concettuale nell'appaiamento, essa era sempre effettuata dall'emisfero sinistro (in condizioni particolari l'emisfero destro che spontaneamente aveva la tendenza a operare una categorizzazione visiva - appaiamento per somiglianza di forme - riusciva anche a realizzare delle categorizzazioni concettuali).
Trevarthen (v., 1974, p. 202), partendo da questi diversi esperimenti, definisce la funzione dell'emisfero sinistro secondo la seguente formula: ‟tutte le esigenze di denominare l'oggetto, di pensare con parole senza parlare, o di ragionare sul significato dell'oggetto in un contesto di oggetti con diverse apparenze fanno sì che l'emisfero sinistro assuma il comando della risposta".
Così gli emisferi si rivelano capaci di trattare l'informazione in modo diverso a seconda delle esigenze della situazione. L'individuo, variando le sue strategie, può utilizzare un emisfero o l'altro per arrivare a risultati in apparenza simili, ma ottenuti con meccanismi diversi. A seconda delle precedenti esperienze l'individuo può utilizzare una strategia piuttosto di un'altra e con questa privilegiare l'attività di un emisfero.
A questo riguardo è dimostrativo un esperimento di Bever e Chiarello (v., 1974). Questi autori hanno in effetti dimostrato che nell'ascolto dicotico in soggetti senza educazione musicale il riconoscimento melodico era, come aveva dimostrato Kimura (v., 1964), superiore nell'orecchio sinistro, mentre nei musicisti era l'orecchio destro che consentiva di ottenere i migliori punteggi. In assenza di educazione musicale il riconoscimento delle melodie è effettuato dalla conformazione melodica generale intesa come Gestalt. Al contrario i musicisti operano un trattamento analitico dell'informazione valutando la combinazione degli elementi che compongono la melodia: è allora chiamata in causa l'attività dell'emisfero sinistro.
Questi risultati concordano d'altronde con quelli di Papçun e altri (v., 1974) sulla percezione di sequenze di lettere in codice Morse. In effetti, in operatori abili e in ascoltatori che non conoscevano il codice Morse era evidente una netta superiorità dell'orecchio destro, cioè dell'emisfero sinistro, per le sequenze comprendenti non più di 7 elementi. Ma per le sequenze con un più elevato numero di elementi, la superiorità dell'orecchio destro persisteva soltanto negli operatori esercitati, in quanto negli ascoltatori inesperti si manifestava la superiorità dell'orecchio sinistro. Gli autori postulano anche che questi ultimi soggetti operino un cambiamento di strategia quando le sequenze diventano più lunghe: incapaci, a causa della loro inesperienza, di valutare gli elementi individuali, essi si basano sui caratteri globali dello stimolo.
Così, partendo dalla competizione rivelata nei soggetti con sezione delle commissure emisferiche (split brain), si capisce meglio come la complementarità tra i due emisferi possa realizzarsi nel soggetto normale grazie all'azione del corpo calloso che permette l'unificazione dell'informazione elaborata in maniera diversa da ciascun emisfero. Un'elaborazione parallela dell'informazione può essere effettuata per certe prestazioni, mentre un'elaborazione più lenta e diversa può sopravvenire secondariamente nell'altro emisfero per assicurare il mantenimento della prestazione (v. Dimond e Beaumont, 1974).
b) Localizzazione della specializzazione emisferica
Questa particolare capacità di ciascuna metà cerebrale è propria dell'emisfero nel suo insieme o dipende da certe zone più o meno definite di ognuno dei due emisferi? Le ricerche anatomiche rivelano che una zona dell'emisfero sinistro, il planum temporale, è più larga di quella corrispondente dell'emisfero destro (v. Geschwind e Levitsky, 1968). Non è ancora stato possibile trovare sull'emisfero destro una zona particolare che sia più estesa in questo emisfero che nel controlaterale e che traduca quindi una capacità funzionale superiore. Esiste anche un certo numero di dati in favore di una diversa distribuzione delle aree funzionali nei due emisferi: maggior focalizzazione delle diverse regioni funzionali con presentazione bilaterale della parte sinistra del corpo, per converso maggiore diffusione a destra con rappresentazione controlaterale del corpo (v. Semmes e altri, 1960; v. Hecaen e Angelergues, 1963; v. Semmes, 1965). Se, a quanto sembra, è possibile ammettere che certe regioni definite di ciascun emisfero siano sede di funzioni specifiche, bisogna notare la partecipazione, per quanto più debole, delle regioni vicine che permette di postulare l'esistenza di un gradiente funzionale intraemisferico.
I risultati delle stimolazioni sottocorticali sembrano confermare ugualmente bene che la specializzazione si estende alle funzioni sottocorticali. Ojemann e altri hanno determinato con stimolazioni a livello del talamo (pulvinar e parte posterocentrale mediana del nucleo ventrolaterale) del lato sinistro, e non di quello destro, risposte di tipo afasico e deficit della memoria non verbale a breve termine (v. Ojemann e altri, 1968; v. Ojemann 1971). Disturbi della memoria non verbale a breve termine sono stati prodotti con la stimolazione del fascio mammillo-talamico e della ‛zona incerta' del lato destro, ma la memoria verbale non era alterata da queste stimolazioni. Si può così pensare che a fianco della rappresentazione orizzontale intraemisferica esista una rappresentazione verticale: le formazioni sottocorticali interverrebbero nella realizzazione delle prestazioni dovute a meccanismi nervosi dell'emisfero con cui esse sono in rapporto. Si verrebbe così a creare un sistema funzionale cortico-sottocorticale proprio di ciascuna metà del cervello.
Bisogna anche porsi il problema delle relazioni tra le funzioni a rappresentazione ben lateralizzata, di carattere strumentale, e le funzioni fondamentali di memoria e di apprendimento, di motivazione, d'attenzione, di programmazione, ecc., che sembrano dipendere da meccanismi organizzati bilateralmente. Ricerche come quelle di Muner e dei suoi collaboratori (v. Corsi, 1972) sugli effetti specifici delle lesioni dell'ippocampo permettono già di intravedere queste interazioni funzionali.
c) Prevalenza manuale e specializzazione emisferica funzionale
Lo studio dei rapporti tra prevalenza manuale e specializzazione emisferica funzionale ha anche tratto vantaggio, naturalmente, sia dagli apporti tecnici della psicologia sperimentale, sia, in patologia, dal metodo di Wada (iniezione intracarotidea di amobarbitalsodico che inattiva per breve tempo l'emisfero del lato dove è stata fatta l'iniezione). Sebbene Broca (v., 1865), stabilendo la predominanza dell'emisfero sinistro nei destrimani, ammettesse che i mancini parlino con l'emisfero destro, ben presto la presenza di osservazioni in contrasto con questa regola, compiute quasi unicamente su mancini, indusse a parlare di afasia crociata. In effetti, soltanto dopo la pubblicazione di Chescher del 1936 si arrivò a pensare a una organizzazione cerebrale delle rappresentazioni del linguaggio nel mancino non inversa rispetto a quella del destrimane, ma diversa, con rappresentazione ripartita in modo quasi uguale nei due emisferi. Alcuni autori avevano pensato che, qualunque fosse la prevalenza manuale, la rappresentazione del linguaggio dovesse essere sempre situata a sinistra. Questa affermazione è smentita dai fatti: se vi è effettivamente un numero rilevante di mancini che presentano disturbi afasici dopo lesioni della zona sinistra del linguaggio, circa il 20% o il 30% dei mancini diventa ugualmente afasico dopo lesioni della zona simmetrica situata sul lato destro. Non vi è dunque una relazione diretta e necessaria tra prevalenza manuale e lateralizzazione cerebrale delle funzioni del linguaggio.
D'altra parte il recupero dei disturbi del linguaggio avviene in modo più completo e rapido nei soggetti mancini, qualunque sia la lateralizzazione della lesione. Infine, la sintomatologia riscontrata dopo una lesione emisferica unilaterale nei soggetti mancini, per quanto riproduca il tipo generale di quella constatata nei destrimani, tuttavia riveste spesso un carattere misto nel senso che comporta deficit funzionali propri ai due emisferi (v. Hecaen e Sauguet, 1971).
La prova di Wada ha anche permesso a Milner e altri (v., 1966) di constatare che il linguaggio era rappresentato bilateralmente in un numero rilevante dei loro soggetti mancini. Queste diverse argomentazioni che parlano in favore di una certa ambilateralità cerebrale nei soggetti mancini normali (v. Zurif e Bryden, 1969; v. Satz e altri, 1967) o nei soggetti mancini con lesioni emisferiche unilaterali (v. Hecaen e Sauguet, 1971) fanno sospettare che questa ambilateralità cerebrale sia propria solo di coloro che hanno ascendenti mancini.
d) Origine della specializzazione emisferica
Resta infine il problema dell'origine della specializzazione emisferica. Come si può porre il problema della ontogenesi della dominanza cerebrale? L'asimmetria è presente sin dalla formazione del cervello umano o viene acquisita nel corso della maturazione?
Per tentare di rispondere a questa domanda abbiamo a disposizione tre ordini di dati: le afasie acquisite nel bambino, i risultati di prove psicometriche nei soggetti che hanno subito delle lesioni neonatali, gli studi anatomici delle asimmetrie emisferiche nel feto e nel neonato.
Da una revisione delle osservazioni compiute sui disturbi acquisiti del linguaggio nei bambini e dallo studio di casi personali, pensiamo si possano desumere i seguenti fatti. In modo particolare grazie ai dati ottenuti da Basser (v., 1962), si sa che le emisferectomie compiute in seguito a lesioni sopravvenute prima dell'acquisizione del linguaggio producono effetti simili qualunque sia il lato sul quale sono state praticate. D'altronde mai è stata constatata una disfasia permanente dopo emisferectomia destra praticata per lesioni sopravvenute dopo l'acquisizione del linguaggio. Possono sopravvenire dei disturbi del linguaggio, dopo lesione dell'emisfero destro in giovane età, soltanto quando il bambino è molto piccolo, ma la loro frequenza in effetti non è più elevata che nell'adulto (v. Krashen, 1973; v. Hecaen, 1976). Le afasie infantili, relativamente frequenti nel caso di lesioni a sinistra, a meno che non siano massive, hanno abitualmente un carattere regressivo. Gli aspetti clinici dei disturbi del linguaggio sono diversi nel bambino e nell'adulto. Nel bambino non vi è mai logorrea, le parafasie verbali sono rare e i disturbi dell'espressione (mutismo o disordini articolatori, disturbi della scrittura) predominano qualunque sia la sede intraemisferica della lesione.
Questi dati sono dunque in favore di una certa equipotenzialità emisferica con possibilità di spostamento all'altro emisfero della rappresentazione del linguaggio durante la maturazione cerebrale. Il periodo critico durante il quale sarebbe possibile il trasferimento durerebbe, secondo Lenneberg (v., 1967), sin quasi alla pubertà. Si tende tuttavia ad abbassarne la data terminale a un'età ben più precoce.
Al contrario, le due altre serie di argomenti sono a favore di una specializzazione emisferica stabilita ben più precocemente, se non innata.
In effetti, gli esami psicologici dei soggetti con lesioni emisferiche unilaterali e perinatali rivelano un tipo di deficit a seconda dell'emisfero leso, simile a quello riscontrato negli adulti; cioè deficit nei compiti verbali nel caso di lesioni a sinistra, deficit nei compiti non verbali, in particolare spaziali, nel caso di lesioni a destra (v. McFie, 1961; v. Fedio e Mirsky, 1969; v. Woods e Teuber, 1973; v. Rudei e altri, 1974).
L'altra serie di argomenti proviene da esami compiuti su cervelli di feti e di neonati. Teszner e altri (v., 1972), Witelson e Pallie (v., 1973) e soprattutto Wada e altri (v., 1975) ritrovano in effetti su questi cervelli la stessa asimmetria destra-sinistra del planum temporale, cioè dell'area posteriore del linguaggio, constatata nei cervelli degli adulti da Oeschwind e Levitsky (v., 1968). Tuttavia Wada e collaboratori osservano che la differenza in favore del planum sinistro è minore nel feto e nel neonato di quanto non lo sia nell'adulto, così che questa differenza sembrerebbe accrescersi con la maturazione.
La discordanza tra questi diversi risultati si può attenuare se si tiene conto di certe recenti constatazioni. Soprattutto, tra i fattori che intervengono nello spostamento della rappresentazione del linguaggio da un emisfero all'altro bisogna considerare, oltre all'età, la sede della lesione. Milner (v., 1974) ha in effetti constatato, con il metodo di Wada, che malgrado ampie e precoci lesioni a sinistra il linguaggio era sempre rappresentato in questo emisfero se la regione parieto-temporale sinistra era integra; al contrario, in caso di lesione anche limitata di questa regione, si produceva lo spostamento all'emisfero destro. Dunque la riorganizzazione può anche aversi sullo stesso emisfero, e forse per la sua comparsa avrebbe importanza una differenza di maturazione delle diverse aree nella zona del linguaggio. Anche lo spostamento della rappresentazione del linguaggio all'emisfero destro può, in certi casi, essere solo parziale (v. Nebes e Sperry, 1971). Infine bisogna almeno accennare al caso particolarissimo di una bambina privata di tutte le comunicazioni verbali dai 20 mesi ai 13 anni, studiato da Fromkin e altri (v., 1974). In questa bambina in cui l'acquisizione del linguaggio resta limitata malgrado il ritorno a una vita normale, l'ascolto dicotico rivela che solo l'emisfero destro tratta il materiale verbale o non verbale. Il problema del periodo critico per il trasferimento e per la maturazione può dunque essere posto in modo diverso: senza lo stimolo adeguato durante la maturazione una zona preformata non potrebbe acquisire la sua capacità funzionale.
L'asimmetria funzionale emisferica è un carattere specifico della specie umana o si possono individuare dei precursori nella scala filogenetica? Sebbene alcuni lavori abbiano rivelato una asimmetria negli Uccelli (v. Notebohm, 1970) e nel topo (v. Collins e Ward, 1970), questi fatti sembrano assai lontani dalla dominanza cerebrale umana. Al contrario, ricerche su animali con sezione delle commissure emisferiche (split brain), sul gatto (v. Robinson e Voneida, 1973; v. Webster, 1972) e soprattutto sulla scimmia (v. Trevarthen 1965; v. Gazzaniga, 1963; v. Butler e Francis, 1973; v. Doty e Yamaga, 1973; v. Hamilton e altri, 1974), suggeriscono la presenza di almeno qualche elemento di una tale specializzazione, non ancora sistematica, talora diversa a seconda degli individui, ma che potrebbe costituire l'abbozzo, nell'evoluzione filogenetica, della dominanza cerebrale umana.
bibliografia
Albert, M., Hecaen, H., Relative movement perception following unilateral brain cerebral damage, in ‟Transactions of the American Neurological Association", 1971, XCVI, pp. 200-202.
Babinski, J., Contribution à l'étude des troubles mentaux dans l'hémiplégie cérébrale (anosognosie), in ‟Revue neurologique", 1914, XXVII, pp. 845-847.
Basser, L. S., Hemiplegia of early onset and the faculty of speech with special reference to the effects of hemispherectomy, in ‟Brain", 1962, LXXXV, pp. 427-460.
Benton, A. L., Hecaen, H., Stereoscopic vision in patients with unilateral cerebral damage, in ‟Neurology", 1970, XX, pp. 1084-1088.
Benton, A. L., Levin, H. S., Varney, N. R., Tactile perception of direction in normal subject: implications for hemispheric cerebral dominance, in ‟Neurology", 1973, XXIII, pp. 1248-1250.
Bever, T. G., Chiarello, R. J., Cerebral dominance in musicians and nonmusicians, in ‟Science", 1974, CLXXXV, pp. 537-539.
Bodamer, J., Die Prosopagnosie, in ‟Archiv für Psychiatrie und Zeitschrift für Neurologie", 1947, CLXXIX, pp. 6-54.
Brain, R., Visual disorientation with special reference to the lesions of the right cerebral hemisphere, in ‟Brain", 1941, LXIV, pp. 244-272.
Broca, P., Perte de la parole, ramollissement chronique et destruction partielle du lobe antérieur gauche du cerveau, in ‟Bulletin de la Société d'Anthropologie", 1865, VI, pp. 337-393.
Butler, C. R., Francis, A. C., Specialization of the left hemisphere in baboon. Evidence from directional preferences, in ‟Neuropsychologia", 1973, XI, pp. 351-354.
Carmon, A., Bechtoldt, H. P., Dominance of the right cerebral hemisphere for stereopsis, in ‟Neuropsychologia", 1969, VII, pp. 29-40.
Chaney, R. B. Jr., Webster, J. C., Information in certain multidimensional sounds, in ‟Journal of the Acoustical Society of America", 1966, XL, pp. 447-455.
Cohen, L., Perception of reversible figures after brain injury, in ‟Archives of neurology", 1959, LXXXI, pp. 765-775.
Collins, R. L., Ward, R., Evidence for an asymmetry of cerebral function in mice tested for audiogenic seizures, in ‟Nature", 1970, CCXXVI, pp. 1062-1063.
Corkin, S., Tactually-guided maze learning in man: effects of unilateral excisions and bilateral hippocampal lesions, in ‟Neuropsychologia", 1965, III, pp. 339-352.
Corsi, P. M., Human memory and the medial temporal region of the brain, Montreal 1972.
Curry, F. K. W., A comparison of left handed and right handed subjects on verbal and non verbal listening tasks,in ‟Cortex", 1967, III, pp. 343-352.
De Renzi, E., Les désorganisations pathologiques de l'espace extra corporel, in De l'espace corporel à l'espace écologique. Symposium de l'Association de Psychologie Scientifique de Langue Française, Paris 1974, pp. 149-186.
De Renzi, E., Faglioni, P., Scotti, G., Hemispheric contribution to exploration of space through the visual and tactile modality, in ‟Cortex", 1970, VI, pp. 191-203.
De Renzi, E., Faglioni, P., Scotti, G., Judgment of spatial orientation in patients with focal brain damage, in ‟Journal of neurology, neurosurgery and psychiatry", 1971, XXXIV, pp. 489-495.
De Renzi, E., Faglioni, P., Spinnler, H., The performance of patients with unilateral brain damage on face recognition tasks, in ‟Cortex", 1968, IV, pp. 17-34.
De Renzi, E., Scotti, G., Spinnler, H., Perceptual and associative disorders of visual recognition: relationship to the side of the cerebral lesion, in ‟Neurology", 1969, XIX, pp. 634-642.
De Renzi, E., Spinnler, H., Visual recognition in patients with unilateral cerebral disease, in ‟Journal of nervous and mental disease", 1966, CXLII, pp. 513-525.
Dide, M., Les désorientations temporospatiales et la prépondérance de l'hémisphère droit dans les agnosies akinésies proprioceptives, in ‟Encéphale", 1938, II, pp. 276-294.
Dimond, S. J., Beaumont, J. G., Experimental studies of hemisphere function in the human brain, in Hemisphere function in the human brain (a cura di S. J. Dimond, J. G. Beaumont e P. Elek), London 1974, pp. 89-120.
Doty, R. W., Yamaga, K., Maze behavior in macaques, in ‟American journal of physical anthropology", 1973, XXXVIII, pp. 403-405.
Dunn, Th. D., Double hemiplegia with double hemianopsia and loss of geographical centre, in ‟Transactions of the College of Physicians of Philadelphia", 1895, XVII, pp. 39-55.
Durnford, M., Kimura, D., Right hemisphere spécialization for depth perception reflected in visual field differences, in ‟Nature", 1971, CCXXXI, pp. 394-395.
Fedio, P., Mirsky, A. F., Selective intellectual deficits in children with temporal lobe or centrencephalic epilepsy, in ‟Neuropsychologia", 1969, VII, pp. 287-300.
Freund, C. S., Über optische Aphasie und Seelenblindheit, in ‟Archiv für Psychiatrie und Nervenkrankheiten", 1888, XX, pp. 276-297.
Fromkin, V. A., Krashen, S., Curtiss, S., Rigler, D., Rigler, M., The development of language in genie: a case of language acquisition beyond the critical period', in ‟Brain and language", 1974, I, pp. 81-108.
Gazzaniga, M. S., Effects of commissurotomy on a preoperatively learned visual discrimination, in ‟Experimental neurology", 1963, VIII, pp. 14-19.
Gazzaniga, M. S., Bogen, J. E., Sperry, R. W., Observations on visual perception after disconnection of the cerebral hemispheres in man, in ‟Brain", 1965, LXXXVIII, pp. 221-236.
Gazzaniga, M., Hillyard, S. A., Language and speech capacity of the right hemisphere, in ‟Neuropsychologia", 1971, IX, pp. 273-280.
Geschwind, N., Disconnexion syndromes in animals and man, in ‟Brain", 1965, LXXXVIII, pp. 237-294, 585-644.
Geschwind, N., Levitsky, W., Human brain, left-right asymetries in temporal speech regions, in ‟Science", 1968, CLXI, pp. 186-187.
Gordon, H. W., Hemispheric asymmetry in the perception of musical chords, in ‟Cortex", 1970, VI, pp. 387-399.
Hamilton, C. R., Tieman, S. B., Farrell, W. S., Cerebral dominance in monkeys?, in ‟Neuropsychologia", 1974, XII, pp. 193-198.
Hecaen, H., Acquired aphasia in chlidren and ontogenesis of hemisphere specialization, in ‟Brain and language", 1976, III, 114-134.
Hecaen, H., Ajuriaguerra, J. de, Massonnet, J., Les troubles visuoconstructifs par lésions pariéto-occipitales droites. Rôle des perturbations vestibulaires, in ‟Encéphale", 1951, I, pp. 122-179.
Hecaen, H., Angelergues, R., Agnosia for faces, in ‟Archives of neurology", 1962, VII, pp. 92-100.
Hecaen, H., Angelergues, R., La cécité psychique, Paris 1963.
Hecaen, H., Angelergues, R., Neuropsychologie clinique des dysfonctionnements des lobes occipitaux, in Rapport du 8ème Congrès International de Neurologie III, Wien 1965, pp. 29-42.
Hecaen, H., Assal, G., A comparisonof constructive deficits following right and left hemispheric lesions, in ‟Neuropsychologia", 1970, VIII, pp. 289-304.
Hecaen, H., Marcie, P., Disorders of written language following right hemisphere lesions: spatial disgraphia, in Hemisphere function in the human brain (a cura di S. J. Dimond, J. G. Beaumont e P. Elek), London 1974, pp. 345-366.
Hecaen, H., Penfield, W., Bertrand, C., Malmo, R., The syndrome of apractognosia due to lesions of the minor cerebral hemisphere, in ‟Archives of neurology and psychiatry" 1956, LXXV, pp. 400-434.
Hecaen, H., Sauguet, J., Cerebral dominance in left-handed subjects, in ‟Cortex", 1971, VII, pp. 19-48.
Hecaen, H., Tzortzis, C., Masure, M. C., Trouble de l'orientation spatiale dans une épreuve de recherche d'itinéraires lors des lésions corticales unilatérales, in ‟Perception", 1972, I, pp. 325-330.
Jackson, H., On the nature of the duality of the brain, in Selected writings II (a cura di J. Taylor), London 1932, pp. 129-145.
Kimura, D., Some effects of temporal-lobe damage on auditory perception, in ‟Canadian journal of psychology", 1961, XV, pp. 156-165.
Kimura, D., Cerebral dominance and the perception of verbal stimuli, in ‟Canadian journal of psychology", 1961, XV, pp. 166-171.
Kimura, D., Right temporal lobe damage, in ‟Archives of neurology", 1963, VIII, pp. 264-271.
Kimura, D., Left-right differences in the perception of melodies, in ‟Quarterly journal of experimental psychology", 1964, XVI, pp. 355-358.
Kimura, D., The asymmetry of the human brain, in ‟Scientific American", 1973, CCXXVIII, 3, pp. 70-78.
Krashen, S., Lateralization, language learning and the critical period. Some new evidence, in ‟Language learning", 1973, XXIII, pp. 63-74.
Lenneberg, E., Biological foundations of language, New York-London-Sydney 1967.
Levy, J., Psychobiological implications of bilateral asymmetry, in Hemisphere function in the human brain (a cura di S. J. Dimond, J. G. Beaumont e P. Elek), London 1974, pp. 121-183.
Levy J., Trevarthen C., Sperry R. W., Perception of bilateral chimeric figures following hemispheric deconnection, in ‟Brain", 1972, XCV, pp. 61-78.
Liepmann, H., Das Krankheitsbild der Apraxie (motorischen Asymbolie), in ‟Monatschrift für Psychiatrie", 1900, VIII, pp. 102-132, 144-145, 182-197.
Lissäuert, H., Ein Fall von Seelenblindheit nebst einem Beitrag zur Theorie derselben, in ‟Archiv für Psychiatrie und Nervenkrankheiten", 1890, XXI, pp. 222-270.
McFie, J., Intellectual impairment in children with localized post infantile cerebral lesions, in ‟Journal of neurology, neurosurgery and psychiatry", 1961, XXIV, pp. 361-365.
McFie, J., Piercy, M. F., Zangwill, O. L., Visual spatial agnosia associated with lesions of the right cerebral hemisphere, in ‟Brain", 1950, LXXIII, pp. 167-190.
Milner, B., Laterality effects in audition, in Interhemispheric relations and cerebral dominance, Baltimore 1962, pp. 177-195.
Milner, B., Visually-guided maze learning in man: effects of bilateral hippocampal, bilateral frontal and unilateral cerebral lesions, in ‟Neuropsychologia", 1965, III, pp. 339-352.
Milner, B., Interhemispheric differences in the localization of psychological processes in man, in ‟British medical bulletin", 1971, XXVII, pp. 272-277.
Milner, B., Hemispheric specialization: scope and limits, in The neurosciences third study program (a cura di F. O. Schmitt e F. G. Worden), Cambridge, Mass., 1974, pp. 75-88.
Milner, B., Branch, C., Rasmussen, Th., Evidence for bilateral speech representation in some non right handers, in ‟Transactions of the American Neurological Association", 1966, XCI, pp. 306-308.
Mullan, S., Penfield, W., Illusions of comparative interpretation and emotion, in ‟Archives of neurology and psychiatry", 1959, LXXXXI, pp. 269-284.
Nebes, R. D., Superiority of the minor hemisphere in commissurotomized man for the perception of part-whole relations, in ‟Cortex", 1971, VII, pp. 333-349.
Nebes, R. D., Sperry, R. W., Hemispheric deconnection syndrome with cerebral birth injury in the dominant arm area, in ‟Neuropsychologia", 1971, IX, pp. 190-216.
Newcombe, F., Missile wounds of the brain, London 1969.
Notebohm, F., Ontogenesis of birdsong, in ‟Science", 1970, CLXVII, pp. 950-956.
Ojemann, G. A., Alteration in non-verbal short term memory with stimulation in the region of the mamillothalamic tract in man, in ‟Neuropsychologia", 1971, IX, pp. 195-202.
Ojemann, G. A., Fedio, P., Van Buren, J. M., Anomia from pulvinar and subcortical parietal stimulation, in ‟Brain", 1968, XCI, pp. 99-116.
Papçun, G., Krashen, S., Terbeek, D., Remington, R., Harshman, R., Is the left hemisphere specialized for speech, language and for something else?, in ‟Journal of the Acoustical Society of America", 1974, LV, pp. 319-327.
Pötzl, O., Die optische-agnostischen Deuticke, Leipzig 1928.
Ratcliff, G., Newcombe, F., Spatial orientation in man: effects of left-right and bilateral posterior lesions, in ‟Journal of neurology, neurosurgery and psychiatry", 1973, XXXVI, pp. 448-454.
Reichardt, M., Die Störungen der optische-räumnlichen Funktionen. Des Erkennens und Wiederkennens, in Allgemeine und spezielle Psychiatrie (a cura di G. Fisher), Iena 1907, pp. 40-49.
Rizzolatti, G., Umiltà, C., Berlucchi, G., Opposite superiorities of the right and left cerbral hemispheres in discriminative reaction time to physiognomical and alphabetical material, in ‟Brain", 1971, XCIV, pp. 431-442.
Robinson, J. S., Voneida, T., Hemisphere differences in cognitive capacity in the split brain cat, in ‟Experimental neurology", 1973, XXXVIII, pp. 123-134.
Rudel, R. G., Denckla, M. B., Relations of forward and backward digit repetition to neurological impairment in children with learning disabilities, in ‟Neuropsychologia", 1974, XII, pp. 109-118.
Rudel, R., Teuber, H. L., Twitchell, T. E., Levels of impairment of sensorimotor early damage, in ‟Neuropsychologia", 1974, XII, pp. 95-108.
Satz, P., Achenback, K., Fennelle, E., Correlations between assessed manual laterality and predicted speech latearlity in a normal population, in ‟Neuropsychologia", 1967, V, pp. 295-310.
Schilder, P., The image and appearance of the human body, London 1935.
Semmes, J., A non tactual factor in astereognosis, in ‟Neuropsychologia", 1965, III, pp. 295-315.
Semmes, J., Weinstein, S., Ghent, L., Teuber, H. L., Somatosensory changes after penetrating brain wounds in man, Cambridge, Mass., 1960.
Shankweiler, D., Effect of temporal-lobe damage on perception of dichotically presented melodies, in ‟Journal of comparative and physiological psychology", 1966, LXII, pp. 115-119.
Shankweiler, D., Studdert-Kennedy, M., Identification of consonants and vowels presented to the left and right ears, in ‟Quarterly journal of experimental psychology", 1967, XIX, pp. 59-63.
Smith, A., Speech and other functions after left dominant hemispherectomy, in ‟Journal of neurology, neurosurgery and psychiatry", 1966, XXIX, pp. 467-471.
Sperry, R. W., Gazzaniga, M. S., Language following surgical disconnection of the hemisphere, in Brain mechanisms underlying speech and language (a cura di C. H. Millikan e F. Darley), New York 1967, pp. 108-118.
Teszner, D., Tzavaras, A., Gruner, J., Hecaen, H., L'asymétrie droite-gauche du planum temporale. A propos de l'étude anatomique de 100 cerveaux, in ‟Revue neurologique", 1972, CXXVI, pp. 444-449.
Teuber, H. L., Battersby, W. S., Bender, M. B., Visual field defects after penetrating missile wounds of the brain, Cambridge, Mass., 1960.
Trevarthen, C. B., Functional interaction between the cerebral hemispheres of the split-brain monkey, in Functions of the corpus callosum (a cura di G. Ettlinger), London 1965, pp. 24-41.
Trevarthen, C. B., Functional relations of disconnected hemispheres with the brain stem and with each other in monkey and man, in Hemispheric disconnection and cerebral function (a cura di M. Kinsbourne e W. L. Smith), Springfield, Ill., 1974, pp. 187-207.
Tzavaras, A., Hecaen, H., Étude des coordonnées visuelles subjectives au cours des lésions corticales unilatérales, in ‟Revue neurologique", 1972, CXXV, pp. 458-461.
Tzavaras, A., Hecaen, H., Le Bras, H., Le problème de la spécificité du déficit de la reconnaissance du visage humain lors des lésions hémisphériques unilatérales, in ‟Neuropsychologia", 1970, VIII, pp. 403-417.
Vignolo, L. A., Auditory agnosia. A review and report of recent evidence, in Contributions to clinical neuropsychology (a cura di A. L. Benton), Chicago 1969, pp. 172-208.
Wada, J. A., Clarke, R., Hamm, A., Cerebral hemispheric asymmetry in humans. Cortical speech zones in 100 adult and 100 infant brains, in ‟Archives of neurology", 1975, XXXII, pp. 239-246.
Warrington, E. K., James, M., Kinsbourne, M., Drawing disability in relation to laterality of cerebral lesions, in ‟Brain", 1966, LXXXIX, pp. 53-82.
Webster, W. G., Functional asymmetry between the cerebral hemispheres of the cat, in ‟Neuropsychologia", 1972, X, pp. 75-87.
Wernicke, C., Der aphasische Symptomencomplex, Breslau 1874.
Witelson, S. F., Pallie, W., Left hemisphere specialization for language in the newborn: neuroanatomical evidence of asymmetry, in ‟Brain", 1973, XCVI, pp. 641-646.
Woods, B. T., Teuber, H. L., Early onset of complementary specialization of cerebral hemispheres in man, in ‟Transactions of the American Neurological Association", 1973, XCVIII, pp. 113-115.
Yin, R. K., Face recognition by brain-injured patients: a dissociate ability, in ‟Neuropsychologia", 1970, VIII, pp. 395-402.
Zurif, E. B., Bryden, M. P., Familial handedness and left-right differences in auditory and visual perception, in ‟Neuropsychologia", 1969, VII, pp. 179-188.
Interazioni interemisferiche cerebrali
di Giovanni Berlucchi
sommario: 1. Introduzione. 2. Anatomia delle connessioni nervose fra gli emisferi. a) Organizzazione generale. b) Connessioni interemisferiche neocorticali. 3. Fisiologia delle commissure telencefaliche. a) Loro ruolo nell'organizzazione interemisferica delle attività elettriche normali e anormali del cervello. b) Interazioni interemisferiche specifiche rivelate aa esperimenti elettrofisiologici. 4. Esperimenti neurocomportamentali sulle comunicazioni interemisferiche. a) Esperimenti sugli animali con cervello bisecato. b) Esperimenti su soggetti umani con cervello bisecato. 5. Conclusione. □ Bibliografia.
1. Introduzione
Il cervello dei Vertebrati è costituito da due organi appaiati, gli emisferi cerebrali, che sono rimarchevolmente simili l'uno all'altro in tutti i dettagli del loro piano strutturale. Fin da quando l'attività degli emisferi cerebrali è stata messa in relazione con la vita mentale, la chiara antinomia fra la dualità morfologica del cervello e l'apparente unità della coscienza e della condotta individuale è stata una costante sorgente di perplessità sia per i filosofi sia per gli scienziati. Dai tempi antichi ad oggi, non solo le idee filosofiche sulle relazioni fra la mente e il cervello, ma anche le teorie fisiologiche sull'organizzazione e sull'attività generale del sistema nervoso centrale sono state spesso ispirate dalla considerazione di questa antinomia e formulate in modo da eliminarla. Per esempio, le innumerevoli illazioni sulla localizzazione della mente o del livello supremo dell'organizzazione nervosa in una varietà di organi o centri intracerebrali singoli e mediani - dalla ghiandola pineale cartesiana al cuore reticolare troncoencefalico delle moderne speculazioni neurofisiologiche - possono essere tutte considerate come dei tentativi più o meno espliciti di attribuire un substrato strutturale unitario a quella che generalmente si presume essere la massima unità funzionale. Il problema viene affrontato in maniera meno speculativa quando si cerca di determinare il ruolo dei due emisferi cerebrali, presi singolarmente o insieme, nel controllo del comportamento e, più specificamente, nella realizzazione di quella unitarietà d'azione che, come disse C. S. Sherrington (v., 1906), ‟è una pietra fondamentale nella costruzione dell'individuo, l'attuazione della cui unità è la funzione specifica del sistema nervoso". In questi termini, il problema rientra chiaramente nella neurofisiologia integrativa, e la sua soluzione dipende dalla misura in cui riusciamo a comprendere: a) le funzioni specifiche servite da ciascun emisfero; b) i meccanismi di coordinazione grazie ai quali le attività emisferiche sono organizzate e integrate nel prodotto finale del lavoro di tutto il sistema nervoso centrale. Per il primo punto, bisogna fare una distinzione fra l'uomo e tutti gli altri Vertebrati. Per quanto ne sappiamo, sia nelle specie non mammifere che nei Mammiferi subumani i due emisferi sono funzionalmente equiparabili da tutti i punti di vista, con la specificazione che ogni emisfero è prevalentemente interessato alle funzioni di senso e di moto del lato controlaterale. Nella specie umana gli emisferi cerebrali non si differenziano solo in ragione delle loro relazioni con il lato destro e sinistro del corpo, ma acquistano anche capacità funzionali proprie, che possono essere alterate selettivamente da lesioni cerebrali unilaterali (v. emisferi cerebrali: Dominanza cerebrale). Il secondo punto è il tema specifico del presente articolo; la sua discussione verrà sviluppata secondo gli aspetti anatomici, fisiologlci e psicologici.
2. Anatomia delle connessioni nervose fra gli emisferi
a) Organizzazione generale
Ogni considerazione sulle interazioni fra gli emisferi deve essere preceduta da una descrizione delle vie anatomiche attraverso le quali le attività emisferiche possono essere coordinate e le influenze interemisferiche possono essere esercitate. Le connessioni più dirette, le commissure, sono compatti sistemi di fibre che attraversano la linea mediana a vari livelli del nevrasse e in genere uniscono aree corrispondenti dei due lati del cervello. Esse sono: le commissure telencefaliche, che includono il corpo calloso, la commissura anteriore e la commissura ippocampale; le commissure diencefaliche e mesencefaliche, che includono la commissura intertalamica, il sistema delle commissure sopraottiche, la commissura abenulare, la commissura posteriore e la commissura intertettale. Altre connessioni, meno dirette, attraverso le quali possono parimenti avvenire comunicazioni interemisferiche, sono i circuiti multisinaptici diffusi della formazione reticolare del talamo e del tronco dell'encefalo e, forse, i sistemi commissurali del midollo spinale. In aggiunta a queste varie vie nervose di interconnessione, gli emisferi cerebrali ovviamente hanno in comune i fluidi circolanti e interstiziali, e non si può escludere che qualche scambio interemisferico di informazioni possa anche prendere questa via non nervosa. Tuttavia, si dimostrerà più avanti che le commissure telencefaliche neocorticali sono il substrato di gran lunga più importante delle interazioni fra gli emisferi, quindi la presente descrizione anatomica sarà in gran parte ristretta a queste commissure.
Il corpo calloso è la commissura della neocorteccia, con la quale esso evolve in parallelo. I Vertebrati non Mammiferi o i bassi Mammiferi come i Monotremi e alcuni dei Marsupiali sono caratterizzati da uno sviluppo scarso o nullo della neocorteccia; in accordo a ciò, il loro cervello non ha corpo calloso. Nei Mammiferi superiori, e specialmente nei Primati, il corpo calloso è un'enorme struttura le cui dimensioni superano largamente quelle di tutti gli altri tratti fibrosi nel cervello. Il corpo calloso dell'uomo contiene circa 200 milioni di fibre, la maggioranza delle quali sono di piccolo diametro (circa 2 μ (v. Tomasch, 1954). Il numero delle fibre callosali nel gatto, nel ratto, nel coniglio e nel cane è rispettivamente 2, 5, 6, e 22 milioni, secondo quanto riportano S. M. Blinkov e I. I. Glezer (v., 1968). La commissura anteriore è fondamentalmente la commissura della paleocorteccia e dei bulbi olfattivi, ma contiene anche fibre di origine neocorticale; di fatto, le scarse connessioni interemisferiche neocorticali nelle specie acallosali dei Mammiferi si effettuano tutte tramite questa commissura. Nell'uomo vi sono circa da 2,5 a 4 milioni di fibre nella commissura anteriore (v. Tomasch, 1957). La commissura ippocampale o psalterium connette reciprocamente le formazioni ippocampali dei due lati; essa può essere considerata la commissura dell'archicorteccia.
b) Connessioni interemisferiche neocorticali
Prima del 1950, le caratteristiche topologiche generali delle connessioni interemisferiche neocorticali attraverso il corpo calloso e la commissura anteriore sono state studiate: a) anatomicamente, con l'esame microscopico di cervelli normali impregnati con i metodi di Golgi o Cajal (v., ad es., Cajal, 1911; v. i contributi di de Villaverde) o di cervelli affetti da lesioni corticali o commissurali, colorati con metodi per la dimostrazione di fibre mieliniche degenerate (v., ad es., Van Valkenburg, 1913; v. Polyak, 1927; v. i contributi di Mettler); b) elettrofisiologicamente, registrando e mappando in tutta la corteccia di un emisfero le risposte elettrobiologiche causate dalla stimolazione elettrica (v. Curtis, Intercortical connections..., 1940; v. Bremer e altri, 1956) o dall'irritazione chimica locale (‛neuronografia stricnica'; v. Bailey e altri, 1941; v. McCulloch e Garol, 1941; v. Garol, 1942) della corteccia dell'altro emisfero, prima e dopo sezione del corpo calloso, della commissura anteriore o di entrambi. Durante l'ultimo ventennio, le moderne tecniche all'argento per impregnare assoni in degenerazione (metodo di Nauta e sue varie modificazioni) sono state applicate intensivamente, in combinazione con lesioni appropriate della corteccia o delle commissure, allo studio delle connessioni interemisferiche neocorticali; dato che i risultati degli studi precedenti sono stati in parte confermati, in parte corretti o precisati e in parte del tutto soppiantati da quelli ottenuti con i nuovi metodi istologici, solo questi ultimi verranno riassunti brevemente nel presente paragrafo.
Prima di entrare nei dettagli dell'organizzazione delle connessioni interemisferiche corticali, è bene considerarne il piano generale. La neuroanatomia aveva dimostrato, e l'elettrofisiologia classica aveva confermato, che la distribuzione delle fibre commissurali attraverso la corteccia è molto ineguale, visto che alcune aree corticali sono riccamente provviste di tali fibre mentre altre mancano di qualsiasi connessione commissurale. Svariati principi di base sono stati proposti per schematizzare le proprietà topologiche delle proiezioni commissurali. Il principio dell'omotopia postula che le fibre commissurali interconnettano aree corticali simmetriche dei due emisferi: esso è saldamente sostenuto da ripetute dimostrazioni che indicano che i legami interemisferici più forti si stabiliscono effettivamente fra punti corticali simmetrici; tuttavia, la sua enunciazione più restrittiva è certamente falsa, visto che esistono connessioni eterotopiche in numero certamente non trascurabile. Vi è poi il principio dell'eterolateralità di Mettler, che sostiene che una data area corticale ha connessioni commissurali non solo con l'area simmetrica corrispettiva, ma anche con le aree controlaterali che sono simmetriche rispetto a tutte le aree dell'emisfero ipsilaterale con cui essa è connessa (v. Mettler, 1935); anche questo principio, però, è almeno in parte contraddetto dalla nota acquisizione che le connessioni interemisferiche sono di regola molto meno diffuse di quelle intraemisferiche. Lo stato attuale delle conoscenze sta in favore di un principio che si può definire di omofunzionalità: un'area della corteccia che rappresenta una parte della periferia motoria o sensitivo-sensoriale proietta fibre commissurali sia all'area simmetrica sia ad altre aree corticali dell'emisfero opposto che rappresentano la stessa parte periferica. Tuttavia, è evidente che parecchie aree corticali non hanno connessioni commissurali. L'applicazione della teoria generale della corteccia di P. Flechsig (v., 1896) all'organizzazione delle connessioni interemisferiche (v. Geschwind, 1965) ha portato all'idea che l'assenza delle connessioni commissurali sia la caratteristica delle aree primarie di senso, e la presenza di molte di queste connessioni sia la caratteristica delle aree di associazione, definendo le due classi di aree corticali sulla base di criteri citoarchitettonici e odologici. Sappiamo oggi che le cose non stanno così e che le fibre commissurali originano e terminano in aree corticali circoscritte qualificate dalla loro relazione con certe parti del corpo o dello spazio visivo o con certe frequenze sonore, anziché dalle loro caratteristiche citoarchitettoniche.
Nelle specie dei Mammiferi che hanno un corpo calloso, la maggior parte delle connessioni commissurali della neo-corteccia sono di natura callosale; è solo la corteccia della parte inferiore del lobo temporale che ha connessioni interemisferiche attraverso la commissura anteriore. L'esistenza di fibre della corteccia occipitale nella commissura anteriore della scimmia è stata discussa, ma non provata al di là di ogni dubbio (v. Doty e Negrào, 1973). Nella seguente descrizione, l'attributo ‛commissurale' verrà usato intercambiabilmente con ‛calloso', salvo diversa indicazione.
Nella scimmia, le regioni corticali che rappresentano la mano e il piede sia nell'area motoria precentrale (M I) che nell'area somatosensitiva primaria nel giro postcentrale (S I) non mandano nè ricevono proiezioni commissurali. Al contrario, le porzioni di queste aree corticali che rappresentano la faccia, il tronco, le membra e il bacino sono densamente connesse attraverso la linea mediana in maniera omotopica. Queste stesse porzioni di M I e di S I mandano fibre commissurali alle porzioni funzionalmente corrispondenti rispettivamente dell'area motoria supplementare sulla faccia mediale dell'emisfero (M II) e dell'area somatosensitiva secondaria nell'opercolo parietale (S II) (v. Jones e Powell, 1969; v. Pandya e Vignolo, 1969; v. Pandya, Gold e Berger, 1969; v. Karol e Pandya, 1971). Secondo Jones e Powell (v., 1969), le connessioni callosali fra le aree S I dei due lati sono strettamente simmetriche (omotopiche) e rispettano le proprietà delle suddivisioni di S I che riguardano sia la topografia sia la modalità; cioè, esse interconnettono aree corticali che rappresentano esattamente la stessa parte del corpo e contengono cellule che rispondono alla stessa classe di afferenze somatosensitive (superficiali della pelle o profonde dei muscoli e delle articolazioni). Pandya e collaboratori sostengono invece il concetto secondo il quale sia M I sia S I proiettano alle aree corrispettive laterali secondo un modello di sovrapposizione, che prevede che ciascuna suddivisione corticale da un lato (per es. l'area della faccia) sia connessa non solo con l'area simmetrica controlaterale, ma anche con aree contigue che rappresentano parti adiacenti della periferia (per esempio l'occipite e il collo). V'è accordo generale sul fatto che le rispettive proiezioni da S I e M I in un emisfero a S II e M II nell'altro emisfero hanno una disposizione omotopica abbastanza poco rigida, che permette forse una mescolanza di proprietà di luogo e di modalità. Una simile disposizione è tipica delle interconnessioni fra le areeS II (v. Jones e Powell, 1969; v. Pandya e Vignolo, 1969) e le aree M II dei due lati (v. Pandya, Gold e Berger, 1969; v. Karol e Pandya, 1971). Anche qui, le aree della mano e del piede sono prive di connessioni callosali.
Risultati molto simili sono stati descritti per le aree motoria e somatosensitiva della corteccia del gatto (v. Ebner e Myers, 1965; v. Jones e Powell, 1968; v. Kawamura e Otani, 1970). Inoltre, F. F. Ebner e R. E. Myers (v., 1965) hanno osservato che l'estesa area della mano nella corteccia somatosensitiva del procione rimane libera da ogni degenerazione dopo sezione completa delle commissure telencefaliche, mentre le aree della faccia, del braccio e della gamba presentano vari gradi di degenerazione.
Le connessioni callosali della corteccia visiva sono state studiate a fondo nel gatto. In generale, è evidente che queste connessioni sono limitate alle aree corticali che rappresentano il meridiano verticale centrale del campo visivo. L'area che rappresenta il meridiano verticale centrale, situata al confine fra le aree 17 e 18, proietta alla regione simmetrica controlaterale e ad aree che rappresentano la stessa parte del campo visivo nell'area 19 e nell'area soprasilviana laterale. Le rappresentazioni del meridiano verticale nelle due aree 19 sono connesse reciprocamente e inoltre mandano fibre commissurali a porzioni correlate funzionalmente dell'area soprasilviana laterale (v. Hubel e Wiesel, 1965; v. Garey e altri, 1968; v. Wilson, 1968; v. Makarov, 1968-1969; v. Heath e Jones, 1970; v. Berlucchi, 1972). Un'eccezione alla regola che limita le connessioni callosali della corteccia visiva alle regioni che rappresentano il meridiano verticale centrale sembra caratterizzare le connessioni reciproche fra le aree soprasilviane laterali dei due lati, che sono estese all'intera rappresentazione del campo visivo (v. Heath e Jones, 1970). Comunque, la stretta relazione fra le connessioni callosali della corteccia visiva e le rappresentazioni corticali del meridiano verticale centrale del campo visivo è stata messa chiaramente in evidenza nelle seguenti specie: scimmia (v. Myers, Commissurai connections..., 1962; v. Cragg e Ainsworth, 1969; v. Zeki, 1970; v. Karol e Pandya, 1971); coniglio (v. Hughes e Wilson, 1969); ratto (v. Nauta e Bucher, 1954; v. Heimer e altri, 1967); opossum (v. Benevento ed Ebner, 1971). Nell'ultimo animale non v'è corpo calloso e le connessioni interemisferiche della corteccia visiva corrono nella commissura anteriore assieme a tutte le altre connessioni interemisferiche neocorticali.
Uno studio dettagliato delle connessioni commissurali della corteccia uditiva del gatto, eseguito da I. T. Diamond, E. O. Jones e T. P. S. Powell (v., 1968), ha dimostrato che le varie aree di rappresentazione (tonotopica) della coclea in cui questa parte della corteccia è suddivisa sono provviste di fibre commissurali in maniera molto inomogenea. La prima area uditiva (A I) manda fibre alla A I controlaterale e alla seconda area uditiva (A II); queste connessioni seguono il principio di omofunzionalità (vedi sopra), cioè uniscono rappresentazioni della stessa parte della coclea. Connessioni omotopiche si stabiliscono fra le aree A II dei due lati; le aree nel giro ectosilviano posteriore (ep) e nel solco soprasilviano (frangia soprasilviana) sono parimenti connesse in maniera omotopica. Non esistono connessioni interemisferiche per l'area uditiva insulare. È evidente che le fibre commissurali in tutte le suddivisioni uditive della corteccia con connessioni interemisferiche sono limitate a certe regioni e ne evitano altre; questa disposizione è chiara al massimo nell'area A I, dove la rappresentazione delle parti della coclea che rispondono a frequenze tonali fra 2 e 12 KHz ha scarse proiezioni interemisferiche o non ne ha affatto. Anche nella scimmia, terminazioni callosali si trovano in certe parti della corteccia uditiva e non in altre. Le connessioni callosali nel lobo temporale sono limitate alle parti mediali delle aree A I e A II; l'area uditiva di associazione nel piano sopratemporale e nel giro temporale superiore ha parimenti connessioni commissurali non omogenee omotopiche ed eterotopiche, per il tramite in parte del corpo calloso, in parte della commissura anteriore (v. Pandya, Hallet e Mukherjee, 1969; v. Karol e Pandya, 1971). Anche l'area di proiezione del sistema vestibolare nel giro postcentrale della scimmia è solo in parte provvista di proiezioni commissurali (v. Karol e Pandya, 1971). L'area inferotemporale nota per le sue funzioni visive è connessa con l'area corrispondente dell'altro lato sia dal corpo calloso che dalla commissura anteriore, ma anche qui la densità delle proiezioni commissurali è ineguale nelle varie parti dell'area (v. Pandya, Hallet e Mukherjee, 1969; v. Karol e Pandya, 1971). Le connessioni interemisferiche delle varie aree corticali sono distribuite in maniera ordinata entro il corpo calloso.
Riassumendo, lo schema delle connessioni callosali della neocorteccia si interpreta meglio in termini di un legame selettivo fra proiezioni interemisferiche e aree corticali correlate funzionalmente ad alcune parti specifiche delle periferie motorie e sensitivo-sensoriali, anziché sulla base di criteri puramente morfologici. Nelle aree corticali visive, così come nelle aree corticali somatosensitive e motorie, le connessioni callosali sono limitate a zone della corteccia che rappresentano, rispettivamente, le porzioni del campo visivo limitrofe al meridiano verticale centrale e le parti assiali del corpo. Come si discuterà più avanti, il significato funzionale di questo schema di interconnessione è stato almeno in parte chiarito da esperimenti di fisiologia. Le aree corticali uditive e vestibolari, così come le aree di associazione, sono ugualmente divise in zone ‛commissurali' e ‛commissurali', ma il significato fisiologico di questa interconnessione selettiva è al momento oscuro. Discussioni teoriche su questo punto si possono trovare nei lavori di Diamond e altri (v., 1968), Pandya e Vignolo (v., 1969) e Karol e Pandya (v., 1971).
Si deve fare anche menzione delle fibre non commissurali (decussanti) del corpo calloso. Vi sono chiare prove in favore dell'esistenza in diverse specie, incluso l'uomo, di una proiezione cortico-striatale che si incrocia nel corpo calloso (v. Locke e altri, 1964; v. Carman e altri, 1965; v. Locke e Yakovlev, 1965; v. Webster, 1965). Nell'uomo la corteccia limbica (cingolo) è un'importante sorgente di fibre callosali per lo striato controlaterale. Nel gatto la corteccia sensoriale-motoria primaria proietta al nucleo caudato controlaterale attraverso il corpo calloso (v. Carman e altri, 1965).
Una considerazione finale riguarda il luogo preciso di terminazione delle fibre commissurali nei vari strati corticali, e le relazioni sinaptiche dei terminali commissurali con le strutture cellulari e subcellulari della corteccia. Marcate differenze fra le specie nella distribuzione laminare dei terminali callosali sono state descritte da S. Jacobson e E. M. Marcus (v., 1970). Terminali callosali possono essere dimostrati in tutti gli strati corticali delle specie studiate, ma nel ratto questi terminali tendono ad essere più concentrati nei tre strati superiori, mentre nel gatto e specialmente nei Primati essi sono più numerosi negli strati III e IV. Lo strato IV è quello dove terminano le fibre talamocorticali; esso presenta uno sviluppo progressivo nelle specie superiori. Karol e Pandya (v., 1971) suggeriscono che questo progressivo sviluppo può essere accompagnato da una convergenza di afferenze corticali (incluse le callosali) verso questo strato. Si dovrebbe tuttavia considerare che la distribuzione laminare dei terminali callosali entro la corteccia varia da un'area corticale all'altra anche nell'ambito della stessa specie (v., per es., Jacobson, 1970). Là dove una convergenza di afferenze callosali e non callosali (talamocorticali, corticocorticali) alla corteccia è stata dimostrata, sorge il problema della distribuzione di queste diverse afferenze ai vari elementi neuronici corticali. A. Globus e A. B. Scheibel (v., 1967) hanno dimostrato nel coniglio che i terminali callosali sono in larga parte in relazione sinaptica con le spine sui rami obliqui dell'albero dendritico delle cellule piramidali, in contrasto con le afferenze talamocorticali che terminano direttamente sulle spine dell'albero dendritico stesso. J. S. Lund e R. D. Lund (v., 1970) hanno riferito che nel ratto i terminali callosali stanno non solo sulle spine e sui rami dendritici delle cellule piramidali e non piramidali, ma anche sui corpi delle cellule non piramidali e sulla parte più grossa del loro tronco dendritico. Sembra che non vi siano contatti assosomatici fra fibre callosali e cellule piramidali. Altri dati sperimentali e altre illazioni sulla convergenza delle afferenze commissurali e non commissurali nel reticolo cellulare corticale si possono trovare nei lavori di E. G. Jones e T. P. S. Powell (v., 1970), L. A. Benevento e F. F. Ebner (v., 1971) e Karol e Pandya (v., 1971). Per quanto riguarda i corpi cellulari che danno origine a fibre callosali, la vecchia indicazione di L. J. Pines e R. M. Maiman (v., 1939), secondo la quale la sezione del corpo calloso causa degenerazione retrograda negli strati III, IV e VI della corteccia, non è stata confermata da Jacobson (v., 1965). Studi elettrofisiologici sulla corteccia visiva del gatto suggeriscono che i neuroni che proiettano al corpo calloso sono elettivamente situati nel III strato (v. sotto, cap. 3, § b).
3. Fisiologia delle commissure telencefaliche
a) Loro ruolo nell'organizzazione interemisferica delle attività elettriche normali e anormali del cervello
Gli studi classici sull'elettroencefalogramma (EEG) nell'uomo e negli animali avevano indicato che le onde elettriche registrate simultaneamente da punti corrispondenti dei due emisferi del cervello normale sono essenzialmente simili in forma, ampiezza e durata (v. Adrian e Yamagiwa, 1935). Le analisi odierne dell'EEG per mezzo di calcolatori, basate su tecniche di correlazione crociata, hanno sostanzialmente confermato la nozione della sincronia bilaterale dell'EEG, anche se si è visto che la corrispondenza fra i due lati non è così precisa come ritenevano i ricercatori precedenti (v. Andersen e Andersson, 1968). Bremer (v., 1958 e Le corps calleux..., 1966) fu uno dei primi a suggerire che le commissure neocorticali sono essenziali per assicurare questa sincronia bilaterale, e la sua idea è stata corroborata da numerosi esperimenti che dimostrano una diminuzione o scomparsa della sincronia bilaterale dei ritmi EEG nel gatto a seguito di commissurotomie acute e croniche (v. Claes, 1939; v. Bremer e Stoupel, 1957; v. Magni e altri, 1960; v. Berlucchi, 1966). D'altro canto C. Batini, M. Radulovacki, R. T. Kado e W. R. Adey (v. Batini e altri, 1967), nella scimmia, e W. Singer e O. Creutzfeldt (v., 1969), nel gatto, hanno riferito che la sincronia bilaterale dell'EEG, misurata su campioni selezionati e analizzati con correlazione crociata per mezzo di calcolatori, non si modificava significativamente in seguito alla sezione delle commissure telencefaliche. Da ciò essi hanno concluso che la sincronia bilaterale dell'EEG dipende interamente da meccanismi sottocorticali. La loro conclusione, tuttavia, perde molto della sua forza quando si consideri che la sincronia bilaterale nei loro controlli normali con commissure intatte era già così bassa che era difficile aspettarsi un aumento dell'asincronia dopo la commissurotomia. La ragione di questi dati controversi non è chiara, ma è possibile che la sincronia bilaterale dell'EEG possa essere stata oscurata negli animali normali da condizioni ambientali stimolanti implicite negli esperimenti in cui si registra da animali liberi di muoversi. B. A. Meyerson (v., 1968) ha riferito che l'EEG di animali allo stato fetale o neonatale presenta una marcata sincronia bilaterale, per la quale non è necessario il corpo calloso; nell'animale adulto la sincronia bilaterale è molto meno evidente, forse perché essa è facilmente mascherata o disturbata dall'impatto asimmetrico di segnali afferenti alla corteccia. A questo stadio, il corpo calloso e la commissura anteriore possono essere importanti per controbilanciare almeno in parte le influenze che disturbano la sincronia bilaterale. Lo studio dell'EEG di soggetti umani che non hanno il corpo calloso (per abnorme sviluppo cerebrale o per sezione chirurgica) dovrebbe almeno in parte contribuire alla soluzione del problema del ruolo delle commissure corticali nella sincronia bilaterale dell'EEG (v. Bremer e altri, 1956; v. Unterharnscheidt e altri, 1968). Purtroppo non si possono trarre conclusioni definitive da questi studi, dato che la quasi certa esistenza di anormalità cerebrali oltre all'assenza del corpo calloso rende difficile l'accertamento della causa di possibili asimmetrie elettroencefalografiche Analogamente l'osservazione che potenziali elettroencefalografici abnormi possono essere bilateralmente sincroni in alcuni di questi pazienti (che spesso sono anche affetti da epilessia) non indica necessariamente che il corpo calloso non serva alla sincronizzazione bilaterale di potenziali elettroencefalografici normali o abnormi. Si ritornerà su questo punto più avanti.
Al momento attuale, sembra ragionevole ritenere che sia le commissure telencefaliche sia meccanismi sottocorticali, come le commissure più basse o un pacemaker troncoencefalico, siano tutti implicati nella sincronizzazione bilaterale dell'EEG. Questa conclusione è suggerita dai seguenti dati sperimentali: 1) attività elettriche diffuse evocate nella corteccia dalla stimolazione talamica unilaterale si diffondono all'emisfero controlaterale attraverso sia il corpo calloso che la commissura intertalamica (v. Enomoto, 1959; v. Giaquinto, 1969), ed è stata ripetutamente confermata l'analogia fra questi potenziali e le attività elettroencefalografiche spontanee; 2) le attività elettriche di aree corticali dei due emisferi isolate dalle afferenze sottocorticali, ma ancora interconnesse attraverso il corpo calloso, presentano chiari segni di mutua interdipendenza (v. Swank, 1949; v. Andersen e altri, 1967; v. Hossman, Untersuchungen..., 1969); 3) in anestesia profonda (v. Aird e Garoutte, 1958) o durante la vita fetale o neonatale (v. Meyerson, 1968) l'EEG mostra un alto grado di sincronia bilaterale che non dipende dal corpo calloso. Altre prove che stanno in favore di questa conclusione sono fornite dai lavori sui meccanismi commissurali della propagazione interemisferica di attività elettriche abnormi.
Un altro problema da considerare, e da non confondere con la precedente questione sui meccanismi della fine corrispondenza bilaterale delle onde dell'EEG, è quello del possibile ruolo delle connessioni interemisferiche nella simultanea insorgenza di ritmi elettroencefalografici del sonno e della veglia nei due emisferi. I risultati nel gatto (v. Berlucchi, 1966; v. Majkowski, 1967; v. Giaquinto, 1969) e nella scimmia (v. Batini e altri, 1967) indicano chiaramente che si possono sezionare le commissure telencefaliche, diencefaliche e mesencefaliche e addirittura parte del tegmento mesencefalico, senza interferire con la normale simultaneità di comparsa nei due lati del cervello dei segni elettroencefalografici del risveglio e dei vari stadi di sonno. È solo quando si seziona mediosagittalmente anche il tegmento pontino e bulbare che cominciano ad apparire asimmetrie emisferiche dei ritmi elettroencefalografici del sonno e della veglia (v. Michei e Roffwarg, 1967; v. Mancia e altri, 1968). Tutti questi dati sono in buon accordo con la teoria che il ciclo sonno-veglia è controllato da un pacemaker situato nel cuore reticolare del tronco dell'encefalo (v. SONNO). Nonostante non siano necessarie per l'insorgenza simultanea dei ritmi del sonno e della veglia nei due emisferi, le commissure telencefaliche potrebbero tuttavia contribuire a modulare l'eccitabilità e la reattività della corteccia alle afferenze di senso. Bremer (v., 1953, 1958, e Étude electrophysiologique..., 1966) ha dimostrato che salve transcommissurali possono aumentare l'ampiezza dei potenziali corticali evocati dalla stimolazione di vie ascendenti corticipete, ed ha avanzato l'idea di un'influenza tonica facilitante da parte dell'attività callosale sull'organizzazione generale della corteccia. Un'attività tonica di fibre callosali, variabile con il livello di veglia, è stata dimostrata nel gatto con la registrazione diretta (v. Berlucchi, 1965). Qualche autore (v., ad es., Toglia, 1961) è arrivato al punto di suggerire che il corpo calloso è un sistema attivante trasversale della corteccia, sovraimposto sull'analogo sistema attivante longitudinale che ascende dal tronco dell'encefalo. K. A. Hossman, (v., Midline section..., 1969) ha visto che il risveglio eléttroencefalografico indotto dalla stimolazione troncoencefalica unilaterale nel suo preparato ‛corpo calloso' nel gatto rimaneva limitato alla corteccia dell'emisfero del lato stimolato. Dato che in questo preparato tutte le connessioni nervose fra i due lati del cervello, eccetto il corpo calloso e la sottostante commissura ippocampale, erano state eliminate chirurgicamente, bisogna concludere che il corpo calloso da solo non poteva trasmettere effetti di risveglio dalla corteccia di un emisfero a quella dell'altro. Altri autori (v., ad es., Chang, Interaction..., 1953; v. Landau e altri, 1961; v. Ajmone Marsan e Morillo, 1963) hanno anche descritto influenze commissurali riducenti l'ampiezza dei potenziali della corteccia evocati sensorialmente indicando così che anche l'inibizione e l'occlusione possono essere implicate. Come verrà discusso nel È b, la microelettrofisiologia moderna ha dimostrato che l'interazione fra le afferenze di senso e le afferenze commissurali alla corteccia non è semplicemente una questione di potenziamento mutuo o di interferenza, ma una complessa addizione d'informazioni che dà luogo a campi recettivi sensoriali più vasti. È quindi probabile che le attività toniche e fasiche delle commissure telencefaliche siano implicate in regolazioni sia facilitatorie sia inibitorie dei neuroni corticali molto più complesse e raffinate di quelle postulate con il concetto semplicistico di un sistema ‛energizzante' o, il che è lo stesso, ‛frenante'.
A partire dalle ricerche precorritrici di M. Gozzano (v., 1935) e G. Moruzzi (v., 1939), il fenomeno della diffusione interemisferica di un'attività elettrica abnorme della corteccia, inizialmente unilaterale, ha costituito un modello importantissimo per lo studio della propagazione sinaptica dell'accesso epilettico nel tessuto nervoso (v. epilessia). Per converso, le caratteristiche topologiche della diffusione interemisferica di segni corticali di epilessia sono state utilizzate con profitto, nell'era della ‛neuronografia stricnica', al fine di determinare l'origine e la terminazione delle vie commissurali entro la corteccia (vedi sopra). I fatti sperimentali sono assai semplici: attività elettriche abnormi indotte in un'area della corteccia dall'applicazione diretta di irritanti chimici o fisici tendono a propagarsi all'area omologa dell'altro emisfero, e la propagazione può essere impedita dalla sezione appropriata del corpo calloso (v. Erickson, 1940; v. Kopeloff e altri, 1950), o della commissura anteriore (v. Poblete e altri, 1959) o di entrambi (v. Moruzzi, 1950; v. Bremer e altri, 1956). Come Moruzzi (v., 1939) ha originariamente dimostrato, all'inizio l'attività epilettica del focolaio secondario o speculare assume la semplice forma di una risposta evocata dalle salve commissurali che originano nell'area accessuale primaria. Con il passar del tempo, tuttavia, il focolaio speculare sviluppa una propria attività abnorme, e diviene indipendente dal focolaio controlaterale. Ciò spiega perché l'attività accessuale del focolaio speculare non scompare se la callosotomia viene effettuata parecchie ore dopo l'instaurazione del focolaio primario (v. Morrell, 1960). Ovviamente non si dovrebbe dimenticare che molte altre vie di comunicazione interemisferica sono ancora disponibili dopo sezione delle commissure telencefaliche, sia nel diencefalo che nel tronco dell'encefalo più caudale. Queste vie sono certamente molto meno importanti dei sistemi commissurali diretti per la propagazione interemisferica dell'attività epilettica, ma che esse possano essere in qualche modo interessate in tale propagazione non è affatto escluso. L. T. Rutledge e T. T. Kennedy (v., 1960) hanno dimostrato che risposte a lunga latenza alla stimolazione di un punto corticale possono essere registrate nel punto simmetrico della corteccia controlaterale anche dopo sezione completa del corpo calloso e della commissura anteriore. La via troncoencefalica che si presume mediare questi effetti può essere ugualmente implicata nei fenomeni residui di propagazione interemisferica di attività epilettiche che sono stati talvolta osservati in animali commissurotomizzati (v. Hoefer e Pool, 1943; v. Straw e Mitchell, 1967).
Il dato sperimentale che indica una chiara relazione fra le vie commissurali e la propagazione interemisferica dell'accesso epilettico non è rimasto senza applicazioni cliniche. Alcuni chirurghi (v. Van Wagenen e Herren, 1940; v. Bogen e Vogel, 1962) hanno suggerito che la sezione radicale delle commissure telencefaliche può essere usata in alternativa alla rimozione del focolaio corticale primario per il controllo degli attacchi epilettici. Questo procedimento può essere efficace non solo per impedire la diffusione specifica delle scariche epilettiche da un emisfero all'altro, ma anche per abolire effetti aspecifici di attivazione generale della corteccia promossi dalle connessioni commissurali. Questi effetti facilitatori interemisferici sono stati dimostrati in esperimenti su animali (vedi sopra), ma vi sono anche indicazioni che una aumentata sensibilità agli agenti epilettogeni può svilupparsi in ratti callosotomizzati sotto forma di fenomeno di supersensitività da denervazione (v. Stavraky, 1961). Qualunque possa essere il meccanismo, i resoconti più recenti sulla commissurotomia terapeutica sono stati sostanzialmente ottimistici, anche se gli autori stessi hanno prudentemente dichiarato che il successo è probabilmente dovuto sia alla commissurotomia in sé, sia alla soppressione, da parte della contemporanea terapia farmacologica, di vie sottocorticali per la propagazione interemisferica di attività epilettiche (v. Bogen e altri, 1969). Un altro dato di interesse clinico è costituito dall'osservazione che gli accessi epilettici indotti transinapticamente in un emisfero dall'attivazione delle sue afferenze commissurali sono generalmente meno intensi di quelli provocati dalla stimolazione diretta. Ciò può spiegare la minore azione perturbante della terapia elettrica convulsiva unilaterale, rispetto a quella bilaterale, sulla memoria e su altre funzioni mentali (v. D'Elia, 1970). Il predominante interessamento delle funzioni sostenute dall'emisfero direttamente stimolato e la persistenza relativamente buona delle funzioni mediate dell'altro emisfero suggeriscono effettivamente che la propagazione controlaterale attraverso i sistemi commissurali è meno efficace dell'applicazione diretta della corrente nel disturbare le attività nervose.
Un approccio un po' diverso al problema del ruolo delle connessioni interemisferiche nel determinare il quadro di attività epilettica nella corteccia è stato seguito da E. M. Marcus e C. W. Watson (v., 1966 e 1968; v. anche Marcus e altri, 1968). In una serie di esperimenti eseguiti sul gatto e sulla scimmia, essi hanno applicato agenti epilettogeni, come stricnina, Metrazol, estrogeni coniugati o raffreddamento locale, ad aree neocorticali corrispondenti di entrambi gli emisferi. I focolai di attività abnorme (di solito punte e complessi punta-onda) causati da questo procedimento tendono in generale a presentare scariche notevolmente simmetriche e sincrone. Controlli oscillografici dimostrano che le singole punte e onde lente dei due focolai sono sincronizzate entro un intervallo da 0 a 20 ms. Tuttavia, questa precisa interazione bilaterale non si realizza se gli agenti epilettogeni sono applicati ad aree corticali prive di connessioni commissurali, come sono ad esempio gran parte delle aree striate. In accordo con questi dati, le scariche di focolai bilaterali instaurati in aree corticali provviste di connessioni commissurali perdono la loro previa sincronizzazione e diventano in larga misura indipendenti dopo sezione del corpo calloso e delle commissure anteriore e ippocampale. Solo occasionalmente si osservano in queste condizioni segni di una sincronizzazione molto più grossolana (entro 50 e 400 ms) fra i due lati. Infine, lembi corticali bilaterali totalmente separati dai centri sottocorticali per mezzo di un procedimento chirurgico che risparmia le connessioni reciproche attraverso il corpo calloso, mostrano, quando vengono trattati con applicazioni epilettogene locali, una capacità di emettere scariche bilateralmente sincrone simili a quelle osservate nel cervello normale. Tutti questi risultati forniscono prove sostanziali in favore dell'idea che le commissure telencefaliche abbiano un'importanza primaria per la sincronizzazione di focolai corticali bilaterali di attività elettriche abnormi. Essi sono stati confermati ed estesi da U. Leibowitz (v., 1969) e da V. M. Mosidze e collaboratori (v., 1971), che hanno aggiunto l'osservazione che la sincronia bilaterale delle scariche epilettiche corticali prodotte da iniezioni endovenose di alte dosi di stricnina è ugualmente abolita dalla callosotomia; e da C. A. Ottino e collaboratori (v., 1971), che hanno descritto che la corrispondenza bilaterale fra le scariche epilettiche insorgenti in punti corticali simmetrici può ricomparire, dopo commissurotomia, se l'intensità delle scariche stesse è appropriatamente aumentata. In queste condizioni la sincronia bilaterale sembra essere dovuta a connessioni crociate diencefaliche e mesencefaliche, dato che una asincronia definitiva si ottiene con una sezione mediosagittale che si estenda caudalmente fino al tegmento mesencefalico.
b) Interazioni interemisferiche specifiche rivelate da esperimenti elettrofisiologici
In accordo con una tendenza generale della ricerca neurofisiologica, l'analisi fisiologica delle funzioni specifiche delle commissure telencefaliche nell'ultimo trentennio ha avuto in larga misura carattere elettrofisiologico e il suo sviluppo è passato attraverso vari stadi chiaramente corrispondenti a progressi metodologici nell'investigazione dell'attività elettrica del sistema nervoso. Dapprima l'attenzione si è concentrata sullo studio dei macropotenziali evocati nella corteccia di un emisfero dalla stimolazione della corteccia dell'emisfero opposto.
Un singolo impulso elettrico applicato a quasi tutte le aree corticali produce un potenziale controlaterale complesso che raggiunge la sua massima ampiezza al punto corticale simmetrico e, almeno per quanto riguarda le sue componenti più precoci, stabili e prominenti, è mediato o dal corpo calloso o dalla commissura anteriore, a seconda dei punti di stimolazione e di registrazione. Questi potenziali evocati transcommissurali sono stati studiati a fondo nel gatto (v., ad es., i contributi di Curtis, 1940, e quelli di Chang, 1953; v. Bremer, 1953 e 1955; v. Peacock, 1957; v. Grafstein, 1959 e 1963; v. Rutledge e Kennedy, 1960; v. Latimer e Kennedy, 1961; v. Hossman, Untersuchungen über..., 1969), ma i risultati ottenuti in altre specie sono essenzialmente simili (v., ad es., Nelson e Lende, 1965; v. Latimer e Wilfred, 1968; v. Meyerson, 1968; v. Putnam e altri, 1968). Queste ricerche sono state estremamente utili per il completamento degli studi anatomici delle connessioni interemisferiche e per la facilitazione dell'interpretazione della fisiologia generale della corteccia sia nel corso dello sviluppo sia dopo la maturazione, ma parecchi problemi sorgono quando i risultati devono essere confrontati con quelli ottenuti con altre tecniche fisiologiche, come la registrazione microelettrodica, o con la sperimentazione neurocomportamentale. Alcune ragioni che stanno alla base di questi problemi sono: 1) la tradizionale difficoltà che si incontra nel tentativo di correlare i macropotenziali elettrici registrati da una data regione cerebrale con le attività individuali dei neuroni sottostanti; 2) la grave incompletezza delle nostre conoscenze della fine anatomia delle strutture corticali che emettono e ricevono proiezioni commissurali; 3) le marcate differenze fra le specie nell'organizzazione corticale e, nello schema delle connessioni interemisferiche, differenze che sono state spesso trascurate a causa delle somiglianze nei potenziali evocati, con il risultato che si sono fatte generalizzazioni illecite; 4) la natura notevolmente complessa dei potenziali transcommissurali che, anche quando sono prodotti dalla stimolazione selettiva di un singolo sistema di fibre, sono sempre contaminati dall'attivazione antifisiologica (antidromica) dei neuriti e dall'eccitazione non-commissurale di collaterali assonici ricorrenti che ne deriva (v. ad es., Asanuma e Okamoto, 1959; v. Clare e altri, 1961; v. Feeney e Orem, 1971). In questo paragrafo, l'accento verrà posto sui dati fisiologici che rivestono importanza per gli esperimenti neurocomportamentali su cui si riferirà in seguito; gli studi sui potenziali evocati saranno menzionati solo quando serviranno alla presente discussione. Eccellenti rassegne sui potenziali evocati transcommissurali sono state pubblicate altrove (v. Bremer e altri, 1956; v. Meyerson, 1968; v. Doty e Negrao, 1973; v. Cuénod, 1972). Un passo importante verso il miglioramento delle correlazioni fra le funzioni delle commissure telencefaliche e i potenziali transcommissurali è stato compiuto da Bremer (v., 1958 e Les corps calleux..., 1966), che ha introdotto il concetto del trasferimento elettrofisiologico d'informazioni sensoriali da parte del corpo calloso: la stimolazione di un nucleo talamico di relè (come i corpi genicolati laterale o mediale) evoca una risposta elettrica non solo nell'area di ricezione ipsilaterale della corteccia (attraverso le vie talamo- corticali), ma anche nell'area corticale corrispondente dell'altro lato, attraverso la corteccia ipsilaterale e il corpo calloso. Questo risultato, che è stato ottenuto per le cortecce somestesica, visiva, uditiva e associativa con la stimolazione dei relè talamici appropriati (v. Bremer, 1953; v. Bremer e Terzuolo, 1955; v. Bremer e Stoupel, 1956; v. Bremer, Étude électrophysiologique..., 1966) indica chiaramente che le informazioni sensoriali che raggiungono la corteccia di un emisfero danno invariabilmente origine a una salva callosale destinata alle aree corticali corrispondenti (e funzionalmente correlate) dell'altro lato. Come vedremo, questo trasferimento elettrofisiologico può avere una considerevole importanza per l'interpretazione degli esperimenti comportamentali sul trasferimento callosale di pratiche apprese. Con l'avvento delle tecniche microelettrodiche e i progressi nell'analisi dei campi recettivi sensoriali di singoli neuroni, il trasferimento elettrofisiologico d'informazioni sensoriali da parte del corpo calloso secondo Bremer è stato fatto oggetto recentemente di numerosi esperimenti in cui si sono utilizzate la stimolazione naturale e la registrazione da unità neuroniche.
Nella sfera visiva, questi esperimenti microfisiologici danno risultati in ottimo accordo con i dati anatomici che indicano una relazione selettiva delle connessioni callosali della corteccia visiva con il meridiano verticale centrale del campo visivo. Il grosso dei dati è stato ottenuto nel gatto, ma ci sono buone ragioni per ritenere che essi valgano complessivamente per molte altre specie di Mammiferi (v. Berlucchi, 1972). In breve: 1) registrazioni macro e microelettrodiche da una corteccia visiva deafferentata con la sezione del tratto ottico ipsilaterale dimostrano che è ancora possibile suscitare risposte agli stimoli luminosi, a patto che questi stimoli siano presentati vicino alla linea mediana del campo visivo; queste risposte dipendono da una eccitazione convogliata dal corpo calloso, perché spariscono quando si raffredda lo spienio o il punto corticale simmetrico dell'altro lato; inoltre esse sono limitate ad aree corticali di cui si sa per certo che ricevono normalmente afferenze dirette (retino-genicolo-corticali) dalla regione del meridiano verticale del campo visivo (v. Choudhury e altri, 1965; v. Vesbayeva e altri, 1967); 2) singole unità isolate dallo splenio del corpo calloso hanno campi recettivi visivi situati invariabilmente sulla linea mediana verticale del campo visivo generale o nelle sue immediate vicinanze, e le caratteristiche di questi campi recettivi callosali non si distinguono da quelle osservate per i neuroni delle varie suddivisioni della corteccia visiva (aree corticali 17, 18, e 19 e area soprasilviana laterale) (v. Berlucchi e altri, 1967; v. Huhel e Wiesel, 1967; v. Berlucchi, 1972): ciò implica che tutte le suddivisioni citoarchitettoniche della corteccia visiva proiettano al corpo calloso, ma solo con le loro porzioni che hanno a che fare con la rappresentazione della linea mediana verticale del campo visivo; 3) le informazioni visive trasmesse dal corpo calloso convergono con quelle portate dalla via diretta dal genicolato laterale alla corteccia visiva, e ciò dà luogo alla formazione di campi recettivi che stanno a cavallo della linea mediana, che sono formati da una porzione controlaterale dovuta alle afferenze genicolo-corticali e da una ipsilaterale dovuta alle afferenze callosali (v. Berlucchi e Rizzolatti, 1968; v. Berlucchi, 1972); simili convergenze sono state descritte per la corteccia associativa visiva nel giro soprasilviano (v. Dow e Dubner, 1971). In conclusione, queste connessioni callosali della corteccia visiva assicurano un legame fra le rappresentazioni corticali degli emicampi visivi destro e sinistro, aventi sede rispettivamente negli emisferi sinistro e destro. Questo legame può servire sia alla continuità percettiva nello spazio visivo (v. Whitteridge, 1965; v. Berlucchi, 1972) sia per la percezione della profondità lungo il piano mediosagittale (v. Berlucchi, 1972, e gli esperimenti comportarnentali descritti più avanti).
Un principio di organizzazione simile sembra valere anche per le connessioni callosali della corteccia motoria e somestesica, stando a quanto indicano lavori eseguiti ancora nel gatto. A. L. Towe e collaboratori (v. Tyner e Towe, 1970; v. Nyquist e Towe, 1970) hanno studiato le influenze interemisferiche sui neuroni nell'area dell'arto anteriore della corteccia pericruciata (motoria). Sulla base delle loro proprietà di risposta, questi neuroni possono essere divisi in tre gruppi. I neuroni sb rispondono alla stimolazione elettrica o naturale (tattile) di un'area ben definita nell'arto anteriore controlaterale; questi neuroni non rispondono alla stimolazione elettrica dell'area corticale simmetrica dell'altro lato, e le loro proprietà di risposta non sono influenzate dalla ablazione della corteccia motoria e somestesica controlaterale. I neuroni sb rispondono alla stimolazione elettrica di entrambi gli arti anteriori, e i loro campi recettivi mappati con la stimolazione naturale hanno la forma di un pullover, dato che includono entrambi gli arti anteriori e si estendono su entrambi i lati della linea mediana della parte superiore del tronco. Questi neuroni rispondono, e con brevi latenze, alla stimolazione corticale controlaterale, e la loro risposta alla stimolazione elettrica della zampa anteriore ipsilaterale va persa dopo ablazione della corteccia motoria e somestesica controlaterale; non sappiamo se la parte ipsilaterale del campo recettivo a forma di pullover scompaia ugualmente in queste condizioni, ma che ciò avvenga è assai probabile. I neuroni m, moltissimi dei quali sono tributari del tratto piramidale, rispondono a stimoli elettrici applicati a quasi tutti i punti della superficie cutanea. Essi rispondono anche alla stimolazione corticale controlaterale, anche se con latenze più lunghe rispetto ai neuroni sb, ma non perdono interamente la loro reattività alla stimolazione elettrica delle zampe ipsilaterali dopo ablazione della corteccia motoria e somestesica controlaterale; tale reattività è semplicemente ridotta, o ristretta a uno solo degli arti ipsilaterali (v. Tyner e Towe, 1970; v. Nyquist e Towe, 1970). Sembra che i neuroni sb manchino nella prima area somatosensitiva (S I), ma ve ne sono molti nella seconda area somatosensitiva (S II) (v. Morse e Vargo, 1970); secondo recenti risultati di D. L. Robinson e R. W. Doty (v., 1972), il corpo calloso sarebbe essenziale per fornire le afferenze ipsilaterali a questi neuroni. Riassumendo, i neuroni sb dell'area motoria e dell'area S II riceverebbero afferenze da un campo recettivo controlaterale attraverso il nucleo talamico di relè ipsilaterale, probabilmente da vie lemniscali, e afferenze combacianti da un campo recettivo ipsilaterale attraverso la corteccia controlaterale e il corpo calloso: come nel sistema visivo (v. sopra) il risultato è un campo recettivo totale che si estende da entrambi i lati della linea mediana. I neuroni m sono presenti sia nell'area S I che in quella S II (v. Innocenti e altri, 1972); essi riceverebbero afferenze periferiche bilaterali dal talamo (possibilmente in relazione a vie extralemniscali) e le loro afferenze callosali rinforzerebbero semplicemente le afferenze periferiche ipsilaterali dal talamo (v. Nyquist e Towe, 1970; v. Innocenti e altri, 1972).
Considerazioni anatomiche (v. sopra, cap. 2, § b) suggeriscono che un piano d'interconnessioni callosali simile a quello della corteccia visiva e somestesica possa esistere anche nella corteccia uditiva; non vi sono tuttavia esperimenti fisiologici indicanti che le connessioni callosali abbiano un ruolo nell'integrazione sensoriale attraverso la linea mediana dello spazio uditivo. Altre possibili funzioni di queste connessioni saranno discusse nel paragrafo sugli esperimenti comportamentali.
Abbiamo qualche informazione sulla possibilità che il corpo calloso causi o moduli scariche corticifughe. Lavori precedenti avevano dimostrato che scariche lungo il tratto piramidale (v. Purpura e Girado, 1959) e lungo la via discendente dalla corteccia visiva al corpo genicolato laterale (v. Ajmone Marsan e Morillo, 1961) possono essere messe in moto da salve transcommissurali dirette rispettivamente alla corteccia sensomotoria e visiva. In gatti anestetizzati con cloralosio, scariche nel tratto piramidale ipsilaterale possono essere causate da attività callosali prodotte dalla stimolazione tattile unilaterale; i neuroni del tratto piramidale implicati in questa risposta sono soprattutto i neuroni del tessuto precruciato, che hanno una connessione abbastanza diretta con i motoneuroni spinali (v. sopra; v. sistema piramidale; v. Patton e altri, 1962; v. Towe e altri, 1969; v. Nyquist e Towe, 1970). Questa attivazione callosale dei neuroni precruciati del tratto piramidale permetterebbe una pronta attivazione bilaterale della muscolatura degli arti alla stimolazione tattile unilaterale. Al contrario, è molto modesta l'influenza callosale sui neuroni del tratto piramidale situati nel tessuto postcruciato, che proiettano agli interneuroni sensoriali nel midollo spinale e probabilmente controllano le afferenze del lato del corpo da cui ricevono informazioni sensitive dirette (v. Nyquist e Towe, 1970; v. sistema piramidale). Alcuni dei circuiti interemisferici non callosali che possono servire all'attivazione bilaterale del tratto piramidale sono stati descritti, sia per il gatto (v. Patton e altri, 1962; v. Eidelberg, 1969) sia per la scimmia (v. Eidelberg, 1969). I dettagli dell'organizzazione delle afferenze callosali ai neuroni del tratto piramidale nel gatto sono stati studiati con metodi intracellulari. In breve: 1) i neuroni del tratto piramidale sono eccitati da neuroni callosali situati in una piccola area nel punto esattamente omologo dell'emisfero opposto, e inibiti da neuroni callosali situati in un area corticale più estesa, concentrica all'area eccitatoria (v. Asanuma e Okuda, 1962); 2) i grandi neuroni del tratto piramidale (neuroni ‛rapidi') ricevono grandi fibre callosali con alta velocità di conduzione, e i piccoli neuroni del tratto piramidale (neuroni ‛lenti') ricevono piccole fibre callosali, con bassa velocità di conduzione (v. Nakamura e altri, 1971 v. Naito e altri, 1971); 2) i neuroni del tratto piramidale sono eccitati monosinapticamente e inibiti disinapticamente dalle fibre callosali: le sinapsi inibitorie stanno sul corpo cellulare o sul tronco dendritico, le sinapsi eccitatorie stanno sul tronco dendritico o su arborizzazioni dendritiche più distali (v. Nakamura e altri, 1971).
Simili studi intracellulari sulla corteccia visiva del gatto (aree 17 e 18) hanno dimostrato che: a) le cellule che proiettano al calloso sono situate principalmente nel III strato corticale, in contrasto alle cellule che proiettano al collicolo superiore e al corpo genicolato laterale, che stanno prevalentemente nel V strato corticale; b) le fibre callosali sono in relazione sinaptica con cellule che danno origine a fibre d'associazione intercorticale; c) gli effetti postsinaptici prodotti in queste cellule dalla stimolazione della corteccia visiva controlaterale o del calloso sono eccitazione monosinaptica e inibizione disinaptica (v. Toyama, Tokashiki e Matsunami, 1969; v. Toyama, Matsunami e Ohno, 1969).
4. Esperimenti neurocomportamentali sulle comunicazioni interemisferiche
Molto di ciò che sappiamo sulle comunicazioni interemisferiche proviene dai cosiddetti esperimenti sul ‛cervello bisecato'. Questi esperimenti, che sono stati eseguiti in molti animali, incluso l'uomo, richiedono la sezione chirurgica di tutte le connessioni interemisferiche o di parte di esse. Prima di descrivere questi esperimenti, è necessario chiarire che le vecchie descrizioni di gravi deficit di senso, di moto e comportamentali, prodotti negli animali dalla commissurotomia telencefalica (v. Mingazzini, 1922; v. Bremer e altri, 1956), possono essere ignorate per il fatto che tutti questi deficit erano stati causati da danni extracommissurali involontariamente inferti durante l'operazione di commissurotomia. La sezione delle commissure cerebrali produce effettivamente disturbi importanti e specifici, ma di solito questi disturbi non si manifestano nel comportamento ordinario e devono essere portati alla luce da esami specializzati.
a) Esperimenti sugli animali con cervello bisecato
Il piano di base di questi esperimenti è il seguente: una risposta viene condizionata con la stimolazione di una via di senso che proietta esclusivamente a un emisfero; si esamina poi la ritenzione della risposta applicando lo stesso stimolo alla via di senso corrispondente che va all'altro emisfero. Se il trasferimento è presente, l'importanza dei sistemi commissurali per la mutuazione interemisferica della risposta viene esaminata eseguendo lo stesso esperimento in animali previamente operati di commissurotomia parziale o completa.
La letteratura su questi esperimenti, a partire dallo studio precursore di Bykoff nel 1924, è cresciuta fino a diventare un enorme corpo di nozioni che, sebbene non sia affatto libero da contraddizioni, favorisce chiaramente l'ipotesi secondo la quale le commissure telencefaliche giuocano un ruolo cruciale nel trasferimento interemisferico dell'esperienza e della pratica uniemisferiche. I risultati più importanti saranno descritti in relazione alle differenti modalità di senso.
Nella visione, gli esperimenti paradigmatici sono stati eseguiti sul gatto da R. E. Myers (v., 1956) nel laboratorio di R. W. Sperry. Nel gatto, come nei Mammiferi superiori, i nervi ottici s'incrociano solo in parte, cosicché ciascuna retina proietta ad entrambi gli emisferi. Tuttavia, le fibre ottiche crociate possono essere interrotte sezionando il chiasma ottico lungo la linea mediana, limitando così le afferenze visive da ciascun occhio all'emisfero ipsilaterale. Il gatto con ‛chiasma bisecato' può essere addestrato ad eseguire una discriminazione di forme visive (per es. a scegliere una croce o un cerchio) con un occhio coperto da una maschera di gomma, e poi sottoposto a un esame di trasferimento interoculare spostando la maschera dall'occhio non allenato o inesperto a quello allenato o esperto. Anche se la prestazione con l'occhio inesperto spesso non raggiunge immediatamente il livello finale ottenuto con l'altro occhio, il gatto con chiasma sezionato presenta sempre un buon trasferimento interoculare della capacità discriminativa specifica. L'esperienza acquisita attraverso l'occhio allenato e l'emisfero corrispondente deve essere messa in qualche modo a disposizione della coppia occhio-emisfero dell'altro lato, e poiché le vie retino-cerebrali dei due lati sono completamente separate alla periferia, lo scambio d'informazioni necessario per il buon esito nelle prove di trasferimento interoculare deve aver luogo a un livello centrale, interemisferico. La struttura necessaria e sufficiente per questo trasferimento interemisferico fu identificata da Myers (v., 1956) ripetendo lo stesso esperimento in gatti che erano stati previamente sottoposti a una sezione combinata del chiasma ottico e del corpo calloso. Quando furono esaminati nella prova di trasferimento interoculare, questi gatti con cervello bisecato dimostrarono di essere incapaci di eseguire con l'occhio inesperto la risposta discriminativa acquisita attraverso l'altro occhio (v. Myers, 1956). Ovviamente essi potevano essere riaddestrati, e il riapprendimento attraverso il secondo occhio richiedeva approssimativamente lo stesso numero di prove che erano state necessarie per l'apprendimento originario con il primo occhio (v. Sperry e altri, 1956). In altre parole, sembra che l'apprendimento visivo monoculare nel gatto con il cervello bisecato avvenga solo nell'emisfero connesso con l'occhio che vede, e che l'altro emisfero non riceva alcuna informazione sul processo d'apprendimento. In uno studio successivo Myers (v., 1959) riuscì a dimostrare che basta sezionare la parte caudale (spleniale) del corpo calloso per bloccare il trasferimento interoculare di discriminazione di forme apprese monocularmente da gatti con chiasma sezionato. Una sezione dei due terzi anteriori del calloso che lasci intatto lo splenio non interferisce con il trasferimento interoculare. Esperimenti simili eseguiti in macachi (v. i contributi di Sperry, 1961; v. Downer, 1962) e in scimpanzé (v. Black e Myers, 1964) hanno dato risultati in buon accordo con quelli ottenuti nel gatto. L'unica differenza di rilievo fra gatto e Primati consiste nel fatto che non solo lo splenio, ma anche la commissura anteriore devono essere tagliati, sia nello scimpanzé (v. Black e Myers, 1964) sia nel macaco (v. Noble, 1968) al fine d'abolire il trasferimento interemisferico di discriminazione di forme. È possibile che l'importanza della commissura anteriore per il trasferimento interemisferico dell'apprendimento visivo nei Primati dipenda dal grande numero di fibre neocorticali che si incrociano in questa commissura e interconnettono aree temporali importanti per la visione. L'indipendenza dei due emisferi separati dei gatti e dei Primati con cervello bisecato nell'apprendimento visivo è brillantemente dimostrata da esperimenti in cui questi animali apprendono, in sequenze (v. Myers, Transmission of visual..., 1962, gatto) o simultaneamente (v. Trevarthen, 1962, scimmia), discriminazioni opposte con i due occhi. Per esempio, un gatto o una scimmia con cervello bisecato possono imparare, senza apparenti segni di conflitto, ad avvicinarsi a una croce e a evitare un cerchio quando usano l'occhio destro, e a fare esattamente l'opposto quando usano l'occhio sinistro. Non c'è bisogno di dire che animali normali o anche con il chiasma sezionato, esaminati nello stesso modo, non riescono ad adattarsi alla situazione conflittuale in quanto tendono a rispondere nello stesso modo indipendentemente da quale occhio impieghino (v. Myers, Transmission of visual..., 1962).
Si dovrebbe ricordare a questo punto che dopo sezione delle commissure telencefaliche, i due emisferi cerebrali potrebbero ancora scambiarsi informazioni visive usando le molte altre vie di comunicazione che sono state già menzionate sopra (v. cap. 2, § a). Tutte le prove sopra descritte indicanti una completa separazione funzionale degli emisferi cerebrali in gatti e Primati con cervello bisecato si riferiscono all'apprendimento di discriminazioni di forme visive. Il problema è ora quello di vedere se altre classi di discriminazioni visive (luminanza, colore ecc.) possano essere trasferite fra gli emisferi anche senza le commissure telencefaliche. Parecchi esperimenti sono stati condotti al fine di chiarire questo punto; per semplificazione, i risultati sono riassunti nella tab. I. Nonostante parecchie contraddizioni, il consuntivo dei dati sembra appoggiare l'idea che, al contrario delle discriminazioni di forme, le discriminazioni basate sulla luminanza o sul colore possono essere trasferite fra gli emisferi attraverso interconnessioni diencefaliche o mesencefaliche. Questa nozione è spesso usata per rinforzare le idee tradizionali neurologiche che localizzano la visione delle forme nella corteccia e la percezione del flusso luminoso in centri sottocorticali, ma questa generalizzazione si espone a varie critiche. La presenza e l'assenza del trasferimento interemisferico dell'apprendimento visivo di animali con cervello bisecato dipendono non solo dalla natura della discriminazione visiva in esame, ma anche dalla sua difficoltà, da variabili relative all'addestramento e alla motivazione, dall'età e dalla predisposizione dell'animale, e così via (v. Ettlinger e Blakemore, 1969). Per esempio, Sechzer (v., 1964) ha dimostrato che il trasferimento interoculare di una discriminazione di forme non avveniva in gatti con sezione del chiasma e del corpo calloso se la motivazione per risolvere il problema era costituita da un premio in cibo, come negli esperimenti originari di Myers (v., 1956; v. sopra); se invece la motivazione consisteva nell'evitare una scossa, il trasferimento interoculare era positivo. Analogamente, in una variante degli esperimenti sul cervello bisecato, R. J. Oavalas e R. W. Sperry (v., 1969) hanno osservato che scimmie con sezione del chiasma e delle commissure telencefaliche potevano eseguire una discriminazione fra due forme presentate separatamente, ma nello stesso tempo, ai due occhi. Questa capacità sembra invece mancare nel gatto con cervello bisecato (v. Robinson e Voneida, 1970). Questi esperimenti suggeriscono che, date le giuste condizioni, anche le informazioni relative alla forma possono essere trasmesse attraverso la linea mediana a livelli sottocorticali. D'altra parte, alcuni dei dati presentati nella tab. I indicano chiaramente che in altre circostanze la commissurotomia telencefalica può impedire il trasferimento di discriminazioni visive non basate sulla forma, così come biocca il trasferimento di discriminazioni di forme. Sembra perciò prematuro cercare di stabilire una corrispondenza definita fra il trasferimento interemisferico di differenti discriminazioni visive e i vari livelli delle connessioni commissurali (v. Trevarthen, 1970).
di Giuseppe Moruzzi
Sonno
sommario: 1. Introduzione storica. 2. Fenomenologia del sonno: a) il ciclo sonno-veglia; b) sonno sincronizzato e sonno desincronizzato. 3. Effetti di lesioni: a) la formazione reticolare ascendente; b) le regioni deattivanti del tronco dell'encefalo; c) alternanza di attività e connessioni reciproche tra sistemi antagonisti. 4. Effetti delle stimolazioni periferiche e centrali: a) stimolazioni periferiche; b) stimolazioni centrali. 5. Ricerche neurochimiche: a) l'approccio umorale alle ricerche sul sonno; b) i mediatori chimici del tronco dell'encefalo; c) la dottrina monoamminergica del ciclo sonno-veglia. 6. Registrazioni elettrofisiologiche da singoli neuroni del tronco dell'encefalo. 7. Il significato funzionale del sonno: a) le teorie del ristoro ipnico; b) significato del sonno desincronizzato. 8. Sonno e comportamento istintivo. 9. Interpretazioni neurofisiologiche nella patologia del sonno: a) narcolessia e cataplessia; b) letargo della malattia del sonno e coma; c) le insonnie. 10. Stati simili al sonno: a) l'ipnosi animale; b) l'ibernazione dei Mammiferi. □ Bibliografia.
1. Introduzione storica
È passato poco più di mezzo secolo da quando C. von Economo (v., 1918) riferiva che nell'encefalite letargica epidemica, o malattia del sonno, le lesioni sono localizzate soprattutto nella regione del mesencefalo che circonda l'acquedotto di Silvio e nell'ipotalamo. L'interpretazione da lui data delle sue osservazioni (v. von Economo, 1929) era errata, ma egli era certo nel giusto quando pensava che le strutture nervose lese dalla malattia epidemica avevano un'importanza critica nella regolazione del ciclo sonno-veglia.
Non molti anni dopo W. R. Hess (v., 1927) trovava che il sonno poteva essere ottenuto, nel gatto senza narcosi e libero nei suoi movimenti, con stimolazioni elettriche appropriate della regione mediale del talamo. Malgrado la possibilità di errori (v. cap. 4), questo indirizzo di ricerca ebbe un influsso profondo sullo sviluppo della fisiologia del sonno. Era la prima dimostrazione che il sonno fisiologico può essere ottenuto con stimolazioni elettriche localizzate del cervello, e i risultati dovevano essere più volte confermati stimolando altre regioni del diencefalo e del tronco dell'encefalo. Inoltre gli esperimenti di Hess aprivano la strada a una feconda linea di ricerca in cui lo studio del comportamento era associato a diversi tipi di stimolazione e di registrazione elettrofisiologica. Naturalmente era impossibile individuare, in quei tempi, un qualsiasi rapporto fra le osservazioni di von Economo e gli esperimenti di Hess. Tutto sembrava tanto differente: casi clinici ed esperimenti su animali, lesioni e stimolazioni, letargia e sonno fisiologico.
La scoperta di H. Berger (v., 1929) che l'attività elettrica della corteccia cerebrale poteva essere registrata, nell'uomo, attraverso i tegumenti cranici (elettroencefalogramma, EEG) apriva l'era degli studi elettrofisiologici nell'uomo e negli animali, in assenza di narcosi. Lo stesso Berger diede un contributo fondamentale agli studi sul sonno quando dimostrò che l'EEG dell'uomo presenta durante questo stato oscillazioni di potenziale lente e di ampio voltaggio, ed eliminò in tal modo dalla discussione l'ingenua ipotesi che i centri superiori dovrebbero essere silenti quando manca la coscienza. Egli dimostrò inoltre che questo tipo d'attività elettrica è nettamente diverso dalle onde rapide di basso voltaggio (onde β) che caratterizzano la veglia attiva; e anche dalle lente oscillazioni di potenziale a 10/s (onde α) che compaiono durante la veglia rilassata. Infine Berger trovò che sia le onde α sia i ritmi del sonno potevano essere aboliti da stimolazioni sensitive o sensoriali e rimpiazzati dalle onde β: due fenomeni designati, rispettivamente, reazione d'arresto di Berger e risveglio elettroencefalografico. Tutti questi effetti erano generalizzati non limitati quindi all'area corticale corrispondente alla modalità sensitiva stimolata. Il loro brusco insorgere impedì allora di riconoscere i legami con i fenomeni tonici responsabili del mantenimento della veglia. Toccherà a E. D. Adrian (v., 1934) spiegare le osservazioni di Berger sulla base di modelli d'attività dei neuroni corticali. Egli suggerì l'ipotesi che i neuroni della corteccia cerebrale pulsino insieme durante il sonno e la veglia rilassata, e che i ritmi sincronizzati vengano disorganizzati (desincronizzazione) durante la veglia attiva.
Da questi primi lavori sono nate due linee di ricerca. La linea di ‛indagine fenomenologica', essenzialmente basata su osservazioni, divenne estremamente feconda con l'introduzione delle tecniche di registrazione elettroencefalografica e microelettrodica in Mammiferi senza narcosi, liberi nei loro movimenti. Essa portò alla dimostrazione che sono presenti, durante il sonno, nel gatto (v. Klaue, 1937) e nell'uomo (v. Dement e Kleitman, 1957), periodi caratterizzati da desincronizzazione dell'EEG e agli studi sui rapporti fra questa fase di sonno, chiamata sonno desincronizzato o paradosso, e i sogni (ibid.). Lungo le stesse linee di ricerca fenomenologica vanno situate le indagini sul comportamento di singole unità della corteccia cerebrale (v. Evarts, 1962 e 1964); e anche i lavori elettrofisiologici sul tono posturale (v. Jouvet, 1962), sui riflessi spinali (v. Pompeiano, 1965, 1966 e The neurophysiological..., 1967) e sulla trasmissione sensitiva durante il sonno desincronizzato (v. Pompeiano, Sensory inhibition..., 1967).
La seconda linea di ricerca, ‛l'indagine sperimentale', si basa sullo studio dei cambiamenti del ciclo veglia-sonno prodotti da lesioni o da stimolazioni. Molti aspetti di questo campo d'indagine sono stati esaminati da F. Bremer nell'articolo sistema reticolare ascendente. Basti ricordare qui le ricerche sull'encéphale isolé e sul cerveau isolé (v. Bremer, 1935, 1937 e 1938), il lavoro di G. Moruzzi e H. W. Magoun (v., 1949) sul sistema reticolare ascendente e gli esperimenti di D. B. Lindsley e altri (v., 1949 e 1950) sugli effetti di lesioni reticolari; e infine la dimostrazione dell'esistenza nella parte inferiore del tronco dell'encefalo di un sistema deattivante, probabilmente ipnogeno (v. Moruzzi, 1963, per la letteratura). Il sistema reticolare ascendente ha permesso di dare una spiegazione unitaria di osservazioni fra loro apparentemente senza rapporto, come quelle che abbiamo riferito nella prima parte di questa introduzione.
Gli sviluppi sperimentali più recenti riguardano lo studio neurochimico delle strutture del tronco dell'encefalo che controllano il ciclo veglia-sonno e l'indagine elettrofisiologica del comportamento di singole cellule nervose del ponte e della corteccia cerebrale.
2. Fenomenologia del sonno
Lo studio dei meccanismi neurofisiologici e neurochimici del ciclo sonno-veglia, che verranno trattati nei successivi capitoli, deve essere preceduto dalla descrizione dei fenomeni che si osservano quando non si fa alcun tentativo di modificare il sonno o la veglia né il loro ritmico alternarsi. ‟Si dà il nome di osservatore a colui che applica i procedimenti di investigazione semplici o complessi allo studio di fenomeni che egli non modifica e che, di conseguenza, raccoglie così come glieli offre la natura" (v. Bernard, 1865, p. 29). Questo è sempre il primo approccio in ogni campo delle scienze naturali.
Mezzo secolo fa la sola strada aperta ai ricercatori era lo studio del comportamento animale e questa linea d'attacco è tuttora seguita in molte ricerche di fisiologia comparata e in un campo delle scienze naturali, l'etologia. Il comportamento animale è basato soprattutto su movimenti o su posizioni, quindi sull'attività fasica e tonica di muscoli scheletrici e dei corrispondenti motoneuroni. Gli sviluppi moderni delle tecniche elettrofisiologiche hanno reso possibile lo studio del comportamento non solo di grandi popolazioni di neuroni cerebrali (elettroencefalografia), ma anche dell'attività di singole cellule nervose nell'animale senza narcosi, libero nei suoi movimenti. Tuttavia lo scopo della ricerca è sempre lo stesso, sia che noi studiamo il comportamento di fibre muscolari sia quello dei neuroni cerebrali. Noi osserviamo semplicemente quello che succede, senza fare alcun tentativo d'influenzare il ciclo fisiologico sonno-veglia. Naturalmente qualche esperimento, nel senso ristretto della parola (v. cap. 3), è inevitabile anche in questo tipo di ricerca. Può essere utile, infatti, porre specificamente qualche domanda alla natura, come facciamo, ad esempio, quando osserviamo gli effetti di stimoli sensitivi o sensoriali. Il punto fondamentale, tuttavia, è che non si cerca minimamente di modificare il ritmo fisiologico dell'alternanza fra sonno e veglia. Nostro scopo è solo quello di descrivere con cura fenomeni naturali e le loro relazioni nel tempo.
Questo approccio ha portato a quella che può essere chiamata la ‛fenomenologia del sonno'. Noi limiteremo la nostra esposizione a due campi di studio: a) l'alternanza fra sonno e veglia, cioè il ciclo sonno-veglia; b) l'alternarsi durante il sonno comportamentale di due stati diversi, caratterizzati rispettivamente da sincronizzazione e desincronizzazione dell'EEG.
a) Il ciclo sonno-veglia
Per ciò che riguarda il ciclo sonno-veglia gli animali possono essere divisi in due classi. Gli animali monociclici sono caratterizzati da una periodicità circadiana di circa 24 ore, mentre quelli policiclici presentano molte alternanze fra riposo e attività, in modo del tutto indipendente dall'alternanza fra giorno e notte. L'uomo adulto e molti mammiferi e uccelli sono esseri monociclici, mentre topi e ratti presentano molti cicli in un giorno. Il bambino neonato presenta un ritmo policiclico, caratterizzato da risvegli periodici legati soprattutto a necessità di nutrizione: è la veglia di necessità di N. Kleitman. L'evoluzione ontogenetica da questa periodicità primitiva al ritmo monociclico è in rapporto con lo sviluppo della corteccia cerebrale e della capacità di reagire in modo critico, mentre il tempo trascorso nel sonno diminuisce continuamente. La veglia protratta dell'uomo adulto è stata chiamata da Kleitman (v., 1963) veglia evoluta. I brevi periodi di sonno che caratterizzano i primi periodi, policiclici, di vita, si consolidano successivamente in un'unica fase di sonno notturno.
Ognuno conosce le differenze comportamentali fra sonno e veglia. Alcune manifestazioni del sonno nell'uomo che tanto hanno impressionato poeti, scultori e pittori - come la chiusura degli occhi, l'incapacità di mantenere la stazione eretta, il rilassamento dei muscoli - sono epifenomeni fisiologici. I buoi dormono con gli occhi aperti e gli Uccelli dormono appollaiati sui rami. La differenza principale fra sonno e veglia è la mancanza di coscienza durante il sonno, o la presenza di una forma rudimentale di coscienza, se prendiamo in considerazione i sogni. Se invece partiamo dal punto di vista obiettivo, comportamentale, dell'etologia, vediamo che il sonno può essere definito, negativamente, come quel periodo in cui tutti i comportamenti di veglia sono impossibili. Un animale reagisce di solito come un tutto, come un individuo, al mondo fisico o biologico che lo circonda, in particolare a certi animali. Può presentare manifestazioni tipiche di difesa o d'aggressione, o i comportamenti legati alla riproduzione o alla cura dei nati; può pulirsi o nutrirsi. Ciò che chiamiamo veglia è uno stato generale del sistema nervoso centrale che è comune a tutti questi tipi di comportamento, e ciò che chiamiamo sonno è la scomparsa, completa e del tutto reversibile, dello stato di veglia.
Va ricordato, infine, che l'approccio obiettivo dell'elettrofisiologia che è stato brevemente ricordato nell'introduzione storica ci permette lo studio dei tracciati elettroencefalografici, desincronizzati e sincronizzati, che si osservano, rispettivamente, nella veglia e nel sonno.
b) Sonno sincronizzato e sonno desincronizzato
Un altro ciclo si svolge durante il sonno: una fase caratterizzata da oscillazioni di potenziale ampie e lente (sincronizzazione dell'EFO), e inoltre da immobilità del corpo e degli occhi, si alterna con un'altra fase caratterizzata da onde rapide e di basso voltaggio (desincronizzazione dell'EEG) e inoltre da scosse cloniche degli arti e da rapidi movimenti oculari (REM: Rapid Eye Movements). La prima fase è il sonno chiamato sincronizzato o anche classico, perché per molti anni esso è stato la sola forma riconosciuta di sonno; la seconda fase è chiamata sonno desincronizzato o REM, o anche sonno paradosso, con un'espressione che indica la nostra sorpresa per il fatto che uno stadio caratterizzato, dal punto di vista comportamentale, da segni manifesti di sonno profondo possa presentare lo stesso tipo di onde rapide e a basso voltaggio che si osserva durante la veglia. In realtà questo paragone non è più giustificato, giacché J. F. Tönnies (v., 1969) ha dimostrato, con il suo analizzatore per EEG, che i tracciati sono di fatto differenti durante il sonno paradosso e durante la veglia. D'altronde anche l'attività di singole unità corticali è differente nei due tipi di sonno, come vedremo fra poco.
In condizioni fisiologiche il sonno comincia sempre con un periodo sincronizzato, che è interrotto più volte durante la notte da brevi episodi di desincronizzazione. Ciascuno di essi dura circa 6 minuti nel gatto adulto, libero nei suoi movimenti; la percentuale di sonno desincronizzato rispetto a tutto il sonno oscilla fra il 20 e il 25%. Nell'uomo adulto la percentuale varia tra il 18 e il 22%, ma può arrivare fino al 50% nel neonato (v. Roffwarg e altri, 1966).
Dal punto di vista dell'esperienza soggettiva la differenza fra i due stadi del sonno è data dal fatto che in genere abbiamo il ricordo di sogni quando siamo risvegliati durante o subito dopo un periodo desincronizzato.
Dal punto di vista obiettivo, il sonno desincronizzato è caratterizzato da molti fenomeni che colpiscono per la loro evidenza; essi meritano di essere ricordati, per quanto noi siamo ancora incapaci di comprenderne il significato fisiologico. V'è completa flaccidità o atonia di tutti quei muscoli che vengono chiamati ‛antigravitari', perché permettono le posizioni di veglia che fanno ostacolo alla forza di gravità (v. Jouvet, 1962); i riflessi spinali e i movimenti prodotti dalla stimolazione elettrica del fascio piramidale sono bloccati da scariche discendenti di impulsi inibitori provenienti dal tronco dell'encefalo (v. Pompeiano 1965, 1966, e The neurophysiological..., 1967); infine, e ancora una volta paradossalmente, queste potenti scariche inibitrici sono incapaci di bloccare l'eruzione di attività motorie fasiche, rappresentate dalle scosse cloniche degli arti e dalle esplosioni di movimenti rapidi oculari (v. Jouvet, 1967, per la letteratura).
È facile, naturalmente, vedere il significato funzionale dei movimenti osservati durante la veglia: come quelli per la difesa o l'aggressione, o in rapporto alla pulizia o alla nutrizione; ma non siamo ancora in grado di comprendere il significato delle attività fasiche che caratterizzano lo stadio desincronizzato del sonno. Forse ciò è dovuto al fatto che partiamo da un punto di vista sbagliato. Quello che importa nell'animale sveglio è il risultato finale, motore o posturale, dell'attività integrata dei muscoli scheletrici. Naturalmente ci rendiamo conto del fatto che posizioni o movimenti altro non sono che l'espressione periferica di scariche ben determinate all'interno del sistema nervoso centrale. Ma il nostro interesse converge soprattutto su questo effetto periferico, il cui significato fisiologico per la sopravvivenza dell'animale o della specie è sempre facile a comprendersi. I modelli di scarica neuronale che si hanno durante il sonno possono avere il loro significato funzionale per il ristoro. Le scosse cloniche del sonno paradosso sono fenomeni che colpiscono dal punto di vista comportamentale, ma possono essere la conseguenza di fenomeni di liberazione prodotti dall'inattività di neuroni inibitori. Anche il sonno, naturalmente, è un comportamento; ma è essenzialmente un tipo di comportamento introverso. Ciò che importa nelle ricerche sul sonno è l'attività dei neuroni, forse solo di certi tipi di neuroni; assai più dei loro effetti periferici, la contrazione delle fibre muscolari.
3. Effetti di lesioni
La parola ‛esperimento' verrà usata in senso ristretto, allo scopo di fare una netta distinzione concettuale tra la fenomenologia del sonno e i lavori sui meccanismi neurofisiologici del ciclo sonno-veglia. ‟Si dà il nome di sperimentatore a colui che usa i procedimenti di investigazione semplici o complessi per far variare o per modificare, a qualsiasi scopo, i fenomeni naturali e farli comparire in circostanze o in condizioni in cui la natura non glieli presenta. In questo senso l'‛osservazione' è l'esame di un fenomeno naturale e l'‛esperimento' è l'esame di un fenomeno modificato dall'esaminatore" (v. Bernard, 1865, p. 29). In questo capitolo riassumeremo i risultati di esperimenti in cui coma, letargia o insonnia, o qualsiasi cambiamento nel ciclo sonno-veglia, vennero ottenuti mediante lesioni nel tronco dell'encefalo o nel cervello.
a) La formazione reticolare ascendente
Questa linea di ricerca è strettamente legata alla fisiologia del sistema reticolare ascendente. Si rimanda, quindi, all'omonimo articolo di Bremer di cui riassumeremo in breve le principali conclusioni come introduzione a questa parte.
Dobbiamo partire dalla desincronizzazione elettroencefalografica prodotta da stimolazioni sensitive o sensoriali (‟reazione di arresto" di Berger, reazione di risveglio), che abbiamo ricordato nella parte storica di questo articolo. Il cambiamento del tracciato elettroencefalografico viene di solito considerato come il segno di un aumento del livello generale di attività del cervello, e il fenomeno è quindi chiamato attivazione. Moruzzi e Magoun (v., 1949) hanno dimostrato che la stessa attivazione può essere prodotta con la stimolazione elettrica della formazione reticolare del tronco dell'encefalo e hanno provato che questa influenza attivante è mediata da vie differenti da quelle dei classici sistemi sensitivi o sensoriali, che rappresentano il substrato anatomico delle percezioni. Si è visto in seguito che anche gli aspetti comportamentali della reazione di risveglio possono essere riprodotti con la stimolazione della formazione reticolare, in animali senza narcosi e liberi nei loro movimenti (v. Moruzzi, 1972, per la letteratura). Questi sono effetti fasici, di breve durata.
Il passo successivo fatto da Moruzzi e Magoun (v., 1949) fu di suggerire l'ipotesi che il sistema reticolare ascendente fosse continuamente, e cioè tonicamente, attivo e che la sua influenza sul cervello dovesse essere al di sopra di un determinato livello critico per mantenere la veglia. L'interruzione di questa influenza ascendente sarebbe la causa del coma che compare nel gatto dopo la sezione del mesencefalo, nel preparato cerveau isolé di Bremer (v. È c). Questa sindrome si osserverebbe anche nell'uomo dopo una lesione del mesencefalo prodotta da trauma. Interruzioni meno complete del mesencefalo, combinate a lesioni ipotalamiche, produrrebbero la letargia, spiegando così le osservazioni di von Economo. La dimostrazione che un'attività continua, ‛tonica', era presente nel sistema reticolare fu ottenuta seguendo due diverse linee di ricerca: a) riproduzione della sindrome di corna del cerveau isolé con interruzione delle proiezioni reticolari ascendenti (v. Magoun, 19632; v. Moruzzi, 1972, per la letteratura); b) dimostrazione mediante registrazione microelettrodica dell'esistenza di una scarica continua, irregolare, nei neuroni reticolari (v. Moruzzi, 1954, per la letteratura; v. sistema reticolare ascendente).
Un terzo passo in avanti fu compiuto da C. Batini e altri (v., 1959), quando dimostrarono che una sindrome comportamentale ed elettroencefalografica d'insonnia, opposta dunque alla sindrome di coma del cerveau isolé, poteva essere ottenuta sezionando il tronco dell'encefalo pochi millimetri all'indietro, a livello pontino. Questo ‛preparato mediopontino pretrigeminale' è caratterizzato da EEG desincronizzato e dal comportamento alerte degli occhi. Questa osservazione e molte altre fatte seguendo differenti vie di ricerca (v. Moruzzi, 1963, 1972; v. Bonvallet, 1966, per la letteratura) condussero alla conclusione che nel tronco dell'encefalo c'è anche un sistema che può essere chiamato ‛deattivante', perché è antagonista al sistema reticolare ascendente che abbiamo visto invece essere attivante. Si tratta di popolazioni di neuroni dotate di effetti sincronizzanti l'EEG e ipnogeni.
Riassumendo, alla fine degli anni quaranta appariva possibile una spiegazione unitaria di osservazioni fra loro apparentemente non legate, quali quelle di von Economo e di Berger. Questo risultato era dovuto alla dimostrazione di un sistema reticolare ascendente, dotato di un'influenza attivante, tonica e fasica, sul cervello. Alla fine degli anni cinquanta, infine, gli effetti ipnogeni classici, ottenuti con la stimolazione elettrica (v. cap. 4), potevano essere messi in qualche modo in rapporto con influenze deattivanti esercitate da altre strutture del tronco dell'encefalo.
b) Le regioni deattivanti del tronco dell'encefalo
Tutti questi risultati erano stati ottenuti con esperimenti acuti e gli effetti cronici di sezioni del tronco dell'encefalo venivano usati solo per controllo. La sperimentazione cronica cominciò a essere usata soprattutto durante gli anni sessanta. Questi esperimenti portarono a risultati importanti per il problema dell'origine del ciclo sonno-veglia. La dimostrazione che sul cervello vengono esercitate due influenze opposte condusse all'ipotesi che il ciclo stesso, e cioè l'alternarsi del sonno e della veglia, potesse nascere nel tronco dell'encefalo.
La storia degli esperimenti di decerebrazione cronica è lunga (v. Moruzzi, 1972, per la letteratura). Tuttavia noi discuteremo solo i risultati ottenuti da J. Villablanca (v., 1966) su gatti in cui il tronco dell'encefalo era stato separato dal cervello con una sezione fatta a livello più alto, e cioè proprio in fronte ai collicoli superiori (decerebrazione precollicolare). Questi gatti mesencefalici alti venivano seguiti per un lungo periodo di tempo. È naturalmente impossibile definire come sonno o come veglia qualsiasi stato osservato in animali senza cervello. Tutto quello che possiamo dire è che, dopo la decerebrazione cronica, è possibile osservare comportamenti che ricordano quelli dell'animale normale durante il sonno e la veglia. Dopo 15-20 giorni i gatti venivano trovati mentre stavano strisciando o sedevano o addirittura facevano tentativi di camminare; le palpebre erano aperte e le pupille dilatate. Questi erano manifestamente sintomi di veglia e tale impressione veniva rafforzata dal fatto che questi periodi si alternavano con stati caratterizzati da manifestazioni corporee di sonno. Villablanca (ibid.) fece una distinzione fra uno stato caratterizzato dalla chiusura delle palpebre, dal sollevamento della membrana nittitante e, soprattutto, dalla fluttuazione del diametro pupillare (miosi fluttuante) - che corrisponde al sonno sincronizzato dell'animale intatto - e uno stato caratterizzato dal restringimento estremo delle pupille (miosi fessurata) e dal collasso generalizzato del tono posturale, che corrisponde al sonno desincronizzato dell'animale intatto. I periodi caratterizzati dalla scomparsa reversibile della rigidità da decerebrazione sono di solito chiamati ‛episodi cataplessici', dal nome di una sindrome clinica che esamineremo in seguito (v. cap. 9, § a).
Il risultato principale ottenuto da Villablanca con i suoi esperimenti non è la dimostrazione che frammenti di comportamento di sonno o di veglia possono essere osservati in assenza del cervello - questo era già stato visto da altri prima di lui - ma piuttosto la dimostrazione del fatto che sia il ciclo sonno-veglia sia il ritmico alternarsi dei due stadi di sonno possono nascere quando il tronco dell'encefalo è separato dal cervello. Naturalmente solo i nervi cranici e il midollo spinale sono disponibili per le manifestazioni di sonno e di veglia, quando il cervello è assente. Ma è un risultato fondamentale l'aver dimostrato che ritmi di questo tipo possono nascere, in un animale senza cervello, nel tronco dell'encefalo.
La spiegazione ovvia di questi risultati, se si trascura il problema del sonno paradosso, è che vi sia un'alternanza di attività fra due sistemi: il sistema reticolare ascendente o attivante e le regioni deattivanti della parte caudale del tronco dell'encefalo. Ricerche recenti hanno permesso di localizzare almeno due di queste regioni deattivanti: 1) la regione del tratto solitario, che è dotata di attività fasiche, come è dimostrato da esperimenti di lesione e di stimolazione (v. Moruzzi, 1963; v. Bonvallet, 1966, per la letteratura); 2) i nuclei del rafe, che sono tonicamente attivi, come è dimostrato dall'insonnia prolungata prodotta dalla loro lesione (v. Jouvet e Renault, 1966). Si tratta di due sistemi indipendenti, e lo dimostra il fatto che è ancora possibile produrre sincronizzazione dell'FEO e miosi mediante la stimolazione di fibre afferenti vagoaortiche che si portano alla regione del fascio solitario. Questo effetto deattivante è presente anche quando l'incrociamento del rafe è stato interrotto da una sezione sagittale (v. Puizillout e Ternaux, 1974).
c) Alternanza di attività e connessioni reciproche tra sistemi antagonisti
Esperimenti cronici sul cerveau isolé mostrano, d'altra parte, che un ciclo sonno-veglia può nascere anche in un cervello isolato, dopo la sezione completa del mesencefalo. I classici esperimenti di Bremer erano acuti e il risultato principale fu la scoperta che il ciclo sonno-veglia era presente dopo la sezione del midollo cervicale a C1, e cioè quando il cervello era ancora connesso al tronco dell'encefalo (encéphale isolé), mentre il ciclo scompariva, ed era sostituito da un coma ‛permanente' dopo la sezione del mesencefalo (cerveau isolé). Manifestamente o l'abolizione del flusso di impulsi sensitivi e sensoriali attraverso i nervi cranici, secondo quanto aveva suggerito originariamente Bremer (v., 1937 e 1938), o la soppressione di un'influenza ascendente che nasce fra le due sezioni potevano spiegare differenze così evidenti fra questi due preparati acuti. Sappiamo oggi che l'eliminazione dell'influenza tonica del sistema reticolare ascendente è responsabile del coma del cerveau isolé acuto.
Il coma, tuttavia, è un sintomo prolungato, ma certo non permanente, del cerveau isolé, come è dimostrato dagli esperimenti cronici. La prima letteratura, dai lavori di U. Genovesi e altri (v., 1956) a quelli di H. L. Batsel (v., 1960 e 1964), è stata già esposta (v. Moruzzi, 1972). Qui parleremo solo degli esperimenti di Villablanca (v., 1965) sul preparato cerveau isolé alto; questa espressione indica che ogni influenza esercitata sul cervello dal tronco dell'encefalo è stata eliminata con una sezione alta (precollicolare) nel mesencefalo. In questo preparato periodi di desincronizzazione comparivano spontaneamente da 7 a 10 giorni dopo la sezione mesencefalica. Molte osservazioni e molti controlli, discussi a fondo in due rassegne critiche (v. Moruzzi, 1972 e 1974), conducono alla conclusione che in un cervello cronicamente isolato possono essere mantenuti veri periodi di veglia, e può apparire un ciclo sonno-veglia.
Le argomentazioni seguite nella discussione fatta sopra, a proposito dei risultati della decerebrazione cronica, condurrebbero all'ipotesi di un'alternanza fra strutture attivanti e deattivanti localizzate nel cervello. Gli esperimenti classici di S. W Ranson (v., 1939) sembrerebbero suggerire che le strutture attivanti siano localizzate nell'ipotalamo. Una sindrome di letargo, che ricorda la malattia del sonno della patologia umana, può essere ottenuta, nella scimmia, con lesioni dell'ipotalamo posteriore. D'altra parte, vi sono nel cervello molte regioni che producono il sonno con la stimolazione elettrica. Molte di esse, tuttavia, non hanno probabilmente importanza critica, giacché il ritmo sonno-veglia non è abolito dalla decorticazione o dalla talamectomia. Forse la struttura deattivante del cervello che più facilmente contribuisce al ritmo è l'area preottica e la parte basale del cervello anteriore, la cui lesione è seguita da insonnia (v. McGinty e Sterman, 1968; v. Moruzzi, 1972, per la letteratura).
Riassumendo, sia nel tronco dell'encefalo sia nel cervello vi sono coppie di sistemi antagonisti che producono effetti opposti sul ciclo sonno-veglia. Nel preparato cronico decerebrato un'alternanza fra i due tipi di attività è dovuta a meccanismi intrinseci al tronco dell'encefalo; nel preparato cerveau isolé cronico la stessa alternanza è dovuta unicamente a meccanismi cerebrali. Questi risultati, naturalmente, dimostrano solo una potenzialità di comportamento ritmico; tuttavia entrambi questi effetti implicano l'esistenza di connessioni reciproche fra, sistemi antagonisti. Il problema principale, ovviamente, e di sapere cosa avviene quando il tronco dell'encefalo è connesso al cervello, come nell'animale normale. Si possono fare due ipotesi. Il ciclo sonno-veglia potrebbe nascere nel tronco dell'encefalo ed essere imposto al cervello attraverso vie ascendenti. Questo è il nucleo della dottrina monoamminergica di Jouvet, che esamineremo nella parte dedicata alla neurochimica. Secondo un'altra ipotesi alternativa, il ciclo sonno-veglia potrebbe nascere nel cervello ed essere semplicemente controllato dall'afflusso di impulsi ascendenti provenienti dal tronco dell'encefalo. Naturalmente si possono verificare molte interazioni fra tronco dell'encefalo e cervello, quando l'animale è intatto. Questi sono tra i maggiori problemi della fisiologia del sonno e verranno di nuovo esaminati alla fine di quest'articolo.
4. Effetti delle stimolazioni periferiche e centrali
a) Stimolazioni periferiche
L'azione ipnogena di stimolazioni sensitive e sensoriali monotone è ben conosciuta. Pavlov aveva spesso osservato la comparsa del sonno negli esperimenti di condizionamento, ma la sua spiegazione del sonno ha ormai solo interesse storico (v. Moruzzi, 1960, 1964 e 1972) Secondo J. Konorski (v., 1967) possono condurre alla sonnolenza e al sonno sia meccanismi innati sia meccanismi condizionati. Tutti i tipi di stimolazione sensitiva e sensoriale monotona appartengono agli stimoli innati. Essi sono stati riprodotti da O. Pompeiano e J. E. Swett (v. i contributi del 1962) con la stimolazione elettrica ripetuta di fibre nervose cutanee del gruppo Il, probabilmente in rapporto a recettori del tatto, della pressione e dei peli. Anche gli stimoli condizionati sono importanti per la nascita del sonno, soprattutto nell'uomo; essi ‟si stabiliscono generalmente in rapporto al tempo, secondo le modalità stereotipe che si verificano ogni giorno, quando ci si addormenta" (v. Konorski, 1967, p 300). Le abitudini individuali riguardanti il modo di addormentarsi, tanto importanti in molti uomini, sono probabilmente in rapporto a meccanismi condizionati.
Impulsi prodotti dalla stimolazione naturale dei barocettori senocarotidei o cardioaortici (v Dell, 1971) o dalla stimolazione elettrica di fibre barocettive vagoaortiche (v. Puizillout e Ternaux, 1974; v. Puizillout e Foutz, 1977) scatenano in via riflessa il sonno. Nel preparato ‛encefalo isolato' la stimolazione di fibre afferenti barocettive vagoaortiche permette di riprodurre a volontà il sonno sincronizzato seguito da episodi di sonno desincronizzato.
È stata prospettata l'ipotesi (v. Moruzzi, 1960) che meccanismi innati, come quelli messi in moto da stimolazioni sensitive e sensoriali monotone, possano agire reclutando progressivamente i neuroni deattivanti della parte caudale del tronco dell'encefalo. I meccanismi condizionati possono interessare le aree basali del cervello anteriore e l'area preottica, come vedremo fra poco. L'effetto opposto, il risveglio, è prodotto dall'attivazione fasica del sistema reticolare ascendente, che è trascinato più facilmente da stimolazioni sensitive o sensoriali, caratterizzate dall'insorgere rapido e inaspettato. Il sistema reticolare ascendente cessa ben presto di rispondere a stimolazioni sensitive ripetute, come dimostra la scomparsa delle manifestazioni elettroencefalografiche e comportamentali del risveglio, un fenomeno chiamato ‛abituazione' (v. Sharpless e Jasper, 1956). Le strutture deattivanti si comportano in modo esattamente opposto: i loro neuroni vengono reclutati a poco a poco da stimolazioni ripetute (v. Moruzzi, 1960) con un meccanismo ben noto nella neurofisiologia classica, la ‛sommazione temporale'. (Il lettore viene rinviato all'articolo sinapsi: Fisiologia della sinapsi centrale, per ciò che riguarda i meccanismi cellulari della ‛sommazione').
b) Stimolazioni centrali
L'opera di W. R. Hess (v. i contributi in bibliografia) è l'introduzione naturale agli esperimenti di stimolazione centrale. Hess dimostrò che la stimolazione elettrica a bassa frequenza di una regione talamica situata lateralmente alla massa intermedia era seguita dalla comparsa del sonno: un genuino fenomeno fisiologico, come dimostra il risveglio che si osserva quando si mette della carne sotto al naso dell'animale. N. Tinbergen (v., 19552) ha osservato che sia il sonno naturale sia quello indotto con il metodo di Hess vengono annunziati da uno stato che ricorda la ‛fase appetitiva' che dà inizio a ogni tipo di comportamento istintivo (v. Craig, 1918): il gatto si guarda attorno, cerca un posto per andare a dormire e infine si acciambella nella posizione naturale del sonno. Manifestamente l'animale tende a raggiungere una situazione che gli permetta, o gli faciliti, l'insorgenza del sonno.
Molte critiche che erano state sollevate contro gli esperimenti di Hess sono state adeguatamente controbattute (v. Moruzzi, 1972). La sola obiezione che merita d'essere discussa, soprattutto in ragione della lunga latenza del fenomeno, è che l'animale avrebbe potuto addormentarsi anche senza stimoli. Che questa sia una causa d'errore nessuno potrebbe negare. Tuttavia gli esperimenti sulla stimolazione dell'area preottica e delle aree basali del cervello anteriore - che verranno discussi fra poco - non lasciano dubbio alcuno che il sonno può essere prodotto, con un meccanismo attivo, dalla stimolazione del cervello. Il decorso nel tempo del sonno indotto da Hess, infatti, è inusitato solo se lo si paragona alle risposte classiche della fisiologia sherringtoniana, che cominciano subito dopo l'inizio della stimolazione (tempo latente) e sopravvivono allo stimolo elettrico che le aveva provocate solo per breve tempo (scarica o effetto postumo). Anche i movimenti prodotti dalla stimolazione della corteccia motoria sono un esempio tipico di questo tipo di risposte: l'effetto periferico è guidato dagli impulsi piramidali prodotti dalla stimolazione elettrica. Tuttavia se il sonno indotto è fenomeno genuinamente fisiologico, come sembra essere, esso deve necessariamente iniziare lentamente e sopravvivere allo stimolo. Il sonno di Hess è in realtà non prodotto, ma piuttosto ‛avviato' dalla stimolazione talamica. In altre parole, le salve d'impulsi nervosi prodotte dagli stimoli elettrici mettono in moto processi nervosi innati, responsabili del comportamento di sonno, purché sia disponibile un tempo sufficiente per i lenti processi di facilitazione centrale. Quando questo risultato è stato ottenuto, probabilmente grazie a un rilassamento dell'attività tonica del sistema reticolare ascendente, il processo va avanti da sé, fino a che è interrotto dal risveglio.
L'importanza dei recenti risultati sull'influenza ipnogena della regione basale del cervello anteriore e sull'area preottica è dovuta a diversi fattori: a) il sonno può essere indotto anche da stimolazioni ad alta frequenza (v. i contributi di Sterman e Clemente, 1962); b) il tempo latente è molto più breve, anche di soli 5 s (ibid.); c) associando al solito modo la stimolazione centrale, utilizzata come stimolo incondizionato, a un tono acustico usato come stimolo condizionato, si vede che è possibile condizionare lo stimolo uditivo, dando così avvio a manifestazioni comportamentali ed elettroencefalografiche di sonno (v. Clemente e altri, 1963); d) lesioni della stessa area producono insonnia (v. McGinty e Sterman, 1968). Bremer ha dato una spiegazione di tutti questi effetti, e forse di tutti i casi di sonno indotto, con la dimostrazione che segni elettrofisiologici d'inibizione appaiono nella formazione reticolare in seguito alla stimolazione elettrica dell'area basale del cervello anteriore e dell'area preottica (v. Bremer, 1970, 1975 e 1977). Queste importanti ricerche sono esposte e documentate nell'articolo sistema reticolare ascendente.
5. Ricerche neurochimiche
a) L'approccio umorale alle ricerche sul sonno
Gli esperimenti sull'ipnotossina (v. Piéron, 1913) possono essere considerati come i precursori del moderno approccio neurochimico. Iniezioni di liquido cerebrospinale (LCS), prelevato da cani privati del sonno, dentro la cisterna magna di un altro cane, ovviamente dopo l'aspirazione di una uguale quantità di LCS, producevano un sonno comportamentale della durata di 2-6 ore. Iniezioni di controllo di LCS normale erano senza effetto. Il fattore ipnogeno venne chiamato ipnotossina. Questi risultati furono confermati da J. R. Pappenheimer e altri (v., 1967 e 1971). Perfusioni intraventricolari di LCS ottenuto da capre private di sonno eseguite su ratti prolungavano i periodi di sincronizzazione dell'EEG e deprimevano l'attività locomotoria per circa 6 ore. Altri fattori sono stati trovati da M. Monner nel plasma del sangue durante il sonno prodotto con la stimolazione elettrica del talamo (v. Jouvet, 1972, per la letteratura).
Non abbiamo finora prova alcuna che sostanze originate dalle strutture cerebrali durante la veglia raggiungano i neuroni attivanti o deattivanti del tronco dell'encefalo attraverso le vie umorali e che il loro accumulo nel sangue o nel LCS, in conseguenza di un periodo fisiologico di veglia, sia sufficiente a produrre o a mettere in marcia un sonno genuino. Le prove ora disponibili sono anzi piuttosto contrarie a tale ipotesi, almeno per ciò che riguarda il sangue, giacché non è stato trovato alcun rapporto fra i cicli di sonno-veglia di gemelli siamesi (v. Alekseyeva, 1958). Questo però non esclude che, quando s'impedisce a un animale di dormire per un lungo periodo di tempo, prodotti endogeni che normalmente eserciterebbero solo azioni locali possano passare nel sangue o nel LCS, dando origine ai fenomeni che abbiamo sopra descritto. Ma non vi sono prove che questo meccanismo entri in azione nella regolazione del ciclo veglia-sonno normale.
L'approccio umorale alle ricerche sul sonno può essere nondimeno importante, perché fornisce, in un certo senso, l'immagine ingrandita di fenomeni che in condizioni normali si svolgono solo nella sostanza grigia. I ‛mediatori chimici' sono per definizione molecole che vengono liberate, agiscono e sono distrutte localmente. Il punto centrale è sapere se dentro i sistemi regolatori del tronco dell'encefalo vi sia un accumulo progressivo di molecole ipnogene per effetto di un periodo fisiologico di veglia; se queste sostanze scompaiano per effetto del sonno fisiologico; se infine questi prodotti ipnogeni coincidano con i mediatori chimici dei sistemi deattivanti.
b) I mediatori chimici del tronco dell'encefalo
I mediatori chimici dei sistemi del tronco dell'encefalo che regolano il ciclo sonno-veglia sono stati identificati durante gli ultimi anni, soprattutto per opera di Jouvet e collaboratori. I risultati sono riassunti in rassegne sintetiche recenti, per ciò che riguarda il problema delle monoammine del cervello (v. Axelrod, 1971), con particolare riferimento al sonno (v. Jouvet, 1972).
Noi conosciamo la maggior parte dei mediatori al servizio dei sistemi che nascono nel tronco dell'encefalo, e i metodi di istofluorescenza hanno permesso di fare una mappa dei neuroni che posseggono lo stesso mediatore. Ci occuperemo in questo paragrafo solo del ciclo vegliasonno sincronizzato. Del ciclo sonno sincronizzato-sonno desincronizzato diremo nel capitolo successivo, esponendo i risultati delle moderne ricerche elettrofisiologiche eseguite su singoli neuroni. Pochi esempi di esperimenti su due sistemi monoamminergici, che utilizzano rispettivamente la 5-idrossitriptammina (5-HT) e la noradrenalina (NA) come mediatore, daranno un'idea delle potenzialità dell'approccio neurochimico, anche senza dettagliate esposizioni e discussioni.
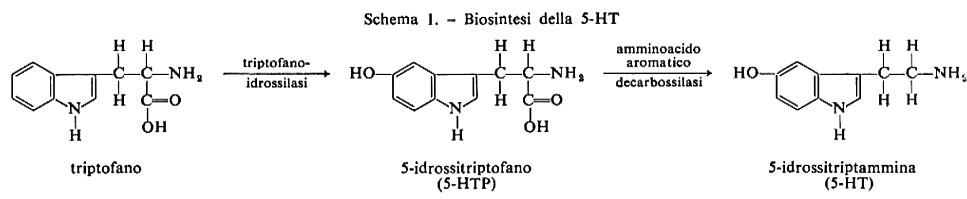
È regola generale per tutti i sistemi monoamminergici che le molecole del mediatore siano sintetizzate dentro il soma del neurone da enzimi specifici, che siano trasportate per flusso assonico alle terminazioni del cilindrasse e che quivi siano immagazzinate, per essere liberate al momento opportuno e messe in condizione di legarsi con molecole recettrici. (Il neurone è costituito da molti dendriti, da un soma e da un assone o cilindrasse. Il soma è la parte del citoplasma che circonda il nucleo: per questa ragione si parla anche di pericari delle cellule nervose). Mentre gli enzimi e le vescicole che immagazzinano le monoammine sono sempre prodotti all'interno del soma, sembra che le terminazioni del cilindrasse possano captare le molecole precorritrici del mediatore e che la sintesi di questo avvenga anche (e forse principalmente) in esse. In ogni modo, dopo la distruzione di pericari monoamminergici, la fluorescenza specifica delle rispettive terminazioni assoniche diminuisce, per scomparire entro 8-10 giorni. Ovviamente i neuroni del tronco dell'encefalo che emettono lunghi cilindrassi ascendenti possono agire su membrane postsinaptiche di cellule nervose localizzate nel cervello; pertanto mediatori sintetizzati nel tronco dell'encefalo e liberati da impulsi nervosi che quivi prendono origine possono combinarsi con appropriate molecole recettrici situate sulle membrane postsinaptiche di neuroni del cervello, esercitando così la loro azione specifica a livello cerebrale. Queste terminazioni di vie ascendenti dal tronco dell'encefalo saranno, ovviamente, degenerate nel cerveau isolé cronico e la stessa situazione si verificherà per le terminazioni di vie cerebrali discendenti, nel preparato decerebrato cronico. Pertanto la tendenza all'attività ritmica che si osserva in questo preparato cronico sarà dovuta a neuroni i cui pericari si trovano, rispettivamente, nel cervello o nel tronco dell'encefalo. Per semplicità solo due sistemi ascendenti verranno presi in esame: i nuclei del rafe, uno dei sistemi deattivanti della parte caudale del tronco dell'encefalo, e il sistema reticolare ascendente, che è di natura attivante. (V. anche sinapsi; sistema nervoso autonomo; neurosecrezione).
5-Idrossitriptammina. - I metodi di fluorescenza istochimica mostrano che la maggior parte dei neuroni il cui soma contiene 5-HT sono localizzati nei ‛nuclei del rafe'. Queste osservazioni suggeriscono che l'azione deattivante, che abbiamo visto nascere in questi nuclei del tronco dell'encefalo, è legata alla liberazione di un mediatore chimico identificato come 5-HT.
La biosintesi della 5-HT cerebrale comincia con l'assunzione dal plasma sanguigno dell'amminoacido triptofano; la prima trasformazione biochimica consiste nella 5-idrossilazione del triptofano a opera della triptofanoidrossilasi. L'amminoacido 5-idrossitriptofano (5-HTP) viene decarbossilato dando origine alla 5-idrossitriptammina (5-HT), una indolammina che è probabilmente il mediatore chimico. Lo schema 1 dà le tappe della biosintesi della 5-HT.
Dopo iniezione di p-clorofenilalanina, che blocca la triptofanoidrossilasi, si ha nel cervello una diminuzione della 5-HT. Una sola iniezione di 400 mg/kg di questa sostanza, nel gatto, è senza alcun effetto sul ciclo sonno-veglia durante le prime 18-24 ore. Ma questo periodo iniziale è seguito dalla diminuzione del sonno e quindi dalla completa insonnia. Il sonno riappare solo dopo 40 ore, ma il normale ciclo sonno-veglia ritorna solo dopo 2.000 ore (v. Jouvet, 1972).
Il fatto che l'insonnia cominci con un ritardo di circa un giorno fa pensare che la p-clorofenilalanina non abbia un'azione diretta sul ciclo sonno-veglia, che verrebbe alterato solo come conseguenza dell'esaurimento delle riserve di 5-HT nel sistema deattivante del rafe. Ulteriori prove del significato fisiologico di queste osservazioni sono date dalla correlazione, nel ratto, fra la diminuzione del sonno e il calo della 5-HT cerebrale e dall'osservazione che l'insonnia viene sostituita, nel gatto, da un normale ciclo sonno-veglia se l'azione di blocco è aggirata con iniezione di 5-HTP (v. Jouvet, 1972).
Noradrenalina. - I metodi di fluorescenza istochimica dimostrano che i pericari del tronco dell'encefalo che contengono NA sono in massima parte localizzati nel tegmento del mesencefalo, ma anche nel ponte e nel bulbo, e cioè nei luoghi d'origine del sistema reticolare ascendente. Questa osservazione fa pensare che la liberazione di un mediatore chimico identificato come NA sia legata all'azione attivante che il tronco dell'encefalo esercita sul cervello.
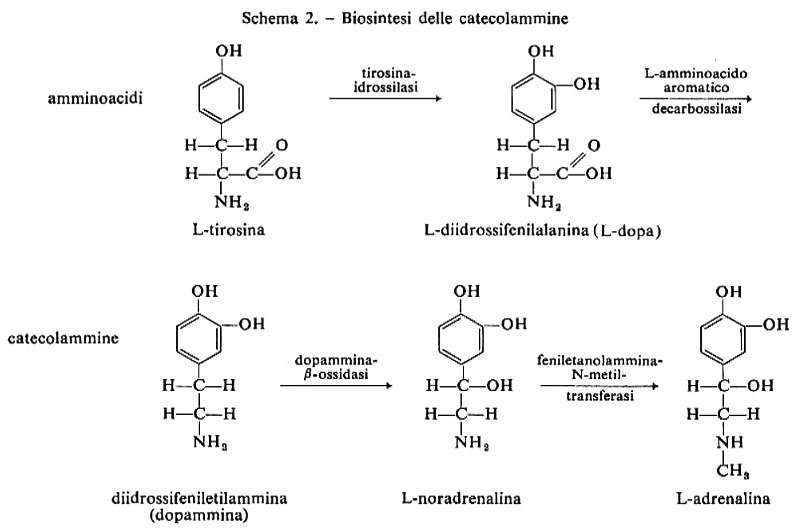
La biosintesi della NA comincia con l'assunzione dell'amminoacido tirosina dal sangue circolante. La prima trasformazione biochimica è rappresentata dalla metaidrossilazione a L-diidrossifenilalanina (L-dopa) per opera della tirosinaidrossilasi. La decarbossilazione di questo amminoacido, per opera della dopadecarbossilasi, porta alla formazione della prima catecolammina, la 3,4-diidrossifeniletilammina o dopammina (DA), che è il mediatore chimico per alcuni neuroni. La DA viene trasformata in un'altra catecolammina, la L-noradrenalina, da una dopammina-β-ossidasi, che è localizzata nelle vescichette sinaptiche in cui s'immagazzina la NA. Un'ulteriore trasformazione, che per altro avviene soprattutto nelle cellule secretorie della midolla surrenale, è la conversione nella catecolammina terminale, la L-adrenalina, per opera di una feniletanolammina-N-metiltransferasi Lo schema 2 dà le tappe della biosintesi della NA.
c) La dottrina monoamminergica del ciclo sonno-veglia
Questi che abbiamo citati sono solo pochi esempi dei risultati che possono essere ottenuti con l'approccio neurochimico. In complesso la natura 5-idrossitriptamminergica del sistema deattivante dei nuclei del rafe e la natura noradrenergica del sistema attivante reticolare sembrano assodate. La ‛teoria monoamminergica' di M. Jouvet (v., 1972) parte dal presupposto che l'alternanza fra veglia e sonno sincronizzato sia dovuta a un ciclo di attività che nasce nel tronco dell'encefalo, essendo legato al risultato di un conflitto fra i due sistemi monoamminergici antagonisti. L'attività dei neuroni cerebrali modificati dalla veglia e dal sonno si limiterebbe a registrare l'esito d'una lotta che è decisa a livello del tronco dell'encefalo.
6. Registrazioni elettrofisiologiche da singoli neuroni del tronco dell'encefalo
I metodi di lesione e di stimolazione e le indagini neurochimiche portano alla conclusione che il ciclo veglia sonno e le fasi stesse del sonno, sincronizzata e desincronizzata, sono legati al ritmico alternarsi dell'attività di strutture nervose localizzate nel tronco dell'encefalo e nel diencefalo. In questi ultimi anni i progressi della tecnica hanno permesso di registrare la scarica di singoli neuroni nell'animale senza narcosi e libero nei suoi movimenti e quindi hanno consentito di mettere in rapporto la scarica unitaria con il comportamento, con l'elettroencefalogramma e con l'elettromiogramma durante la veglia e le fasi sincronizzata e desincronizzata del sonno. Non tutte le strutture nervose il cui significato funzionale è stato chiarito con i metodi di lesione o di stimolazione sono state studiate a livello cellulare, ma per alcune di esse i progressi conseguiti sono tali da giustificare un'esposizione a parte dei risultati di questo nuovo metodo d'indagine.
Abbiamo visto che il ciclo veglia-sonno è legato all'attività di strutture antagoniste, dette attivanti e deattivanti. Recentemente M. Steriade e altri (v., 1980) sono riusciti a registrare le scariche di singoli neuroni del tegmento mesencefalico e a dimostrarne con metodi fisiologici l'appartenenza al sistema reticolare ascendente. La scarica di queste cellule nervose si accelera infatti quando si passa dal sonno sincronizzato alla veglia, mentre diminuisce poco prima dell'inizio del sonno sincronizzato. Il passo successivo sarà verosimilmente quello di analizzare a livello cellulare le strutture deattivanti localizzate nelle regioni basali del cervello anteriore, nel rafe e nella regione del tratto solitario.
Le nostre conoscenze sono molto più soddisfacenti per ciò che riguarda lo studio a livello cellulare delle strutture pontine che assicurano l'alternarsi delle fasi sincronizzata e desincronizzata del sonno.
Jouvet (v., 1962) aveva dimostrato che la distruzione bilaterale e simmetrica della parte rostrale del nucleus reticulapis pontis caudalis e della parte caudale del nucleus reticularis pontis oralis sopprimeva il sonno paradosso. I neuroni ‛generatori' di questa fase del sonno si trovano dunque nel tegmento pontino. Hobson e altri (v., 1975), con ricerche sul gatto, hanno dimostrato che nel nucleo tegmentale gigantocellulare del ponte esistono neuroni la cui frequenza di scarica aumenta selettivamente durante il sonno desincronizzato. Altre ricerche eseguite da Pivik e altri (v., 1977) hanno dimostrato che i movimenti rapidi oculari che caratterizzano il sonno desincronizzato sono preceduti, e verosimilmente causati, da uno spiccato aumento selettivo della scarica degli stessi neuroni. Le cellule nervose del nucleo tegmentale gigantocellulare del ponte sono dunque i generatori delle manifestazioni toniche e fasiche del sonno desincronizzato. Questo non significa, naturalmente, che i predetti neuroni non svolgano la loro azione anche durante la veglia.
Il problema che ora si pone è quello del meccanismo fisiologico che dà il via alla scarica dei neuroni del nucleo reticolare pontino e quindi alla comparsa delle manifestazioni toniche e fasiche del sonno desincronizzato. Hobson e altri (v., 1975) hanno trovato, nel locus coeruleus e nel nucleus subcoeruleus, neuroni che hanno un comportamento reciproco a quello del nucleo gigantocellulare. Durante il passaggio dal sonno sincronizzato a quello desincronizzato la loro frequenza di scarica si riduce, e questo avviene proprio mentre la scarica dei neuroni del nucleo gigantocellulare si accelera. Queste osservazioni suggeriscono l'ipotesi che per queste ultime cellule nervose si verifichi in quel momento la liberazione da un freno inibitore.
Esperimenti recenti fanno ritenere che i neuroni del nucleo gigantocellulare siano colinergici, mentre quelli del locus coeruleus e del nucleus subcoeruleus sarebbero noradrenergici. T. T. Amatruda e altri (v., 1975) hanno dimostrato infatti che l'iniezione locale nelle regioni del nucleo gigantocellulare di un agente colinergico a lunga durata, il carbacolo, aumenta fortemente le manifestazioni del sonno paradosso del gatto normale, quali l'atonia muscolare e la desincronizzazione del tracciato elettroencefalografico. Se il gatto è stato decerebrato si possono avere solo le manifestazioni del sonno paradosso legate a scariche discendenti dei neuroni generatori pontini. Hoshino e Pompeiano (v., 1976) hanno dimostrato che l'iniezione endovenosa di un farmaco anticolinesterasico, quale l'eserina, fa comparire nel gatto decerebrato acuto fenomeni cataplessici caratterizzati da atonia, sempre preceduti da un aumento rilevante delle scariche dei neuroni del nucleo gigantocellulare. Quando la rigidità ricompare, spontaneamente o per effetto di stimoli cutanei, essa si accompagna a una marcata diminuzione della frequenza di scarica di questi neuroni. Pompeiano e Hoshino (v., 1976) hanno anche osservato che neuroni del nucleo del locus coeruleus si comportano in modo reciproco, ossia riducono la loro frequenza di scarica durante gli episodi cataplessici: quando la rigidità ricompate, la scarica dei neuroni del locus coeruleus si intensifica di nuovo. Questi esperimenti e altri ricordati nei lavori originali fanno ritenere che i neuroni del nucleo gigantocellulare siano colinocettivi e colinergici e scatenino con la loro scarica l'insorgenza delle manifestazioni del sonno paradosso, e che invece i neuroni del locus coeruleus, colinocettivi ma noradrenergici, inibiscano periodicamente i neuroni generatori del nucleo gigantocellulare.
7. Il significato funzionale del sonno
Fino a pochi anni or sono il sonno veniva considerato come un fenomeno omogeneo. Si diceva che compito del sonno era di ristabilire, con meccanismi che potevano essere genericamente definiti ‛di ristoro', le condizioni che si avevano nel cervello all'inizio del precedente periodo di veglia. La teoria del ristoro presupponeva che durante la veglia si verificasse un logoramento nei centri nervosi (v. Sherrington, 1946, p. 252). Si trattava forse di alterazioni limitate a un gruppo o ad alcuni gruppi di neuroni, ma la loro caratteristica fondamentale era che esse richiedevano un lungo tempo (alcune ore) per essere riparate.
Sappiamo ora che vi sono due periodi di sonno, profondamente diversi nelle loro manifestazioni e nel loro significato funzionale. Questo diverso significato funzionale è dimostrato dal fatto che nella notte successiva a quella in cui un soggetto era stato svegliato ogniqualvolta il tracciato elettroencefalografico si desincronizzava, e quindi dopo una privazione selettiva del sonno paradosso, la percentuale di tempo trascorsa nel sonno desincronizzato si accresceva. L'aumento compensatorio del sonno paradosso dimostra che questa fase di sonno ha un significato funzionale suo proprio. Le azioni di ristoro svolte durante il sonno desincronizzato non possono essere sostituite da meccanismi fisiologici che si svolgono in altre fasi del sonno.
Esamineremo separatamente le teorie del ristoro ipnico e in seguito il significato del sonno desincronizzato.
a) Le teorie del ristoro ipnico
Pressoché tutte le attività del cervello e del corpo sono influenzate dal ciclo sonno-veglia. Tuttavia alcune manifestazioni corporee del sonno sono chiaramente epifenomeni, non particolarmente interessanti per ciò che riguarda il problema fisiologico principale. Il rilassamento muscolare è assente nel treno anteriore e nel collo del bestiame bovino. La chiusura degli occhi è estremamente rara nel bestiame bovino, e d'altra parte brevi manifestazioni di lagoftalmo sono state descritte persino nell'uomo. L'abolizione dei riflessi di raddrizzamento, infine, non compare durante il sonno degli Uccelli, come è dimostrato dal fatto che questi animali possono dormire appollaiati su un ramo senza mai cadere. Possiamo terminare con una semplice considerazione. Quando giaciamo insonni su un letto, sappiamo perfettamente che scopo principale del sonno non è dare un periodo di riposo al nostro corpo.
Scopo del sonno è il ristoro del sistema nervoso centrale. Ma occorre fare una distinzione fra i neuroni la cui attività è modificata come conseguenza dello stato di sonno e quelli il cui ristoro è il principale compito del sonno. Il sonno influenza fortemente, per esempio, il respiro, il ritmo cardiaco, il tono dei muscoli scheletrici; ma tutti sappiamo che il ristoro del centro respiratorio o dei neuroni cardioregolatori o del cervelletto o del midollo spinale non rappresenta lo scopo principale del sonno. In realtà sintomo principale del sonno è la mancanza di coscienza o la sua grave alterazione. Come giustamente ha messo in evidenza Evarts (v., Relation of cell..., 1965, p. 89) ‟le vivide esperienze dei sogni non lasciano tracce permanenti nella memoria, a meno che il soggetto non sia svegliato durante il sogno o poco dopo. Pertanto le scariche neuroniche necessarie per la percezione immediata permangono durante il sogno, mentre invece cessano i processi necessari al consolidamento delle tracce della memoria". E, infine, è un fatto ben conosciuto che i sintomi prodotti dalla privazione del sonno appartengono alla sfera psichica (v. Kleitman, 1963, pp. 215-229).
È stata operata una distinzione fra processi rapidi e lenti di ristoro delle attività nervose. I processi di ristoro rapidi sono in rapporto alla conduzione e alla trasmissione sinaptica degli impulsi nervosi, e quindi, essenzialmente, alla permeabilità agli ioni della membrana (v. elettrofisiologia; v. sinapsi). Questo è il compito principale del ristoro nel cervelletto, ad esempio, o nel centro respiratorio bulbare o nel midollo spinale. I processi di ristoro lenti, invece, occupano una parte molto maggiore della vita, almeno in molti Mammiferi e negli Uccelli: quasi un terzo della vita umana viene speso nel sonno. Un andamento nel tempo tanto differente deve essere in rapporto a meccanismi del tutto differenti di ristoro. È stata prospettata l'ipotesi che il sonno non riguardi tutto il cervello, e neppure tutta la neocorteccia (la parte filogeneticamente nuova della corteccia cerebrale, che contiene aree di proiezione - motorie e sensitive - e di associazione), ma solo quei neuroni e quelle sinapsi, e forse quelle cellule gliali, che durante la veglia sono responsabili delle funzioni cerebrali a cui è dovuto il comportamento cosciente, o sono in qualche modo in rapporto con esse (v. Moruzzi, 1972, p. 121). Cambiamenti di natura plastica, probabilmente macromolecolare, è verosimile avvengano durante la veglia nelle strutture nervose, in rapporto con i processi di apprendimento.
I. Feinberg ed E. V. Evarts (v., 1969, p. 338) hanno giustamente rilevato che l'idea di una funzione positiva del sonno ‟implica la nozione di complementarità, e cioè che la quantità e l'intensità del sonno debbano essere in rapporto all'intensità di quei processi di veglia che richiedono, appunto, un'azione di completamento da svolgersi durante il sonno". Tra le funzioni positive che riconoscono al sonno, anche in base ai loro esperimenti di psicofisiologia nell'uomo, essi citano non solo il consolidamento degli engrammi, ma anche la scomparsa di certe categorie di memorie. Questa è indubbiamente un'ipotesi stimolante, se si tiene presente l'importanza ben conosciuta dell'oblio selettivo dei ricordi non significativi. È chiaro che la parola ‛ristoro' ha significati differenti per i muscoli scheletrici e per il cervello. Non può essere scopo del sonno il ristabilire o il restaurare esattamente le stesse condizioni che erano presenti all'inizio del precedente periodo di veglia: ciò impedirebbe l'apprendimento. L'ottenere condizioni costanti mediante il riposo può essere possibile per le fibre muscolari, non per il cervello, anche qualora si prescinda dal problema della possibilità di periodi caratterizzati dall'assenza di attività specifica nel cervello, cosa invece che accade nel caso delle fibre di un muscolo scheletrico completamente rilassato.
D'altra parte la durata del sonno non ci dà una misura soddisfacente dell'ammontare dei processi neurochimici che si svolgono in esso. Ciò è dimostrato dal fatto che la sindrome imponente prodotta in un uomo da un'insonnia prolungata per ben 200 ore può scomparire pressoché completamente solo dopo 13 ore di sonno (v. Luce, 1965, p. 20). Non v'è dubbio, inoltre, che molti animali, quali il gatto domestico, dormano in eccesso rispetto ai loro bisogni. Secondo Jouvet (v., 1972) questa mancanza di una relazione lineare fra le durate della veglia e del sonno riguarda solo la fase caratterizzata da EEG sincronizzato, che sarebbe funzione del log della durata della veglia. La durata del sonno paradosso, invece, è direttamente proporzionale alla durata della veglia, nella misura in cui la veglia implica deprivazione di sonno paradosso. Abbiamo detto sopra che la privazione selettiva della fase paradossale è seguita da un debito specifico per questa fase del sonno, che può essere pagato solo con un aumento della durata del tempo trascorso nei periodi di sonno desincronizzato durante la notte successiva.
Durante il periodo di ristoro il muscolo scheletrico è rilassato; esso quindi è inattivo, almeno per ciò che riguarda la sua funzione specifica, la contrazione. Per analogia si potrebbe pensare che durante il sonno i neuroni della corteccia cerebrale, dediti alle complesse operazioni metaboliche necessarie al ristoro, siano essi pure fisiologicamente inattivi e quindi non scarichino impulsi. Questa era effettivamente la concezione di Pavlov, che presupponeva un silenzio generalizzato della corteccia cerebrale durante il sonno. L'obliterazione della coscienza, che caratterizza il sonno, poteva spiegarsi con l'inattivazione funzionale del mantello neocorticale. La registrazione elettrofisiologica di singoli neuroni della corteccia cerebrale ha reso insostenibile questa ipotesi. Sappiamo con certezza che i neuroni corticali sono spontaneamente attivi durante il sonno, anche se la loro attività appare nettamente diversa da quella che si osserva in condizioni di veglia.
b) Significato del sonno desincronizzato
Evarts è stato il primo a registrare sistematicamente durante le varie fasi della veglia e del sonno le scariche dei neuroni della corteccia visiva e anche di quelli della corteccia motoria che danno origine al fascio piramidale (v. Evarts, 1962, 1964 e 1965). Si sapeva da tempo che i neuroni motori corticospinali sono tonicamente attivi durante la narcosi barbiturica, e quindi in assenza di movimenti (v. Adrian e Moruzzi, 1939); la scarica d'impulsi piramidali diventa sopraliminare per i motoneuroni spinali solo quando raggiunge una frequenza critica e solo allora appaiono i movimenti. Nella scimmia senza narcosi, Evarts (v., 1964) ha dimostrato che i neuroni del fascio piramidale non sono affatto inattivi durante le fasi sincronizzata e desincronizzata del sonno, ma semplicemente si scaricano in modo diverso rispetto alla veglia rilassata. Le modalità di scarica che si osservano in questi esperimenti di Evarts non hanno però validità generale, a differenza di quanto si riteneva un tempo. Lavori successivi di G. Berlucchi (v., 1965) e di L. M. Mukhametov e altri (v., 1970), e infine diverse ricerche ricordate da Steriade (v., 1978) hanno dimostrato che per altri tipi di neuroni le modalità di scarica durante le fasi del sonno sono diverse. V'è però un punto fondamentale su cui tutti sono d'accordo: una scarica cellulifuga è sempre presente nel sonno in tutti i neuroni corticali studiati in queste condizioni sperimentali.
Resta il problema di vedere come il ristoro possa intervenire se i neuroni che dovrebbero beneficiarne sono sempre attivi. In un tentativo di risolvere queste difficoltà, Evarts (v., 1964) e Moruzzi (v., 1966) avevano suggerito che questa incessante attività fosse limitata ai soli neuroni del I tipo, a cilindrasse lungo. I neuroni del Il tipo o interneuroni, che assicurano i circuiti nervosi all'interno della sostanza grigia, erano sfuggiti fino allora allo studio microelettrodico, almeno nelle condizioni sperimentali. Queste cellule nervose avrebbero potuto essere silenti durante il sonno. Recentemente Steriade e collaboratori sono riusciti a identificare entrambi i tipi di neuroni nella corteccia cerebrale e a registrarne separatamente l'attività nell'animale senza narcosi, libero nei suoi movimenti (v. Steriade, 1978). Essi hanno visto che la scarica dei neuroni corticali del Il tipo è addirittura maggiore nel sonno sincronizzato rispetto alla veglia. Questi risultati rimettono in discussione tutte le teorie sui meccanismi nervosi che si svolgono nella corteccia cerebrale durante il sonno. È certo, in ogni caso, che durante il sonno le cellule nervose corticali non riposano, ma presentano un'organizzazione diversa della loro attività.
Sembra dunque chiaro che durante il sonno non si possano avere fenomeni di ristoro secondo lo schema classico del ristoro del muscolo scheletrico. Almeno per il sonno desincronizzato sembra addirittura che esso sia necessario per permettere attività del sistema nervoso che nulla hanno a che fare con i fenomeni di ristoro, nel senso che in fisiologia si dà a questa parola. Secondo Jouvet (v., 1978) il sonno desincronizzato, con i suoi potenti meccanismi inibitori che isolano il cervello dalla periferia motoria e forse (in parte almeno) dalla periferia sensitiva, potrebbe fornire un quadro ideale per una programmazione genetica e periodica del cervello. J. P. Sastre (v., 1978) ha continuato le ricerche di Jouvet e Delorme (v., 1965) sugli effetti di lesioni bilaterali delle strutture nervose responsabili della comparsa delle manifestazioni inibitrici negli episodi di sonno desincronizzato. In queste condizioni vengono alla luce i comportamenti motori corrispondenti ai sogni che, come è noto, compaiono nell'uomo durante il sonno desincronizzato. L'animale sembra allora cercare un oggetto immaginario o combattere un nemico immaginario. Il cervello eccita dunque, direttamente o indirettamente, le sinapsi, ‛provando' in tal modo i circuiti sensitivi e motori messi in gioco (durante la veglia) nei comportamenti innati. La soppressione dei fenomeni inibitori porta alla luce gli effetti motori della scarica programmata che agiscono sui motoneuroni spinali. In condizioni normali solo frammenti di attività motoria sfuggono all'inibizione e possono venire alla luce. Il resto dell'attività onirica dei centri motori è bloccato dalle scariche inibitrici che discendono al midollo spinale dal tronco dell'encefalo. Il sogno si riduce così, in gran parte almeno, ad allucinazioni visive, con scarse manifestazioni motorie.
8. Sonno e comportamento istintivo
La regolazione del ciclo sonno-veglia è dovuta a meccanismi nervosi innati, che possono essere modificati sia da impulsi incondizionati sia dal condizionamento (v. condizionamento, meccanismo del).
Molti etologi (v. Holzapfel, 1940; v. Tinbergen, 1952 e 19552; v. Hediger, 1959 e 1969) hanno rilevato che il comportamento degli animali nel periodo che precede il sonno ricorda la fase appetitiva che preannunzia ogni tipo di comportamento istintivo.
Ai fini del presente articolo possiamo trascurare qui il problema delle componenti acquisite del comportamento animale o quello della differenza fra attività istintive e attività puramente riflesse (v. istinto).
Secondo Tinbergen (v., 1952) il sonno potrebbe essere considerato come la fase finale, o atto consummatorio, di un comportamento istintivo. Come ha osservato W. Craig (v., 1918), quando il comportamento istintivo è stato completato con un atto consummatorio, l'animale appare ‛sazio' o ‛soddisfatto'.
Questi concetti etologici sono stati sviluppati da un punto di vista neurofisiologico. È stato detto (v. Moruzzi, 1969) che il sonno, con le sue fasi sincronizzata e desincronizzata, può essere piuttosto paragonato a una catena di atti consummatori. Questa catena di risposte inizierebbe sempre con la fase caratterizzata da sincronizzazione dell'EEG, non appena l'attività tonica del sistema reticolare ascendente cade al di sotto del livello critico.
In tutti i tipi di comportamento istintivo che si osservano durante la veglia, la fase consummatoria è caratterizzata da movimenti stereotipati; essi implicano l'esistenza di modelli neuronici di scarica stereotipata che regolano, nello spazio e nel tempo, l'attività dei motoneuroni corrispondenti. Scariche stereotipate di neuroni cerebrali caratterizzano le due fasi di sonno, peraltro in modo del tutto indipendente dagli effetti periferici sui muscoli.
Dal punto di vista del tema di quest'articolo abbiamo la scelta fra due ipotesi: 1) vi potrebbe essere un rapporto di causalità tra la scarica stereotipata di neuroni corticali e il compito fondamentale del sonno, il ristoro del cervello. Abbiamo definito questo modello di comportamento neuronico un tipo introverso di comportamento, per mettere l'accento sulla differenza fondamentale rispetto al comportamento di veglia. Nell'animale sveglio lo scopo principale della scarica dei neuroni è la produzione di determinati movimenti, e cioè di effetti periferici di cui è facile comprendere il significato rispetto al mondo che circonda l'animale. I modelli di scarica stereotipata che si hanno nel sonno sarebbero necessari, per ragioni ancora sconosciute, allo scopo di avere il ristoro ipnico, o di assicurare la programmazione periodica dei circuiti nervosi, mentre i loro effetti periferici, quali le scosse cloniche della fase paradossa, sarebbero solo epifenomeni; 2) vi potrebbe anche essere un'associazione fra manifestazioni del sonno, di natura istintiva, e i processi neurochimici e neurofisiologici di ristoro, che di per sé non sarebbero istintivi. La situazione sarebbe analoga all'associazione fra il comportamento istintivo che porta all'assunzione di acqua e i meccanismi neuroendocrini, puramente fisiologici, che regolano la diuresi. Due tipi diversi di attività, un comportamento istintivo e meccanismi neurochimici o neuroendocrini, potrebbero convergere nella produzione dello stesso risultato: rispettivamente, il ristoro ipnico dei neuroni cerebrali e la costanza della pressione osmotica nel mezzo interno.
Indubbiamente il sonno è una forma unica di regolazione fisiologica, perché implica la perdita temporanea della coscienza o un suo forte indebolimento (se si considerano i sogni). Tutte le forme conosciute di comportamento istintivo scompaiono durante il sonno, e quindi gli animali devono adattare il ciclo sonno-veglia alle loro esigenze per ciò che riguarda la difesa o l'approvvigionamento di cibo o acqua. Di conseguenza ogni animale deve pagare un forte scotto allo scopo di beneficiare degli effetti del sonno, come appare chiaro a chi consideri cosa può apportare la perdita della coscienza negli erbivori allo stato selvaggio. Pertanto la regolazione dei periodi dedicati al sonno ha un grande valore per la sopravvivenza, e non è sorprendente che tale controllo diventi parte di un modello di attività istintiva. Esso è infatti il risultato di un conflitto fra gli istinti della veglia, da una parte, e il comportamento istintivo del sonno dall'altra. Le differenze tanto marcate fra i vari Mammiferi, per ciò che riguarda il ritmo sonno-veglia, possono essere spiegate in questo modo. È improbabile che il fatto che in molti erbivori si osservano rari e brevi episodi di sonno, mentre il gatto può dedicarvi il 72% del suo tempo, possa essere spiegato da differenze nell'esigenza di ristoro ipnico (v. Moruzzi, 1972, per la letteratura etologica).
Queste considerazioni portano a una concezione più ampia del significato fisiologico del controllo del cervello da parte del tronco dell'encefalo (ibid.). I risultati delle ricerche sul sonno, in grande sviluppo nell'ultimo quarto del secolo, potrebbero dare l'impressione che compito principale dei sistemi ascendenti del tronco dell'encefalo sia il controllo del ciclo sonno-veglia. Ma in realtà appare improbabile che siano richieste tante strutture nervose per un compito comparativamente così semplice.
9. Interpretazioni neurofisiologiche nella patologia del sonno
Le principali sindromi della patologia del sonno possono ora essere reinterpretate in base alle nostre conoscenze di fisiologia reticolare.
a) Narcolessia e cataplessia
La narcolessia è una malattia caratterizzata da un bisogno irresistibile di dormire. Ricerche elettroencefalografiche (v. Dement e Rechtschaffen, 1968) hanno dimostrato che si tratta di casi di sonno paradosso, non preannunciati da una fase sincronizzata. La cataplessia, invece, è caratterizzata da episodi brevi (da 3 a 5 minuti) di atonia completa, senza perdita di coscienza. Una scarica improvvisa del centro pontino responsabile degli episodi paradossi potrebbe spiegare entrambi i fenomeni. Durante la cataplessia, tuttavia, sarebbe presente, per ragioni ignote, solo l'influenza discendente inibitrice esercitata dal tronco dell'encefalo sul midollo spinale (v. Pompeiano, 1965 e The neurophysiological..., 1967). La sensazione di essere paralizzati durante gli incubi notturni e la paralisi da sonno che si osserva al momento del risveglio potrebbero essere pure in rapporto con episodi di sonno paradosso, o con una scarica postuma delle salve d'impulsi discendenti inibitori di solito legate a questi episodi. L'ipotesi è suggerita dall'arresto dei movimenti prodotti dalla stimolazione elettrica del fascio piramidale, fenomeno che si osserva costantemente nel gatto durante gli episodi di sonno desincronizzato (v. Marchiafava e Pompeiano, 1964).
b) Letargo della malattia del sonno e coma
Il letargo che si osserva nella malattia del sonno (encefalite letargica epidemica) differisce nettamente dal coma, perché il paziente può essere facilmente risvegliato, e allora appare ben orientato rispetto all'ambiente. La sindrome è stata riprodotta nella scimmia con lesioni ipotalamiche (v. Ranson, 1939).
Nei casi clinici questo risultato è probabilmente dovuto alla distruzione parziale del tegmento del mesencefalo, che determina un minore afflusso di impulsi reticolari attivanti combinato alla lesione delle strutture ipotalamiche in rapporto con la veglia. Il coma è uno stato caratterizzato dalla completa perdita di coscienza, che non può essere interrotto neppure con le più forti stimolazioni sensitive o sensoriali (v. coscienza: Alterazioni della coscienza). Esso compare ogni qualvolta il sistema reticolare ascendente è interrotto, a livello del mesencefalo, da traumi o tumori, o è inattivato funzionalmente da dosi narcotiche di barbiturici. Questo stato è riprodotto sperimentalmente nel preparato cerveau isolé (v. Bremer, 1935, 1937 e 1938). H. Orthner (v., 1969) ha dimostrato che le sindromi sperimentali sia del cerveau isolé sia del preparato pretrigeminale sono riprodotte nella patologia umana. (Per la patologia del sonno v. Kleitman, 1963; v. Orthner, 1969; v. Otter, 1970; per la farmacologia del sonno, v. Holm, 1970).
c) Le insonnie
Gli sviluppi recenti della neurochimica hanno gettato luce sulla patologia e sulla farmacologia delle insonnie. Di notevole interesse è la marcata insonnia osservata da Jouvet e collaboratori in seguito alla distruzione dei nuclei del rafe (v. cap. 3) o al blocco farmacologico della biosintesi di 5-HT (v. cap. 5). Oltre che dalla depressione del sistema deattivante indolamminergico, l'insonnia può risultare anche dall'iperattività primaria dei neuroni attivanti, catecolamminergici. Un esempio farmacologico di questo tipo di squilibrio del ciclo sonno-veglia è rappresentato dall'azione dell'anfetammina. Il lettore viene rinviato alla rassegna critica di Jouvet (v., 1972) per la discussione e per l'esposizione di molti altri effetti neurochimici e neurofarmacologici sul sonno e sulla veglia.
10. Stati simili al sonno
Cercheremo ora di riassumere brevemente i principali dati fisiologici sull'ipnosi animale e sull'ibernazione. Entrambi questi fenomeni sono nettamente differenti dal sonno fisiologico. Tuttavia in essi potrebbero intervenire alcuni aspetti dei meccanismi neurofisiologici e neurochimici interessati nel ciclo sonno-veglia. In particolare vedremo che il sistema inibitore discendente, attivo durante la fase paradossale del sonno fisiologico, può essere selettivamente messo in azione durante l'ipnosi animale, mentre l'ibernazione potrebbe essere messa in rapporto all'arresto del turnover di catecolammine nel sistema reticolare ascendente. Pertanto una breve analisi di tali problemi può essere giustificata alla fine di questo articolo.
a) L'ipnosi animale
L' ipnosi animale è caratterizzata dall'assenza di movimenti e dall'abolizione dei riflessi di raddrizzamento. Viene facilmente ottenuta ponendo sul dorso conigli, galline o altri animali che, immobilizzati per un tempo molto breve, rimangono senza muoversi nella nuova, inusitata posizione. La vecchia letteratura su questo argomento - e anche quella sull'ipnosi umana, tanto meno soddisfacente dal punto di vista scientifico - è data da Kleitman (v., 1963), mentre la letteratura più moderna si trova in un lavoro di G. Carli (v., 1971).
Risultati contraddittori sono stati ottenuti con studi elettroencefalografici sui conigli (v. Gerebtzoff, 1941; v. Svorad, 1957), ma sembra comunque assai improbabile che l'ipnosi animale possa essere paragonata a una forma qualsiasi di sonno (v. Klemm, 1966). Per ciò che riguarda il midollo spinale, invece, è stato dimostrato nel coniglio che i comuni riflessi sono molto depressi e che questi effetti non trovano rispondenza nel tracciato dell'EEG (v. Carli, 1969). Si è visto inoltre che l'ipnosi animale può ancora essere ottenuta, sempre nel coniglio, dopo decerebrazione precollicolare (v. Carli, 1971).
P. Buser e G. Viala (v., 1968) hanno avanzato l'ipotesi che l'arresto dei movimenti sia dovuto all'influenza inibitoria del sistema reticolo-spinale di Magoun, che probabilmente è anche responsabile dell'inibizione dei riflessi spinali e dei movimenti prodotti dalla stimolazione del fascio piramidale (v. Pompeiano, 1965, 1966 e The neurophysiological..., 1967). Pertanto meccanismi inibitori discendenti interessati al ciclo sonno-veglia possono essere messi selettivamente in attività durante l'ipnosi animale. Questo fenomeno è manifestamente in rapporto con i casi clinici di cataplessia (v. cap. 9, § a) e con gli episodi cataplessici del gatto decerebrato (v. cap. 3, § b).
b) L'ibernazione dei Mammiferi
L' ibernazione è un fenomeno stagionale che può essere osservato durante l'inverno in alcuni mammiferi, quali lo scoiattolo e la marmotta. L'argomento è stato oggetto di una rassegna critica di Sàtinoff (v., 1970).
Dopo un periodo di preparazione che si svolge nella tarda estate e nel primo autunno, durante il quale aumenta l'ingestione di cibo e si accresce il peso dell'animale, la temperatura corporea dello scoiattolo comincia a cadere e può giungere a valori assai bassi, fino a 4-5 °C. L'instaurarsi dello stato di letargo è in rapporto a questa caduta della temperatura. Dopo circa due settimane d'ibernazione l'animale si sveglia completamente, pur nell'assenza di qualsiasi cambiamento apprezzabile nell'ambiente, mentre la temperatura sale agli abituali livelli omeotermici di 37 °C. Il livello di vigilanza è eccellente durante questi periodi di veglia, che possono durare anche qualche giorno all'inizio o alla fine dell'ibernazione, mentre a mezzo inverno durano solo poche ore. In questi periodi l'animale orina e può anche mangiare, bere o defecare, per poi cadere nuovamente nello stato d'ibernazione. Solo dall'inizio della primavera all'autunno l'animale ritorna al normale ciclo sonno-veglia.
L'ibernazione non può essere considerata come uno stato di coma, giacché gli animali possono risvegliarsi, quando vengono stimolati, e rispondere a stimoli acustici voltando la testa in direzione del suono. Si tratta tuttavia di un tipo differente di sonno, almeno per ciò che riguarda le fasi sincronizzata e paradossale che si osservano nello scoiattolo non ibernante. Gli studi elettroencefalografici hanno dato risultati contrastanti, che possono anche essere spiegati con differenze di specie. È certo comunque che quando lo scoiattolo raggiunge lo stadio finale dell'ibernazione, caratterizzato da temperature che possono scendere a 6-8 °C e da una caduta della frequenza cardiaca a 6-7 pulsazioni al minuto, vi possono essere lunghi periodi di silenzio elettrico nella corteccia cerebrale. È molto improbabile che l'azione ristoratrice del sonno possa svolgersi nei neuroni corticali a temperatura così bassa. Il significato funzionale della letargia dell'animale ibernante sembrerebbe quindi essere del tutto differente da quello del sonno fisiologico.
Vi è tuttavia un lento ritmo ciclico fra i periodi di risveglio che interrompono l'ibernazione e le lunghe fasi di letargo. Questo ritmo è però del tutto differente dal ciclo dell'animale non ibernante. A mezzo inverno l'animale può rimanere in letargo per un periodo che dura fino a 14 giorni ed essere sveglio per sole 12-14 ore. Questo lento ciclo deve essere dovuto a meccanismi ipotalamici o del tronco dell'encefalo, giacché non è verosimile che la corteccia cerebrale silente possa intervenire nel risveglio.
bibliografia
Adrian, E. D., Electrical activity of the nervous system, in ‟Archives of neurology and psychiatry", 1934, XXXII, pp. 1125-1136.
Adrian, E. D., Moruzzi, G., Impulses in the pyramidal tract, in ‟Journal of physiology", 1939, XCVII, pp. 153-199.
Alekseyeva, T. T., Correlation of nervous and humoral factors in the development of sleep in non-disjointed twins, in ‟Zhurnal vysshei nervnoi deyatel'nosti imeni I. P. Pavlova", 1958, VIII, pp. 835-844.
Amatruda, T. T. III, Black, D. A., McKenna, T. M., McCarley, R. W., Hobson, J. A., Sleep cycle control and cholinergic mechanisms: differential effects of carbachol injections at pontien brain stem sites, in ‟Brain research", 1975, XCVIII, pp. 501-515.
Axelrod, J., Brain monoamines, in ‟Neurosciences research program bulletin", 1971, IX, pp. 188-196.
Batini, C., Moruzzi, G., Palestini, M., Rossi, G. F., Zanchetti, A., Effects of complete pontine transections on the sleep-wakefulness rhythm: the midpontine pretrigeminal preparation, in ‟Archives italiennes de biologie", 1959, XCVII, pp. 1-12.
Batsel, H. L., Electroencephalographic synchronization and desynchronization in the chronic ‛cerveau isolé' of the dog, in ‟Electroencephalography and clinical neurophysiology", 1960, XII, pp. 421-430.
Batsel, H. L., Spontaneous desynchronization in the chronic cat ‛cerveau isolé', in ‟Archives italiennes de biologie", 1964, CII, pp. 547-566.
Baust, W., Die Phänomenologie des Schafles, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 99-144.
Belugou, J. L., Benoit, O., Leygonie, F., Décharges neuronales de l'hippocampe au cours de la veille et du sommeil, in ‟Journal de physiologie", 1968, LX, suppl. 2, p. 399.
Berger, H., Über das Elektroenzephalogramm des Menschen, in ‟Archiv für Psychiatrie", 1929, LXXXVII, pp. 527-570.
Berlucchi, G., Callosal activity in unrestrained, unanesthetized cats, in ‟Archives italiennes de biologie", 1965, CIII, pp. 623-634.
Berlucchi, G., Mechanismes von Schlafen und Wachen, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 145-203.
Bernard, Cl., Introduction à l'étude de la médecine expérimentale, Paris 1865.
Bonvallet, M., Système nerveux et vigilance, Paris 1966.
Bremer, F., Cerveau ‛isolé' et physiologie du sommeil, in ‟Comptes rendus des séances de la Société de Biologie", 1935, CXVIII, pp. 1235-1241.
Bremer, F., L'activité cérébrale au cours du sommeil et de la narcose. Contribution à l'étude du mécanisme du sommeil, in ‟Bulletin de l'Académie Royale de Médecine de Belgique", 1937, IV, pp. 68-86.
Bremer, F., L'activité électrique de l'écorce cérébrale et le problème physiologique du sommeil, in ‟Bollettino della Società Italiana di Biologia Sperimentale", 1938, XIII, pp. 271-290.
Bremer, F., Preoptic hypnogenic focus and mesencephalic reticular formation, in ‟Brain research", 1970, XXI, pp. 132-134.
Bremer, F., Existence of a mutual tonic inhibitory interaction between the preoptic hypnogenic structures and the midbrain reticular formation, in ‟Brain research", 1975, XCVI, pp. 71-75.
Bremer, F., Cerebral hypnogenic centers, in ‟Archives of neurology", 1977, II, pp. 1-6.
Brunelli, M., Magni, F., Moruzzi, G., Musumeci, D., Effetti di stimolazioni elettriche localizzate del ponte sulle attività istintive del piccione talamico e del piccione integro, in ‟Atti dell'Accademia Nazionale dei Lincei" (serie VIII), 1971, II, pp. 603-606.
Brunelli, M., Magni, F., Moruzzi, G., Musumeci, D., Effects of pontine stimulations on sleep and waking behaviors of the pigeon, in Basic sleep mechanisms (a cura di O. Petre-Quadens e J. D. Schlag), New York-London 1974, pp. 33-49.
Buser, P., Viala, G., Analyse du mécanisme de l'akinésie induite ‛hypnotique' chez le lapin, in Actualités neurophysiologiques (a cura di A. M. Monnier), vol. VIII, Paris 1968, pp. 179-196.
Carli, G., Dissociation of electrocortical activity and somatic reflexes during rabbit hypnosis, in ‟Archives italiennes de biologie", 1969, CVII, pp. 219-234.
Carli, G., Subcortical mechanisms of rabbit hypnosis, in ‟Archives italiennes de biologie", 1971, CIX, pp. 15-26.
Clemente, C. D., Sterman, M. B., Wyricka, W., Forebrain inhibitory mechanisms: conditioning of basal forebrain induced EEG synchronization and sleep, in ‟Experimental neurology", 1963, VII, pp. 404-417.
Craig, W., Appetites and aversions as constituents of instincts, in ‟Biological bulletin", 1918, XXXIV, pp. 91-107.
Dell, P., Lower brain stem mechanisms and sleep induction, in Proceedings of the XXV international congress of physiology, München 1971, pp. 29-30.
Dement, W. C., Kleitman, N., Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming, in ‟Electroencephalography and clinical neurophysiology", 1957, IX, pp. 673-690.
Dement, W. C., Rechtschaffen, A., Narcolepsy: polygraphic aspects, experimental and theoretical considerations, in The abnormalities of sleep in man (a curadi H. Gastaut, E. Lugaresi, G. Berti Ceroni e G. Coccagna), Bologna 1968, pp. 147-164.
Draskóczy, P. R., Lyman, C. P., Turnover of catecholamines in active and hibernating ground squirrels, in ‟Journal of pharmacology and experimental therapeutics", 1967, CLV, pp. 101-111.
Economo, C. von, Die Encephalitis lethargica, Wien 1918.
Economo, C. von, Schlaftheorie, in ‟Ergebnisse der Physiologie", 1929, XXVIII, pp. 312-339.
Evarts, E. V., Activity of neurons in visual cortex of the cat during sleep with low voltage fast EEG activity, in ‟Journal of neurophysiology", 1962, XXV, pp. 812-816.
Evarts, E. V., Temporal patterns of discharge of pyramidal tract neurons during sleep and waking in the monkey, in ‟Journal of neurophysiology", 1964, XXVII, pp. 152-171.
Evarts, E. V., Relation of cell size to effects of sleep in pyramidal tract neurons, in Progress in brain research, vol. XVIII, Sleep mechanisms (a cura di K. Akert, C. Bally e J. P. Schadé), Amsterdam 1965, pp. 81-91.
Evarts, E. V., Relation of discharge frequency to conduction velocity in pyramidal tract neurons, in ‟Journal of neurophysiology", 1965, XXVIII, pp. 215-228.
Evarts, E. V., Activity of individual cerebral neurons during sleep and arousal, in Association for Research in Nervous and Mental Disease, vol. XLV, Sleep and altered states of consciousness, Baltimore 1967, pp. 319-337.
Feinberg, I., Evarts, E. V., Changing concepts of the function of sleep: discovery of intense brain activity during sleep calls for revision of hypotheses as to its function, in ‟Biological psychiatry", 1969, I, pp. 331-348.
Genovesi, U., Moruzzi, G., Palestini, M., Rossi, G. P., Zanchetti, A., EEG and behavioral patterns following lesions of the mesencephalic reticular formation in chronic cats with implanted electrodes, in Abstracts of communications. 20th international physiological congress, Bruxelles 1956, pp. 335-336.
Gerebtzoff, M., État fonctionnel de l'écorce cérébrale au cours de l'hypnose animale, in ‟Archives internationales de physiologie", 1941, LI, pp. 365-378.
Hediger, H., Wie Tiere schlafen, in ‟Medizinische Klinik", 1959, LIV, pp. 938-946, 965-968.
Hediger, H., Comparative observations on sleep, in ‟Proceedings of the Royal Society of Medicine", 1969, LXII, pp. 153-156.
Heller, H. C., Glotzbach, S. F., Thermoregulation during sleep and hibernation, in ‟International review of physiology", 1977, XV, pp. 147-188.
Hess, W. R., Stammganglien-Reizversuche. Tagung der deutschen physiologischen Gesellschaft, in ‟Berichte über die gesamte Physiologie und experimentelle Pharmakologie", 1927, XLII, pp. 554-555.
Hess, W. R., Das Schlafsyndrom als Folge dienzephaler Reizung, in ‟Helvetica physiologicva et pharmacologica acta", 1944, II, pp. 305-344.
Hess, W. R., Das Zwischenhirn. Syndrome, Lokalisationen, Funktionen, Basel 1949.
Hess, W. R., Psychologie in biologisher Sicht, Stuttgart 1968.
Hess, W. R., Hypothalamus und Thalamus, Stuttgart 1969.
Hobson, J. A., McCarley, R. W., Discharge patterns of cat pontine brain stem neurons during desynchronized sleep, in ‟Journal of neurophysiology", 1975, XXXVIII, pp. 751-766.
Hobson, J. A., McCarley, R. W., Wyzinski, P. W., Sleep cycle oscillation: reciprocal discharge by two brain stem neuronal groups, in ‟Science", 1975, CLXXXIX, pp. 55-58.
Holm, E., Die pharmakologische induzierte Schlaf, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 247-308.
Holzapfel, M., Triebbedingte Ruhezustände als Ziel von Appetenzhandlungen, in ‟Naturwissenschaften", 1940, XXVIII, pp. 273-280.
Hoshino, K., Pompeiano, O., Selective discharge of pontine neurons during the postural atonia produced by an anticholinesterase in the decerebrate cat, in ‟Archives italiennes de biologie", 1976, CXIV, pp. 244-277.
Jouvet, M., Recherches sur les structures nerveuses et les mécanismes responsables des différentes phases du sommeil physiologique, in ‟Archives italiennes de biologie", 1962, C, pp. 125-206.
Jouvet, M., Neurophysiology of the states of sleep, in ‟Physiological reviews", 1967, XLVII, pp. 117-177.
Jouvet, M., The role of monoamine and acetylcholine containing neurons in the regulation of the sleep-waking cycle, in ‟Ergebnisse der Physiologie", 1972, LXIV, pp. 166-307.
Jouvet, M., Le sommeil paradoxal est-il responsable d'une programmation génétique du cerveau?, in ‟Comptes rendus des séances de la Société de Biologie", 1978, CLXXII, pp. 9-32.
Jouvet, M., Delorme, F., Locus coeruleus et sommeil paradoxal, in ‟Comptes rendus des séances de la Société de Biologie", 1965, CLIX, pp. 895-899.
Jouvet, M., Renault, J., Insomnie persistante après lésions des noyaux du raphé chez le chat, in ‟Comptes rendus des séances de la Société de Biologie", 1966, CLX, pp. 1461-1465.
Klaue, R., Die bioelektrische Tätigkeit der Grosshirnrinde in normalen Schlaf und in der Narkose durch Schlafmittel, in ‟Journal für Psychologie und Neurologie", 1937, XLVII, pp. 510-531.
Kleitman, N., Sleep and wakefulness, Chicago 1963.
Klemm, W. R., Electroencephalographic-behavioral dissociations during animal hypnosis, in ‟Electroencephalography and clinical neurophysiology", 1966, XXI, pp. 365-372.
Konorski, J., Integrative activity of the brain, Chicago-London 1967.
Lindsley, D. B., Bowden, J. W., Magoun, H. W., Effect upon the EEG of acute injury to the brain stem activating system, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 475-486.
Lindsley, D. B., Schreiner, L. H., Knowles, W. B., Magoun, H. W., Behavioral and EEG changes following chronic brain stem lesions in the cat, in ‟Electroencephalography and clinical neurophysiology", 1950, II, pp. 483-498.
Luce, G. G., Current research on sleep and dreams, Public Health Service Publication No. 1389, Bethesda 1965.
McGinty, D. J., Sterman, M. B., Sleep suppression after basal forebrain lesions in the cat, in ‟Science", 1968, CLX, pp. 1253-1255.
Magoun, H. W., The waking brain, Springfield 19632.
Mantegazzini, P., Glässer, A., Action de la DL-3,4-dioxyphenylalanine (Dopa) et de la dopamine sur l'activité électrique du chat ‛cerveau isolé', in ‟Archives italiennes de biologie", 1960, XCVIII, pp. 367-374.
Marchiafava, P. L., Pompeiano, O., Pyramidal influences on spinal cord during desynchronized sleep, in ‟Archives italiennes de biologie", 1964, CII, pp. 500-529.
Moruzzi, G., The physiological properties of the brain stem reticular system, in Brain mechanisms and consciousness (a cura di E. D. Adrian, F. Bremer e H. H. Jasper), Oxford 1954, pp. 21-53.
Moruzzi, G., Synchronizing influences of the brain stem and the inhibitory mechanisms underlying the production of sleep by sensory stimulation, in ‟Electroencephalography and clinical neurophysiology", 1960, suppl. 13, pp. 231-256.
Moruzzi, G., Active processes in the brain stem during sleep, in ‟Harvey lectures", 1963, LVIII, pp. 233---297.
Moruzzi, G., The historical development of the deafferentation hypothesis of sleep, in ‟Proceedings of the American Philosophical Society", 1964, CVIII, pp. 19-28.
Moruzzi, G., Summary statement, in Progress in brain research, vol. XVIII, Sleep mechanisms (a cura di K. Akert, C. Bally e J. P. Schadé), Amsterdam 1965, pp. 241-243.
Moruzzi, G., The functional significance of sleep with particular regard to the brain mechanisms underlying consciousness, in Brain and conscious experience (a cura di J. C. Eccles), Berlin-Heidelberg-New York 1966, pp. 345-388.
Moruzzi, G., Sleep and instinctive behavior, in ‟Archives italiennes de biologie", 1969, CVII, pp. 175-216.
Moruzzi, G., The sleep-waking cycle, in ‟Ergebnisse der Physiologie", 1972, LXIV, pp. 1-165.
Moruzzi, G., Neural mechanisms of the sleep-waking cycle. Basic sleep mechanism (a cura di O. Petre-Quadens e J. D. Schlag), New York-London 1974, pp. 13-31.
Moruzzi, G., Magoun, H. W., Brain stem reticular formation and activation of the EEG, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 455-473.
Mukhametov, L. M., Rizzolatti, G., Tradardi, V., Spontaneous activity of neurones of nucleus reticularis thalami in freely moving cats, in ‟Journal of physiology", 1970, CCX, pp. 651-667.
Orthner, H., Neuroanatomische Gesichtspunkte der Schlaf-Wach-Regelung, in Der Schlaf (a cura di U. J. Jovanović), München 1969, pp. 49-84.
Otter, H. P., Die pathologische Physiologie des Schlafes, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 217-245.
Pappenheimer, J. R., Koski, G., Fench, V., Induction of slow wave sleep by a factor released into cerebrospinal fluid (CSF) during sleep deprivation, in Proceedings of the XXV international congress of physiology. Abstracts of volunteer papers and films, vol. IX, München 1971, p. 439.
Pappenheimer, J. R., Miller, T. B., Goodrich, C. A., Sleep-promoting effects of cerebro-spinal fluid from sleep-deprived goats, in ‟Proceedings of the National Academy of Sciences", 1967, LVIII, pp. 513-517.
Parmeggiani, P. L., Rabini, C., Shivering and panting during sleep, in ‟Brain research", 1967, VI, pp. 789-791.
Piéron, H., Le problème physiologique du sommeil, Paris 1913.
Pivik, R. T., McCarley, R. W., Hobson, J. A., Eye movement-associated discharge in brain stem neurons during desynchronized sleep, in ‟Brain research", 1977, CXXI, pp. 59-76.
Pompeiano, O., Ascending and descending influences of somatic afferent volleys in unrestrained cats; supraspinal inhibitory control of spinal reflexes during natural and reflexly induced sleep, in Aspects anatomofonctionnels de la physiologie du sommeil (a cura di M. Jouvet), Paris 1965, pp. 309-395.
Pompeiano, O., Muscular afferents and motor control during sleep, in Muscular afferents and motor control (a cura di R. Granit), Stockholm 1966, pp. 415-436.
Pompeiano, O., Sensory inhibition during motor activity in sleep, in Neurophysiological basis of normal and abnormal motor activities (a cura di M. D. Yahr e D. P. Purpura), New York 1967, pp. 323-375.
Pompeiano, O., The neurophysiological mechanisms of the postural and motor events during desynchronized sleep, in Association for Research in Nervous and Mental Diseases, vol. XLV, Sleep and altered states of consciousness, Baltimore 1967, pp. 351-423.
Pompeiano, O., Hoshino, K., Tonic inhibition of dorsal pontine neurons during the postural atonia produced by an anticholinesterase in the decerebrate cat, in ‟Archives italiennes de biologie", 1976, CXIV, pp. 310-340.
Pompeiano, O., Swett, J. E., EEG and behavioral manifestations of sleep induced by cutaneous nerve stimulation in normal cats, in ‟Archives italiennes de biologie", 1962, C, pp. 311-342.
Pompeiano, O., Swett, J. E., Identification of cutaneous and muscular afferent fibers producing EEG synchronization or arousal in normal catas, in ‟Archives italiennes de biologie", 1962, C, pp. 343-380.
Puzillout, J. J., Foutz, A. S., Characteristic of the experimental reflex sleep induced by vago-aortic nerve stimulation, in ‟Journal of electroencephalography and clinical neurophysiology", 1977, XLII, pp. 552-563.
Puizillout, J. J., Ternaux, J. P., Endormement vago-aortique après section sagittale médiane du tronc cérébral et après administration de p-chlorophenylalanine ou destruction des noyaux du raphé, in ‟Brain research", 1974, LXX, pp. 19-42.
Purpura, D. P., Frigyesi, T. L., McMurtry, J. C., Scarf, T., Synaptic mechanisms in thalamic regulation of cerebello-cortical projection activity, in The talamus (a cura di D. P. Purpura e M. D. Yahr), New York-London 1966, pp. 153-172.
Ranson, S. W., Somnolence caused by hypothalamic lesions in the monkey, in ‟Archives of neurology and psychiatry", 1939, XLI, pp. 1-23.
Roffwarg, H. P., Muzio, J. N., Dement, W. C., Ontogenetic development of the human sleep-dream cycle, in ‟Science", 1966, CLII, pp. 604-619.
Sastre, J. P., Effets des lésions du tegmentum pontique sur l'organisation des états de sommeil chez le chat, Lyon 1978.
Sàtinoff, E., Hibernation and the central nervous system, in Progress in physiological psychology (a cura di E. Stellar e J. M. Sprague), vol. III, New York 1970, pp. 201-236.
Sharpless, S., Jasper, H., Habituation of the arousal reaction, in ‟Brain", 1956, LXXIX, pp. 655-680.
Sherrington, C. S., Man on his nature, Cambridge 1946.
Steriade, M., Cortical long-axoned cells and putative interneurons during the sleep-waking cycle, in ‟The behavioral and brain sciences", 1978, III, pp. 465-514.
Steriade, M., Ropert, N., Kitsikis, A., Oakson, G., Ascending activating neuronal networks in midbrain reticular core and related rostral systems in The reticular formation revisited (a cura di J. A. Hobson e M. A. Brazier), New York 1980, pp. 125-167.
Sterman, M. B., Clemente, C. D., Forebrain inhibitory mechanisms: cortical synchronization induced by basal forebrain stimulation, in ‟Experimental neurology", 1962, VI, pp. 91-102.
Sterman, M. B., Clemente, C. D., Forebrain inhibitory mechanisms: sleep patterns induced by basal forebrain stimulation in the behaving cat, in ‟Experimental neurology", 1962, VI, pp. 103-117.
Svorad, D., ‛Animal hypnosis' (Totstellreflex) as experimental model for psychiatry. Electroencephalographic and evolutionary aspects, in ‟American Medical Association. Archives of neurology and psychiatry", 1957, LXXVII, pp. 533-539.
Tinbergen, N., ‛Derived' activities: their causation, biological significance, origin, and emancipation during evolution, in ‟Quarterly review of biology", 1952, XXVII, pp. 1-32.
Tinbergen, N., The study of instinct, Oxford 19552.
Tönnies, J. F., Automatische EEG-Intervall-Spektrumanalyse (EISA) zur Langzeitdarstellung der Schlafperiodik und Narkose, in ‟Archiv für Psychiatrie und Nervenkrankheiten", 1969, CCXII, pp. 423-445.
Villablanca, J., The electrocortigram in the chronic cerveau isolé cat, in ‟Electroencephalography and clinical neurophysiology", 1965, XIX, pp. 576-586.
Villablanca, J., Behavioral and polygraphic study of ‛sleep' and ‛wakefulness' in chronic decerebrate cats, in ‟Electroencephalography and clinical neurophysiology", 1966, XXI, pp. 562-577.
Il trasferimento interoculare e interemisferico è stato esaminato anche in Mammiferi inferiori, come il ratto (v. Sheridan, 1965), il coniglio (v. Van Hof, 1970) e la cavia (v. Levinson, 1972). In queste specie non si è cercato di produrre chirurgicamente una lateralizzazione delle afferenze visive, perché le vie ottiche sono già quasi interamente crociate. Risultati in accordo con quelli precedentemente ottenuti in gatti e scimmie con cervello bisecato sono stati descritti per i ratti albini (v. Sheridan, 1965) e le cavie (v. Levinson, 1972): il trasferimento interoculare di discriminazioni apprese con un occhio è gravemente disturbato dalla sezione del corpo calloso. Al contrario, i ratti con il pelo cefalico pigmentato presentano un buon trasferimento interoculare anche dopo callosotomia: ciò è dovuto con ogni probabilità al fatto che questi animali hanno un contingente di fibre ottiche non crociate più numeroso rispetto ai ratti albini, cosicché con la stimolazione monoculare si ha sempre un invio d'informazioni visive a entrambi gli emisferi (v. Sheridan, 1965; v. Levinson, 1972). Un trasferimento interoculare molto deficiente è stato osservato nel coniglio normale (v. Van Hof, 1970); le ragioni di queste differenze tra le specie non sono chiare.
Una brillante tecnica per lo studio del trasferimento interemisferico dell'apprendimento visivo nel ratto è stata realizzata da Bureš e collaboratori (v. Bureš e Burešovà, 1960; v. Burešovà e Bureš, 1969), che sostengono i vantaggi del preparato cervello bisecato reversibile rispetto al preparato cervello bisecato chirurgico. In breve, le attività nervose in un emisfero vengono bloccate temporaneamente dall'applicazione locale di KCl (depressione propagata), mentre il ratto apprende a fare una discriminazione con l'altro emisfero. Successivamente, la ritenzione della discriminazione viene esaminata quando l'emisfero previamente depresso è ritornato normale, e l'emisfero previamente normale è a sua volta sottoposto alla depressione propagata. La ritenzione da parte di quest'ultimo emisfero può poi essere ancora esaminata permettendone il recupero e spostando la depressione propagata di nuovo all'altro emisfero. I risultati di queste prove indicano che, almeno per quanto riguarda i compiti difficili, la capacità di discriminare è presente solo quando l'emisfero usato durante l'apprendimento originario funziona normalmente. L'emisfero inizialmente depresso è ovviamente in grado di apprendere la discriminazione, ma lo fa dimostrando di non aver tratto alcun vantaggio dall'esperienza dell'altro emisfero. Quindi, il preparato cervello bisecato reversibile mostra un'indipendenza dei due emisferi nell'apprendimento visivo simile a quello del preparato cervello bisecato chirurgico.
Una rassegna degli esperimenti sul trasferimento interoculare nei Pesci e negli Uccelli è stata recentemente pubblicata da M. Cuénod (v., 1972). Questi animali hanno vie ottiche completamente crociate e non hanno il corpo calloso: è perciò interessante conoscere per qual via il trasferimento oculare, quando sia presente, possa avvenire. Vi sono certe limitazioni al trasferimento interoculare di discriminazioni visive, dovute probabilmente alla natura degli stimoli (v. Sperry e Clark, 1949; v. Schulte, 1957), alla loro posizione nel campo visivo (v. Levine, 1945; v. Ingie, 1968) e a variabili relative all'addestramento e ai metodi d'esame (v. Mc Cleary, 1960). Tuttavia, in condizioni sperimentali appropriate, si dimostra che il trasferimento interoculare di varie discriminazioni visive è presente; esso viene abolito dalla sezione delle commissure sopraottiche negli Uccelli, come, per es., nel piccione (v. Cuénod e Zeier, 1967; v. Meier, 1971), e della commissura intertettale nei Pesci (v. Mark, 1966).
Gli esperimenti sul cervello bisecato e la visione sono ormai innumerevoli e per limiti di spazio è impossibile presentarne una rassegna anche superficiale. È tuttavia interessante discutere i problemi sollevati da alcuni di questi esperimenti, sia perché essi interessano la ricerca sul cervello bisecato in genere, sia in vista d'importanti sviluppi futuri nello studio delle comunicazioni interemisferiche.
Risulta chiaramente da molti degli studi sopra descritti che nell'animale normale le commissure telencefaliche devono in qualche modo permettere ai due emisferi di scambiarsi informazioni visive inizialmente raccolte da uno solo di essi. La prima domanda che si presenta riguarda la seguente alternativa: 1) l'emisfero che riceve direttamente l'informazione visiva ne manda immediatamente un duplicato all'altro emisfero, in modo che l'apprendimento basato su tale informazione procede in parallelo nei due emisferi, e ciascuno di essi forma una traccia mnemonica (engramma) propria; oppure, 2) l'engramma si stampa solo nell'emisfero direttamente stimolato e le commissure telencefaliche lo mettono semplicemente a disposizione dell'emisfero opposto quando le informazioni visive appropriate sono inviate all'altro lato. Espressa in altre parole, la domanda è: il trasferimento d'informazioni visive da parte delle commissure telencefaliche è di natura esclusivamente sensoriale, con conseguente engramma bilaterale, o di natura esclusivamente mnemonica, in relazione con un engramma unilaterale? Non si può dare una risposta definitiva a questa domanda. La fisiologia dimostra che il corpo calloso comunica informazioni visive dirette (v. sopra, cap. 3, § b), ma sembra improbabile che il trasferimento interemisferico della pratica visiva studiato negli esperimenti sul cervello bisecato dipenda da questo scambio d'informazioni visive (v. Berlucchi, 1972). Gli esperimenti neurocomportamentali forniscono prove in favore d'entrambe le ipotesi: in certe condizioni, è del tutto chiaro che afferenze visive lateralizzate portano alla formazione sia di un engramma diretto nell'emisfero bersaglio, sia, attraverso le commissure telencefaliche, di un engramma indiretto nell'emisfero opposto (e l'engramma indiretto è in genere più debole di quello diretto) (v., ad es., Myers, Transmission of visual..., 1962; v. Gazzaniga, 1966; v. Butler, 1968). D'altra parte, risultati che fanno pensare ad engrammi unilaterali sono stati ottenuti in esperimenti in cui le variabili critiche erano probabilmente la difficoltà della discriminazione (v. Myers e Sperry, 1958; v. Myers, Transmission of visual..., 1962) e l'esperienza precedente dell'animale (v. Gazzaniga, 1966). L'unilateralità degli engrammi è la regola nel preparato cervello bisecato reversibile (v. sopra). Un nuovo approccio che dimostra la possibilità di lateralizzare gli engrammi a un emisfero è stato proposto da Doty e Negrão nella scimmia (v., 1973). La commissura anteriore e il corpo calloso, salvo lo splenio, vengono tagliati e un laccio viene posto attorno allo splenio intatto. La scimmia viene poi addestrata a premere una leva quando si stimola selettivamente la corteccia visiva (area striata) di un lato. Dopo che l'animale ha appreso, si può dimostrare che la risposta condizionata è suscitata anche dalla stimolazione del punto corrispondente nella corteccia visiva opposta. Tuttavia, se il laccio attorno allo splenio viene tirato, completando così la callosotomia, la risposta alla stimolazione della corteccia visiva non allenata sparisce, mentre la stimolazione della corteccia visiva allenata continua a produrre con la stessa efficacia di prima la risposta condizionata. Questo risultato sta chiaramente in favore dell'idea che, almeno in queste condizioni sperimentali, l'engramma è unilaterale, e lo splenio è necessario per renderlo accessibile all'emisfero controlaterale. Quando la commissura anteriore è intatta, e il calloso completamente sezionato, sembra invece che si stabiliscano engrammi bilaterali; se si tira il laccio attorno alla commissura anteriore la risposta alla stiolazione della corteccia non allenata non viene abolita, ma si può dimostrare, con un procedimento di estinzione, che l'engramma in questa corteccia è molto più debole di quello nella corteccia allenata (v. Doty e Negrão, 1973).
Varie domande possono essere poste in relazione agli aspetti anatomici del trasferimento interemisferico della pratica visiva. Quali sono le aree corticali implicate nel trasferimento commissurale d'informazioni, sia esso sensoriale o mnemonico, che è necessario perché l'esperienza visiva acquisita tramite un solo emisfero sia a disposizione di entrambi gli emisferi? N. Geschwind (v., 1965) ha esaminato in una rassegna vecchi e nuovi studi sulle disconnessioni corticali nell'uomo e negli animali, giungendo alla conclusione che le connessioni anatomiche fra la corteccia visiva e le strutture limbiche e temporali sono essenziali per la percezione, per l'apprendimento e per la memoria visiva. M. Mishkin (v., 1966), J. A. Horel e E. G. Keating (v., 1969) hanno dimostrato che, dopo lesione del lobo occipitale in un emisfero e del lobo temporale nell'altro, l'integrità del corpo calloso diventa critica per le discriminazioni visive, per il riconoscimento degli oggetti e per il comportamento di paura in risposta a stimoli visivi. Per spiegare questi risultati, Mishkin (v., 1966 e 1972) ha proposto l'ipotesi che l'informazione visiva sia trasmessa dall'area striata alla corteccia prestriata ipsilaterale, e dalla corteccia prestriata venga inviata sia alla corteccia ipsilaterale inferotemporale, sia, attraverso il corpo calloso, alla corteccia prestriata controlaterale. L'idea che questa stessa via sia usata per la trasmissione interemisferica dell'esperienza visiva è molto interessante, e vi è qualche prova diretta in favore d'essa (v. Zeki, 1967). Nel gatto con chiasma sezionato, lesioni che distruggono le porzioni delle aree corticali visive 17, 18 e 19, che rappresentano le parti del campo visivo in prossimità del meridiano verticale centrale (v. sopra, cap. 3, § b), non interferiscono con il trasferimento interoculare di discriminazioni apprese con un solo occhio. Sembrerebbe perciò che questo trasferimento venga effettuato da altre aree corticali connesse attraverso il corpo calloso (v. Berlucchi, 1972).
Sperry (v., per es., Some developments..., 1961) ha ripetutamente suggerito l'uso dei preparati con cervello bisecato per lo studio degli effetti delle lesioni cerebrali sull'apprendimento, sia nella modalità visiva che in altre modalità. Poiché l'apprendimento visivo può svolgersi indipendentemente nei due emisferi dell'animale con cervello bisecato, un emisfero può essere sottoposto a lesioni sperimentali che si suppone interferiscano con la visione, e le sue capacità visive possono essere poi paragonate a quelle dell'emisfero intatto. In questi esperimenti lo stesso animale funge sia da soggetto sperimentale che da controllo, a seconda dell'emisfero che si esamina. L'utilità di questo interessante approccio è stata ripetutamente dimostrata (v., ad es., Sperry e altri, 1960; v. Barrett, 1969; v. Berlucchi e altri, 1972).
Per quanto riguarda la somatoestesia e la coordinazione sensorio-motoria, si considera che l'era della ricerca sul cervello bisecato sia iniziata con il famoso esperimento fatto da K. Bykoff (1924) nel laboratorio dii. P. Pavlov. Si sapeva che cani condizionati a secernere saliva in risposta alla stimolazione tattile di un punto particolare della superficie di un lato del corpo presentavano una perfetta generalizzazione della risposta alla stimolazione del punto corrispondente dell'altro lato del corpo (v. Pavlov, 1927). Bykoff (v., 1924) dimostrò che a seguito della sezione del corpo calloso questa generalizzazione controlaterale era assente, e che riflessi indipendenti potevano essere instaurati con stimoli uguali applicati ai due lati del corpo. Vari anni dopo, le osservazioni di Bykoff furono ripetute da J. 5. Stamm e R. W. Sperry (v., 1957) e da T. H. Meikle jr., J. A. Sechzer e E. Stellar (v., 1962) in una specie differente (gatto) e usando un diverso procedimento di condizionamento (strumentale anziché classico). L'abolizione del trasferimento intermanuale dell'apprendimento unimanuale di varie classi di discriminazioni somestesiche è stata osservata anche in Primati commissurotomizzati. Tuttavia, è stato anche riferito che in alcuni casi il trasferimento può persistere (v. tab. II per un riassunto di risultati). Poiché si sa che in aggiunta alla componente controlaterale, molto più cospicua, vi è anche una piccola proiezione ipsilaterale a ciascun emisfero, è probabile che il trasferimento intermanuale positivo osservato negli animali commissurotomizzati dipenda dalla possibilità che i segnali sensitivi raggiungano entrambi gli emisferi durante l'addestramento, anziché da uno scambio interemisferico d'informazioni che avvenga a livello sottocorticale (v. Ettunger e Blakemore, 1969; v. Gazzaniga, 1970). Non è chiaro comunque perché le afferenze periferiche ipsilaterali debbano essere utilizzate in certi casi e non in altri. A ogni modo, non v'è dubbio che l'apprendimento somestesico può spesso procedere indipendentemente negli emisferi sconnessi dell'animale con cervello bisecato. H. Teitelbaum, S. K. Sharpless e R. Byck (v., 1968) hanno descritto che l'ablazione della seconda area somatosensitiva (S II) nel gatto non abolisce l'apprendimento d'una discriminazione tattile-cinestesica con la zampa controlaterale, ma abolisce il trasferimento della discriminazione alla zampa corrispondente dell'altro lato. È quindi probabile che i neuroni dell'area S II con campi recettivi bilaterali (v. sopra, cap. 3, § b) siano un substrato critico per il trasferimento interemisferico dell'esperienza tattile. Il curioso conflitto fra i dati anatomici che dimostrano l'assenza di connessioni callosali fra le porzioni dell'area SI e S II che rappresentano la mano e il piede (v. sopra, cap. 2, § b) e i dati comportamentali che indicano che il trasferimento intermanuale e interpedale dipende dal corpo calloso, è stato esaminato in dettaglio da C. R. Butler e A. C. Francis (v., 1973). I loro esperimenti indicano che quando l'informazione tattile è rigorosamente limitata alle dita, il trasferimento intermanuale è assente perfino nella scimmia normale! Forse i trasferimenti positivi osservati nei precedenti esperimenti dipendevano da informazioni somestesiche provenienti dalle parti prossimali nell'arto usato nell'apprendimento.
di Franco Angeleri e Zdenæk Servít
Epilessia
Sommario: 1. Introduzione. 2. Fisiopatologia (patogenesi) dell'epilessia: a) patogenesi della crisi epilettica; b) patogenesi del processo epilettico. 3. Aspetti clinici dell'epilessia: a) predisposizione ed ereditarietà; b) correlazioni anatomo-elettrocliniche; c) epilessie a scatenamento o ad arresto sensoriale; d) terapia dell'epilessia. □ Bibliografia.
1. Introduzione
L'epilessia può essere definita in modo diverso a seconda dell'obiettivo di ricerca che ci si propone. Dal punto di vista medico è utile una definizione clinica, nel tentativo di delimitare e distinguere l'epilessia da altre malattie che hanno sintomi simili. In questo senso essa risulta una malattia che si manifesta prevalentemente con crisi a differenti intervalli di tempo. Una crisi epilettica è caratterizzata da un improvviso e transitorio disturbo dell'attività cerebrale, accompagnato da un insieme di sintomi parossistici che, separatamente o in varie combinazioni, possono interessare ogni funzione neurologica o psichica.
Scopo della definizione fisiopatologica (o patogenetica) è quello di chiarire i meccanismi della genesi della malattia e dei suoi sintomi. Una simile definizione evidentemente è condizionata dalle conoscenze scientifiche su tali meccanismi. In base a quanto oggi noto, la crisi epilettica può essere definita come espressione di un'attività automatica parossistica e ritmica di estese popolazioni di cellule nervose, escluse così dall'attività fisiologica del cervello.
Sia dal punto di vista clinico sia da quello fisiopatologico è importante distinguere una crisi epilettica dalla ‛malattia epilessia' la cui base fisiopatologica è un processo epilettico. Le componenti fondamentali di questo processo sono rappresentate da una progressiva modificazione della predispozione alle crisi e dalla formazione di focolai epilettici secondari (v. cap. 2, È b).
La storia dell'epilessia riflette non soltanto il livello di sviluppo della medicina (sia sul piano clinico sia su quello fisiologico), ma anche gli aspetti sociologici e filosofici che tale malattia ha chiamato in causa per la strana e drammatica sintomatologia delle sue crisi (compromissione o perdita di coscienza, convulsioni o altri comportamenti involontari che colpiscono il paziente improvvisamente, mentre sembra essere perfettamente normale).
Nell'antichità, nel Medioevo e oltre, questi sintomi, per la loro stessa natura, indussero a una interpretazione magica della crisi, come espressione di un intervento di forze esterne e irrazionali. Sono facilmente immaginabili le conseguenti implicazioni sociali e ideologiche. La storia dell'epilessia dall'antichità al tempo presente è stata passata in rassegna in modo completo nella monografia di O. Temkin (v., 1945).
La seconda metà del sec. XIX è stata contrassegnata da una svolta importante nelle ricerche sull'epilessia, soprattutto a causa delle scoperte neuroanatomiche. La struttura ma- cr0- e microscopica del cervello veniva gradualmente a chiarirsi mentre lentamente si conducevano le prime ricerche sulle funzioni cerebrali. In questo contesto hanno particolare importanza i contributi di Fritsch e Hitzig (v., 1870) sull'eccitabilità della corteccia, gli studi sperimentali di Ferrier (v., 1873) e la concezione fisiopatologica di Jackson (v., 1870) secondo la quale una convulsione è ‟una scarica del tessuto nervoso sui muscoli, occasionale, eccessiva e disordinata".
Nel sec. XX la fisiologia e l'anatomia funzionale (per es., la descrizione dell'anatomia cerebrale macro- e microscopica da un punto di vista funzionale) hanno contribuito in modo decisivo alla ricerca sull'epilessia. Ciò è stato reso possibile anche dal perfezionarsi dei metodi di indagine, specialmente con l'affermarsi e lo svilupparsi dell'elettrofisiologia (v. elettrofisiologia; v. Brazier, 1961), di alcune tecniche di biochimica (specialmente quelle di citochimica) e di microscopia elettronica (v. De Robertis, 1967).
Il progresso nella fisiopatologia dell'epilessia è stato influenzato specialmente dalle seguenti scoperte o ipotesi: il concetto di epilessia centroencefalica (v. cap. 2, È a, 3; v. cap. 3, È b, 3; v. Jasper, 1965), le correlazioni tra sintomi delle crisi e localizzazione cerebrale (v. cap. 2, È a, 5; v. cap. 3, È b; v. Penfield e Jasper, 1954), l'estesa applicazione dell'elettroencefalografia clinica e sperimentale (v. Berger, 1929; v. Adrian, 1934; v. Bremer, 1949; v. Gibbs e altri, 1935), la scoperta del sistema reticolare ascendente, cioè di un doppio sistema afferente al cervello (v. cap. 2, È a, 1; v. Jasper, 1949; v. sistema reticolare ascendente), la delucidazione dei meccanismi eccitatori e inibitori a livello cellulare (v. cap. 2, È a, 1; v. Eccles 1964 e 1969) e gli studi delle sinapsi al microscopio elettronico (v. De Robertis, 1967).
2. Fisiopatologia (patogenesi) dell'epilessia
Il cervello di qualsiasi vertebrato (compreso quello dell'uomo sano) può reagire a un certo stimolo e a certe condizioni con una crisi epilettica. La predisposizione alla crisi può essere favorita da determinate circostanze. In tal caso le crisi possono essere scatenate da differenti stimoli o persino intervenire senza alcuna causa apparente. Le crisi spesso si originano in una determinata regione cerebrale che viene definita come focolaio epilettico. Quando le crisi si ripetono, e a maggior ragione se sono ricorrenti, la predisposizione a esse spesso si modifica, aumentando o diminuendo. In tal modo si può sviluppare nell'organismo, soprattutto a carico del cervello, un processo patologico, il processo epilettico, che è la base fisiopatologica della malattia epilettica.
a) Patogenesi della crisi epilettica.
1. Basi fisiologiche dei fenomeni epilettici. - Nel XX secolo le conoscenze sulla morfologia e sulla funzione dell'elemento fondamentale del tessuto nervoso, il neurone, si sono arricchite notevolmente. La microelettrofisiologia, per mezzo di registrazioni microelettrodiche (intra- ed extracellulari) dell'attività elettrica di singoli neuroni, ha dato importanti informazioni sulla genesi dell'impulso nervoso. Il potenziale elettrico cellulare insorge quando sulla membrana si realizza un certo grado di depolarizzazione. Per certe circostanze (dovute per es. a fattori metabolici o farmacologici) il livello di polarizzazione della membrana può essere inizialmente più elevato o più basso. Ad esempio la membrana può essere parzialmente depolarizzata per uno squilibrio del metabolismo ossidativo, cioè per mancanza di ossigeno o di glucosio (v. Tower, 1960; v. È b, 3).
Una membrana parzialmente depolarizzata si trova allora più prossima a quel livello di depolarizzazione capace di generare un impulso nervoso, oppure - se viene raggiunto tale livello - da essa si generano impulsi automaticamente, in assenza cioè di una eccitazione sinaptica. Questa situazione è particolarmente importante nella genesi dei fenomeni epilettici.
È stato inoltre dimostrato che in alcune condizioni l'attività elettrica di una estesa popolazione di neuroni può divenire sincrona. In tal caso può essere registrata anche da macroelettrodi posti sulla superficie del cuoio capelluto (elettroencefalografia; v. elettrofisiologia). L'attività sincrona parossistica di estesi aggregati neuronali (‛aggregati neuronali epilettici'; v. Ajmone-Marsan, 1961) è espressione tipica della presenza di attività epilettica nel cervello. Ne consegue che le anormalità bioelettriche registrabili con la elettroencefalografia rappresentano sintomi importanti dell'epilessia (v. punto 4; v. cap. 3, È b).
Da ricerche condotte negli ultimi anni risulta che nella genesi della sincronizzazione dei potenziali elettrici di aggregati neuronali epilettici può giocare un ruolo importante un disturbo dell'equilibrio del doppio feedback dei neuroni cerebrali (v. Dichter e Spencer, 1969). L'esistenza di questo doppio feedback è stata messa in evidenza in strutture corticali relativamente semplici (per es., nell'ippocampo); sembra comunque che esso possa essere considerato quale principio funzionale generale di tutte le strutture nervose (v. Eccles, 1969). Esso è schematizzato in modo semplice nella fig. 1. La cellula piramidale (B) è un importante elemento strutturale della corteccia cerebrale. Le cellule piramidali sono disposte nella corteccia a strati (A); i loro dendriti apicali (B, d.a.), paralleli, risultano verticali rispetto alla superficie corticale. I feedbacks si realizzano per mezzo delle cosiddette ‛collaterali ricorrenti' (B, c.r.). Una fibra collaterale ricorrente di tipo eccitatorio (C, c.r.e.) ha contatti sinaptici con diverse cellule. Ciò rappresenta un'importante base strutturale per la sincronizzazione. In tal modo infatti l'impulso in uscita da una cellula può indurre la simultanea attivazione di un certo numero di altri neuroni. Le fibre ricorrenti collaterali di tipo inibitorio (c.r.i.) entrano in contatto sinaptico con i neuroni inibitori (n.i.), che, insieme alle cellule piramidali, rappresentano un altro elemento importante nella struttura corticale. Gli assoni delle cellule inibitrici stabiliscono delle giunzioni sinaptiche di tipo inibitorio con il soma delle cellule piramidali (s.i.). Il feedback inibitorio controlla l'attività delle cellule piramidali agendo, tra l'altro, come un freno di sicurezza, per prevenire o arrestare la loro attività ritmica automatica.
L'attività normale, ben bilanciata, del neurone piramidale dipende quindi in particolar modo da due condizioni: dal livello basale intrinseco della polarizzazione della sua membrana e dalla funzione proporzionale dei feedbacks eccitatono e inibitorio. Il neurone piramidale agisce come integratore degli impulsi eccitatori e inibitori che lo raggiungono mediante le sinapsi eccitatorie e inibitorie. Questi messaggi gli sono convogliati dalle fibre afferenti provenienti sia dai neuroni di altri centri encefalici, sia dai recettori che mediano stimoli estero- e intracettivi.
La neurofisiologia del XX secolo ha dimostrato che le vie afferenti che stabiliscono sinapsi con le cellule nervose sono di due tipi, quelle specifiche e quelle aspecifiche (v. fig. 1, C, v.a.s. e v.a.a.; v. Moruzzi e Magoun, 1949). Le afferenze specifiche probabilmente terminano in maggioranza in prossimità del soma cellulare. Esse mediano informazioni specifiche, per esempio una sensazione somestesica relativa a una determinata area della superficie corporea. Le afferenze aspecifiche terminano con tutta probabilità sui dendriti apicali, lontano dai corpi cellulari; in maggioranza prenderebbero contatto sinaptico con le ramificazioni dei dendriti apicali che nella corteccia cerebrale occupano gli strati superficiali. Esse non conducono informazioni specifiche, modulano invece il livello basale di reattività dei neuroni, probabilmente elevando o diminuendo il grado di polarizzazione della loro membrana. In questo modo le afferenze aspecifiche partecipano in modo sensibile al controllo sia dello stato di vigilanza sia della convulsività del cervello. Ciò spiega, ad esempio, perché spesso le crisi epilettiche nell'uomo siano legate al ritmo veglia-sonno.
2. Fattori patogenetici fondamentall dell'epilessia. - I fattori patogenetici dell'epilessia possono essere schematicamente suddivisi in tre gruppi: a) la predisposizione alle crisi (convulsività); b) il focolaio epilettico; c) lo stimolo epilettogeno.
La convulsività è condizionata da uno stato della membrana neuronale facilitante l'attività ritmica automatica e da una facilitazione verso l'attività sincrona e ritmica di estese popolazioni neuronali. A questo riguardo assume importanza fondamentale l'equilibrio del doppio feedback dei neuroni cerebrali (v. punto 1). Dal punto di vista funzionale ogni altra componente cerebrale può influenzare la convulsività: per esempio, l'esistenza di vie preformate o predisposte che sono sfruttate dall'attività epilettica del focolaio primario per propagare in altre strutture, talora verso regioni distanti ed estese dell'encefalo, oppure l'attività del sistema aspecifico che può influenzare la predisposizione verso crisi generalizzate (v. punto 1; v. cap. 3, È c).
Il focolaio epilettico è il punto reale o potenziale di origine e di propagazione della crisi; è localizato nella corteccia o in altre strutture cerebrali (degli aspetti clinici si dirà nel cap. 3, È b, 1). Nell'area che lo delimita la predisposizione alle crisi è aumentata. Di tanto in tanto i potenziali elettrici dei neuroni in essa compresi vanno incontro a una sincronizzazione il cui corrispettivo elettrofisiologico è rappresentato dall'attività epilettica focale che può essere registrata talora anche dal cuoio capelluto (spikes elettroencefalografici focali). A livello cellulare essa corrisponde a un'improvvisa e transitoria depolarizzazione della membrana accompagnata da una scarica di impulsi (spikes cellulari). Questa scarica è generalmente interrotta da una ripolarizzazione o iperpolarizzazione della membrana neuronale indotta principalmente dal feedback inibitorio (v. punto 1; v. Ajmone-Marsan, 1961).
Nella patologia umana la causa più frequente di un focolaio epilettico è rappresentata da un trauma cerebrale o da lesioni infiammatorie o circolatorie del tessuto nervoso (v. cap. 3, È b). Negli esperimenti su animali è possibile provocare un focolaio epilettogeno per mezzo di alcune sostanze (applicazione locale di penicillina, idrossido di alluminio, cobalto, ecc.). Recenti ricerche morfologiche su questi focolai sperimentali, eseguite al microscopio elettronico (v. Fischer, 1969), hanno dimostrato che nell'area attiva di un focolaio epilettico è possibile spesso osservare neuroni privi di sinapsi esclusivamente o prevalentemente nel soma, cioè nella zona dove probabilmente è situata la maggioranza delle sinapsi inibitorie. Si può quindi supporre che in tali neuroni sia disturbato il feedback inibitorio; ma essi sono anche privati del contatto con cellule di altro tipo, le cellule della nevroglia (astrociti) che rappresentano un'altra importante componente del tessuto nervoso. Esse partecipano al metabolismo dei neuroni contribuendo al mantenimento del normale livello di base della polarizzazione della loro membrana. Nelle cicatrici cerebrali gli astrociti sono rimpiazzati da un diverso tipo di cellule gliali (glia fibrosa) alle quali con tutta probabilità mancano le proprietà metaboliche sopra riferite. Complessivamente tutto ciò contribuisce a un notevole calo della polarizzazione della membrana neuronale, depolarizzazione che giunge così a valori assai prossimi al livello responsabile di un'attività autoritmica (v. Trachtenberg e Pollen, 1970; v. Pollen e Trachtenberg, 1970).
In esperimenti su animali è stato dimostrato che è possibile scatenare la scarica di un focolaio epilettico per mezzo di stimoli epilettogeni adeguati (stimolazione di un recettore sensoriale, stimolazione elettrica di una via afferente). Focolai localizzati nelle varie regioni cerebrali risentono dei diversi stimoli scatenanti a seconda dei rapporti che contraggono con le vie nervose afferenti. I contributi sperimentali di ricercatori italiani (v. Amantea, 1920; v. Clementi, 1929) sono stati, in questo settore, particolarmente significativi. La scarica epilettica talvolta è scatenata in modo predominante o esclusivo da certi organi di senso. Anche nell'epilessia umana talvolta certi stimoli acquistano un ruolo francamente epilettogeno, decisivo cioè nello scatenamento di una crisi (epilessia sensoriale riflessa; v. cap. 3, È c), ma quest'ultima è più frequente negli animali. Può essere, ad esempio, indotta in alcuni ratti, topi e conigli per mezzo di forti stimolazioni acustiche (epilessia audiogenica dei Roditori; v. Bevan, 1955) o da stimoli ottici in alcune specie di scimmie (epilessia fotogenica dei babuini; v. Killam, 1969).
Si può infine supporre, sulla base sia dei dati sperimentali sia delle conoscenze cliniche, che impulsi afferenti possano influenzare l'attività di un focolaio epilettico e l'origine delle crisi anche quando queste ultime non sono scatenate da uno stimolo costante. Abbiamo già ricordata l'influenza del sistema aspecifico sul grado di convulsività cerebrale (v. punto 1). Alcuni risultati sperimentali suggeriscono che anche una deafferentazione parziale di una certa zona cerebrale può aumentarne la convulsività (v. Stavraky, 1961; v. Echlin, 1959). Sembra che, quando l'attività dei neuroni non è sufficientemente stimolata da impulsi afferenti, in essi si realizzi una tendenza all'attività automatica spontanea.
3. Propagazione della crisi. - In molti casi la scarica epilettica rimane limitata alla zona del focolaio epilettico. Ciò può osservarsi nell'epilessia umana e può essere riprodotto sperimentalmente nell'animale. Comunque per certe condizioni l'attività epilettica si propaga dal focolaio ad altre regioni cerebrali, talora interessando vasti distretti degli emisferi cerebrali e in modo particolare del diencefalo (propagazione e generalizzazione delle scariche epilettiche). Questo fenomeno è stato ampiamente studiato con ricerche sperimentali. È stato così dimostrato che le scariche e la crisi si propagano in modo preferenziale attraverso alcune vie preformate dell'encefalo. Anche la convulsività delle diverse regioni cerebrali non è uguale. Oltre alla corteccia cerebrale anche alcune regioni sottocorticali hanno una maggiore convulsività (v. cap. 3, È b, 1). Vie importanti per la propagazione delle scariche sono le connessioni interemisferiche, il corpo calloso ed altre commissure (v. emisferi cerebrali: Interazioni interemisferiche cerebrali).
Alcune regioni encefaliche sembrano avere speciale importanza nella propagazione e generalizzazione delle crisi nei due emisferi. Esse sono rappresentate in specie da alcuni distretti diencefalici; in particolare dal talamo. Le connessioni tra la corteccia cerebrale e il talamo (circuiti talamo-corticali) senza dubbio giocano un ruolo importante nella genesi delle oscillazioni ritmiche dei potenziali elettrici registrati dall'EEG (v. elettrofisiologia), per esempio nella sincronizzazione dell'attività elettrica di estese popolazioni di neuroni. Tali strutture partecipano anche alla sincronizzazione dell'attività elettrica dei due emisferi. In elettroencefalografia ed elettrofisiologia esse sono indicate come pacemakers, cioè come promotrici di certi ritmi (v. cap. 3, È b, 2). Un altro importante pacemaker, oltre quello talamico, si trova nella regione del setto (v. Petsche e altri, 1962). Esso sembra avere un'influenza importante sull'attività ippocampale nella regione dei lobi temporali. Le regioni con funzione di pacemaker interverrebbero anche nel controllo dello stato di coscienza. Essendo tutte situate alla base dell'encefalo lungo la linea mediana ne formano quasi il centro topografico. Da ciò deriva il termine di ‛crisi centroencefaliche' con il quale sono state spesso indicate quelle crisi che si sviluppano con una sincronia biemisferica. Il termine è stato introdotto da Penfield e Jasper (v., 1954; v. Jasper, 1965; v. anche cap. 3, È b, 2).
Va tenuto presente comunque che la sincronizzazione dei potenziali elettrici cerebrali non coinvolge mai tutti gli elementi di una regione, come dimostrano precise misure elettrofisiologiche. Gli stessi neuroni compresi in un'area cerebrale o quelli appartenenti ai suoi differenti strati non partecipano in egual misura al fenomeno di sincronizzazione. È stato dimostrato inoltre che il pacemaker talamico non è indispensabile al realizzarsi di un sincronismo biemisferico, bastando a ciò le commissure interemisferiche. Nondimeno alle strutture con funzione di pacemaker deve essere attribuita una notevole importanza nella generalizzazione dell'attività epilettica.
4. Elettrogenesi delle alterazioni parossistiche nell'elettroencefalogramma. - Nelle ultime due decadi numerosi studi elettrofisiologici sono stati dedicati al problema dell'elettrogenesi dei potenziali elettroencefalografici, in particolare di quelli epilettici.
L'EEG dell'uomo a riposo è caratterizzato da alcuni ritmi fondamentali. Caratteristica delle alterazioni epilettiche (parossistiche) è in genere una maggiore ampiezza dei potenziali e una loro evidente maggiore ritmicità (v. fig. 2). Frequenze a 10 c/s, 4-5 c/s e 3 c/s si riscontrano molto spesso. Un'espressione assai tipica di epilessia nell'EEG è rappresentata dai complessi punta-onda con frequenza sui 3 c/s. Tale attività può essere facilmente provocata (quale fenomeno epilettico) in esperimenti su animali per mezzo di differenti tecniche e ciò non solo nei Mammiferi ma anche in animali con struttura cerebrale filogeneticamente più bassa (v. Servít e altri, 1965; v. Servít, 1965). Tale attività epilettica sembra dunque un aspetto basale e generale della reattività cerebrale.
Attraverso metodi elettrofisiologici è stato possibile correlare potenziali epilettici, derivati da macroelettrodi posti sulla superficie della corteccia cerebrale, con l'attività di singole cellule nervose (attività unitaria) comprese nella stessa regione. È stato così dimostrato che l'elettrocorticogramma di superficie è la risultante di una sincronizzazione dell'attività elettrica di vaste quantità di neuroni.
Comunque non tutti i neuroni di una certa regione partecipano in egual misura all'attività del tracciato macroelettrodico nè essi influenzano in modo eguale le modificazioni che risultano nel campo elettrico (v. Ajmone-Marsan, Microstructural..., 1965; v. Sawa e altri, 1968; v. Morrell, 1969). Alla genesi delle modificazioni del campo elettrico, derivabili con macroelettrodi persino dal cuoio capelluto, concorrono in particolar modo le cellule piramidali con i loro lunghi dendriti apicali disposti in parallelo. Da simile disposizione si generano dipoli elettrici, cioè differenze di potenziali tra corpo cellulare e terminazioni dendritiche apicali, campi elettrici discretamente intensi che insorgono durante la sincronizzazione dell'attività di tali neuroni.
Nel corso di una crisi epilettica interviene una progressiva e profonda depolarizzazione della membrana dei neuroni coinvolti nella crisi. A tale depolarizzazione si associa una temporanea eliminazione del meccanismo neuronale a feedback di tipo inibitorio che verosimilmente ne è anche la causa (v. punto 1). Questo problema è stato recentemente approfondito da Dichter e Spencer (v., 1969) anche per mezzo di modelli elettronici.
5. Correlazioni tra quadri clinici ed elettroencefalografici delle crisi epilettiche. - L'attività ritmica e automatica che caratterizza la crisi epilettica impedisce ai neuroni interessati di esplicare la loro funzione, spiccatamente differenziata; in tal modo essi vengono eliminati dai processi normali di informazione che si svolgono nel cervello. Ciò sembra un fattore importante nel determinismo dei sintomi clinici della crisi. I quadri clinici accessuali variano in rapporto alla localizzazione delle scariche e alla funzione dei neuroni coinvolti (v. cap. 3, È b).
Se la regione interessata dalla crisi epilettica non è molto estesa e se essa è abbastanza distante dal cuoio capelluto, può succedere che la crisi clinica non si accompagni a modificazioni dell'EEG. Al contrario quando la regione coinvolta dalla crisi è prossima al cuoio capelluto e di dimensioni tali da non compromettere funzioni cerebrali importanti, si verifica un fenomeno opposto, cioè la ricorrenza di crisi elettriche subcliniche rivelate dall'EEG. In molti epilettici si verifica in realtà che numerose crisi elettriche siano palesi solo alla registrazione elettroencefalografica.
Nel periodo iniziale della ricerca elettroencefalografica sono stati compiuti molti sforzi per individuare correlazioni elettrocliniche (v. Gastaut, 1954); confrontando criticamente quadri clinici ed EEG, in un primo tempo si concluse ottimisticamente per un rapporto abbastanza costante tra gli uni e gli altri. Tuttavia successive osservazioni cliniche e sperimentali hanno dimostrato che tali correlazioni non possono essere accettate come regola generale. La sintomatologia critica clinica è determinata principalmente dalla localizzazione delle scariche epilettiche. Tale localizzazione può condurre a particolari quadri elettroencefalografici che per altro non sempre indicano la stessa focalità (v. cap. 3, È b, 4).
b) Patogenesi del processo epilettico.
Abbiamo già accennato come da un punto di vista patogenetico sia importante distinguere la crisi epilettica dal processo epilettico. Solo quando quest'ultimo fenomeno si verifica si può parlare di epilessia come malattia. Anche sulla base della semplice esperienza clinica è possibile concludere che una crisi epilettica isolata non è necessariamente espressione di malattia epilettica. Una crisi isolata, infatti, può essere indotta anche in un cervello perfettamente sano per mezzo di particolari tecniche. Negli ultimi decenni la ricerca sperimentale, sulla falsariga dell'esperienza clinica, elettroencefalografica e neurochirurgica, ha offerto alcuni dati che delucidano i meccanismi del processo epilettico.
1. Propagazione dell'attività focale e origine di focolai secondari. - Registrazioni elettroencefalografiche seriate nell'epilessia umana documentano che un focolaio epilettico, localizzato in una regione di un emisfero cerebrale, molto spesso provoca l'origine di un altro focolaio epilettico nella regione speculare dell'emisfero opposto. Questo ‛focolaio speculare' (mirror focus) talora diviene un ‛focolaio secondario indipendente o autonomo', cioè rimane attivo anche quando il focolaio primario sia stato rimosso chirurgicamente. Focolai speculari e più in generale focolai secondari indipendenti sono riproducibili con relativa facilità anche nell'epilessia sperimentale. Nei Mammiferi il fenomeno è stato ampiamente studiato nel gatto e nelle scimmie (v. Udvarhelyi e Walker, 1965). Da tali ricerche è scaturito come primo dato fondamentale che la formazione di focolai secondari è favorita da una predisposizione di alcune strutture (ippocampo, amigdala e certe regioni corticali) e da alcune vie preformate (per es., le vie commissurali interemisferiche).
Ancor oggi l'origine di un focolaio autonomo secondario può essere spiegata solo in via ipotetica. Si sospetta che due meccanismi entrino in gioco nella sua patogenesi: a) una deafferentazione parziale dell'area interessata, conseguente all'interruzione delle normali afferenze che dovrebbero pervenire dalla regione del focolaio primario; b) un intenso e iterativo bombardamento di impulsi sincronizzati che provengono da questo alla regione del focolaio secondario. In quest'ultima potrebbero verificarsi anche processi inquadrabili nell'ambito della traccia mnesica (v. Morrell, 1960). Certo è che lo sviluppo di un focolaio secondario autonomo, a parte i suoi meccanismi patogenetici, rappresenta una tappa importante del processo epilettico.
2. Modificazione progressiva della predisposizione alle crisi. - Nel corso di un processo epilettico può realizzarsi una progressiva modificazione della predisposizione alle crisi, non soltanto nella sede dei focolai epilettici ma anche in più estese regioni cerebrali dove si verifica una propagazione delle scariche, come dimostrano alcune ricerche sperimentali. Analogamente si può sospettare che a modificare la predisposizione concorra una variazione delle reattività delle regioni con funzione di pacemaker (v. È a, 3). È infine verosimile che alterazioni metaboliche tissutali provocate dal processo epilettico inducano a loro volta modificazioni in senso epilettico sulla polarizzazione della membrana neuronale (v. È a, 3; v. cap. 3, È b, 3).
3. Biochimica dell'epilessia. - Negli ultimi cinquant'anni sono stati compiuti ripetuti tentativi per ricondurre l'epilessia a un disordine metabolico primario e per dare una definizione dei caratteri specifici di tale disturbo.
Una completa rassegna dei problemi concernenti i meccanismi biochimici che stanno alla base della patogenesi dell'epilessia è offerta dai lavori di Tower (v., 1960 e 1969) ai quali si rimanda per ulteriori informazioni bibliografiche. Certamente molti problemi rimangono ancora insoluti. Allo stato attuale delle nostre conoscenze può essere stabilito quanto segue.
L'ultimo anello di ogni influenza metabolica o biochimica capace di modificare la predisposizione epilettica è rappresentato dal livello di polarizzazione della membrana del neurone o, in altri termini, dalla differenza di potenziale tra l'interno e l'esterno della cellula (v. È a, 1; v. cap. 3, È a, 3). Numerosi e assai differenti fattori metabolici possono agire su tale bersaglio finale. Non è stato possibile invece scoprire un disturbo metabolico specifico dell'epilessia.
Il livello di polarizzazione della membrana neuronale è mantenuto dal trasporto attivo di ioni, specialmente di quelli sodio e potassio. Costantemente vi è un passaggio dello ione sodio all'esterno della membrana assicurato da un meccanismo noto come ‛pompa del sodio'. La principale sorgente dell'energia necessaria per assicurare il gradiente di membrana è rappresentata dal processo di ossidazione del glucosio. Una depolarizzazione della membrana può essere provocata da un'insufficienza relativa della pompa del sodio con conseguente accumulo di ioni sodio nell'interno del neurone. La membrana può inoltre essere depolarizzata, ripolarizzata ed iperpolarizzata dall'azione delle sinapsi eccitatorie e inibitorie (v. È a, 1; v. fig. 1). I segnali sinaptici sono trasmessi da mediatori chimici; tra essi l'aceticolina è impiegata in prevalenza dalle sinapsi eccitatorie e l'acido gamma-ammino-butirrico (GABA) è, con tutta probabilità, il mediatore delle sinapsi inibitorie. Queste inducono una iperpolarizzazione della membrana, mentre quelle eccitatorie la depolarizzano.
Gli agenti biochimici epilettogeni possono interferire su ognuno di questi meccanismi fondamentali della polarizzazione della membrana neuronale. Ciò spiega perché, ad esempio, la soglia epilettica può essere abbassata da differenti condizioni patologiche che diminuiscono l'apporto al tessuto nervoso di ossigeno o di glucosio, sostanze indispensabili per il metabolismo generatore dell'energia necessaria al trasporto attivo di ioni. In campo clinico si descrivono vari casi nei quali si verificano crisi epilettiche in conseguenza di insufficienza o arresto di circolazione ematica cerebrale, di ipossia cerebrale (eccessiva altitudine) o di ipoglicemia (terapia con insulina, adenomi del pancreas, ecc.).
La catena metabolica preposta alla sintesi del GABA sembra avere una speciale importanza nella biochimica dell'epilessia. Tale sostanza, oltre a svolgere funzioni di mediatore chimico ad effetto inibitorio, concorre al metabolismo ossidativo del glucosio, garantendo una alternativa al ciclo ossidativo di Krebs o rappresentandone un processo collaterale, e pertanto concorre alla produzione neuronale di energia. La piridossina (vitamina B6) entra nel ciclo metabolico del GABA. La grave ipovitaminosi B6 da carenza dietetica, o la cattiva utilizzazione metabolica di tale sostanza, sono talora causa di epilessia.
Fattori metabolici, farmacologici o tossici possono inoltre interferire con la sintesi, il deposito, l'attivazione e l'inattivazione dei mediatori chimici delle sinapsi favorendo di conseguenza i processi epilettici. Per esempio il diisopropilfluorofosfato (DFP) e il tetraetilpirofosfato (TEPP) bloccano l'inattivazione provocando crisi epilettiche.
Altri fattori metabolici possono direttamente influenzare il trasporto di ioni attraverso la membrana neuronale. Ciò può realizzarsi ad esempio a seguito di disordini del ricambio idrosalino che s'accompagnano ad alterazioni ormoniche e metaboliche (v. anche cap. 3, È a, 3). In particolare tutte le condizioni cliniche che inducono una ritenzione di acqua e di NaCl nell'organismo possono favorire o indurre accessi convulsivi.
Recentemente sono state offerte prove convincenti sul difettoso funzionamento della pompa del sodio nei neuroni epilettici e sulla capacità dei farmaci anticonvulsivi (specialmente dintoina e suoi derivati) nel migliorare tale funzione (v. Woodbury, 1969). Ulteriori delucidazioni dei meccanismi biochimici coinvolti nella genesi dell'epilessia sicuramente contribuiranno a far progredire la terapia farmacologica di questa malattia.
4. Diminuzione della predisposizione alle crisi, meccanismi di difesa antiepilettica. - Oltre ai meccanismi che creano e mantengono il processo epilettico, e distinti da essi, devono essere ricordati altri meccanismi che possono rientrare nella definizione di ‛meccarnsmi di difesa antiepilettica'. Di essi in realtà si conosce assai poco, ma la loro esistenza è assai probabile. Il cervello è un organo con una elevata omeostasi (capacità di mantenere uno stabile stato di equilibrio funzionale e metabolico). Numerosi meccanismi cerebrali ed extracerebrali controllano tale omeostasi. Poiché un focolaio epilettico altera tale equilibrio, vi devono pur essere alcune reazioni regolatrici.
Ciò è confermato dall'esperienza clinica. Un focolaio epilettico elettroencefalografico, anche se accompagnato da crisi elettriche, induce crisi cliniche solo in alcuni casi. Casi di epilessia postraumatica, anche con crisi cliniche, presentano una tendenza spontanea alla remissione, avendosi parallelamente un miglioramento del quadro elettroencefalografico nel quale le alterazioni vengono a circoscriversi al focolaio primario e dal quale infine possono anche scomparire.
In questo senso si può dare una spiegazione degli effetti favorevoli della terapia anticonvulsiva. In linea generale si ritiene che i farmaci antiepilettici svolgano una influenza diretta o indiretta sulla polarizzazione della membrana neuronale. La relativa specificità degli effetti dei farmaci può essere spiegata dal fatto che le membrane dei differenti neuroni risultano sensibili o accessibili in modo diverso alla loro azione. Il miglioramento della predisposizione alle crisi che si instaura nel modo indicato crea certamente condizioni favorevoli per l'intervento di più profondi meccanismi omeostatici del cervello (per gli aspetti clinici della terapia v. cap. 3, È d).
3. Aspetti clinici dell'epilessia
Ai fini della discussione che segue è sufficiente distinguere da una parte epilessie primarie e secondarie (suddivisione eziologica), dall'altra crisi generalizzate e crisi parziali (suddivisione sintomatica).
Le epilessie primarie (sinonimi: idiopatiche, genuine, essenziali, genetiche, asintomatiche, criptogenetiche, endogene) sul piano clinico sono definite: a) dall'inizio delle crisi nell'infanzia o adolescenza; b) dall'assenza di cause neonatali e postnatali che possano in qualche modo giustificarle; c) dalla negatività obiettiva neurologica e strumentale per l'esistenza di lesioni cerebrali. Le epilessie secondarie possono conseguire a una noxa cerebrale da cause extracerebrali (intossicazioni eso-endogene, ipossia, ipoglicemia, ecc.) o a lesioni cerebrali (epilessie lesionali).
Le due suddivisioni proposte (eziologica e sintomatica) non sono in alcun modo esattamente sovrapponibili. Infatti confrontando ciascun membro della prima con quelli della seconda si osserva che: 1) le epilessie primarie sono caratterizzate più spesso da crisi generalizzate, ma possono accompagnarsi anche solo a crisi parziali come nell'epilessia rolandica dell'infanzia e adolescenza (v. Y. Gastaut, 1952) o in altre epilessie focali la cui familiarità indica una ereditarietà (v. Bray e Wiser, 1965; v. Rodin e Gonzales, 1966). Una simile evenienza si può realizzare anche indirettamente sulla base di una predisposizione, come nel caso dell'epilessia del lobo temporale che consegue a convulsioni febbrili (v. Ounsted e altri, 1966); 2) le epilessie secondarie spesso sono caratterizzate da crisi parziali, ritenute espressione della localizzazione della lesione focale (epilessie lesionali), ma con altrettanta frequenza esse possono esprimersi sul piano elettroclinico soltanto con crisi generalizzate; per le epilessie lesionali ciò si realizza con il bisincronismo secondario (v. Penfield e Jasper, 1954) o con le lesioni diffuse corticosottocorticali (epilessie reticolo-corticali di Gloor; v., Generalized..., 1968) e in campo sperimentale con lesioni corticali biemisferiche (v. Marcus e altri, 1968); quanto alle epilessie sintomatiche da cause extracerebrali (intossicazioni eso-endogene, ipossia, ipoglicemia, ecc.) esse notoriamente danno luogo più spesso a crisi generalizzate.
Non è tra gli scopi del presente lavoro l'analisi completa e dettagliata delle classificazioni proposte per le epilessie. Informazioni di tal genere possono reperirsi nei testi di neurologia, elettroencefalografia ed epilettologia (v. Jasper e Kershman, 1949; v. Penfield e Jasper, 1954; v. Lennox e Lennox, 1960; v. Bates, 1963; v. Gastaut e altri, 1964; v. Janz, 1969; v. Sutherland e Tait, 1969; v. Niedermeyer, 1970).
Sebbene le classificazioni anatomo-elettrocliniche non possano essere considerate una soluzione definitiva al problema (v. È b, 4), esse si fondano su basi neurologiche indiscutibili e continuano ad essere una pietra miliare lungo il cammino delle nostre conoscenze sull'epilessia. Per questo, con il solo scopo di richiamare i quadri critici clinici che corrispondono alle varie localizzazioni cerebrali, le crisi di più frequente riscontro clinico sono state riassunte nella fig. 3. Nella fig. 2 possono essere osservati anche alcuni quadri elettroencefalografici che caratterizzano le crisi generalizzate e parziali. Tenendo presenti questi principi si può procedere a un breve esame di alcuni punti fondamentali per lo studio clinico delle epilessie.
a) Predisposizione ed ereditarietà
La predisposizione convulsiva è stata dimostrata da Amantea (v., 1920) e da Clementi (v., 1929) negli animali con il modello dell'epilessia sperimentale di Baglioni e Magnini (v., 1909) e di Baglioni e Amantea (v., 1914). Non è difficile metterla in evidenza anche nell'uomo (v. Longo, 1952). In tema di predisposizione convulsiva si pongono tre direttrici di indagine: a) il suo riconoscimento clinico; b) la sua trasmissione genetica; c) le sue implicazioni cerebrali ed extracerebrali.
1. Diagnosi di predisposizione. - Appare ormai ben documentato da una serie di ricerche che il quadro elettroencefalografico della punta-onda 3 c/s diffusa e sincrona rappresenta l'aspetto più tipico di un fattore di predisposizione epilettica. Tale quadro infatti non necessariamente esprime la malattia in atto (v. Metrakos e Metrakos, 1970). In questo stesso ambito sono da ricordare anche i quadri di poli-punta-onda diffusi e sincroni. Nei soggetti predisposti all'epilessia gli uni e gli altri possono essere presenti spontaneamente nel tracciato di veglia o attivati dal sonno, da iperpnea, da stimolazione luminosa intermittente o da ipossia indotta da inalazione di azoto.
Le alterazioni elettroencefalografiche possono rimanere infracliniche o tradursi in crisi cliniche di tipo convulsivo (grande male, attacchi mioclonici massivi) o di tipo non convulsivo (assenze, crisi amiotoniche). Le cause capaci di scatenare le crisi sono sempre di intensità molto bassa, talora comprese nell'ambito delle funzioni fisiologiche (epilessia catameniale ed epilessia al risveglio, ad esempio).
Oltre a questi casi con una obiettività subclinico-clinica, ve ne sono altri in cui la predisposizione risulta ancora ben evidente, ma certamente di grado inferiore. In tali condizioni l'EFG risulta normale o con alterazioni aspecifiche sia in stato di veglia che nel sonno. Le comuni attivazioni non inducono alterazioni apprezzabili, ma è invece presente una bassa soglia convulsiva che si può evidenziare con l'attivazione fotocardiazolica. Tali soggetti vanno incontro con facilità a convulsioni febbrili nell'infanzia, mentre nella vita adulta possono presentare crisi sporadiche in seguito ad aggressioni cerebrali di un certo rilievo (iperazotemia, eclampsia, ecc.). Del modo in cui una simile predisposizione può dar luogo ad epilessie lesionali si è già detto più sopra. Comunque una identica convulsività è stata riscontrata anche in soggetti con affezioni non epilettiche (isterismo, startie disease, fotosensitività, schizofrenia; v. Gastaut, 1969).
2. Ereditarietà della predisposizione. - Sul problema della ereditarietà delle epilessie il fronte degli studiosi è, sin dal secolo scorso, diviso tra sostenitori e oppositori. Appartengono al primo gruppo A.-L. Foville (i cui studi vennero pubblicati nel 1868), J. Grasset e Razier (1879), J. Dejerine (1866), Feré (1890) e W. R. Gowers (v., 1881); al secondo, J. H. Jackson (v., 1873) e P. Marie (1887). Il dialogo è continuato in questo secolo anche se mai alcuna delle opposte tesi è stata radicalizzata: così ad esempio Abadie (v., 1932), Marchand (v., 1952) e Penfield e Jasper (v., 1954) insistono soprattutto sul ruolo eziologico della lesione cerebrale relegando in sottordine la predisposizione, mentre Conrad (v., 1936), Nachtsheim (v., 1940), Hall (v., 1947), Longo (v., 1952) sottolineano l'importanza della predisposizione e degli elementi genetici sia nell'instaurarsi sia nell'evoluzione della sindrome.
Con l'avvento dell'elettroencefalografia e il miglioramento delle possibilità di diagnosi nei singoli casi, il problema dell'ereditarietà dell'epilessia è stato affrontato con maggior precisione soprattutto valutando l'incidenza familiare di alcune forme, quella idiopatica in particolare, gli aspetti che esse assumono in coppie di gemelli mono- ed eterocoriali e le alterazioni elettroencefalografiche infracliniche specifiche dell'epilessia riscontrabili in un alto numero di familiari degli epilettici (per una rassegna bibliografica, v. Gastaut e altri, 1969). Ai dati così acquisiti hanno fatto seguito considerazioni statistiche entro gruppi di forme particolari di epilessia e tra famiglie di epilettici e famiglie di non epilettici (v. Sorel, 1969).
Studi e considerazioni di questo genere, che non possono essere qui riferiti per esteso, hanno definitivamente fatto luce sull'esistenza di una trasmissione ereditaria della predisposizione all'epilessia nell'uomo. Parallelamente essa è stata dimostrata anche in alcune specie animali (Nachtsheim, Hall). Negli ultimi anni solo Alström (v., 1950) si è opposto a tale conclusione, ma la sua ricerca non appare convincente anche se riesce apprezzabile il suo sforzo di non rinfrancare prevenzioni e preclusioni sociali verso gli epilettici. Le acquisizioni teoriche devono ovviamente servire a far comprendere meglio la malattia senza danneggiare i malati; inoltre altro è parlare di ereditarietà, altro è stabilire il rischio reale di progenie epilettica per il singolo caso di epilessia. Sorel, per esempio, ha calcolato che tale rischio è valido solo per una parte delle epilessie genuine che, a loro volta, rappresentano il 7,5% di tutte le epilessie. I casi di epilessia genuina che hanno un rischio effettivamente superiore di progenie epilettica rispetto alla popolazione generale sono quelli con altri casi di epilessia nella famiglia, in particolare fratelli.
Qualche discordanza, nell'ambito della riconosciuta ereditarietà di una predisposizione all'epilessia, esiste a proposito della natura di tale predisposizione e delle forme di epilessia nelle quali essa risulta operante. H. Gastaut (v., 1969), per esempio, distingue una predisposizione all'epilessia e una predisposizione convulsiva. La prima sarebbe espressa dai quadri elettroencefalografici e clinici sopra descritti: punta- e polipunta-onda diffuse e sincrone, grande male, mioclonie massive, assenze, amiotonie, crisi tutte provocate da noxae cerebrali minime e anche da situazioni fisiologiche. Solo per essa, molto rara in realtà, può essere invocata una trasmissione monogenetica, cioè un'eredità monomerica a penetranza irregolare quale quella stabilita da J. D. e K. Metrakos (v., 1960, 1961 e 1970).
Le indagini di questi autori, condotte in famiglie di epilettici con crisi generalizzate, hanno infatti dato prove convincenti sull'esistenza di un gene responsabile della trasmissione delle scariche generalizzate intercritiche presenti in molti membri di tali famiglie; si tratterebbe di un gene autosomico la cui espressione clinica varia con l'età, essendo molto debole alla nascita, quasi completa tra i 4 e i 6 anni e di nuovo via via più debole negli anni successivi.
A tale predisposizione epilettica Gastaut contrappone la predisposizione convulsiva (clinicamente espressa da convulsioni febbrili o da altre valide aggressioni cerebrali e da bassa soglia convulsiva). Quest'ultima sarebbe espressione di una eredità multifattoriale, corrispondente a una semplice variazione di un carattere normale (un soggetto, in altre parole, sarebbe più o meno predisposto alla convulsione, nello stesso modo in cui è più o meno alto o più o meno intelligente). In tal modo si nega che possa esistere una ereditarietà per le epilessie focali, con la sola eccezione di quelle sindromi neurologiche ereditarie che conducono a lesioni cerebrali epilettogene (malattia di Crouzon, di Sturge-Weber, ecc.) nelle quali evidentemente l'epilessia è solo epifenomeno. Le casistiche che riportano una familiarità per epilessie lesionali (v. Bray e Wiser, 1965; v. Rodin e Gonzales, 1966) devono essere spiegate alla luce delle lesioni focali che possono essere provocate da una semplice predisposizione convulsiva.
Recentemente J. D. e K. Metrakos (v., 1970) hanno insistito sul fatto che un fattore genetico di predisposizione è il comune denominatore sia delle epilessie cosiddette centroencefaliche, sia delle convulsioni febbrili e sia infine dei casi di epilessia focale. Se ciò è vero, allora gli aspetti subclinici e clinici, distinti da Gastaut in predisposizione epilettica e predisposizione convulsiva, più verosimilmente sarebbero espressione solo quantitativamente diversa dello stesso fattore genetico. È inoltre plausibile che quest'ultimo, nel suo estrinsecarsi, risulti evidente clinicamente in modo maggiore o minore a seconda del suo differente grado di penetranza e della sua interazione con l'intero genotipo e con le stesse condizioni ambientali che su di esso agiscono.
3. Implicazioni umorali della predisposizione. - Ricerche in questa direzione cominciarono in Italia (v. Longo, 1952) con lo studio endocrino e umorale di animali che dimostravano una predisposizione epilettica con il metodo di Baglioni e Amantea. Successivamente le segnalazioni di modificazioni endocrine, umorali e metaboliche nell'epilessia umana sono divenute così numerose che non è pensabile di poterle qui riassumere e nemmeno semplicemente citare. Talora esse rappresentano acquisizioni cliniche di notevole interesse anche per le implicazioni biochimiche ed ereditarie (come per es. i quadri di epilessia fenilpiruvica, o le epilessie sensibili al trattamento piridossinico), tuttavia nel loro insieme esse rappresentano una enorme congerie di segnalazioni dalle quali riesce difficile trarre un filo conduttore o una ricostruzione che direttamente conduca all'intima comprensione della eziopatogenesi della crisi epilettica. Né è facile stabilire quanto sia da attribuirsi a fattori ereditari, quanto a fattori acquisiti e quanto infine sia epifenomeno della stessa epilessia e della terapia prolungata che essa impone.
Con riferimento a quanto detto nel paragrafo relativo alla biochimica dell'epilessia (v. cap. 2, È b, 3) è evidente che i molteplici processi che conducono alla crisi epilettica finiscono per incidere sulla membrana cellulare e sulla sua trasmissione. Ne consegue che anche l'elemento genetico responsabile delle epilessie nel senso più sopra indicato deve trovare il suo ultimo anello eziopatogenetico a tale livello. Per questo molto interessante appare il recente contributo di McKhann e Shooter (v., 1969) anche come importante ipotesi di lavoro per future ricerche: essi hanno passato in rassegna vari modelli mediante i quali fattori genetici potrebbero indurre modificazioni proteiche capaci a loro volta di alterare la soglia di trasmissione delle membrane neuronali. Sfortunatamente una conoscenza biochimico-genetica di tal natura è ben lungi dall'essere completa nei suoi dettagli, almeno per ciò che concerne il neurone.
b) Correlazioni anatomo-elettrocliniche.
Un intenso lavoro di selezione elettroclinica e anatomo-elettroclinica (v. Hill, 1953 e 1963) e ricerche e osservazioni su materiale di diagnostica preoperatoria in neurochirurgia (v. Hill, 1953; v. Brazier, 1956; v. Bickford, 1956; v. Walker e Ribstein, 1957; v. Angeleri e altri, 1961 e 1964; v. Bancaud e altri, 1965) hanno consentito di acquisire un patrimonio di conoscenze diagnostiche di notevole valore pratico. Non va taciuto che a una simile ricostruzione anatomo-elettroclinica hanno concorso ricerche anatomo-fisiopatologiche e soprattutto i risultati tratti da numerosi modelli di epilessia sperimentale (v. Gastaut e Fischer-Williams, 1959). Anzi, dal continuo confronto dei dati sperimentali con le osservazioni cliniche sono derivati alcuni concetti fondamentali per la comprensione stessa dei fenomeni osservati (v. figg. 2 e 3).
1. Il focolaio epilettico. - Tralasciamo di parlare dei focolai funzionali e delle modificazioni stabili che in essi si possono verificare (v. cap. 2, È b, 1) nè ci soffermiamo sulle implicazioni istopatologiche, neurochimiche e fisiopatologiche a livello neuronale relative ai focolai lesionali (v. cap. 2, È a, 2). Da un punto di vista clinico per questi ultimi è importante prendere in considerazione i seguenti elementi: a) la causa che li produce; b) la loro topografia ed estensione; c) le tecniche e la diagnostica idonee a porli in evidenza nel malato; d) l'evoluzione in senso epilettico del focolaio organico (intervallo tra lesione e insorgenza di scariche elettroencefalografiche intercritiche o di crisi elettrocliniche, frequenza delle crisi, scatenamento di esse, ingravescenza o remissione nel tempo).
I focolai epilettogeni delle forme lesionali di epilessia umana sono rappresentati da danni tissutali localizzati o diffusi, primitivi o secondari alla stessa epilessia, di origine degenerativa, tossica, ischemica, infiammatoria, neoplastica, traumatica o parassitaria. È evidente che molte delle affezioni comprese nei gruppi di tale elenco rappresentano, indipendentemente dall'epilessia cui possono dare origine, malattie neuropsichiatriche così importanti da far passare in secondo piano la stessa epilessia.
In questi casi, ai fini clinici, è essenziale sapere non solo che le epilessie molto spesso (sempre nei casi di epilessia tardiva) sono espressione di altra affezione neurologica, ma sapere anche, con buona approssimazione, per quali è frequentemente sintomo inaugurale o tardivo, quando è segno localizzatorio, quando non lo è. Utili in questo senso sono i contributi clinico-statistici sul differente grado di convulsività dei vari tipi di processi espansivi endocranici e in particolare delle differenti neoplasie cerebrali (v. Penfield e Jasper, 1954).
I focolai che più frequentemente stanno alla base delle forme di epilessia in cui essa è disturbo predominante o esclusivo sono le sequele cerebrali da traumi, ischemie o flogosi. Penfield descrive numerosi quadri anatomo-patologici che o sono già presenti (malformazioni) o si realizzano alla nascita oppure sono sequele di molte affezioni endocraniche che hanno rappresentato una malattia preminente dell'infanzia, dell'adolescenza e della vita adulta.
La latenza tra l'instaurarsi della lesione e la comparsa delle crisi epilettiche varia in rapporto a molti fattori. Talora essa dura molti anni e dipende anche dalla convulsività dei settori cerebrali coinvolti dai focolai anatomici del tipo sopra descritto. È noto che la soglia convulsiva aumenta progressivamente a partire dalle strutture archi-paleopalliali, passando a quelle neocorticali associative e motorie sino a quelle sottocorticali (v. Ajmone-Marsan, A newly proposed..., 1965). Questa differente suscettibilità epilettica zonale è condizionata dalla citoarchitettonica (v. cap. 2, È a, 1) e aggravata da particolari condizioni di angiotettonica (per es., l'ippocampo risente precocemente di alterazioni anche non gravi del circolo cerebrale).
D'altra parte il concetto di focolaio organico è accettabile, a livello fisiopatologico, solo se ad esso viene attribuita una terza dimensione. È noto che una cicatrice epilettogena di un'area corticale specifica necessariamente coinvolge i circuiti talamo-cortico-talamici corrispondenti e che crisi ippocampali sempre propagano all'amigdala ipsilaterale (v. Angeleri e altri, 1961). Tuttavia su questi sistemi, precocemente interessati a causa delle proiezioni del focus, agiscono altri meccanismi cerebrali non direttamente coinvolti dalle scariche epilettiche. Un esempio in questo senso è offerto dallo studio delle scariche elettrocliniche nella corteccia motoria rolandica quando si registrino i potenziali negli strati corticali a differente profondità e si valutino i quadri elettrici e quelli clinici nella veglia e in narcosi (v. Penfield e Jasper, 1954). Simili dati sottintendono implicazioni relative a importanti problemi fisiopatologici, quali la propagazione e/o generalizzazione delle scariche, i loro meccanismi di arresto e le loro correlazioni elettrocliniche (v. cap. 2, È a, 5 e È b, 4).
Infine la funzione e le connessioni di una determinata area cerebrale sono fondamentali al quadro clinico finale che conseguirà dopo l'insorgenza di un focolaio epilettogeno in tale sede. È noto, per esempio, che lesioni epilettogene della corteccia interemisferica (aree limbiche anteriori) conducono con estrema facilità a un bisincronismo secondario (v. Penfield e Jasper, 1954).
2. Crisi generalizzate (bisincronismo primario e secondario). - L'ipotesi di Penfield e Jasper (1954) formulata sui risultati sperimentali di Jasper e Drogleever Fortuyn (v., 1946), ampiamente confermata successivamente (v. Pollen, 1968), propone l'esistenza di un pacemaker talamico e reticolare (sistema talamico aspecifico e formazione reticolare ascendente del tronco encefalico - ARAS) responsabile della generalizzazione simultanea della crisi all'intero encefalo in alcune forme di epilessia (epilessie centroencefaliche) le cui scariche avrebbero inizio in tali strutture (bisincronismo primario) e si generalizzerebbero per le caratteristiche funzionali intrinseche ad esse (sistema a proiezione diffusa). In particolare i nuclei aspecifici del talamo sono interessati nella genesi dei complessi punta-onda a differenza dell'ARAS, che può essere importante nel controllare l'attività dei nuclei talamici aspecifici, ma non è in grado di evocare nè complessi punta-onda nè fenomeni di reclutamento bilaterali e sincroni (Pollen; v. anche cap. 2, È a, 3).
Casi di epilessia focale delle regioni corticali mediali interessanti le aree limbiche realizzano precocemente quadri elettroencefalografici con potenziali punta-onda diffusa e sincrona che spesso mascherano l'attività intercritica e critica focale. Questo interessamento secondario del sistema centroencefalico è stato indicato come bisincronismo secondario (v. Tückel e Jasper, 1952). Esso può essere invocato non solo a spiegazione dei casi nei quali vi è una evoluzione stabile verso una forma generalizzata, tale sovente da mascherare la focalità, ma anche per quelli assai più numerosi in cui i due tipi di crisi e di alterazioni elettroencefalografiche (focali e generalizzate) si associano, nonché per le crisi parziali a più o meno rapida successiva generalizzazione.
Contro l'ipotesi di Jasper e Penfield stanno altri risultati sperimentali. Quadri punta-onda diffusa e sincrona sono stati evocati con vari tipi di stimolazione, anche dopo distruzione del sistema talamico aspecifico o dopo completa talamotomia (v. Angeleri, 1958; v. Marcus e Watson, 1964; v. Servít e altri, 1965). Lesioni corticali biemisferiche (v. Marcus e Watson, Bilateral..., 1966; v. Marcus e altri, 1968) o sottocorticali multiple nella scimmia (v. Udvarhelyi e Walker, 1965) sono sufficienti a produrre alterazioni elettroencefalografiche di tipo epilettico diffuse e sincrone; tale sincronismo è mantenuto dall'integrità delle commissure interemisferiche. A ciò si aggiunge che con la stimolazione del talamo aspecifico non è possibile evocare scariche epilettiche autointrattenute (v. Ralston e Ajmone-Marsan, 1956).
In campo clinico altre obiezioni all'ipotesi del pacemaker talamico derivano dalle osservazioni stereoelettroencefalografiche in casi di epilessia generalizzata primaria. In essi sono stati dimostrati focolai frontali profondi (v. Bickford, 1956; v. Bancaud e altri, 1965) ippocampali e limbici (v. Angeleri e altri, 1961 e 1964; v. Rossi e altri, 1968), corticali e sottocorticali diffusi (v. Walker e Marshall, 1964). In casi di epilessia dello stesso tipo, la stimolazione cadenzata di nuclei talamici aspecifici non ha mai evocato scariche diffuse e sincrone; talora invece è stata in grado di interromperle (v. Angeleri e altri, 1961 e 1964; v. Bancaud e altri, 1965). Su questa linea altre obiezioni di carattere clinico vengono avanzate: in primo luogo l'assenza della massa intermedia talamica così spesso documentabile con la pneumoencefalografia proprio negli epilettici con crisi generalizzate e inoltre il fatto che nessuna affezione del talamo ha mai prodotto crisi epilettiche generalizzate (v. Williams, 1965).
Interessanti a questo punto le conclusioni di Gloor (v., 1968) derivate da una serie di ricerche istopatologiche su casi di encefalopatie diffuse caratterizzate da anormalità elettroencefalografiche bilaterali e sincrone (anche se non necessariamente tipo punta-onda) e dallo studio di casi di epilessia con scariche diffuse nei quali si era proceduto all'introduzione endocarotidea o endovertebrale di Amytal sodico o di convulsivanti. Le conclusioni di Gloor sembrano superare la contradditorietà delle osservazioni sperimentali e cliniche più sopra riferite. Da esse scaturisce come nuova ipotesi un controllo cortico-sottocorticale (in particolare delle strutture talamiche aspecifiche e dell'ARAS) dell'elettrogenesi cerebrale. Una rottura di tale integrazione o bilanciamento cortico-sottocorticale produrrebbe i quadri elettroencefalografici e clinici di epilessia con crisi generalizzate che Gloor denomina epilessie ‛cortico-reticolari', le quali comprendono sia forme primitive sia secondarie (v. fig. 4).
3. Scariche e crisi epilettiche nell'infanzia e adolescenza. - Alla nascita e per tutto il periodo evolutivo la sintomatologia epilettica di ordine clinico e i quadri elettroencefalografici sono condizionati dal grado di maturazione cerebrale. Nei primi mesi di vita ad esempio non si osservano crisi tonico-cloniche, assenze del piccolo male, automatismi del lobo temporale né attacchi di tipo jacksoniano. Nell'uomo, a differenza di quanto si verifica in altri Mammiferi, il cervello del neonato manca ancora delle funzioni che condizionano la propagazione intraemisferica e intracorticale, manca ancora il bisincronismo interemisferico. Dopo i primi mesi di vita vi e una tendenza alla sincronizzazione dei bioritmi cerebrali con un accresciuto potere di diffusione delle scariche epilettiche (v. Angeleri e altri, 1958; v. Massa e Niedermeyer, 1968). Espressione elettroencefalografica dell'immaturità anatomo-funzionale sono la tendenza alla sincronizzazione dei potenziali cerebrali (quindi una maggiore convulsività), una più facile propagazione e diffusione delle scariche epilettiche e una loro minore stabilità nei circuiti nervosi (v. Gibbs e altri, 1954; v. Kellaway, 1954; v. Pinelli, 1954). In definitiva da tali caratteristiche e dalla capacità dei centri nervosi di organizzare certe risposte conseguono: a) quadri elettroclinici particolari dell'epilessia dell'infanzia; b) certe età obbligate per l'insorgenza di crisi epilettiche del tipo osservabile nell'adulto; c) certe dissociazioni tra quadri elettroencefalografici e sintomatologia clinica.
Nel neonato e nei primissimi mesi di vita le crisi più frequenti sono di tipo clonico; iniziano di solito in un'emifaccia o in un arto, si diffondono poi ad altre regioni ipsilaterali, presentano il fenomeno di waxing and waning e non sembrano determinare una compromissione della vigilanza del bambino.
Più rare sono le crisi emiconvulsive che divengono invece dominanti dopo i primi mesi. Determinano uno spasmo adversivo della testa e degli occhi contemporaneo all'interessamento di un emicorpo che può essere alternativamente di un lato o dell'altro. Raramente le crisi focali sottendono un processo patologico cerebrale circoscritto, sono invece spesso espressione di condizioni dismetaboliche (ipocalcemia, ipomagnesemia, ipo- e ipernatriemia, deficienza o dipendenza piridossinica, ipoglicemia, amminoaciduria). Le crisi emigeneralizzate, emicloniche, più raramente emitoniche, specie se a ricorrenza subentrante o in forma di stato di male, a causa dell'ipossia ed edema cerebrale che inducono determinano una lesione ischemica secondaria: si inizia con emiconvulsioni, si finisce con emiplegia ed epilessia (sindrome H.H.E. di H. Gastaut e altri; v., 1959-1960).
Altre crisi caratteristiche del periodo neonatale e dei primissimi mesi di vita sono le scariche erratiche o migratorie che investono settori limitati degli emisferi trasferendosi da una regione all'altra e determinando movimenti clonici che interessano ora un arto ora l'altro anche in differenti lati del corpo; se la migrazione è rapida con successivo interessamento dei 4 arti, può essere simulato un accesso generalizzato. In questo stesso periodo possono ricorrere anche crisi ‛minime', caratterizzate cioè da brevi periodi di apnea associata a lievi contrazioni toniche o a tremori di uno o più arti, o a brevi deviazioni degli occhi con scialorrea. L'indagine elettroencefalografica è di fondamentale aiuto nella diagnosi dei quattro tipi di crisi descritte e serve anche dal punto di vista prognostico. Si osservano quadri elettroencefalografici multifocali, tracciati periodici o parossistici e tracciati piatti. Questi ultimi insieme alle crisi ‛minime' sono di significato prognostico gravissimo per la stessa vita del piccolo o per il suo successivo sviluppo psicomotorio.
Il quadro elettroclinico dell'ipsiaritmia (v. Gastaut e altri, 1964), che inizia nei primi 6 mesi di vita, dà luogo a mioclonie massive e brevi ed evolve sfavorevolmente se non trattato. Una possibile evoluzione epilettica può essere la sindrome di Lennox (v. Lennox e Davis, 1950) per la quale però si prospettano cause molteplici. Essa è considerata come esempio tipico di bisincronismo secondario, insorge tra il primo e il sesto anno di età, dà luogo ad assenze atipiche (con automatismo) e a crisi generalizzate toniche per lo più notturne. La sua prognosi è sfavorevole sia per l'evoluzione epilettica, sia per la grave compromissione psicomotoria. Favorevole invece è il decorso dell'epilessia rolandica: insorge a 7-8 anni, recede alla pubertà e si manifesta con focalità intercritica rolandica e crisi parziali che intervengono specie nel sonno (v. Y. Gastaut, 1952; v. Bancaud e altri, 1958; v. Nayrac e Beaussart, 1958).
Anche il piccolo male tipico è una forma caratteristica dell'infanzia; scariche organizzate da complessi punta-onda a 3 c/s diffusi e sincroni non sono per altro osservabili prima dei tre anni. Spesso esse sono infracliniche; l'età in cui più frequentemente insorgono assenze è quella scolare.
Le convulsioni febbrili dell'infanzia ricorrono in una percentuale elevata di bambini (3-7%) tra i 6 mesi e i 5 anni, con massima incidenza tra i 9 e i 20 mesi. Hanno carattere ereditario ed evoluzione differente dall'epilessia, dalla quale vanno tenute distinte, pur rappresentando una condizione che la favorisce, di solito creando una condizione epilettogena stabile (v. Ounsted ed altri, 1966). Al momento della prima o delle prime crisi convulsive febbrili non esistono criteri validi di previsione per quei casi destinati a trasformarsi nella sindrome più grave.
Le crisi caratteristiche dell'adulto non sono state osservate prima di una certa età. Così le crisi di grande male, nella loro caratteristica successione tonica e clonica, fanno la loro comparsa solo dopo i 3-4 anni; precedentemente si hanno crisi o toniche o cloniche generalizzate o emigeneralizzate. Ancor più tardiva è la comparsa di crisi jacksoniane (intorno ai 7-9 anni). La sintomatologia delle crisi psicomotorie e del lobo temporale è scarsa e poco elaborata nella prima infanzia, assumendo le sue caratteristiche parallelamente al processo della maturazione anatomo-funzionale del cervello.
Una terza espressione elettroclinica del particolare substrato anatomo-funzionale del cervello in maturazione è rappresentata dalla discordanza tra sintomi accessuali e quadri elettroencefalografici e dalla migrazione dei foci epilettici descritta per la prima volta da Gibbs e altri (v., 1954). Le dissociazioni elettrocliniche sono assai frequenti: crisi generalizzate in alcuni casi presentano un tracciato intercritico focale e viceversa. La fig. 5 ne mostra un esempio.
4. Limiti del criterio localizzatorio anatomo-elettroclinico. - I dati clinici e fisiopatologici sino ad oggi acquisiti sull'epilessia si basano anche sulla conoscenza dei limiti dei mezzi di indagine con cui sono stati raggiunti e delle ipotesi formulate su essi. Tali aspetti negativi sono d'altronde stimolanti per perfezionare tecniche e metodiche e per formulare nuove ipotesi di lavoro.
Pochi esempi sono forse sufficienti a dimostrare la complessità del problema. Un primo gruppo di limitazioni nel campo della ricerca clinica può essere considerato di ordine tecnico. Ciò che offre l'elettroencefalografia di superficie è una rappresentazione parziale e distorta dell'attività elettrica cerebrale (v. Abraham e Ajmone-Marsan, 1958). Non solo scariche intercritiche parossistiche possono non essere raccolte dalla registrazione dal cuoio capelluto, ma anche crisi cliniche possono mancare di un correlato elettrico o perché investono un settore corticale troppo ristretto o perché si svolgono prevalentemente nel polo profondo del focolaio. La registrazione dal cuoio capelluto raccoglie solo una parte dell'attività elettrica cerebrale, essendo fuori della sua portata la maggior parte della sostanza grigia. Infine, la lettura visiva degli stessi tracciati elettroencefalografici consente di estrarre solo una minima parte delle informazioni che essi contengono.
Alcuni di questi limiti sono superabili in condizioni particolari. In campo clinico solo in caso di interventi chirurgici è consentito di registrare direttamente dalla corteccia cerebrale o da strutture grigie profonde (corticografia e stereoelettroencefalografia), mentre con attrezzature elettroniche più o meno complesse si cerca di superare i limiti della semplice lettura visiva dei tracciati (analizzatore di frequenza di Gray-Walter, toposcopio di Petsche o di Rémond, ecc.; v. Storm van Leeuwen e Magnus, 1961).
Trattandosi di condizioni privilegiate, rimane il fatto che il maggior numero di informazioni sull'epilessia dell'uomo proviene da registrazioni dal cuoio capelluto; su di esse si è spesso costretti a risolvere i problemi diagnostici del singolo caso e su di esse sono spesso basate considerazioni di ordine generale. Ciò naturalmente non esclude il costante, talora implicito, confronto di tali informazioni con i dati più completi offerti dalle metodiche cliniche avanzate, nè la loro valutazione critica alla luce di quanto offrono i modelli dell'epilessia sperimentale.
Anche la metodologia impiegata nelle registrazioni elettroencefalografiche sottintende implicazioni non sempre chiare. È noto, ad esempio, che le scariche epilettiche critiche e intercritiche possono essere attivate o deattivate in modo diverso nella veglia e nelle differenti fasi del sonno. Ciò può dipendere dal tipo e distribuzione delle scariche, ma anche dalla topografia del focolaio. Per contro, veglia e fasi del sonno possono a loro volta modificare morfologia e distribuzione delle scariche intercritiche e delle stesse crisi elettrocliniche (v. Angeleri, 1968). Tutto ciò dimostra la stretta compenetrazione tra dinamica dei processi epilettici e meccanismi cerebrali. Siccome tali interdipendenze sono lungi dall'essere chiare, le interpretazioni dei differenti quadri elettroencefalografici rimangono spesso ipotetiche.
Considerazioni in parte analoghe valgono per i risultati che si ottengono con le metodiche speciali di attivazione, tra le quali più comuni quelle con farmaci convulsivanti (bemegride, metrazol, ecc.). La mancanza, o quasi, di inconvenienti (se eseguite lege artis) e la rapidità con cui si ottengono le informazioni elettrocliniche che dovrebbero servire alla diagnosi hanno consentito un largo impiego di tali metodiche. Tuttavia già da tempo si è sottolineato che con esse non è possibile andare molto oltre la semplice diagnosi di epilessia. Esaltando globalmente la convulsività cerebrale, ma in particolare quella di alcune strutture, si deforma notevolmente la realtà clinica del singolo caso. Trarre conclusioni particolari o generali da tali registrazioni è pertanto erroneo (per ulteriori dati e bibliografia, v. Andrioli e altri, 1966). Secondo l'esperienza di chi scrive, con le attivazioni da convulsivanti non solo si verifica un mascheramento delle crisi focali per una più o meno precoce generalizzazione (v. figg. 6 e 7), ma anche, nel singolo caso, si realizza un polimorfismo elettroclinico che non corrisponde a quello delle crisi spontanee.
Un terzo gruppo di limiti alla valutazione dei dati elettroencefalografici investe l'aspetto teorico del problema; al di là dell'elettroencefalografia tale aspetto coinvolge lo stesso problema patogenetico dei processi epilettici. In primo luogo deve essere citato il dubbio significato localizzatorio delle scariche elettriche. Sia in campo clinico (v. Ajmone-Marsan e Lewis, 1960; v. Scherman e Abraham, 1963; v. Madsen e Bray, 1966) sia in campo sperimentale (v. Ralston, 1961) è dimostrato che ristrette lesioni organiche epilettogene possono manifestarsi con alterazioni parossistiche diffuse, senza alcun segno localizzatorio o di lateralizzazione. Parimenti l'‛attivazione a distanza', dimostrata sperimentalmente e ricorrente anche in clinica (possono essere ricordati a titolo di esempio certi quadri di attivazione epilettica della corteccia del lobo temporale per foci rinencefalici), rende talora impossibile stabilire se una certa scarica è realmente fenomeno locale o indotto a distanza da un processo generatosi altrove. Infine, un'area potenzialmente epilettogena può mancare di manifestarsi con segni elettrici tipici, mentre inequivocabili scariche epilettiche non sempre corrispondono a un processo epilettico locale, almeno a un processo che necessariamente si manifesterà con crisi cliniche (v. i contributi di Ajmone-Marsan, 1965).
Altro aspetto importante è rappresentato dalle correlazioni tra scariche elettriche e sintomatologia clinica della crisi. Attivazione, inibizione o interferenza ed esaurimento dell'area investita dalla scarica epilettica sono termini che non hanno ancora una interpretazione fisiopatologica soddisfacente e non spiegano perché si verifichi ora l'uno ora l'altro effetto (attivazione o inibizione) per scariche elettriche apparentemente identiche, anche se è noto che certe regioni prediligono l'uno piuttosto che l'altro fenomeno. Inoltre per certe regioni, come il lobo temporale, riesce assai difficile stabilire quanto della sintomatologia clinica dipenda da attivazione di alcuni settori o da loro liberazione (in senso jacksoniano) per inibizione critica o estinzione postcritica di altri.
Sotto tutti questi punti insoluti deve necessariamente nascondersi la spiegazione delle numerose apparenti contraddizioni tra dati elettroencefalografici, clinici e localizzazioni anatomiche: a) perché, ad esempio, si possano osservare crisi cliniche parziali e concomitanti scariche critiche elettriche generalizzate; b) perché certi epilettici abbiano solo crisi generalizzate pur presentando quadri elettroencefalografici esclusivamente focali nei periodi intercritici; c) perché l'asportazione chirurgica del focolaio non sia sempre seguita dalla scomparsa delle crisi; d) perché vi siano soggetti con quadri elettroencefalografici differenti nel tempo, ora solo focali ora esclusivamente generalizzati (v. Strobos e Kavallinis, 1968); e) perché addirittura si possa realizzare una migrazione delle scariche, come dimostrato dai Gibbs, nell'infanzia.
In realtà l'insieme di queste osservazioni non indica soltanto i limiti dell'indagine elettroencefalografica, ma pone in discussione anche il problema dei rapporti tra focolaio lesionale, scariche elettroencefalografiche e sintomatologia critica. Senza rinunciare alle acquisizioni sin qui raggiunte, è compito dell'indagine futura rivedere tali correlazioni per dar loro un'interpretazione più valida alla luce di quanto oggi appare contradditorio, ma che in realtà potrebbe essere la chiave per la soluzione dell'intero problema.
c) Epilessie a scatenamento o ad arresto sensoriale.
Il modello di epilessia sperimentale di Amantea e Clementi lascia sospettare un ruolo importante delle afferenze sensitivo-sensoriali nello scatenamento delle crisi nel focolaio epilettico (v. anche cap. 2, È a, 2). La risposta epilettica ottenibile in alcune specie di animali con determinati stimoli, ad esempio la crisi audiogena del ratto albino e la risposta fotomioclonica del babbuino (Papio papio) riscontrate in un alto numero di adolescenti e giovani soggetti (v. Killam e altri, 1966), dimostrano che vi è anche una predisposizione di specie.
Quanto all'epilessia umana occorre subito precisare che, per ciò che concerne le epilessie ‛riflesse' da abnormi stimolazioni periferiche o midollari, la casistica è andata riducendosi con il passare degli anni e con il perfezionarsi della diagnostica, tanto che oggi può esserne messa in dubbio l'esistenza (v. Tassinari, 1968). Il Tassinari sottolinea invece che si sono moltiplicate le segnalazioni di epilessia con crisi a scatenamento sensitivo-sensoriale o emozionale. Le innumerevoli varietà di situazioni-stimolo possono essere raggruppate come segue: a) crisi epilettiche provocate da situazioni stimolo complesse (leggere, ascoltare una musica, fare un calcolo, ecc.); b) crisi provocate da stimolazioni sensoriali e propriocettive indipendenti da ogni reazione di sorpresa; c) crisi scatenate dagli stessi stimoli sensitivo-sensoriali, ma in quanto realizzano una reazione di sorpresa; d) crisi di coreo-atetosi parossistica a scatenamento emotivo-sensoriale la cui natura epilettica per altro è posta in dubbio.
Altrettanto numerose, anche se non così varie, sono le situazioni-stimolo capaci di arrestare una crisi al suo inizio (v. anche cap. 2, È a, 2).
Un dato molto importante ai fini della valutazione del ruolo eziopatogenetico degli stimoli è l'osservazione clinica che la maggioranza delle epilessie cosiddette ‛riflesse' sono di tipo generalizzato. Ciò dimostra che non vi è corrispondenza tra settore cerebrale raggiunto dalle afferenze e inizio delle crisi.
Questo fatto, insieme a quello che nega la possibile genesi di una epilessia da stimolazioni afferenti abnormi (se esistono, tali casi sono rarissimi; la letteratura non possiede sicure dimostrazioni in tale senso), dimostra che il meccanismo di scatenamento sensitivo-sensoriale ed emozionale nell'epilessia umana deve essere relegato nella sua giusta posizione nel lungo elenco di fattori intrinseci ed estrinseci capaci di un'attivazione critica di un focus epilettico. H. Gastaut (v., 1963) enumera i seguenti fattori estrinseci: a) quelli che modificano l'eccitabilità corticale (modificazioni del circolo cerebrale con conseguente ischemia e ipossia, stati tossici esogeni o endogeni o fattori ormonali, stati funzionali del cervello quali sincronizzazione o desincronizzazione dei suoi potenziali elettrici); b) quelli che attivano il focus aumentandone l'eccitabilità (impulsi di origine centrale convergenti sul focus, impulsi afferenti provenienti dalla periferia che raggiungono il focus attraverso le vie specifiche o che agiscono tramite strutture sottocorticali, tra cui quelle aspecifiche). Oltre ai fattori estrinseci ve ne sono molti altri intrinseci al focolaio che vengono omessi per brevità. Complessivamente dunque lo scatenamento sensitivo-sensoriale è solo un aspetto parziale del complesso meccanismo che può entrare in gioco nello scatenamento della crisi. Ciò nonostante esso rappresenta un modello clinico di epilessia interessante per le sue implicazioni fisiopatologiche e per i suoi possibili accostamenti con analoghi modelli di epilessia sperimentale.
d) Terapia dell'epilessia.
Con riferimento all'epilessia come processo sindromico, le indicazioni terapeutiche di ordine chirurgico o medico richiedono una diagnosi eziologica e localizzatoria quanto più esatta possibile e una valutazione della gravità e della frequenza delle crisi.
Sebbene i primi tentativi razionali di interventi chirurgici in casi di epilessia (riservati a quella postraumatica) risalgano alla seconda metà del secolo scorso (v. Horsley, 1887-1888), la moderna neurochirurgia si è sviluppata negli ultimi quarant'anni con le sue équipes neurofisiologiche e neuroradiologiche preposte alla diagnosi pre- e intraoperatoria (v. Bates, 1963).
Alcuni interventi hanno subito ormai il vaglio dei controlli postoperatori su casistiche abbastanza ampie. Essi sono rappresentati da topectomie corticali su cicatrici meningo-cerebrali o su focolai atrofici, da lobectomie temporali o da ippocampectomie in casi di epilessia del lobo temporale e infine da emisferectomie in bambini o giovani con cerebropatia lateralizzata ed estesa a un emisfero (v. Walker, 1965; v. Rasmussen, 1969).
Altri interventi sono ancora in una fase interlocutoria, anche se fondati su interessanti ipotesi teoriche (né deve essere trascurato il notevole contributo che hanno dato alla fisiopatologia delle epilessie). A titolo di esempio ricordiamo alcuni interventi che sono stati definiti di ‛neurochirurgia funzionale' come la coagulazione di alcuni nuclei talamici aspecifici (v. Spiegel e Wycis, 1950), della amigdala temporale (v. Angeleri e altri, 1961) e di commissurotomia interemisferica (v. Sperry, 1966). Tuttavia i loro risultati terapeutici restano incerti.
I vari tipi di intervento non possono essere discussi in questa sede. I risultati che sono stati riferiti sono condizionati da numerosi fattori, tra i quali in primo luogo: a) la scelta dei malati; b) la definizione fisiopatologica che di ciascuno di essi è stata data. Bancaud e Talairach (v., 1970) insistono in particolare su quest'ultimo punto. Solo un adeguato studio spazio-temporale delle scariche epilettiche, eseguito mediante stereoelettroencefalografia, può dare precise informazioni sulla dinamica del focolaio e quindi consentire di compiere - se possibile - exeresi razionali, capaci cioè di interrompere le vie principali del traffico epilettico. Prima ancora però dello studio stereoelettroencefalografico, che deve essere considerato già un tempo operatorio, si impone un'accurata scelta dei casi sulla base di indagini preoperatorie assai complesse (v. Andrioli e altri, 1966).
Sta di fatto che anche nei casi di maggior successo chirurgico non può essere abbandonato un successivo discreto trattamento farmacologico anticonvulsivo. Pertanto l'indicazione chirurgica nel trattamento dell'epilessia deve seguire sempre quella medica comprendente regole igieniche, internistiche, psicoterapiche e infine l'impiego di farmaci anticonvulsivi o antiepilettici. Accenniamo solo a questi ultimi, che sono l'aspetto di gran lunga più importante della terapia antiepilettica dal punto di vista pratico.
Sebbene l'impiego del bromo e dei bromuri risalga al secolo scorso, si può affermare che l'era della terapia anticonvulsiva rientra nel nostro secolo. Essa è iniziata con l'impiego del fenobarbital (v. Hauptmann, 1912) ed è in pieno sviluppo.
I farmaci di cui oggi si dispone sono numerosi e numerosi anche i dati di farmacologia clinica che ne valutano efficacia, effetti secondari e tossicità (v. De Jong e Magee, 1965). Purtroppo nessuno di tali farmaci è frutto di una ricerca inizialmente diretta contro l'epilessia; al contrario il loro impiego è stato ed è del tutto empirico, anche se il loro meccanismo di azione a livello del tessuto nervoso è ipotizzato su basi scientifiche accettabili (v. cap. 2, È b, 4).
Al momento della scelta del farmaco o dei farmaci (giacchè vengono usati anche in associazione) occorre avere in mente tre principî fondamentali: a) vi è una selettività di efficacia di un farmaco verso forme particolari di epilessia; b) vi possono essere effetti secondari sfavorevoli tali da imporre la sospensione di farmaci utili nel loro effetto primario; c) in alcuni soggetti si possono verificare effetti tossici, anche gravi; tali pericoli sono maggiori per alcuni tipi di farmaci.
La concentrazione ematica di un farmaco dipende dalla quantità somministrata, dalla quota assorbita dall'intestino, dalla quantità della sua distribuzione nel sangue e nei tessuti e infine dalla quota di eliminazione di esso per mezzo del metabolismo e per mezzo dell'escrezione con gli emuntori. L'assorbimento intestinale rappresenta una barriera lenta, l'equilibrio sangue-tessuti si realizza più rapidamente.
La principale via di escrezione dei farmaci anticonvulsivi è rappresentata dal rene; il fegato è invece l'organo prevalentemente interessato alla loro metabolizzazione. Con la sola eccezione della fenitoina, il grado di eliminazione dei farmaci antiepilettici è proporzionale alla loro concentrazione nel siero, cioè l'eliminazione segue una cinetica di primo grado.
Quando vengono somministrate dosi ripetute, a intervalli più lunghi del tempo richiesto per la totale eliminazione dal sangue di un farmaco, le concentrazioni ematiche subiscono oscillazioni che vanno da zero a valori di poco inferiori al rapporto tra dose (mg/kg) e ‛volume di distribuzione'. Se invece il farmaco non è stato completamente eliminato al momento di ciascuna successiva dose, si verifica un accumulo di esso nel sangue. La concentrazione ematica aumenta con ciascuna dose successiva sino a un equilibrio che viene detto ‛livello ematico' e che è caratterizzato da un'eliminazione giornaliera uguale alla dose ingerita. Più basso è il grado di eliminazione più elevato risulta il livello ematico.
Acido fenilbarbiturico e dintoinici sono le sostanze a più ampio spettro di azione (crisi di grande male, crisi parziali a semeiologia elementare elaborata o complessa). Risultano però del tutto inefficaci contro le assenze del piccolo male. Il primidone (Mysoline) è efficace sia nelle crisi di grande male sia in quelle del lobo temporale. Il 5-carbamil-5H-dibenz-(bf)azepina (Tegretol) e l'acido N-(4′-sulfamoilfenil)-1,4-butansultame (Ospolot) sono efficaci soprattutto in queste ultime. Succinimmidici e dionici hanno efficacia contro le assenze del piccolo male ma nessun effetto sulle crisi generalizzate e parziali convulsive. Il Diazepam è efficace nelle assenze atipiche del bisincronismo secondario (per es., sindrome di Lennox). Le mioclonie massive della sindrome di West sono dominate dall'ACTH, che è efficace nel combattere anche gli altri sintomi di questa encefalopatia: il farmaco ha pertanto un meccanismo di azione diverso dagli anticonvulsivi, così come la piridossina in certi casi di epilessia ad essa sensibili.
L'elenco potrebbe continuare, ma non rientra tra gli scopi del presente scritto dare indicazioni dettagliate su questa materia, indicazioni che tra l'altro dovrebbero essere integrate dai dosaggi. L'uso corretto dei farmaci anticonvulsivi si basa su una esperienza, talora anche individuale, che non può essere riferita in poche righe.
Tra gli effetti collaterali indesiderabili devono menzionarsi in particolare: astenia psicofisica e sonnolenza (specie con Luminal e dintoinici); vertigini, vomito, cefalea (Mysoline, Tegretol, ecc.); eruzioni cutanee di tipo allergico (Luminai, dintoinici, ecc.); fotofobia (dionici). Talora questi effetti possono scomparire con la sola riduzione dei dosaggi, tuttavia in tal caso occorre valutare se per posologie ridotte viene conservato un efficace effetto anticonvulsivo.
Ben più importanti gli effetti tossici che sono abbastanza frequenti per alcuni farmaci. In particolare ricordiamo quelli causati da Fenurone, dionici e Tegretol (agranulocitosi, anemia, degenerazione epatica), senza tacere che anche per gli altri esistono tali possibili effetti negativi, donde la necessità di controlli seriati clinici e di laboratorio (per es., emocromo, prove di funzionalità epatica, ecc.).
Altri problemi terapeutici sono quello del trattamento dello stato di male epilettico, quello del significato degli esami elettroencefalografici seriati nella valutazione a distanza dell'efficacia delle cure e, per contro, il problema degli effetti tossici a distanza e dell'assuefazione.
Bibliografia
Abadie, J., Conceptions, étiologiques modernes sur les épilepsies, in ‟Revue neurologique", 1932, LXIV, pp. 1049-1138.
Abraham, K., Ajmone-Marsan, C., Patterns of cortical discharges and their relation to routine scalp electroencephalography, in ‟Electroencephalography and clinical neurophysiology", 1958, X, pp. 447-461.
Adrian, E. D., Electrical activity of the nervous system, in ‟Archives of neurology and psychiatry", 1934, XXXII, pp. 1125-1134.
Ajmone-Marsan, C., Electrographic aspects of ‛epileptic' neuronal aggregates, in ‟Epilepsia", 1961, II, pp. 22-38.
Ajmone-Marsan, C., A newly proposed classification of epileptic seizures. Neurophysiological basis, in ‟Epilepsia", 1965, VI, pp. 275-296.
Ajmone-Marsan, C., Microstructural mechanisms of seizure susceptibility, in Symposium of comparative and cellular pathophysiology of epilepsy (a cura di Z. Servit), Praha-Amsterdam 1965, pp. 47-59.
Ajmone-Marsan, C., Lewis, W. R., Pathologic findings in patients with centrecephalic electroencephalographic patterns, in ‟Neurology", 1960, X, pp. 922-930.
Alström, C. H., A study of epilepsy in its clinical, social and genetic aspects, in ‟Acta psychiatrica", 1950, suppl. 63, pp. 5-284.
Amantea, G., Rapporto fra eccitamenti afferenti ed epilessia sperimentale, in ‟Bollettino dell'Accademia Medica", 1920, VIII, pp. 46-121.
Andrioli, G., Angeleri, F., Bergonzi, P., Cantore, P., Ferroni, A., Gentilomo, A., Mingrino, S., Ricci, G. B., Rosadini, G., Rossi, G. F., Inquadramento diagnostico dell'epilettico in neurochirurgia, in ‟Minerva neurochirurgica, 1966, X, pp. 49-144.
Angeleri, F., Il problema dei potenziali parossistici bilaterali e sincroni in elettroencefalografia. A proposito di osservazioni sperimentali, in ‟Rivista di neurobiologia", 1958, IV, pp. 217-240.
Angeleri, F., Il sonno notturno negli epilettici, in L'epilessia. Argomenti di fisiopatologia, clinica e terapia (a cura di E. Lugaresi e P. Pazzaglia), Bologna 1968, pp. 268-303.
Angeleri, F., Boncompagni, P., Sacchetti, E., Quadri elettroencefalografici di epilessia focale in bambini senza corrispondenti manifestazioni accessuali, in ‟Rivista di neurobiologia", 1958, IV, pp. 315-324.
Angeleri, F., Ferro-Milone, F., Parigi, S., Contributi delle registrazioni profonde simultanee dal cervello umano al problema della elettroencefalografia dell'epilessia con particolare riguardo per quella del lobo temporale, in ‟Rivista di neurobiologia", 1961, suppl. 4, pp. 901-1094.
Angeleri, F., Ferro-Milone, F., Parigi, S., Electrical activity and reactivity of the rhinencephalic pararhinencephalic and thalamic structures. Prolonged implantation of electrodes in man, in ‟Electroencephalography and clinical neurophysiology", 1964, XVI, pp. 100-129.
Baglioni, S., Amantea, G., Il metodo della locale stimolazione chimica nello studio dei centri corticali, in ‟Archivio di farmacologia sperimentale", 1914, XVII, pp. 302-335.
Baglioni, S., Magnini, M., Azione di alcune sostanze chimiche sulle zone eccitabili della corteccia cerebrale del cane, in ‟Archivio di fisiologia", 1909, VI, pp. 240-258.
Bancaud, J., Colomb, D., Dell, M. B., Les pointes rolandiques. Un symptome EEG propre à l'enfant, in ‟Revue neurologique", 1958, IC, pp. 206-209.
Bancaud, J., Talairach, J., L'électroencéphalographie de profondeur, in Epilepsy, modern problems of pharmacopsychiatry (a cura di E. Niedermeyer), Basel 1970, pp. 29-41.
Bancaud, J., Talairach, J., Bonis, A., Schaub, C., Szikla, G., Morel, P., Bordas-Ferer, M., La stéréo-électroencéphalographie dans l'épilepsie, Paris 1965.
Bates, J. A. V., Special investigation techniques. Indwelling electrodes and electrocoticography, in Electroencephalography (a cura di D. Hill e G. Parr), London 1963, pp. 429-479.
Berger, H., Über das Elektroenzephalogram des Menschen, in ‟Archiv für Psychiatrie", 1929, LXXXVII, pp. 527-570.
Bevan, W., Sound-precipitated convulsions, in ‟Psychological bulletin", 1955, LII, pp. 473-502.
Bickford, R. G., The application of depth electrography in some varieties of epilepsy, in ‟Electroencephalography and clinical neurophysiology", 1956, VIII, pp. 526-527.
Bray, P. F., Wiser, W. C., The relation of focal to diffuse epileptiform EEG discharges in genetic epilepsy, in ‟Archives of neurology", 1965, XIII, pp. 223-237.
Brazier, M. A. B., Depth recordings from the amygdaloid region in patients with temporal lobe epilepsy, in ‟Electroencephalography and clinical neurophysiology", 1956, VIII, pp. 532-533.
Brazier, M. A. B., A history of the electrical activity of the brain. The first half-century, London 1961.
Bremer, F., Considérations sur l'origine et la nature des ‛ondes' cérébrales, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 177-193.
Clementi, A., Stricnizzazione della sfera corticale visiva ed epilessia sperimentale da stimoli luminosi e acustici, in ‟Archivio di fisiologia", 1929, XXVII, pp. 356-414.
Conrad, K., Die Bedeutung der Erbanlage bei der Epilepsie. Untersuchung an 253 Zwillingspaaren, in ‟Zentralblatt für die gesamte Neurologie und Psychiatrie", 1936, LXXVIII, pp. 160-192.
De Jong, R. N., Magee, K. R., Trattamento medico dell'epilessia nell'adulto, in Terapia moderna, vol. I, Roma 1965, pp. 36-51.
De Robertis, E., Ultrastructure and cytochemistry of the synaptic region, in ‟Science", 1967, CLVI, pp. 907-914.
Dichter, M., Spencer, A. W., Penicillin induced interictal discharge from the cat hippocampus, in ‟Journal of neurophysiology", 1969, XXXII, pp. 649-687.
Eccles, J. C., The physiology of synapses, Berlin 1964.
Eccles, J. C., The inhibitory path-ways of the central nervous system, Springfield, Ill., 1969.
Echlin, F. A., The supersensitivity of chronically ‛isolated' cerebral cortex as a mechanism in focal epilepsy, in ‟Electroencephalography and clinical neurophysiology", 1959, XI, pp. 697-722.
Ferrier, D., Experimental researches in cerebral physiology and pathology, in ‟West Riding lunatic asylum medical reports", 1873, III, pp. 30-96.
Fischer, J., Electron microscopic changes in the perikarya and in the processes of ganglion cells in the cobalt gelatine epileptogenic focus, in ‟Physiologia bohemoslovenica", 1969, XVIII, pp. 387-394.
Fritsch, G., Hitzig, H., Über die elektrische Erregbarkeit des Grossgehirns, in ‟Archiv für Anatomie, Physiologie und wissenschaftliche Medicin", 1870, pp. 300-332.
Gastaut, H., The epilepsies. Electro-clinical correlations, Springfield, Ill., 1954.
Gastaut, H., Conclusions, in Reflex mechanism in the genesis of epilepsy (a cura di Z. Servit), Amsterdam 1963, pp. 249-250.
Gastaut, H., On genetic transmission of epilepsies, in ‟Epilepsia", 1969, X, pp. 3-6.
Gastaut, H., Fischer-Williams, M., The physiopathology of epileptic seizures, in Handbook of physiology (a cura di J. Filed, H. W. Magoun e V. F. Hall), vol. I, Neurophysiology, Washington 1959, pp. 329-363.
Gastaut, H., Glaser, G. M., Magnus, O., Symposium on epilepsy and heredity, in ‟Epilepsia", 1969, numero speciale 10, pp. 1-100.
Gastaut, H., Poirier, F., Payan, H., Salamon, G., Toga, M., Vigoroux, M. H., H. H. E. syndrome. Hemiconvulsions, hemiplegia, epilepsy, in ‟Epilepsia", 1959-1960, I, pp. 418-447.
Gastaut, H., Soularyol, R., Roger, J., Pinsard, N., L'encephalopathie myoclonique infantile avec hypsaritmie (Syndrome de West), Paris 1964.
Gastaut, Y., Un élément déroutant de la séméiologie électroéncephalographique. Les pointes pre-rolandiques sans signification focale, in ‟Revue neyrologique", 1952, LXXXVII, pp. 488-490.
Gibbs, E. L., Gillen, H. W., Gibbs, F. A., Disappearance and migration of epileptic foci in childhood, in ‟American journal of diseases of children", 1954, LXXXVIII, pp. 596-603.
Gibbs, F. A., Davis, H., Lennox, W. G., The electroencephalogram in epilepsy and in conditions of impaired consciousness, in ‟Archives of neurology and psychiatry", 1935, XXXIV, pp. 1133-1148.
Gloor, P., Contribution of the intracarotid sodium amytal and metrazol tests to the understanding of the mechanisms of ‛centrencephalic' epilepsy, in ‟Electroencephalography and clinical neurophysiology", 1968, XXIV, pp. 289-290.
Gloor, P., Generalized cortico-reticular epilepsies. Some considerations on the pathophysiology of generalized bilaterally synchronous spike and wave discharge, in ‟Epilepsia", 1968, IX, pp. 249-263.
Gowers, W. R., Epilepsy and other chronic convulsive diseases, London 1881.
Hall, C. S., Genetic differences in fatal audiogenic seizures between two inbred strains of house mice, in ‟Journal of heredity", 1947, XXXVIII, pp. 3-6.
Hauptmann, A., Luminal bei Epilepsie, in ‟Münchner medizinische Wochenschrift", 1912, LIX, pp. 1907-1909.
Hill, D., Discussion on the surgery of temporal lobe epilepsy. The clinical study and selection of patients, in ‟Proceedings of the Royal Society of Medicine", 1953, XLVI, pp. 965-976.
Hill, D., Epilepsy. Clinical aspects, in Electroencephalography (a cura di D. Hill e G. Parr), London 1963, pp. 250-294.
Horsley, V., Trephining in the neolitic period, in ‟Journal of the Anthropology Institute", 1887-1888, XVII, pp. 100-106.
Hughes, J. R., Bilateral EEG abnormalities on corresponding areas, in ‟Epilepsia", 1966, VII, pp. 44-52.
Jackson, J. H., A study of convulsion, in ‟Transaction of the St. Andrews Medical Graduates Association", 1870, III, pp. 1-45 (reprint: Selected writings of J. H. Jackson, a cura di J. Taylor, London 1931, pp. 8-36).
Jackson, J. H., On the anatomical, physiological and pathological investigations of the epilepsies, in ‟West Riding lunatic asylum medical reports", 1873, III, pp. 315-339.
Janz, D., Die Epilepsien. Spezielle Pathologie und Therapie, Stuttgart 1969.
Jasper, H. H., Diffuse projections systems. The integration action of the thalamic reticular system, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 405-420.
Jasper, H. H., Review of the concept of centrencephalic epilepsy (Kershman memorial lecture), in ‟Electroencephalography and clinical neurophysiology", 1965, XIX, pp. 320-327.
Jasper, H. H., Drogleever Fortuyn, J., Experimental studies on the functional anatomy of petit mal epilepsy, in ‟Proceedings of the Association for Nervous and Mental Diseases", 1946, XXVI, pp. 272-298.
Jasper, H. H., Kershman, J., Classification of the EEG in epilepsy, in ‟Electroencephalography and clinical neurophysiology", 1949, suppl. 2, pp. 123-131.
Kellaway, P., The characteristics of epileptogenic foci in children. Projecte dand other secondary effects produced by focal discharge, in ‟Electroencephalography and clinical neurophysiology", 1954, VI, pp. 698-699.
Killam, K. F. Jr., Genetic model of epilepsy with special reference to the syndrome of the Papio papio, in ‟Epilepsia", 1969, X, pp. 229-238.
Killam, K. F. Jr., Naquet, R., Bert, J., Paroxysmal responses to intermittent light stimulation in a population of baboons (Papio papio), in ‟Epilepsia", 1966, VII, pp. 215-219.
Lennox, W. G., Davis, J. P., Clinical correlates of the fast and the slow spike-wave electroencephalogram, in ‟Pediatrics", 1950, V, pp. 626-644.
Lennox, W. G., Lennox, M. A., Epilepsy and related disorders, Boston 1960.
Longo, V., Il contributo dell'epilessia sperimentale allo studio della patogenesi dell'epilessia umana, in Enciclopedia medica italiana, vol. I, Firenze 1952, pp. 142-144.
McKhann, G. M., Shooter, E. M., Genetics of seizure susceptibility, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper e altri), Boston 1969, pp. 347-373.
Madsen, J. A., Bray, P. F., The coincidence of diffuse electroencephalographic spike-wave paroxysms and brain tumors, in ‟Neurology", 1966, XVI, pp. 546-555.
Marchand, L., Comment apprécier le rôle de l'hérédité dans les épilepsies, in ‟Presse médicale", 1952, LX, pp. 1472-1473.
Marcus, E. M., Watson, C. W., Reproduction in isolated cortex of cat brain of bilateral synchronous and symmetrical spike and slow wave pattern resembling petit mal, in ‟Transactions of the American Neurological Association", 1964, LXXXIX, pp. 221-222.
Marcus, E. M., Watson, C. W., Bilateral synchronous spike wave electrographic patterns in the cat. Interaction of bilateral cortical-calcolosal and diencephalic preparation, in ‟Archives of neurology", 1966, XIV, pp. 601-610.
Marcus, E. M., Watson, C. W., Studies of the bilateral corticocallosal preparation, in ‟Transactions of the American Neurological Association", 1966, XCI, pp. 291-293.
Marcus, E. M., Watson, C. W., Simon, S. A., An experimental model of some varieties of petit mal epilepsy. Electrical-behavioral correlations of acute bilateral epileptogenic foci in cerebral cortex, in ‟Epilepsia", 1968, IX, pp. 233-248.
Massa, T., Niedermeyer, E., Convulsive disorders during the first three months of life, in ‟Epilepsia", 1968, IX, pp. 1-9.
Metrakos, J. D., Metrakos, K., Genetics of convulsive disorders. I. Introduction, problems, methods and baselines, in ‟Neurology", 1960, X, pp. 228-240.
Metrakos, J. D., Metrakos, K., Genetics of convulsive disorders. II. Genetic and electroencephalographic studies in centrencephalic epilepsy, in ‟Neurology", 1961, XI, pp. 474-483.
Metrakos, J. D., Metrakos, K., Genetic factors in epilepsy, in Epilepsy. Modern problems of pharmacopsychiatry (a cura di E. Niedermeyer), Basel 1970, pp. 71-86.
Morrell, F., Secondary epileptogenic lesions, in ‟Epilepsia", 1960, II, pp. 538-560.
Morrell, F., Cellular pathophysiology of focal epilepsy, in ‟Epilepsia", 1969, X, pp. 495-505.
Moruzzi, G., Magoun, H. W., Brain stem reticular formationand activation of the EEG, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 455-473.
Nachtsheim, H., Krampfberitschaft und Erbbild des Epileptikers, in ‟Erbarzt", 1940, VIII, pp. 12-25.
Nayrac, P., Beaussart, M., Les pointes-ondes prérolandiques, expression électroencéphalographique trés particulière, in ‟Revue neurologique", 1958, IC, pp. 203-205.
Niedermeyer, E., Common generalized epilepsy and generalized synchronous seizure discharges, in Epilepsy. Modern problems of pharmacopsychiatry (a cura di E. Niedermeyer), Basel 1970, pp. 139-154.
Ounsted, C., Lindsay, J., Norman, R., Biological factors in temporal lobe epilepsy, London 1966.
Penfield, W., Jasper, H., Epilepsy and the functional anatomy of the human brain, London 1954.
Petsche, H., Stumpf, Ch., Gogolak, G., The significance of the rabbit's septum as a relay station between the midbrain and the hippocampus. I. Control of hippocampus arousal activity by septum cells, in ‟Electroencephalography and clinical neurophysiology", 1962, XIV, pp. 202-211.
Pinelli, P., L'elettroencefalografia nelle convulsioni dell'infanzia, in Atti del XXIII congresso della Società Italiana di Pediatria, Bologna 1954, pp. 63-96.
Pollen, D. A., Experimental spike and wave responses and petit mal epilepsy, in ‟Epilepsia", 1968, IX, pp. 233-248.
Pollen, D. A., Trachtenberg, M. C., Neuroglia. Gliosis and focal epilepsy, in ‟Science", 1970, CLXVII, pp. 1252-1254.
Pond, D. A., The EEG in pediatrics, in Electroencephalography. A symposium on its various aspects (a cura di D. Hill e G. Parr), London 1963, pp. 207-231.
Ralston, B. L., Cingulate epilepsy and secondary bilateral synchrony, in ‟Electroencephalography and clinical neurophysiology", 1961, XIII, pp. 591-598.
Ralston, B. L., Ajmone-Marsan, C., Thalamic control of certain normal and abnormal cortical rhythms, in ‟Electroencephalography and clinical neurophysiology", 1956, VIII, pp. 559-582.
Rasmussen, T., The role of surgery in the treatment of focal epilepsy, in ‟Clinical neurosurgery", 1969, XVI, pp. 288-314.
Rodin, E., Gonzales, S., Hereditary components in epileptic patients. Electroencephalographic family studies, in ‟Journal of the American Medical Association", 1966, CXCVIII, pp. 221-225.
Rossi, G. F., Walter, R. D., Crandall, P. H., Generalized spike and wave discharge and nonspecific thalamic nuclei. A stereotaxic analysis, in ‟Archives of neurology", 1968, XIX, pp. 174-183.
Sawa, M., Nakamura, K., Naito, H., Intracellular phenomena and spread of epileptic seizure discharges, in ‟Electroencephalography and clinical neurophysiology", 1968, XXIV, pp. 146-154.
Scherman, R. G., Abraham, K., Centrencephalic electroencephalographic patterns in precocious puberty, in ‟Electroencephaloraphy and clinical neurophysiology", 1963, XV, pp. 559-567.
Servít, Z., Comparative and cellular pathology as method of research in pathogenesis of epilepsy, in Symposium of comparative and cellular pathophysiology of epilepsy (a cura di Z. Servít), Praha-Amsterdam 1965, pp. 7-15.
Servít, Z., Mackek, J., Fischer, J., Electrical activity of the frog brain during electrically induced seizures. A comparative study of the spike and wave complex, in ‟Electroencephalography and clinical neurophysiology", 1965, XIX, pp. 162-171.
Sorel, L., The descendants of epileptic patients, in ‟Epilepsia", 1969, X, pp. 91-96.
Sperry, R. W., Brain bisection and consciousness, in Brain and conscious experience (a cura di J. C. Eccles), Berlin 1966, pp. 298-307.
Spiegel, E. A., Wycis, H. T., Thalamic recordings in man with special reference to seizure discharges, in ‟Electroencephalography and clinical neurophysiology", 1950, II, pp. 23-27.
Stavraky, C. W., Supersensitivity following lesions of the nervous system, Toronto 1961.
Storm van Leeuwen, W., Magnus, D., La signification de l'analyse automatique de l'électroencéphalogramme pour le diagnostic clinique, in ‟Revue neurologique", 1961, CIV, pp. 177-203.
Strobos, R. J., Kavallinis, G. P., Changes in repeated electroencephalograms in epileptics, in ‟Neurology", 1968, XVIII, pp. 622-633.
Sutherland, J. M., Tait, H., The epilepsies. Modern diagnosis and treatment, Edinburgh 1969.
Tassinari, C. A, Le crisi epilettiche riflesse, in L'epilessia. Argomenti di fisiopatologia, clinica e terapia (a cura di E. Lugaresi e P. Pazzaglia), Bologna 1968, pp. 87-112.
Temkin, O., The falling sickness. A history of epilepsy from the Greeks to the beginning of modern neurology, Baltimore 1945.
Tower, D. B., Neurochemistry of epilepsy. Seizure mechanisms and their management, Springfield, Ill., 1960.
Tower, D. B., Neurochemical mechanisms, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper e altri), Boston 1969, pp. 611-638.
Trachtenberg, M. C., Pollen, D. A., Neuroglia. Biophysical properties and physiologic function, in ‟Science", 1970, CLXVII, pp. 1248-1251.
Tückel, K., Jasper, H. H., The elctroencephalogram in parasagital lesions, in ‟Electroencephalography and clinical neurophysiology", 1952, IV, pp. 481-494.
Udvarhelyi, G. B., Walker, A. E., Dissemination of acute focal seizures in the monkey, in ‟Archives of neurology", 1965, XII, pp. 333-380.
Walker, A. E., Trattamento chirurgico dell'epilessia, in Terapia moderna, vol. I, Roma 1965, pp. 69-77.
Walker, A. E., Marshall, C., The contribution of depth recordings to clinical medicine, in ‟Electroencephalography and clinical neurophysiology", 1964, XVI, pp. 88-89.
Walker, A. E., Ribstein, M., Enregistrements et stimulations des formations rhinencéphaliques avec électrodes profondes à demeure chez l'homme, in ‟Revue neurologique", 1957, XCVI, pp. 453-459.
Williams, D., The thalamus and epilepsy, in ‟Brain", 1965, LXXXVIII, pp. 539-556.
Woodbury, D. M., Mechanisms of action of anticonvulsants, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper e altri), Boston 1969, pp. 647-681.
Considerando ora la coordinazione motoria, sembra che, se si eccettuano quelle risposte motorie complesse che vengono apprese sulla base d'informazioni tattili e cinestesiche con un arto e non sono trasferite all'altro arto, sia nel gatto con cervello bisecato (premere un pedale; v. i contributi di Sperry, 1961), che nei Primati con cervello bisecato (aprire un chiavistello alla cieca; v. Myers e Henson, 1960; v. Kohn e Myers, 1969), afferenze sensitive limitate a un emisfero possono evocare attività motorie egualmente efficienti nei due lati del corpo (v. Gazzaniga, 1970). In generale, una scimmia con cervello bisecato e un occhio coperto non ha alcuna difficoltà a usare entrambe le mani per prendere del cibo. Eppure, la combinazione occhio-mano dello stesso lato dovrebbe almeno in principio chiamare in causa la corteccia visiva di un emisfero e la corteccia motoria dell'altro (v. Downer, 1959; v. Lehman, 1968). Gazzaniga (v., 1970) ha escluso la possibilità che interazioni a livello del tronco dell'encefalo possano mediare la prensione controllata dalla visione, dimostrando che bisecando il cervello non si altera questo aspetto del comportamento. È chiaro che possono essere in gioco vie motorie ipsilaterali, ma secondo Gazzaniga queste sono solo una parte del meccanismo. Nelle prove di prensione guidate dall'occhio ipsilaterale alla mano impiegata, l'emisfero veggente della scimmia con cervello bisecato potrebbe semplicemente dare un suggerimento all'altro emisfero, che controlla la mano impegnata nella risposta, dirigendo la testa e gli occhi verso il bersaglio. Le afferenze propriocettive della testa e degli occhi, in larga misura bilaterali, possono poi guidare l'emisfero cieco nel dirigere esattamente la mano verso la mira. Gazzaniga ha portato prove sperimentali in favore di questa teoria, dimostrando che l'immobilizzazione della testa ostacola notevolmente la capacità della scimmia con cervello bisecato di usare la mano ipsilaterale all'occhio che vede per compiere un movimento piuttosto complesso indirizzato dalla visione. Altri esempi di questi suggerimenti periferici verranno discussi nella descrizione del comportamento di soggetti umani con cervello bisecato (v. sotto, § b). Un uso corretto delle due mani sotto controllo visivo è stato osservato anche in scimmie con cervello bisecato che hanno un prisma che deflette la luce davanti a un occhio e l'altro occhio coperto, a patto che all'animale venga permesso di usare entrambe le mani nel periodo iniziale di adattamento motorio alla distorsione delle afferenze visive. In questi animali, il trasferimento intermanuale dell'adattamento motorio a una mano senza esperienza non avviene mai, mentre il trasferimento interoculare è sempre positivo. Ciò starebbe a indicare che l'adattamento motorio alla distorsione delle afferenze visive implica aggiustamenti sul versante motorio-propriocettivo anziché sul versante visivo (v. Hamilton, 1967). Un altro importante problema nella integrazione motoria bilaterale nei Primati con cervello bisecato è la coordinazione dell'attività bimanuale nella manipolazione di oggetti e in altri atti motori fini. Sebbene si possa osservare qualche deficit subito dopo l'operazione di bisezione cerebrale (v. Trevarthen, 1965), la regola è che scimmie con cervello bisecato non differiscono apprezzabilmente da questo punto di vista da controlli normali (v. Mark e Sperry, 1968). La rappresentazione somestesica e il controllo motorio ipsilaterale, e la strategia dei suggerimenti crociati di Gazzaniga sono tutti fattori che possono concorrere, in diverse misure, a rendere possibile la persistenza di una buona coordinazione bimanuale dopo la bisezione cerebrale.
Le informazioni sul ruolo delle commissure telencefaliche nell'olfatto e nell'udito sono piuttosto scarse. H. Teitelbaum (v., 1971) ha dimostrato nel ratto che discriminazioni olfattive apprese con una narice non vengono trasferite all'altra narice se la commissura anteriore viene tagliata prima dell'apprendimento. Riguardo all'udito, l'affermazione di Pavlov (v., 1927) che cani condizionati a rispondere differenzialmente a suoni provenienti dalla destra o dalla sinistra perdono questa capacità dopo callosotomia non è stata confermata (v. Neff, 1961). Un ostacolo anatomico all'applicazione della tecnica del cervello bisecato al sistema uditivo è costituito dall'ampia rappresentazione di ciascuna coclea in entrambi gli emisferi: questa rappresentazione bilaterale, con una certa predominanza delle connessioni crociate, è dovuta ai molti parziali incrociamenti delle vie uditive ai livelli sottocorticali. È praticamente impossibile eliminare chirurgicamente la rappresentazione bilaterale delle coclee negli emisferi. Le ipotesi sulle attività commissurali nell'udito sono state descritte in una rassegna di J. O. Wegener (v., 1965). Più recentemente, J. Kaas, S. Axelrod e I. T. Diamond (v., 1967) hanno descritto risultati che indicano che il corpo calloso può giocare un ruolo insospettato nella percezione uditiva. Alcuni gatti venivano allenati a rispondere usando un orecchio a un cambio di frequenza tonale, non prestando attenzione a simili cambi di frequenza presentati simultaneamente all'altro orecchio. L'ablazione della corteccia uditiva controlaterale all'orecchio ‛attento' portava alla perdita della capacità di eseguire il compito, mentre non v'erano deficit dopo ablazione della corteccia uditiva ipsilaterale allo stesso orecchio. Questo risultato suggerisce che l'engramma era lateralizzato in un emisfero. Tuttavia, se il corpo calloso veniva sezionato prima dell'apprendimento, la capacità di risolvere il problema persisteva dopo ablazione della corteccia uditiva controlaterale all'orecchio ‛attento'. In questi esperimenti, il corpo calloso era perciò necessario per mantenere l'engramma localizzato in un emisfero, del tutto in contrasto con gli esperimenti sulla visione, sulla somatoestesia e sull'olfatto di cui abbiamo già detto e con i quali si è dimostrato che il corpo calloso è necessario per permettere l'uso dell'engramma da parte d'entrambi gli emisferi. Resta da vedere se questa funzione restrittiva del corpo calloso possa valere anche per altre modalità quando le afferenze sensoriali vengono dirette a entrambi gli emisferi.
b) Esperimenti su soggetti umani con cervello bisecato
In alcuni casi della patologia umana il corpo calloso non si sviluppa nel corso dell'ontogenesi del sistema nervoso (agenesia callosale), mentre in altri casi un corpo calloso che si è sviluppato normalmente viene distrutto nel corso della vita da processi degenerativi o tumorali. Inoltre, il neurochirurgo deve talvolta tagliare un corpo calloso normale o per poter accedere a formazioni patologiche sottostanti (cisti, tumori e così via) che devono essere rimosse, o per alleviare il decorso di epilessie altrimenti incurabili, interrompendo una delle più importanti vie per la propagazione intracerebrale di attività nervose abnormi (v. sopra, cap. 3, § a). È perciò possibile studiare le funzioni callosali nell'uomo esaminando questi vari tipi di soggetti acallosali. Fino a pochi anni fa, il quadro generale era del tutto oscuro, ma recentemente, grazie ai suggerimenti forniti dalla sperimentazione sugli animali, molte lacune nella nostra conoscenza delle attività interemisferiche nell'uomo sono state colmate e molte delle precedenti contraddizioni in quest'area sono state risolte. Un breve resoconto storico faciliterà forse la descrizione dello stato attuale delle conoscenze in questo campo di ricerca.
Nel 1922 G. Mingazzini riassunse in un'ampia monografia una varietà di studi anatomici, fisiologici e clinici sulle funzioni del corpo calloso. Sulla base dei dati in suo possesso, Mingazzini ritenne di poter concludere che il corpo calloso ha un ruolo essenziale nelle attività della mente e che, nel cervello umano, questa struttura è indispensabile per la coordinazione dell'emisfero sinistro, dominante, e dell'emisfero destro, subordinato, nel linguaggio e nella prassi. Aderendo strettamente alla concezione dei Leitungsstörungen di Liepmann e Déjerine, egli tentò perfino di suddividere, in termini anatomofunzionali, il corpo calloso in una parte anteriore, o portio verbalis, una parte intermedia, portio praxica, e una parte posteriore, portio gnostica. Circa venti anni dopo, una lunga serie di ricerche di A. J. Akelaitis e collaboratori (v. Akelaitis, 1944; v. Smith, 1952) su pazienti sottoposti a sezione del corpo calloso e della commissura anteriore per ragioni terapeutiche diede risultati in netto contrasto con la conclusione di Mingazzini sull'importanza funzionale del corpo calloso. In breve, i soggetti con cervello bisecato di Akelaitis non presentavano in generale sintomi afasici, alessici e aprassici, e mostravano un'integrazione percettiva normale di informazioni sensitivo-sensoriali che originavano dai due lati del corpo o dalle due metà del campo visivo. I sintomi patologici eventualmente presenti potevano essere attribuiti a danni extracommissurali preesistenti o intraoperatori, anziché alla sezione delle commissure di per sé. Il fatto che la commissurotomia non producesse i disturbi da attendersi sulla base delle classiche teorie connessionistiche era in buon accordo con le concezioni olistiche e ‛gestaltiche' del cervello, allora prevalenti. Di conseguenza, il problema delle comunicazioni interemisferiche nell'uomo fu trascurato per parecchi anni, a dispetto di occasionali reperti ‛positivi', come quello riportato da P. E. Maspes (v., 1948) relativo a un caso di alessia nel campo visivo di sinistra a seguito della sezione dello splenio del corpo calloso. Furono i successi della sperimentazione sugli animali con cervello bisecato che spinsero Sperry e collaboratori a riconsiderare il problema delle funzioni commissurali nell'uomo. Gli esperimenti furono nuovamente eseguiti in un numero di pazienti epilettici che erano stati sottoposti alla sezione completa del corpo calloso e della commissura anteriore con lo scopo di controllare accessi non trattabili con altri mezzi. L'operazione ha avuto successo dal punto di vista terapeutico (v. Bogen e altri, 1969) e al tempo stesso ha offerto un'occasione unica per confermare e ampliare nell'uomo i risultati della sperimentazione sugli animali. Come gli animali con cervello bisecato, i soggetti umani con cervello bisecato non sono disturbati nelle loro attività ordinarie, e presentano sintomi di disconnessione emisferica solo quando sono esaminati con prove sperimentali speciali. Ampie rassegue sono state pubblicate da Sperry (v., 1966 e i contributi del 1968 e del 1970), Hécaen e Assal (v., 1968), J. E. Bogen (v., The other side of the brain. I, II, 1969), J. E. Bogen e G. M. Bogen (v., 1969) e Gazzaniga (v., 1970); si rimanda ad esse il lettore per informazioni dettagliate. In breve, la sindrome di disconnessione commissurale e le prove per metterla in evidenza sono le seguenti. Per limitare le afferenze ottiche a un singolo emisfero, gli stimoli visivi sono presentati nella metà controlaterale del campo visivo, a pochi gradi dal punto di fissazione e al suo stesso livello. Gli stimoli (lettere, parole, immagini di oggetti e così via) vengono proiettati su uno schermo di fronte al soggetto. Il tempo di esposizione è molto breve (circa 100 ms), cosicché il soggetto non può usare i movimenti oculari per portare lo stimolo nell'area della visione centrale e ottenere così una stimolazione biemisferica. Quando si chiede ai pazienti con cervello bisecato di riconoscere lo stimolo proiettato sullo schermo dicendone il nome, si vede che essi lo possono fare solo quando lo stimolo viene presentato nel campo visivo di destra (e quindi all'emisfero sinistro). Cioè, parole, numeri, lettere, colori, immagini di oggetti e figure geometriche sono sempre prontamente identificati con il loro nome da questi pazienti quando le informazioni visive sono dirette all'emisfero che è dominante per il linguaggio (v. emisferi cerebrali: Dominanza cerebrale). Risultati del tutto differenti si ottengono quando gli stessi stimoli vengono proiettati nell'emicampo visivo di sinistra, incanalando così l'informazione visiva nell'emisfero destro. In questo caso, i pazienti con cervello bisecato non solo sono incapaci di identificare lo stimolo con il suo nome, ma sostengono anche che non hanno ricevuto alcuno stimolo, eccetto forse un lampo di luce. Tuttavia, se in base a ciò si concludesse che questi pazienti hanno un'agnosia visiva nel campo sinistro si commetterebbe un grave errore, perché il deficit si manifesta solo quando si richiede che il riconoscimento degli stimoli sia di natura verbale. Infatti, se si trascura quello che i soggetti dicono di ciò che vedono, e si chiede loro di riconoscere lo stimolo proiettato in modo non verbale, è possibile dimostrare che la percezione nel campo visivo di sinistra è buona come quella nel campo destro. Per esempio, i pazienti con cervello bisecato non hanno difficoltà a scegliere da una collezione di oggetti diversi l'oggetto che corrisponde allo stimolo proiettato nel campo visivo di sinistra; o possono identificare al tatto questo oggetto e prelevarlo da dietro una tenda che impedisce loro di vedere i diversi oggetti della collezione. Tuttavia questa prova di associazione visivo-tattile riesce solo quando il soggetto usa la mano sinistra, cioé la mano rappresentata nell'emisfero destro. Analogamente, il riconoscimento tattile di oggetti che corrispondono a stimoli proiettati nel campo visivo di destra è possibile solo con la mano destra. Come nel caso del riconoscimento di stimoli visivi lateralizzati, l'identificazione verbale (orale o scritta) di oggetti posti fuori del campo visivo ed esaminati al tatto può avvenire solo quando l'informazione sensitiva è diretta all'emisfero sinistro, cioè quando viene usata la mano destra. Il riconoscimento con la mano sinistra (e l'emisfero destro) non può essere che averbale (v. i contributi di Sperry, 1968; v. Gazzaniga, 1970). Risultati del tutto simili si ottengono nella modalità olfattiva. Poiché la sezione della commissura anteriore interrompe gran parte delle connessioni crociate, ciascun bulbo olfattivo resta connesso quasi esclusivamente con i centri olfattivi nell'emisfero ipsilaterale. In accordo a ciò, i pazienti con cervello bisecato possono dire il nome di sostanze odorose introdotte nella narice sinistra, ma non di sostanze introdotte nella narice destra. Tuttavia, l'identificazione corretta di stimoli presentati attraverso la narice destra si dimostra ancora solo con prove non verbali (v. Gordon e Sperry, 1969). Per quanto riguarda l'udito, è già stato sottolineato il fatto che la lateralizzazione emisferica delle afferenze acustiche è quasi impossibile da ottenersi (v. sopra, § a). Tuttavia, il metodo della stimolazione simultanea dei due orecchi con materiale verbale diverso è stato usato con successo per esaminare le interazioni emisferiche nella sfera uditiva. I pazienti con cervello bisecato possono ripetere correttamente cifre presentate separatamente all'orecchio destro e a quello sinistro, ma se cifre differenti sono presentate simultaneamente ai due orecchi, essi possono ripetere solo le cifre dell'orecchio destro e poche o nessuna dell'orecchio sinistro. Una lieve predominanza dell'orecchio destro in questa prova si osserva anche in soggetti normali, ma questa differenza fra gli orecchi è molto piccola quando la si paragona alla quasi totale soppressione dell'orecchio sinistro nei pazienti con cervello bisecato. Se si davano ordini diversi di prendere oggetti nascosti alla vista simultaneamente ai due orecchi, e si chiedeva ai pazienti di usare la mano sinistra, gli oggetti nominati attraverso l'orecchio destro erano ora quasi totalmente ignorati, e solo le istruzioni date all'orecchio sinistro venivano eseguite correttamente (v. Milner e altri, 1968). Secondo questi risultati, è come se le commissure telencefaliche permettessero una registrazione di segnali diversi provenienti simultaneamente dai due orecchi; in loro assenza, solo le afferenze controlaterali sono registrate a spese di quelle ipsilaterali.
In conclusione, è chiaro che in tutte le modalità le informazioni di senso inviate esclusivamente a un emisfero del paziente commissurotomizzato sono analizzate solo entro quell'emisfero e non vengono comunicate all'altro emisfero. Dato che i centri del linguaggio sono lateralizzati nell'emisfero sinistro, l'emisfero destro può esternare ciò che sa solo con mezzi non verbali. Per citare Sperry (v., Mental unity..., 1968), i pazienti con cervello bisecato hanno ‟[...] un emisfero cosiddetto dominante o maggiore che può parlarci e scrivere, e un emisfero subordinato o minore che non può esprimersi a parole". Tuttavia, questo non significa che l'emisfero destro sia inferiore a quello sinistro da tutti i punti di vista, o che non abbia capacità di comprendere il linguaggio. In prove di capacità motoria e prassica non vi sono differenze fra gli emisferi nel controllo della mano controlaterale. In prove di disegno che implichino capacità visuo spaziali specifiche la mano sinistra spesso supera la destra (v. Bogen, The other side of the brain. I, 1969) indicando così, in accordo con i dati neurologici (v. emisferi cerebrali: Dominanza cerebrale), una superiorità dell'emisfero destro in questo tipo di prove. Riguardo al linguaggio, il solo fatto che i pazienti con cervello bisecato possono scegliere una carta con il nome di un oggetto dopo che un'immagine di questo oggetto è stata proiettata all'emisfero destro indica l'esistenza di qualche comprensione linguistica in questo emisfero. Dati recenti indicano tuttavia che i limiti della capacità dell'emisfero destro a comprendere e a ‛pensare' in termini verbali sono molto ridotti (v. Gazzaniga e Hillyard, 1971).
Molte altre interessanti caratteristiche percettive dei pazienti con cervello bisecato vengono continuamente portate alla luce da procedimenti d'esame appositamente escogitati. Per esempio è stato dimostrato che questi pazienti non hanno stereopsia binoculare nella parte centrale del campo visivo (v. Mitchell e Blakemore, 1970) né mostrano risposte convergenti o divergenti degli occhi a stimoli presentati al davanti o al di dietro del punto di fissazione (v. Westheimer e Mitchell, 1969). Poiché sia la stereopsia binoculare che i movimenti convergenti e divergenti sono del tutto normali quando gli stimoli sono presentati eccentricamente, e quindi sono proiettati a un solo emisfero, sembra logico attribuire i deficit nelle risposte a stimoli lungo la linea mediana, alla mancanza dei meccanismi callosali che estendono i campi recettivi della corteccia visiva attraverso il meridiano verticale centrale. Tuttavia, dato che le prove della stereopsia nei pazienti con cervello bisecato sono sempre state eseguite con stimoli con grandi disparità, è stato suggerito che, al contrario della stereopsia grossolana, la stereopsia fine lungo il piano mediosagittale sia dovuta alla imprecisione della decussazione chiasmatica e non alle connessioni callosalì (v. Bishop, 1973; v. Berlucchi, 1972).
Questo breve riassunto dei risultati delle ricerche sul cervello bisecato nell'uomo indica chiaramente che la sezione delle commissure cerebrali produce nel cervello umano una sindrome da disconnessione fondamentalmente simile a quella osservata negli animali commissurotomizzati: se le afferenze di senso sono lateralizzate a un emisfero, le informazioni portate da queste afferenze in genere non possono essere comunicate all'altro emisfero. In queste condizioni, ogni emisfero ha la sua sfera privata di percezione e di conoscenza. Ciò che differenzia la sindrome da disconnessione interemisferica nell'uomo da quella negli animali è la rimarchevole differenziazione funzionale fra gli emisferi del cervello umano, con la lateralizzazione dei processi verbali e analitici nell'emisfero sinistro e delle capacità visuospaziali e gestaltiche nell'emisfero destro (v. emisferi cerebrali: Dominanza cerebrale; v. i contributi di Bogen, 1969; v. Bogen e Bogen, 1969).
Questi risultati pongono domande molto importanti per il problema generale delle relazioni fra la mente e il cervello. Vi sono letteralmente due menti nel paziente con cervello bisecato, una in ciascun emisfero? Questo è ovviamente un problema semantico, che Sperry (v., 1966), MacKay (v., 1966), Eccles (v., 1965), Bogen (v. i contributi del 1969), Bogen e Bogen (v., 1969) e Gazzaniga (v., 1970 e 1972) hanno discusso partendo da diverse posizioni filosofiche. Poiché molte delle questioni in gioco sono di natura epistemologica, esse non possono essere risolte dall'esperimento. Vi sono invece altri aspetti delle ricerche sul cervello bisecato nell'uomo che possono indubbiamente portare a progressi scientifici sperimentali. Una linea di sviluppo può forse partire dalla considerazione delle nette differenze frai pazienti con cervello bisecato di Sperry, Bogen e Gazzaniga da un lato, e i pazienti con cervello bisecato di Akelaitis e i casi di agenesia callosale (v. Unterharnscheidt e altri, 1968) dall'altro lato. Nei pazienti con cervello bisecato di Akelaitis, come nella maggior parte dei soggetti con agenesia callosale, non v'era sindrome da disconnessione interemisferica. Ciò può essere spiegato da vari fattori: a) vi potrebbe essere una completa equivalenza funzionale fra gli emisferi di questi soggetti, con una bilateralizzazione dei centri del linguaggio (v. Sperry, Cerebral dominance..., 1970); b) i fattori anatomici, come la possibile permanenza nei pazienti di Akelaitis di qualche fibra nella commissura anteriore o nel calloso, sono probabilmente molto importanti, quando si considerano i recenti dati di Gordon, Bogen e Sperry (v., 1971) che indicano una completa assenza di sintomi di disconnessione interemisferica in due soggetti con sezione della commissura anteriore, della commissura ippocampale e di tutto il corpo calloso eccetto lo splenio; c) i soggetti con cervello bisecato chirurgicamente o con agenesia callosale possono imparare ad usare nuove strategie di comunicazione interemisferica. Queste strategie potrebbero implicare l'uso delle rimanenti commissure o di altre vie interemisferiche (v. Trevarthen, 1970) o il meccanismo dei suggerimenti crociati periferici, considerato in dettaglio da Gazzaniga (v., 1970).
Un'altra linea di sviluppo sperimentale, indubbiamente ispirata dalle ricerche sul cervello bisecato, è lo studio delle comunicazioni interemisferiche in soggetti umani normali. Questo studio è stato eseguito essenzialmente con l'analisi dei tempi di reazione a stimoli visivi lateralizzati, cioè a stimoli proiettati a un singolo emisfero (v., ad es., Gazzaniga, 1970; v. Rizzolatti e altri, 1971; v. Geffen e altri, 1971). È possibile, per esempio, dimostrare che fotografie di facce sono riconosciute prima e producono risposte più veloci nel soggetto destrimane normale quando sono proiettate all'emisfero destro. Al contrario, gli stimoli di natura verbale sono riconosciuti prima e producono risposte più rapide quando sono proiettati all'emisfero sinistro (v. Rizzolatti e altri, 1971). Anche se non è ancora stato definitivamente provato che le differenze nel tempo di risposta fra i due emisferi riflettono effettivamente un processo di comunicazione interemisferica, sembra chiaro che questo approccio è un utile complemento alle ricerche sul cervello bisecato.
Una questione finale riguarda la possibilità che la bisezione cerebrale possa aumentare la capacità di analisi di informazioni da parte del sistema nervoso (v. Gazzaniga, 1970; v. Burešovà e Bureš, 1969). Esperimenti eseguiti su scimmie con cervello bisecato (v. Gazzaniga e Young, 1967) e su pazienti commissurotomizzati (v. Gazzaniga e Sperry, 1966) hanno dimostrato che i due emisferi divisi possono utilizzare informazioni visive diverse presentate simultaneamente ai due occhi (nella scimmia) e nei due emicampi visivi (nell'uomo). In queste condizioni sperimentali il cervello bisecato è più efficiente del cervello intatto, probabilmente perché ciascuno degli emisferi disconnessi agisce come se fosse un cervello separato. D'altra parte, dati della sperimentazione sugli animali (v. Meikle jr. e altri, 1962; v. Larsen jr. e altri, 1969; v. Robinson e Voneida, 1970; v. Sechzer, 1970) hanno indicato che la bisezione cerebrale disturba, come del resto ci si aspetterebbe, l'organizzazione nervosa, visto che l'apprendimento con un emisfero singolo può essere assai peggiore dell'apprendimento con i due emisferi normalmente connessi. C. B. Trevarthen (v., 1968 e 1970) ha anche suggerito che nei pazienti con cervello bisecato meccanismi unificanti sensoriali e percettivi, probabilmente localizzati nel mesencefalo, non sezionato, possano impedire l'attuarsi di una completa indipendenza interemisferica. Questa indipendenza è postulata con l'ipotesi secondo la quale l'analisi simultanea di informazioni visive diverse può avvenire negli emisferi separati dei soggetti con cervello bisecato.
5. Conclusione
Il vasto interesse contemporaneo nei meccanismi delle comunicazioni interemisferiche è testimoniato dalla seguente bibliografia, la cui mole è in larga misura dovuta a lavori pubblicati nell'ultimo ventennio. La presente rassegna, assieme a quella sulla dominanza cerebrale (v. emisferi cerebrali: Dominanza cerebrale), indica chiaramente che lo studio delle comunicazioni interemisferiche sta procedendo sia nel campo della scienza neurologica di base, fondata prevalentemente sulla sperimentazione sugli animali, sia in quello dell'analisi clinica e neuropsicologica sperimentale dell'uomo. Le ricerche morfologiche e fisiologiche continuano a rivelare l'ordine e la complessità delle connessioni interemisferiche e le influenze sinaptiche transcommissurali. Il trasferimento interemisferico di segnali di senso e di comandi motori, dimostrato dall'elettrofisiologia, può ora essere correlato con 4a trasmissione interemisferica di messaggi percettivi e mnemonici studiata negli esperimenti neurocomportamentali. L'affascinante analisi dei soggetti umani con cervello bisecato, intrapresa negli ultimi anni, non solo permette il confronto con esperimenti simili eseguiti su altri Mammiferi, ma fornisce anche un mezzo unico per esplorare le differenze fra l'emisfero destro e l'emisfero sinistro del cervello umano, e per dedurre i meccanismi commissurali che effettuano normalmente l'integrazione delle differenti capacità degli emisferi. Questo è chiaramente uno dei campi in cui la combinazione delle più svariate tecniche per studiare l'attività del sistema nervoso non solo ha dato risposte esaurienti a importantissime domande scientifiche, ma continua tuttora a presentare allo studioso del sistema nervoso aspetti nuovi e ugualmente interessanti dell'attività e dell'organizzazione del cervello.
bibliografia
Adrian, E. D., Yamagiwa, K., The origin of the Berger rhythm, in ‟Brain", 1935, LVIII, pp. 323-351.
Aird, R. B., Garoutte, B., Studies on the ‛cerebral pacemaker', in ‟Neurology", 1958, VIII, pp. 581-589.
Ajmone Marsan, C., Morillo, A., Cortical control and callosal mechanisms in the visual system of cat, in ‟Electroencephalography and clinical neurophysiology", 1961, XIII, pp. 553-563.
Ajmone Marsan, C., Morillo, A., Callosal and specific response in the visual cortex of cat, in ‟Archives italiennes de biologie", 1963, CI, pp. 1-29.
Akelaitis, A. J., A study of gnosis, praxis and language following section of the corpus callosum and anterior commissure, in ‟Journal of neurosurgery", 1944, I, pp. 94-101.
Andersen, P., Andersson, S. A., Physiological basis of the alpha rhythm, New York 1968.
Andersen, P., Andersson, S. A., Lømo, T., Nature of thalamocortical relations during spontaneous barbiturate spindles, in ‟Journal of physiology", 1967, CXCII, pp. 283-307.
Asanuma, H., Okamoto, K., Unitary study on evoked activity of callosal neurons and its effect on pyramidal tract cell activity in cats, in ‟Japanese journal of physiology", 1959, IX, pp. 473-483.
Asanuma, H., Okuda, O., Effects of transcallosal volleys on pyramidal tract cell activity of cat, in ‟Journal of neurophysiology", 1962, XXV, pp. 198-208.
Bailey, P., Garol, H. W., McCulloch, W. S., Cortical origin and distribution of corpus callosum and anterior commissure in chimpanzee (‛Pan satyrus'), in ‟Journal of neurophysiology", 1941, IV, pp. 564-571.
Barrett, T. W., Studies of the function of the amygdaloid complex in ‛Macaca mulatta', in ‟Neuropsychologia", 1969, VII, pp. 1-12.
Batini, C., Radulovacki, M., Kado, R. T., Adey, W. R., Effect of interhempispheric transection on the EEG patterns in sleep and wakefulness in monkeys, in ‟Electroencephalography and clinical neurophysiology", 1967, XXII, pp. 101-112.
Benevento, L. A., Ebner, F. F., The areas and layers of cortico-cortical terminations in the visual cortex of the Virginia opossum, in ‟Journal of comparative neurology", 1971, CXLI, pp. 157-184.
Berlucchi, G., Callosal activity in unrestrained, unanesthetized cat, in ‟Archives italiennes de biologie", 1965, CIII, pp. 623-634.
Berlucchi, G., Electroencephalographic studies in ‛split brain' cats, in ‟Electroencephalography and clinical neurophysiology", 1966, XX, pp. 348-356.
Berlucchi, G., Anatomical and physiological aspects of visual functions of corpus callosum, in ‟Brain research", 1972, XXXVII, pp. 371-392.
Berlucchi, G., Gazzaniga, M. S., Rizzolatti, G., Microelectrode analysis of transfer of visual information by the corpus callosum, in ‟Archives italiennes de biologie", 1967, CV, pp. 583-596.
Berlucchi, G., Rizzolatti, G., Binocularly driven neurons in visual cortex of split-chiasm cats, in ‟Science", 1968, CLIX, pp. 308-310.
Berlucchi, G., Sprague, J. M., Levy, J., Di Berardino, A., The pretectum and superior colliculus in visually guided behavior and in flux and form discrimination in the cat, in ‟Journal of comparative and physiological psychology", 1972, LXXVIII, 123-172.
Bishop, P. O., Neurophysiology of binocular single vision and stereopsis, in Handbook of sensory physiology (a cura di R. Jung), vol. VII, Berlin 1973, pp. 255-305.
Black, P., Myers, R. E., Visual function of the forebrain commissures in the chimpanzee, in ‟Science", 1964, CXLVI, pp. 799-800.
Black, P., Myers, R. E., Visual function of the forebrain commissures in the chimpanzee, in ‟Science", 1964, CXLVI, pp. 799-800.
Black, P., Myers, R. E., Brainsistem mediation of visual perception in a higher primate, in ‟Transactions of the American Neurological Association", 1968, XCIII, pp. 191-193.
Blakemore, C., Binocular depth perception and the optic chiasm, in ‟Vision research", 1970, X, pp. 43-47.
Blinkov, S. M., Glezer, I. I., The human brain in figures and tables. A quantitative handbook, New York 1968.
Bogen, J. E., The other side of the brain. I. Dysgraphya and dyscopia following cerebral commissurotomy, in ‟Bulletin of the Los Angeles Neurological Society", 1969, XXXIV, pp. 73-105.
Bogen, J. E., The other side of the brain. II. An appositional mind, in ‟Bulletin of the Los Angeles Neurological Society", 1969, XXXIV, pp. 135-162.
Bogen, J. E., Bogen, G. M., The other side of the brain. III. The corpus callosum and creativity, in ‟Bulletin of the Los Angeles Neurological Society", 1969, XXXIV, pp. 191-220.
Bogen, J. E., Sperry, R. W., Vogel, J. P., Commissural section and propagation of seizures, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper, A. A. Ward e A. Pope), Boston 1969, pp. 439-440.
Bogen, J. E., Vogel, P. J., Cerebral commissurotomy in man. Preliminary case report, in ‟Bulletin of the Los Angeles Neurological Society", 1962, XXVII, pp. 169-172.
Bremer, F., Un aspect de la physiologie du corps calleux, in ‟Archives internationales de physiologie", 1953, LXI, pp. 110-112.
Bremer, F., Response of visual area to callosal impulses in the cat, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1955, XC, pp. 22-25.
Bremer, F., Physiology of the corpus callosum, in ‟Research pubblications. Association for Research in Nervous and Mental Diseases", 1958, XXXVI, pp. 424-448.
Bremer, F., Étude électrophysiologique d'un transfert interhémisphérique callosal, in ‟Archives italiennes de biologie", 1966, CIV, pp. 1-29.
Bremer, F., Les corps calleux dans la dynamique cérébrale, in ‟Experientia", 1966, XXII, pp. 201-208.
Bremer, F., Bryhaye, J., André-Balisaux, G., Physiologie et pathologie du corps calleux, in ‟Schweizer Arciv für Neurologie und Psychiatrie", 1956, LXXVIII, pp. 31-87.
Bremer, F., Stoupel, N., Transmission interhémisphérique des influx visuels par le corps calleux, in ‟Journal de physiologie", 1956, XLVIII, pp. 411-414.
Bremer, F., Stoupel, N., Étude des mécanismes de la synergie bioélectrique des hémisphères cérébraux, in ‟Acta physiologica et pharmacologica neerlandica", 1957, VI, pp. 487-496.
Bremer, F., Terzuolo, C., Transfert interhémisphérique d'informations sensorielles par le corps calleux, in ‟Journal de physiologie", 1955, XLVII, pp. 105-107.
Bures, J., Buresová, O., The use of Leao's spreading depression in the study of interhemispheric transfer of memory traces, in ‟Journal of comparative and physiological psychology", 1960, LIII, pp. 558-563.
Burešová, O., Bures, J., Can the brain be improved?, in ‟Endeavour", 1969, XXVIII, pp. 139-145.
Burešová, O., Bures, J., Rustová, M., Conditions for interhemispheric transfer of initially lateralized visual engrams in hooded rats, in ‟Journal of comparative and physiological psychology", 1971, LXXV, pp. 200-205.
Butler, C. R., A memory-record for visual discrimination habits produced in both cerebral hemispheres of monkey when only one hemisphere has received direct visual information, in ‟Brain research", 1968, X, pp. 152-167.
Butler, C. R., Francis, A. C., Split-brain behaviour without splitting. Tactile discriminations in monkeys, in ‟Israel journal of medical sciences", 1973, IX (Suppl.), pp. 79-84.
Bykoff, K., Versuche an Hunden mit Durchschneiden des Corpus callosum, in ‟Zentralblatt für die gesamte Neurologie und Psychiatrie", 1924, XXXIX, p. 199.
Cajal, S. R., Histologie du système nerveux de l'homme et des vertébrés, 2 voll., Paris 1911 (reprint, Madrid 1955).
Carman, J. B., Cowan, W. M., Powell, T. P. S., Webster, K. E., A bilateral cortico-striate projection, in ‟Journal of neurology, neurosurgery and psychiatry", 1965, XXVIII, pp. 71-77.
Catania, A. C., Interocular transfer of discriminations in the pigeon, in ‟Journal of experimental analysis and behavior", 1965, VIII, pp. 147-155.
Chang, H. T., Cortical response to activity of callosal neurons, in ‟Journal of neurophysiology", 1953, XVI, pp. 117-131.
Chang, H. T., Interaction of evoked cortical potentials, in ‟Journal of neurophysiology", 1953, XVI, pp. 133-144.
Choudhury, B. P., Whitteridge, D., Wilson, M. E., The function of the callosal connections of the visual cortex, in ‟Quarterly journal of experimental physiology", 1965, L, pp. 214-219.
Claes, E., Contribution a l'étude physiologique de la fonction visuelle. I. Analyse oscillographique de l'activité spontanée et sensorielle de l'aire visuelle cortical chez le chat non anesthésié, in ‟Archives internationales de physiologie", 1939, XLVIII, pp. 181-237.
Claire, H. M., Landau, W. M., Bishop, G. H., The cortical response to direct stimulation of the corpus callosum in the cat, in ‟Electroencephalography and clinical neurophysiology", 1961, XIII, pp. 21-33.
Cragg, B. G., Ainsworth, A., The topography of the afferent projections in the circumstriate cortex of the monkey studied by the Nauta method, in ‟Vision research", 1969, IX, pp. 733-747.
Cuénod, M., Split brain studies. Functional interaction between bilateral central nervous structures, in The structure and function of nervous tissue (a cura di G. H. Bourne), vol. V, New York 1972, pp. 455-506.
Cuénod, M., Zeier, H., Transfert interhemisphérique et commissurotomic chez le pigeon, in ‟Schweizer Archiv für Neurologie, Neurochirurgie und Psychiatrie", 1967, C, pp. 365-380.
Curtis, H. J., Intercortical connections of corpus callosum as indicated by evoked potentials, in ‟Journal of neurophysiology", 1940, III, pp. 407-413.
Curtis, H. J., An analysis of cortical potentials mediated by the corpus callosum, in ‟Journal of neurophysiology", 1940, III, pp. 414-422.
D'Elia, G. (a cura di), Unilateral electroconvulsive therapy, in ‟Acta psychiatrica scandinavica", 1970, suppl. 215, pp. 1-98.
Diamond, I. T., Jones, E. G., Powell, T. P. S., Interhemispheric fiber connections of the auditory cortex of the cat, in ‟Brain research", 1968, XI, pp. 177-193.
Doty, R. W., Negrão, N., Forebrain commissures and vision, in Handbook of sensory physiology (a cura di R. Jung), vol. VII, Berlin 1973, pp. 543-582.
Dow, B. M., Dubner, R., Single unit responses to moving visual stimuli in middle suprasylvian gyrus of the cat, in ‟Journal of neurophysiology", 1971, XXXIV, pp. 47-55.
Downer, J. L. de C., Changes in visually guided behavior following midsagittal division of optic chiasm and corpus callosum in monkey (Macaca mulatta'), in ‟Brain", 1959, LXXXII, pp. 251-259.
Downer, J. L. de C., Interhemispheric integration in the visual system, in Interhemispheric relations and cerebral dominance (a cura di V. B. Mountcastle), Baltimore 1962, pp. 87-100.
Ebner, F. F., Myers R. E., Corpus callosum and the interhemispheric transmission of tactual learning, in ‟Journal of neurophysiology", 1962, XXV, pp. 380-391.
Ebner, F. F., Myers, R. E., Distribution of corpus callosum and anterior commissure in cat and raccoon, in ‟Journal of comparative neurology", 1965, CXXIV, pp. 353-375.
Eccles, J. C., The brain and the unity of conscious experience (Eddington lecture), London 1965.
Eidelberg, E., Callosal and non-callosal connexions between the sensor-motor cortices in cat and monkey, in ‟Electroencephalography and clinical neurophysiology", 1969, XXVI, pp. 557-564.
Enomoto, T. F., Unilateral activation of the non-specific thalamic system and bilateral cortical responses, in ‟Electroencephalography and clinical neurophysiology", 1959, XI, pp. 219-232.
Erickson, T. C., Spread of the epileptic discharge. An experimental study of the after-discharge induced by electrical stimulation of the cerebral cortex, in ‟Archives of neurology and psychiatry", 1940, XLIII, pp. 429-452.
Ettlinger, G., Blakemore, C. B., The behavioral effects of commissural section, in Contribution to clinical neuropsychology (a cura di A. L. Benton), Chicago 1969, pp. 30-72.
Ettlinger, G., Morton, H. B., Tactile discrimination performance in the monkey. Transfer of training between the hands after commissural section, in ‟Cortex", 1966, II, pp. 30-49.
Feeney, D. M., Orem, J. M., Influence of antidromic callosal volleys on single units in visual cortex, in ‟Experimental neurology", 1971, XXXIII, pp. 310-321.
Flechsig, P., Gehirn und Seele, Leipzig 1896.
Garey, L.J., Jones, E. G., Powell, T. P. S., Interrelationships of striate and exstrastriate cortex with the primary relay sites of the visual cortex, in ‟Journal of neurology, neurosurgery, psichiatry", 1968, XXXI, pp. 135-157.
Garey, L. J., Powell, T. P. S., An experimental study of the termination of the lateral geniculo-cortical pathway in the cat visual cortex, in ‟Proceedings of the Royal Society" (Series B), 1971, CLXXIX, pp. 41-63.
Garol, H. W., Cortical origin and distribution of corpus callosum and anterior commissure in the cat, in ‟Journal of neuropathology and experimental neurology", 1942, I, pp. 422-429.
Gavalas, R. J., Sperry, R. W., Central integration of visual halffields in split-brain monkeys, in ‟Brain research", 1969, XV, pp. 97-106.
Gazzaniga, M. S., Interhemispheric communication of visual learning, in ‟Neuropsychologia", 1966, IV, pp. 183-189.
Gazzaniga, M. S., The bisected brain, New York 1970.
Gazzaniga, M. S., One brain-two minds?, in ‟American scientist", 1972, LX, pp. 311-317.
Gazzaniga, M. S., Bogen, J. E., Sperry, R. W., Observations on visual perception after disconnexion of the cerebral hemispheres in man, in ‟Brain", 1965, LXXXVIII, pp. 221-236.
Gazzaniga, M. S., Bogen, J. E., Sperry, R. W., Dyspraxia following division of the cerebral commissures, in ‟Archives of neurology", 1967, XVI, pp. 606-612.
Gazzaniga, M. S., Hillyard, S. A., Language and speech capacity of the right hemisphere, in ‟Neuropsychologia", 1971, IX, pp. 273-280.
Gazzaniga, M. S., Sperry, R. W., Simultaneous double discrimination responses following brain bisection, in ‟Psychonomic science", 1966, IV, pp. 261-262.
Gazzaniga, M. S., Young, E. D., Effects of commissurotomy on the processing of increasing visual information, in ‟Experimental brain research", 1967, III, pp. 368-371.
Geffen, G., Bradshaw, J. L., Wallace, G., Interhemispheric effects on reaction time to verbal and nonverbal visual stimuli, in ‟Journal of experimental psychology", 1971, LXXXVII, pp. 415-422.
Geschwind, N., Disconnexion syndromes in animals and man, in ‟Brain", 1965, LXXXVIII, pp. 237-294.
Giaquinto, S., Effects of medial thalamic stimulations in chronic split-brain cats, in ‟Electroencephalography and clinical neurophysiology", 1969, XXVI, pp. 50-61.
Glickstein, M., Sperry, R. W., Intermanual somesthetic transfer in split-brain Rhesus monkey, in ‟Journal of comparative and physiological psychology", 1960, LIII, ppp. 322-327.
Globus, A., Scheibel, A. B., Synaptic loci on parietal cortical neurons. Termination of corpus callosum fibers, in ‟Science", 1967, CLVI, pp. 1127-1129.
Gordon, H. W., Bogen, J. E., Sperry, R. W., Absence of deconnexion syndrome in two patients with partial section of the neocommissures, in ‟Brain", 1971, XCIV, pp. 327-336.
Gordon, H. W., Sperry, R. W., Lateralization of olfactory perception in the surgically separated hemispheres in man, in ‟Neuropsychologia", 1969, VII, pp. 111-120.
Gozzano, M., Ricerche sui fenomeni elettrici della corteccia cerebrale, in ‟Rivista di neurologia", 1935, VIII, pp. 212-261.
Grafstein, B., Organization of callosal connections in suprasylvian gyrus of cat, in ‟Journal of neurophysiology", 1959, XXII, pp. 504-515.
Grafstein, B., Postnatal development of the transcallosal evoked response in the cerebral cortex of the cat, in ‟Journal of neurophysiology", 1963, XXVI, pp. 79-99.
Hamilton, C. R., Effects of brain bisection on eye-hand coordination in monkeys wearing prisms, in ‟Journal of comparative and physiological psychology", 1967, LXIV, pp. 434-443.
Hamilton, C. R., Gazzaniga, M. S., Lateralization of learning of colour and brightness discriminations following brain bisection, in ‟Nature", 1964, CCI, p. 220.
Hamilton, C. R., Hillyard, S. A., Sperry, R. W., Interhemispheric comparison of color in split-brain monkeys, in ‟Experimental neurology", 1968, XXI, pp. 486-494.
Hamilton, C. R., Lund, J. S., Visual discrimination of movement. Midbrain or forebrain?, in ‟Science", 1970, CLXX, pp. 1428-1430.
Heath, C. J., Jones, E. G., Connexions of area 19 and the lateral suprasylvian area of the visual cortex of the cat, in ‟Brain research", 1970, XIX, pp. 302-305.
Hécaen, H., Assal, G., Les relations interhémisphériques et le problème de la dominance cérébrale d'après les recherches sur les setions calleuses chez l'animal et chez l'homme, in ‟Année psychologique", 1968, LXVIII, pp. 491-523.
Heimer, L., Ebner, F. F., Nauta, W. J. H., A note on the termination of commissural fibers in the neocortex, in ‟Brain resarch", 1967, V, pp. 171-177.
Hoefer, P. F. A., Pool, J. L., Conduction of cortical impulses and motor management of convulsive seizures, in ‟Archives of neurology and psychiatry, Chicago", 1943, L, pp. 381-400.
Horel, J. A., Keating, E. G., Partial Klüver-Bucy syndrome produced by cortical disconnection, in ‟Brain research", 1969, XVI, pp. 281-284.
Hossman, K. A., Midline section of the cat brain with sparing of the corpus callosum, in ‟Electroencephalography and clinical neurophysiology", 1969, XXVI, pp. 627-629.
Hossmann, K. A., Untersuchungen über transcallosale Potentiale an der akuten Corpus Callosum-Katze, in ‟Deutsche Zeitschrift für Nervenheilkunde", 1969, CXCV, pp. 79-102.
Hubel, D. H., Wiesel, T. N., Receptive fields and functional architecture in two nonstriate visual areas (18 and 197 of the cat), in ‟Journal of neurophysiology", 1965, XXVIII, pp. 229-289.
Hubel, D. H., Wiesel, T. N., Cortical and callosal connections concerned with the vertical meridian of visual fields in the cat, in ‟Journal of neurophysiology", 1967, XXX, pp. 1561-1573.
Hughes, A., Wilson, M. E., Callosal termination along the boundary between visual areas I and II in the rabbit, in ‟Brain research", 1969, XII, pp. 19-25.
Ingle, D., Interocular integration of visual learning by gold-fish, in ‟Brain, behavior and evolution", 1968, I, pp. 58-85.
Innocenti, G. M., Manzoni, T., Spidalieri, G., Peripheral and transcallosal reactivity of neurones within SI and SII cortical areas (segmental divisions), in ‟Archives italiennes de biologie", 1972, CX, pp. 415-443.
Jacobson, S., Intralaminar, interlaminar, callosal and thalamo-cortical connections in frontal and parietal areas of the albino rat cerebral cortex, in ‟Journal of comparative neurology", 1965, CXXIV, pp. 131-145.
Jacobson, S., Distribution of commissural axon terminals in the rat neocortex, in ‟Experimental neurology", 1970, XXVIII, pp. 193-205.
Jacobson, S., Marcus, E. M., The laminar distribution of fibers of the corpus callosum. A comparative study in the rat, cat, Rhesus monkey and chimpanzee, in ‟Brain research", 1970, XXIV, pp. 517-520.
Jones, E. G., Powell, T. P. S., The commissural connexions of the somatic sensory cortex in the cat, in ‟Journal of anatomy", 1968, CIII, pp. 433-455.
Jones, E. G., Powell, T. P. S., Connexions of the somatic sensory cortex of the Rhesus monkey. II. Contralateral cortical connexions, in ‟Brain", 1969, XCII, pp. 717-730.
Jones, E. G., Powell, T. P. S., An electron microscopic study of the laminar pattern and mode of termination of afferent fibre pathways in the somatic sensory cortex of the cat, in ‟Philosophical transactions of the Royal Society" (Series B), 1970, CCLVII, pp. 45-62.
Kaas, J., Axelrod, S., Diamond, I. T., An ablation study of the auditory cortex in the cat using binaural tonal patterns, in ‟Journal of neurophysiology", 1967, XXX, pp. 710-724.
Karol, E. A., Pandya, D. N., The distribution of the corpus callosum in the Rhesus monkey, in ‟Brain", 1971, XCIV, pp. 471-486.
Kawamura, K, Otani, N., Corticocortical fiber connections in the cerebrum. The frontal region, in ‟Journal of comparative neurology", 1970, CXXXIX, pp. 434-448.
Kohn, B., Myers, R. E., Visual information and intermanual transfer of latch box problem solving in monkeys with commissures sectioned, in ‟Experimental neurology", 1969, XXIII, pp. 303-309.
Kopeloff, N., Kennard, M. A., Pacella, B. L., Kopeloff, L. M., Chusid, J. G., Section of corpus callosum in experimental epilepsy in the monkey, in ‟Archives of neurology and psychiatry", 1950, LXIII, pp. 719-727.
Landau, W. M., Bishop, G. H., Clare, M. H., The interaction of several varieties of evoked responses in visual and association cortex of the cat, in ‟Electroencephalography and clinical neurophysiology", 1961, XIII, pp. 43-53.
Larsen, J. W. Jr., Winans, S. S., Meikle, T. H. Jr., The effect of forebrain commissurotomies on the rate of learning a dark-light discrimination by cats, in ‟Brain research", 1969, XIV, pp. 717-731.
Latimer, C. N., Kennedy, T. T., Cortical unit activity following transcallosal volleys, in ‟Journal of neurophysiology", 1961, XXIV, pp. 66-79.
Latimer, C. N., Wilfred, M. C., Motor, sensory and callosal responses from cortex of Cercopithecus aethiops, in ‟Physiology and behavior", 1968, III, pp. 687-692.
Lee-Teng, E., Sperry, R. W., Intermanual stereognostic size discrimination in split-brain monkeys, in ‟Journal of comparative and physiological psychology", 1966, LXII, pp. 84-89.
Lehman, R. A. W., Motor coordination and hand preference after lesions of the visual pathway and corpus callosum, in ‟Brain", 1968, XCI, pp. 525-538.
Leibowitz, U., Interhemispheric spread of drug-induced electrical cortical activity, in ‟Electroencephalography and clinical neurophysiology", 1969, XXVI, pp. 515-523.
Levine, J., Studies in the interrelations of central nervous structures in binocular vision. II. The conditions under which interocular transfer of discriminative habits takes place in the pigeon, in ‟Journal of genetic psychology", 1945, LXVII, pp. 131-142.
Levinson, D. M., Interocular transfer in guinea pigs following section of the corpus callosum, in ‟Journal of comparative and physiological psychology", 1972, LXXVIII, pp. 26-31.
Locke, S., Kruper, D. C., Yakovlev, P. I., Limbic nuclei of thalamus and connections of limbic cortex. VII. Transcallosal connections of cerebral hemisphere with striatum in monkey and man, in ‟Archives of neurology", 1964, XI, pp. 571-582.
Locke, S., Yakovlev, P. I., Transcallosal connections of the cingulum in man, in ‟Archives of neurology", 1965, XIII, pp. 471-476.
Lund, J. S., Adaptation to visual field displacement by monkeys with optic chiasm sectioned, in ‟Experimental neurology", 1970, XXVII, pp. 334-343.
Lund, J. S., Lund, R. D., The termination of callosal fibers in the paravisual cortex of the rat, in ‟Brain research", 1970, XVII, pp. 25-45.
McCleary, R. A., Type of response as a factor in interocular transfer in the fish, in ‟Journal of comparative and physiological psychology", 1960, LIII, pp. 311-321.
McCulloch, W. S., Garol, H. W., Cortical origin and distribution of corpus callosum and anterior commissure in the monkey (‛Macaca mulatta'), in ‟Journal of neurophysiology", 1941, IV, pp. 555-563.
MacKay, D. M., Discussion of Sperry's paper, in Brain and conscious experience (a cura di J. C. Eccles), New York 1966, pp. 312-313.
Magni, F., Melzack, R., Smith, L. J., A stereotaxic method for sectioning the corpus callosum of the cat, in ‟Electroencephalography and clinical neurophysiology", 1960, XII, pp. 517-518.
Majkowski, J., Electrophysiological studies of learning in split-brain cats, in ‟Electroencephalography and clinical neurophysiology", 1967, XXIII, pp. 521-531.
Makarov, F. N., Commissural connections of the cat visual cortex (areas 17, 18, and 19), in ‟Neuroscience translations", 1968-1969, VII, pp. 785-790.
Mancia, M., Desirau, T., Chhina, G. S., The monkey split brainstem. Effects on the sleep-wakefulness cycle, in ‟Electroencephalography and clinical neurophysiology", 1968, XXIV, pp. 409-416.
Marcus, E. M., Watson, C. W., Bilateral synchronous spike wave electrographic patterns in the cat, in ‟Archives of neurology", 1966, XIV, pp. 601-610.
Marcus, E. M., Watson, C. W., Symmetrical epileptogenic foci in monkey cerebral cortex, in ‟Archives of neurology", 1968, XIX, pp. 99-116.
Marcus, E. M., Watson, C. W., Simon, S. A., An experimental model of some varieties of petit mal epilepsy. Electrical-behavioral correlations of acute bilateral epilpetogenic foci in cerebral cortex, in ‟Epilepsia", 1968, IX, pp. 233-248.
Mark, R. F., The tectal commissure and interocular transfer of pattern discrimination in cichlid fish, in ‟Experimental neurology", 1966, XVI, pp. 215-225.
Mark, R. F., Sperry, R. W., Bimanual coordination in monkeys, in ‟Experimental neurology", 1968, XX, pp. 92-104.
Maspes, P. E., La syndrome expérimental chez l'homme de la section du splénium du corps calleux. Alexie visuelle pure hémianopsique, in ‟Revue neurologique, 1948, LXXX, pp. 100-113.
Meier, R. E., Interhemisphärischen Transfer visueller Zweifachwahlen bei commissureotomierten Taube, in ‟Phychologische Forschung", 1971, XXXIV, pp. 220-245.
Meikle, T. H. Jr., Failure of interocular transfer of brightness discrimination, in ‟Nature", 1964, CCII, pp. 1243-1244.
Meikle, T. H. Jr., Seczher, J. A., Interocular transfer of brightness discrimination in ‛split-brain' cats, in ‟Science", 1960, CXXXII, pp. 734-835.
Meikle, T. H. Jr., Sechzer, J. A., Stellar, E., Interhemispheric transfer of tactile conditioned responses in corpus callosum sectioned cats, in ‟Journal of neurophysiology", 1962, XXV, pp. 530-542.
Mettler, F. A., Corticofugal fiber connections of the cortex of ‛Macaca mulatta'. The occipital region, in ‟Journal of comparative neurology", 1935, LXI, pp. 221-256.
Mettler, F. A., Corticofugal fiber connections of the cortex of ‛Macaca mulatta'. The frontal region, in ‟Journal of comparative neurology", 1935, LXI, pp. 509-542.
Mettler, F. A., Corticofugal fiber connections of the cortex of ‛Macaca mulatta'. The parietal region, in ‟Journal of comparative neurology", 1935, LXII, pp. 263-291.
Meyerson, B. A., Ontogeny of interhemispheric functions. An electrophysiological study in pre- and postnatal sheep, in ‟Acta physiological scandinavica", 1968, suppl. 312, pp. 1-111.
Michel, F., Roffwarg, M. P., Chronic split brain-stem preparation. Effect on the sleep-waking cycle, in ‟Experientia", 1967, XXIII, pp. 126-128.
Milner, B., Taylor, L., Sperry, R. W., Lateralized suppression of dichotically presented digits after commissural section in man, in ‟Science", 1968, CLXI, pp. 184-186.
Mingazzini, G., Der Balken. Eine anatomischen, physiopathologischen und klinische Studie, Berlin 1922.
Mishkin, M., Visual mechanisms beyond the striate cortex, in Frontiers in physiological psychology (a cura di R. W. Russell), New York 1966, pp. 93-119.
Mishkin, M., Cortical visual areas and their interactions, in Brain and human behavior (a cura di A. G. Karczman e J. C. Eccles), Berlin 1972, pp. 187-208.
Mitchell, D. E., Blakemore, C., Binocular depth perception and the corpus callosum, in ‟Vision research", 1970, X, pp. 49-54.
Morrell, F., Secondary epileptogenic lesions, in ‟Epilepsia", 1960, I, pp. 538-560.
Morse, R. W., Vargo, R. A., Functional neuronal subsets in the forepaw focus of somatosensory area II of the cat, in ‟Experimental neurology", 1970, XXVII, pp. 125-138.
Moruzzi, G., Contribution à l'electrophysiologie du cortex moteur. Facilitation, after-discharge et épilepsie corticale, in ‟Archives internationales de physiologie", 1939, IL, pp. 33-100.
Moruzzi, G., L'épilepsie expérimentale, Paris 1950.
Mosidze, V. M., Rishinashvili, R. S., Totibadze, N. K., Kenavishvili, Z. S., Akbardia, K. K., Some results of studies on split brain, in ‟Physiology and behavior", 1971, VII, pp. 763-772.
Myers, R. E., Function of corpus callosum in interocular transfer, in ‟Brain", 1956, LXXIX, pp. 358-363.
Myers, R. R., Localization of function in the corpus callosum. Visual gnostic transfer, in ‟Archives of neurology", 1959, I, pp. 74-77.
Myers, R. E., Corpus callosum and visual gnosis, in Brain mechanisms and learning (a cura di J. F. Delafresnaye), Oxford 1961, pp. 481-505.
Myers, R. E., Commissural connections between occipital lobes of the monkey, in ‟Journal of comparative neurology", 1962, CXVIII, pp. 1-16.
Myers, R. E., Transmission of visual information within and between the hemispheres. A behavioral study, in Interhemispheric relations and cerebral dominance (a cura di V. B. Mountcastle), Baltimore 1962, pp. 51-73.
Myers, R. E., Henson, C. O., Role of corpus callosum in transfer of tactuokinesthetic learning in chimpanzee, in ‟Archives of neurology", 1960, III, pp. 404-409.
Myers, R. E., Sperry, R. W., Interhemispheric communication through the corpus callosum. Mnemonic carry-over between the hemispheres, in ‟Archives of neurology and psychiatry", 1958, LXXX, pp. 298-303.
Myers, R. E., Sperry, R. W., McCurdy, N. M., Neural mechanisms in visual guidance of limb movement, in ‟Archives of neurology", 1962, VII, pp. 195-202.
Nadel, L., Interhemispheric transfer. Monocular input and varied sensory conditions, in ‟Physiology and behavior", 1971, VI, pp. 655-661.
Naito, H., Miyakawa, F., Ito, N., Diameter of callosal fibers interconnecting cat sensorimotor cortex, in ‟Brain research", 1971, XXVII, pp. 396-372.
Naito, H., Nakamura, K., Kurosaki, T., Tamura, Y., Transcallosal excitatory postsynaptic potentials of fast and slow pyramidal tract cells in cat sensorimotor cortex, in ‟Brain research", 1970, XIX, pp. 299-301.
Nakamura, K., Naito, H., Kurasaki, T., Tamura, Y., Effect of polarizing currents ont ranscallosal postsynaptic potentials of cat pyramidal tract cells, in ‟Brain research", 1971, XXXV, pp. 547-550.
Nauta, W. J. H., Bucher, V. M., Efferent connections of the striate cortex in the albino rat, in ‟Journal of comparative neurology", 1954, C, pp. 257-286.
Neff, W. D., Neural mechanisms of auditory discrimination, in Sensory communication (a cura di W. A. Rosenblith), New York-London 1961, pp. 259-278.
Nelson, L. R., Lende, R. A., Interhemispheric responses in the opossum, in ‟Journal of neurophysiology", 1965, XXVIII, pp. 189-199.
Noble, J., Paradoxical interocular transfer of mirror-image discrimination in the optic chiasm sectioned monkey, in ‟Brain research", 1968, X, pp. 127-151.
Nyquist, J. K., Towe, A. L., Neuronal activity evoked in cat precruciate cerebral cortex by cutaneous stimulation, in ‟Experimental neurology", 1970, XXIX, pp. 494-512.
Ottino, C. A., Meglio, M., Rossi, G. F., Tercero, E., An experimental study of the structures mediating bilateral synchrony of epileptic discharges of cortical origin, in ‟Epilepsia", 1971, XII, pp. 299-311.
Pandya, D. N., Gold, D., Berger, T., Interhemispheric connections of the precentral motor cortex in the Rhesus monkey, in ‟Brain research", 1969, XV, pp. 594-596.
Pandya, D. N., Hallet, M., Mukherjee, S. K., Intra- and interhemispheric connections of the neocortical auditory system in the Rhesus monkey, in ‟Brain research", 1969, XIV, pp. 49-65.
Pandya, D. N., Karol, E. A., Heilbronn, D., The topographical distribution of interhemispheric projections in the corpus callosum of the Rhesus monkey, in ‟Brain research", 1971, XXXII, pp. 31-43.
Pandya, D. N., Vignolo, L. A., Interhemispheric projections of the parietal lobe in the Rhesus monkey, in ‟Brain research", 1969, XV, pp. 49-65.
Patton, H. D., Towe, A. L., Kennedy, T. T., Activation of pyramidal tract neurons by ipsilateral cutaneous stimuli, in ‟Journal of neurophysiology", 1962, XXV, pp. 501-514.
Pavlov, I. P., Conditioned reflexes. An investigation of the physiological activity of the cerebral cortex, Oxford 1927 (reprint, New York 1960).
Peacock. S.M. Jr., Activity of anterior suprasylvian gyrus in response to transcallosal afferent volleys, in ‟Journal of neurophysiology", 1957, XX, pp. 140-155.
Pines, L. J., Maiman, R. M., Cells of origin of fibers of corpus callosum. Experimental and pathologic observations, in ‟Archives of neurology and psychiatry", 1939, XLII, pp. 1076-1081.
Poblete, R., Ruben, R. J., Walker, A. E., Propagation of after discharge between temporal lobes, in ‟Journal of neurophysiology", 1959, XXII, pp. 538-553.
Polyak, S., An experimental study of the association, callosal and projection fibers of the cerebral cortex of the cat, in ‟Journal of comparative neurology", 1927, XLIV, pp. 197-258.
Purpura, D. P., Girado, M., Synaptic mechanisms involved in transcallosal activation of cortico-spinal neurons, in ‟Archives italiennes de biologie", 1959, XCVII, pp. 111-139.
Putnam, S. J., Megirian, D., Manning, J. W., Marsupial interhemispheric relation, in ‟Journal of comparative neurology", 1968, CXXXII, pp. 227-233.
Rizzolatti, G., Umiltà, C., Berlucchi, G., Opposite superiorities of the right and left cerebral hemispheres in discriminative reaction time to physiognomical and alphabetical material, in ‟Brain", 1971, XCIV, pp. 431-442.
Robinson, D. L., Doty, R. W., Transcallosal effects on single units in second somatosensory area (S II) of the cat, in ‟Federation proceedings", 1972, XXXI, p. 303.
Robinson, J. S., Voneida, T. J., Central cross-integration of visual inputs presented simultaneously to the separate eyes, in ‟Journal of comparative and physiological psychology", 1964, LVII, pp. 22-28.
Robinson, J. S., Voneida, T. J., Quantitative differences in performance on abstract discriminations using one or both hemispheres, in ‟Experimental neurology", 1970, XXVI, pp. 72-83.
Russell, I. S., Neurological basis of complex learning, in ‟British medical bulletin", 1971, XXVII, pp. 278-285.
Rutledge, L. T., Kennedy, T. T., Extracallosal delayed responses to cortical stimulation in chloralosed cat, in ‟Journal of neurophysiology", 1960, XXIII, pp. 188-196.
Schrier, A. M., Sperry, R. W., Visuo-motor integration in split-braint cats, in ‟Science", 1959, CXXIX, pp. 1275-1276.
Schulte, A., Transfer-und Transposition-Versuche mit monokular dressierten Fischen, in ‟Zeitschrift für vergleichende Physiologie", 1957, XXXIX, ppp. 432-476.
Sechzer, J. A., Successful interocular transfer of pattern discrimination in ‛split-brain' cats with shock-avoidance motivation, in ‟Journal of comparative and physiological psychology", 1964, LVIII, pp. 76-93.
Sechzer, J. A., Successful interocular transfer of pattern discrimination in ‛split-brain' cats with shock-avoidance motivation, in ‟Journal of comparative and physiological psychology", 1964, LVIII, pp. 76-83.
Sechzer, J. A., Prolonged learning and split-brain cats, in ‟Science", 1970, CLXIX, pp. 889-892.
Sheridan, C. L., Interocular transfer of brightness and pattern discriminations in normal and corpus callosum sectioned rats, in ‟Journal of comparative and physiological psychology", 1965, LIX, pp. 292-294.
Sherrington, C. S., The integrative action of the nervous system, New Haven 1906 (reprint, London 1910).
Singer, W., Creutzfeldt, O., Die Bedeutung der Vorderhinkommissuren für die Koordination bilateraler EEG-Muster, in ‟Experimental brain research", 1969, VII, pp. 195-213.
Smith, K. U., Experimental analysis of the associative mechanism of the human brain in learning functions, in ‟Journal of comparative and physiological psychology", 1952, XLV, pp. 66-72.
Spatz, W. B., Tigges, J., Tigges, M., Subcortical projections, cortical associations and some intrinsic interlaminar connections in the striate cortex in the squirrel monkey (Saimiri'), in ‟Journal of comparative neurology", 1970, CXL, pp. 155-174.
Sperry, R. W., Cerebral organization and behavior, in ‟Science", 1961, CXXXIII, pp. 1747-1757.
Sperry, R. W., Some developments in brain lesion studies of learning, in ‟Vederation proceedings", 1961, XX, pp. 609-616.
Sperry, R. W., Brain bisection and mechanisms of consciousness, in Brain and conscious experience (a cura di J. C. Eccles), New York 1966, pp. 298-314.
Sperry, R. W., Hemisphere deconnection and unity in conscious awareness, in ‟American psychologist", 1968, XXIII, pp. 723-833.
Sperry, R. W., Mental unity following surgical disconnection of the cerebral hemispheres, in ‟Harvey lectures", 1968, LXII, pp. 293-323.
Sperry, R.W., Cerebral dominance in perception, in Early experience in visual information processing in perceptual and reading disorders (a cura di F. A. Young e D. B. Linbley), Washington 1970, pp. 167-178.
Sperry, R. W., Perception in the absence of the neocortical commissures, in ‟Research publications. Association for Research in Nervous and Mental Diseases", 1970, XLVIII, pp. 123-138.
Sperry, R. W., Clark, E., Intercular transfer of visual discrimination habits in a teleost fish, in ‟Physiological zoology", 1949, XXII, pp. 372-378.
Sperry, R. W., Myers, R. E., Schrier, A. M., Perceptual capacity of the isolated visual cortex in the cat, in ‟Quarterly journal of experimental psychology", 1960, VII, pp. 65-71.
Sperry, R. W., Stamm, J. S., Miner, N., Relearning tests for interocular transfer following division of optic chiasm and corpus callosum in cats, in ‟Journal of comparative and physiological psychology", 1956, IL, pp. 529-533.
Stamm, J. S., Sperry, R. W., Function of corpus callosum in contralateral transfer of somesthetic discrimination in cats, in ‟Journal of comparative and physiological psychology", 1957, L, pp. 138-143.
Stavraky, G. W., Supersensitivity following lesions of the nervous system. An aspect of the relativity of nervous integration, Toronto 1961.
Straw, R. N., Mitchell, C.L., Effect of section of the corpus callosum on cortical after-discharge patterns in the cat, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1967, CXXV, pp. 128-132.
Swank, R. L., Synchronization of spontaneous electrical activity of cerebrum by barbiturate narcosis, in ‟Journal of neurophysiology", 1949, XII, pp. 159-172.
Teitelbaum, H., Lateralization of olfactory memory in the split-brain rat, in ‟Journal of comparative and physiological psychology", 1971, LXXV, pp. 51-56.
Teitelbaum, H., Sharpless, S K., Byck, R., Role of somatosensory cortex in interhemispheric transfer of tactile habits, in ‟Journal of comparative and physiological psychology", 1968, LXVI, pp. 623-632.
Toglia, J. U., The corpus callosum, in ‟Diseases of the nervous system", 1961, XXII, pp. 1-6.
Tomasch, J., Size, distribution and number of fibres in the human corpus callosum, in ‟Anatomical record", 1954, CXIX, pp. 119-135.
Tomasch, J., A quantitative analysis of the human anterior commissure, in ‟Acta anatomica", 1957, XXX, pp. 902-906.
Towe, A. L., Nyquist, J. K, Tyner, C. F., Composition of the corticofugal reflex, in ‟Brain research", 1969, XVI, pp. 530-534.
Toyama, K., Matsunami, K., Ohno, T., Antidromic identification of association, commissural and corticofugal efferent cells in cat visual cortex, in ‟Brain research", 1969, XIV, pp. 513-517.
Toyama, K., Tokashiki, S., Matsunami, K., Synaptic action of commissural impulses upon association efferent cells in cat visual cortex, in ‟Brain research", 1969, XIV, pp. 518-520.
Trevarthen, C. B., Double visual learning in split-brain monkeys, in ‟Science", 1962, CXXXVI, pp. 258-259.
Trevarthen, C. B., Functional interactions between the cerebral hemispheres of the split-brain monkey, in Functions of the corpus callosum (a cura di E. G. Ettlinger), London 1965, pp. 24-41.
Trevarthen, C. B., Two mechanisms of vision in primates, in ‟Psychologische Forschung", 1968, XXXI, pp. 299-337.
Trevarthen, C. B., Experimental evidence for a brain-stem contribution to visual perception in man, in ‟Brain, behavior and evolution", 1970, III, pp. 338-352.
Tyner, C. F., Towe, A. L., Interhemispheric influences on sensorimotor neurons, in ‟Experimental neurology", 1970, XXVIII, pp. 88-105.
Unterhardnscheidt, F., Jachnik, D., Gött, H., Der Balkenmangel, Berlin 1968.
Van Hof, M. W., Interocular transfer in the rabbit, in ‟Experimental neurology", 1970, XXVI, pp. 103-108.
Van Valkenburg, C. T., Experimental and pathologico-anatomical researches on the corpus callosum, in ‟Brain", 1913, XXXVI, pp. 119-165.
Van Wagenen, W. P., Herren, R. Y., Surgical division of commissureal pathways in the corpus callosum, in ‟Archives of neurology and psychiatry", 1940, XLIV, pp. 740-759.
Vesbayeva, C., Whitteridge, D., Wilson, M. E., Callosal connexions of the cortex representing the area centralis, in ‟Journal of physiology", 1967, CXCI, pp. 79P-80P.
Villaverde, J. M. de, Contribution à la connaissance du système commissural de l'aire motrice du lapin, in ‟Travaux des recherches biologiques de l'Université de Madrid", 1924, XXII, pp. 1-21.
Villaverde, J. M. de, Les connexions commissurales des régions postérieures du cerveau du lapin, in ‟Travaux des recherches biologiques de l'Université de Madrid", 1924, XXII, pp. 99-141.
Villaverde, J. M. de, Sur la terminaison de fibres calleuses dans l'écorce cérébrale, in ‟Travaux des recherches biologiques de l'Université de Madrid", 1932, XXVII, pp. 275-297.
Villaverde, J. M. de, Quelques détails sur la manière dont les fibres calleuses se distribuent dans l'écorce cérébrale, in ‟Travaux des recherches biologiques de l'Université de Madrid", 1932, XXVII, pp. 345-375.
Voneida, T. J., Performance of a visual conditioned response in split-brain cats, in ‟Experimental neurology", 1963, VIII, pp. 473-504.
Voneida, T. J., Effect of brain bisection on capacity for cross comparison of patterned visual input, in ‟Experimental neurology", 1970, XXVI, pp. 60-71.
Webster, K. E., The cortico-striatal projection in the cat, in ‟Journal of anatomy", 1965, IC, pp. 329-337.
Wegener, J. G., A note on auditory discrimination behaviour and the corpus callosum, in Functions of the corpus callosum (a cura di E. G. Ettlinger), London 1965, pp. 69-72.
Westheimer, G., Mitchell, D. E., The sensory stimulus for disjunctive eye movements, in ‟Vision research", 1969, IX, pp. 749-755.
Whitteridge, D., Area 18 and the vertical meridian of the visual field, in Functions of corpus callosum (a cura di E. G. Ettlinger), London 1965, pp. 115-120.
Wilson, M. E., Cortico-cortical connexions of the cat visual areas, in ‟Journal of anatomy", 1968, CII, pp. 375-386.
Zeki, S. M., Visual deficits related to size of lesion in prestriate cortex of optic chiams sectioned monkeys, in ‟Life science", 1967, VI, pp. 1627-1638.
Zeki, S. M., Interhemispheric connections of prestriate cortex in monkey, in ‟Brain research", 1970, XIX, pp. 63-75.