
Sinapsi
Sinapsi
Fisiologia della sinapsi centrale, di John C. Eccles
Fisiologia della sinapsi periferica, di John C. Eccles
Farmacologia della sinapsi neuromuscolare, di William D. M. Paton
Fisiologia della sinapsi centrale
SOMMARIO: 1. Introduzione storica. □ 2. Struttura della sinapsi. □ 3 Azione sinaptica eccitatoria: a) registrazione elettrica intracellulare; b) potenziali postsinaptici eccitatori (PPSE); c) effetti della variazione del potenziale di membrana sui PPSE. □ 4. Generazione dell'impulso per azione sinaptica: a) potenziali d'azione dei motoneuroni; b) motoneuroni tonici e motoneuroni fasici. □ 5. Azione sinaptica inibitoria: a) potenziali postsinaptici inibitori (PPSI); b) meccanismi ionici dell'azione sinaptica inibitoria. □ 6. Vie eccitatorie e inibitorie nel sistema nervoso centrale: a) introduzione; b) il sistema delle cellule di Renshaw; c) l'inibizione mediata dalle cellule a canestro dell'ippocampo; d) circuiti neuronali nel cervelletto; e) caratteristiche operative delle vie inibitorie; f) inibizione per mezzo di sinapsi reciproche. □ 7. Inibizione presinaptica. □ 8. Sostanze trasmettitrici sinaptiche: a) trasmettitore eccitatorio; b) trasmettitori inibitori. □ 9. Attivazione sinaptica ripetitiva: a) potenziamento di frequenza delle sinapsi; b) potenziamento posttetanico delle sinapsi. □ Bibliografia.
1. Introduzione storica.
La struttura funzionale della sinapsi periferica (v. l'articolo seguente, SINAPSI: Fisiologia della sinapsi periferica) risolve il problema fisiologico essenziale della trasmissione di impulsi attraverso lo spazio sinaptico con il minimo di ritardo e di distorsione, e anche con un fattore di sicurezza abbastanza alto da assicurare regolarità di funzionamento. Queste sinapsi a trasmissione chimica sono modellate in modo tale da compensare l'asimmetria elettrica fra le componenti pre- e postsinaptica della sinapsi, come per esempio la piccolissima terminazione nervosa e la vasta superticie della membrana della fibra muscolare con la sua alta capacità (v. fig. 1A dell'articolo seguente). Sebbene queste stesse funzioni siano espletate dalle sinapsi del sistema nervoso centrale, la loro struttura funzionale deve essere del tutto diversa a causa del problema generale delle interrelazioni tra le diverse parti del sistema nervoso. Le differenze principali sono infatti dovute alla funzione integrativa di tale sistema.
All'inizio di questo secolo Ch.S. Sherrington (v., 1906) ha fatto dell'integrazione il tema fondamentale del suo classico libro The integrative action of the nervous system: egli ha sviluppato il concetto di integrazione centrale come risultato di uno studio sui riflessi selezionati in modo da comprendere questo o quell'aspetto dell'integrazione. Venivano usati stimoli sensitivi specifici e venivano condotte osservazioni sulle conseguenti contrazioni muscolari. Era già stato provato che il muscolo è un esecutore fedele del sistema nervoso: infatti le contrazioni che si osservano riproducono fedelmente i segnali in uscita dal sistema nervoso centrale, che sono ovviamente le scariche di impulsi dei motoneuroni. Così Sherrington ha potuto definire il modo di operare di semplici sistemi di neuroni - le cosiddette vie riflesse - mettendolo in relazione specificamente con le proprietà delle sinapsi fra i neuroni. Alcuni anni prima egli aveva sviluppato il concetto funzionale di sinapsi da quello anatomico del neurone e delle minute regioni di contatto che esistono fra i neuroni. È stata una magnifica acquisizione di S. Ramòn y Cajal (v., 1909 e 1911) l'aver mostrato che il sistema nervoso è composto di unità anatomicamente separate, i neuroni, in relazione fra loro per mezzo di stretti contatti - e perciò per contiguità, e non per continuità come proposto dalla teoria reticolare. Dal momento che queste aree sinaptiche erano molto piccole, Sherrington suppose che per far scaricare un neurone fosse necessario attivarne molte contemporaneamente: ne derivarono pertanto i concetti di convergenza e di sommazione. Una proprietà complementare deriva dalla ricca diramazione delle fibre nervose, cui consegue infatti una notevole diffusione spaziale dell'attività che entra nel sistema nervoso attraverso una qualunque via afferente. Il principio funzionale di divergenza, come è stato chiamato, assicura che ogni particolare afferenza abbia almeno la possibilità di determinare una vasta influenza sulle scariche riflesse. Il concetto più rivoluzionario introdotto da Sherrington fu che alcune delle sinapsi su una membrana avessero un'azione inibitoria, e pertanto impedissero la scarica che altrimenti sarebbe stata evocata dall'azione sinaptica eccitatoria. È sorprendente il fatto che fino a 20-30 anni fa si riteneva che l'azione inibitoria fosse esercitata solo sui motoneuroni!
Una pubblicazione successiva, Reflex activity in the spinal cord di R. S. Creed e altri (v., 1932), fornisce un resoconto dello studio analitico compiuto più tardi sui riflessi dalla scuola neurofisiologica di Sherrington a Oxford. Il concetto di unità motrice era già stato sviluppato e studiato quantitativamente (v. fig. 1 dell'articolo seguente). Si definisce come unità motrice l'insieme dell'assone di un motoneurone e delle fibre muscolari da questo innervate (v. Sherrington, 1929). A causa della notevole ramificazione dell'assone, generalmente il numero di fibre muscolari che esso innerva è superiore a 100 e ciascuna fibra è innervata da un solo motoneurone. Il lavoro di Creed è stato essenzialmente un tentativo di descrivere diversi tipi di riflessi del midollo spinale in termini di motoneuroni che ne sono alla base. Le contrazioni riflesse venivano spiegate quantitativamente mediante il numero di unità motrici individuali implicate e la sequenza temporale della loro attivazione: in realtà questa asserzione può essere estesa a tutte le reazioni comportamentali che comportano la contrazione di un muscolo striato.
Nell'articolo seguente sarà descritta la trasmissione attraverso la sinapsi neuromuscolare, per cui un impulso nervoso genera un impulso muscolare e pertanto una contrazione del muscolo. In questo articolo è più opportuno cominciare con alcuni semplici concetti sulle modalità di azione del neurone che rende operante l'unità motrice, cioè il motoneurone, poiché questo studio si collega con il classico lavoro sui riflessi (v. Creed e altri, 1932) e anche perché il motoneurone è stato estensivamente studiato con le moderne tecniche analitiche.
La fig. 1A mostra le vie più semplici di eccitazione e inibizione attraverso il sistema nervoso centrale da muscoli che agiscono in maniera antagonista in corrispondenza dell'articolazione del ginocchio. I recettori anulospirali da stiramento (AS) del muscolo estensore del ginocchio (E) sono connessi al midollo spinale da fibre a grande diametro (gruppo Ia) che esercitano una potente azione sinaptica eccitatoria direttamente sui motoneuroni di quel muscolo (E) e dei muscoli collegati. Se il muscolo viene stirato in maniera abbastanza forte, le terminazioni anulospirali, mediante le scariche lungo le loro fibre, indurranno nei motoneuroni un'eccitazione abbastanza potente da causare la scarica di impulsi lungo gli assoni, e pertanto il muscolo si contrae. Questo è il noto riflesso tendineo o riflesso da stiramento della neurofisiologia ed è l'esempio classico dei riflessi monosinaptici. Il breve stiramento prodotto da un leggero colpo sul tendine provoca un riflesso tendineo come il riflesso patellare o il riflesso tendineo della caviglia. Uno stiramento costante e prolungato provoca una contrazione prolungata del riflesso da stiramento (v. Creed e altri, 1932). La fig. 1C mostra un esempio di facilitazione temporale in questa via monosinaptica. La doppia stimolazione delle fibre la mediante l'elettrodrodo SE evoca la scarica di un impulso da molti motoneuroni (la risposta di potenziale), mentre ciascuno stimolo isolato è inefficace.
Nella fig. 1A vi è un'altra via verso il motoneurone E, quella che proviene dalle terminazioni anulospirali (AS) del muscolo flessore antagonista del ginocchio (F). Ciascuna di queste fibre afferenti emette collaterali nel midollo spinale, le quali formano poi sinapsi eccitatorie su piccole cellule nervose (interneuroni, IN); questi a loro volta formano sinapsi inibitorie sui motoneuroni estensori, che impediscono o riducono la probabilità che si produca una loro scarica riflessa. Questa riduzione appare come una diminuzione nella risposta riflessa monosinaptica (v. fig. 1D), registrata come nella fig. 1B sotto forma di ‛potenziale di popolazione' dall'insieme degli assoni motori che innervano il muscolo. Il potenziale in risposta alla stimolazione SF (v. fig. 1A) è ridotto a meno della metà da uno stimolo SE applicato precedentemente. La fig. 1A schematizza la via più semplice per un'azione inibitoria dalla periferia, e inoltre mostra la relazione reciproca delle vie che provengono da muscoli antagonisti, gli estensori e i flessori dell'articolazione del ginocchio. Tutte le sinapsi delle fibre afferenti dalle terminazioni anulospirali, sia sui motoneuroni sia sugli altri neuroni, sono eccitatorie. Per avere un'azione inibitoria si trova inserita una speciale classe di interneuroni (IN) le cui sinapsi sono esclusivamente inibitorie. È stato osservato per la prima volta da Lloyd nel 1946 che le fibre afferenti da un muscolo hanno un'azione inibitoria sui motoneuroni dei muscoli antagonisti, come indicato dalla riduzione del potenziale di popolazione registrato dagli assoni motori nella radice ventrale (v. fig. 1D). Solo molto più tardi fu stabilito che vi è interpolazione di un neurone inibitore, come schematizzato nella fig. 1A (v. Eccles, 1957 e 1969).
È chiaro che il diagramma della fig. lA è notevolmente semplificato, sia nella via monosinaptica eccitatoria per il singolo motoneurone sia nella via inibitoria attraverso un singolo interneurone. Vi sono più di 100 terminazioni anulospirali in un grande muscolo, il quale naturalmente è l'insieme di centinaia di unità motrici (v. Creed e altri, 1932). Ciascuna fibra afferente da una terminazione anulospirale si dirama estesamente nel midollo spinale (il ‛principio di divergenza' ricordato prima), cosicché essa innerva un grande numero di motoneuroni con le rispettive unità motrici. In maniera reciproca, su ciascun motoneurone vi è una larga convergenza di fibre eccitatorie, in genere da 10 a 50 (v. Kuno, 1964), su ciascun motoneurone delle centinaia che innervano il muscolo. La divergenza e la convergenza sarebbero presenti in ciascuno stadio delle vie inibitorie della fig. 1A, le quali si devono immaginare come tante linee in parallelo. Tuttavia, la fig. lA indica correttamente che, nonostante questa massiva disposizione in parallelo, vi è soltanto un interneurone (IN) in serie nella via inibitoria: pertanto tale via è disinaptica.
2. Struttura della sinapsi.
Dopo questa introduzione generale sulle vie più semplici, è opportuno descrivere le caratteristiche strutturali essenziali dei neuroni del sistema nervoso centrale: quelli le cui connessioni sono implicate nelle semplici azioni riflesse, e quelli con collegamenti molto più complessi ai più alti livelli del sistema nervoso. Esamineremo inoltre la morfologia dettagliata di tipiche sinapsi eccitatorie e inibitorie, quale risulta dall'osservazione al microscopio elettronico.
Come illustrato nella fig. 2A, su un motoneurone converge un gran numero di fibre, ciascuna delle quali forma uno o alcuni contatti sinaptici piccoli - in genere di non più di 1 μm di diametro. Questi si vedono sparsi sulla superficie del soma e delle diramazioni dendritiche che da esso originano, ma le sinapsi sono presenti anche sulle sottili diramazioni dendritiche terminali. Su un grande motoneurone vi sarebbero decine di migliaia di sinapsi, ma solo poche di queste appartengono alle vie riflesse schematizzate nella fig. 1A. Tecniche sperimentali raffinate hanno dimostrato che le sinapsi eccitatorie monosinaptiche si trovano per la maggior parte sui dendriti, in genere nella regione intermedia, sebbene alcune siano vicine al soma (v. Burke, 1967). Tuttavia vi è un enorme numero di altre vie in rapporto con i motoneuroni. La fig. 2B illustra una cellula piramidale dell'ippocampo con alcuni contatti sinaptici disegnati sul soma, sui grandi dendriti e perfino sui piccoli dendriti terminali. Le strutture sinaptiche sono mostrate in dettaglio negli ingrandimenti a destra.
Le fotografie prese al microscopio elettronico mostrano che le sinapsi centrali assomigliano alle sinapsi neuromuscolari (v. articolo seguente, fig. 2) nelle loro caratteristiche fondamentali. La fibra nervosa presinaptica si espande in un bulbo terminale, oppure può attraversare diverse zone sinaptiche espandendosi in ciascuna di queste per formare un bouton de passage. Nella fibra presinaptica vi sono vescicole sinaptiche ammassate verso lo spazio sinaptico, spesso in gruppi discreti. I numerosi mitocondri si trovano più lontano. La larghezza dello spazio sinaptico, notevolmente uniforme, è di circa 200 Å. Le membrane pre- e postsinaptica che delimitano la fessura tendono a colorarsi intensamente, e frequentemente appaiono spesse e dense dalla parte presinaptica. Sui dendriti di alcuni neuroni, per esempio delle cellule piramidali della corteccia cerebrale (v. fig. 2B), vi sono numerose spine, ciascuna delle quali costituisce l'elemento postsinaptico di una sinapsi. Queste sinapsi sulle spine possono essere molto numerose - fino a 20.000 su una grande cellula piramidale della corteccia cerebrale, e fino a 200.000 su una cellula di Purkinje del cervelletto umano.
Nelle fotografie prese al microscopio elettronico è ora generalmente possibile distinguere le sinapsi eccitatorie da quelle inibitorie. La fig. 3A illustra un valido criterio scoperto da Uchizono (v., 1967). Quando sono fissate in glutaraldeide, le vescicole di alcune sinapsi (come la sinapsi sulla spina nella fig. 3A) hanno una forma sferica abbastanza uniforme, come nella sinapsi neuromuscolare (v. articolo seguente, fig. 2D). In altre sinapsi le vescicole sono più piccole e tendono a essere ellissoidali, come si può vedere nell'altra sinapsi della fig. 3A. Uchizono ha dimostrato che le vescicole di tipo ellissoidale si trovano nelle sinapsi che con criteri fisiologici sono state identificate come inibitorie, mentre le vescicole sferiche sono caratteristiche delle sinapsi eccitatorie. Questo criterio di differenziazione sinaptica, basato sulla forma delle vescicole, è stato oggetto di approfonditi esami sperimentali ed è ora generalmente accettato. Per es., la fig. 3B mostra due sinapsi adiacenti su di un motoneurone, con vescicole sferiche l'una, ellissoidali l'altra. Prima che questo criterio fosse scoperto, vi erano indicazioni a favore del fatto che i due tipi di sinapsi nervose descritte da Gray (v. Whittaker e Gray, 1962) corrispondessero alle due funzioni sinaptiche, e che i cosiddetti tipi 1 e 2 fossero rispettivamente eccitatori e inibitori. Si può ora ritenere che questa identificazione sia in genere corretta, come provato in base alla forma delle vescicole, ma vi sono eccezioni.
Il diagramma della fig. 4A illustra questi due tipi di sinapsi: la forma delle vescicole indica che il tipo 1 è eccitatorio e il tipo 2 inibitorio. Le sinapsi sulle spine, come quelle marcate da a a d nella fig. 2B, sono quasi costantemente del tipo 1, come mostrato nella fig. 4B, e hanno vescicole sferiche; pertanto si pensa che siano sinapsi eccitatorie. Le sinapsi e ed f della fig. 2B sono del tipo 2. Le vescicole nella fig. 2B sono tutte sferiche, e ciò perché non è stata usata la fissazione con glutaraldeide. È ora riconosciuto che la sinapsi indicata con e è formata dalle cellule a canestro ed è inibitoria (v. fig. 20), con vescicole ellissoidali. Anche la sinapsi f può essere inibitoria, ma g è eccitatoria e prende origine dalle collaterali di Schaffer (v. fig. 11A). La fig. 4C mostra schematicamente che le vescicole sinaptiche sono disposte in maniera regolare in un mezzo strutturato che si estende a guisa di dense proiezioni dalla membrana presinaptica. Le vescicole sinaptiche vicine alla membrana sono pertanto tenute in una specie di griglia vescicolare che può avere a che fare con il metabolismo delle vescicole o può servire a guidarle verso la membrana presinaptica pronte per l'emissione (v. Gray, 1966 e 1969; v. Akert e Sandri, 1969; v. Akert e altri, 1969).
Una nuova eccellente tecnica per mettere in risalto l'ultrastruttura delle sinapsi è illustrata nella fig. 4D (v. Akert e altri, 1969). Le vescicole sinaptiche (vs) e un mitocondrio (m) si trovano nella superficie di taglio del bottone sinaptico, del quale si vedono anche la superficie esterna (se) e l'assone (ass.). Più in basso si vede la stretta fessura sinaptica (fs), che separa il bottone dall'elemento postsinaptico (po).
3. Azione sinaptica eccitatoria.
a) Registrazione elettrica intracellulare.
Le ricerche sperimentali sulla trasmissione sinaptica nella via più semplice del midollo spinale (v. fig. 1) furono rivoluzionate nel 1951 dall'applicazione della tecnica di registrazione elettrica dall'interno delle cellule nervose, mediante l'uso di microelettrodi intracellulari di vetro e di un inseguitore catodico; tale metodo era stato usato circa un anno prima da Fatt e Katz nelle ricerche sulla trasmissione neuromuscolare (v. Brock e altri, 1952). Per il motoneurone vi era una difficoltà in più: infatti il microelettrodo doveva essere collocato all'interno della cellula senza l'aiuto del controllo visivo e ancor più difficile era mantenere rigidamente la sua posizione; ciò nonostante, già i primi esperimenti eseguiti con una tecnica ancora primitiva ebbero sorprendentemente successo, e presto entrammo in possesso di conoscenze assolutamente nuove sulla natura dell'azione sinaptica eccitatoria e inibitoria sul motoneurone (v. Eccles, 1957). In seguito risultò chiaro che questa scelta iniziale di sperimentazione intracellulare era stata particolarmente fortunata; infatti di tutti i neuroni del sistema nervoso centrale i motoneuroni sono quelli che tollerano meglio la penetrazione di un microelettrodo, e dopo alcuni minuti molti di essi si stabilizzano e danno per ore risposte che non sembrano essere alterate dalla penetrazione. Gli elettrodi che danno migliori risultati sono quelli che si assottigliano molto gradualmente e hanno la punta di circa 0,5 μm di diametro, il che conferisce loro una resistenza di 10-15 MΩ quando sono riempiti con una soluzione di KCl 3M.
b) Potenziali postsinaptici eccitatori (PPSE).
La fig. 5A mostra schematicamente l'elettrodo stimolante appoggiato sulle numerose e grandi fibre nervose che prendono origine dalle terminazioni anulospirali del muscolo (v. fig. 1A), le quali convergono e formano sinapsi eccitatorie sul motoneurone di quel muscolo. Nella fig. 5, B-J, viene applicata una stimolazione progressivamente più forte attraverso l'elettrodo stimolante; il numero di fibre eccitate in tal modo è stato determinato mediante l'elettrodo registrante posto in maniera tale da registrare gli impulsi nel momento in cui entrano nel midollo spinale (tracciati superiori). Nei tracciati inferiori vi sono i potenziali registrati intracellularmente, prodotti dall'azione sinaptica eccitatoria. Essi assomigliano ai potenziali di placca (PP) descritti nell'articolo seguente, in quanto sono in direzione depolarizzante; infatti vi è una temporanea diminuzione del potenziale di riposo di membrana, che è di circa -70 mV. Aumentando il numero delle fibre stimolate, si ha un cambiamento nell'ampiezza, ma non nel decorso temporale del potenziale intracellulare, come indicato dai picchi di potenziale delle tracce superiori. Questo illustra bene il modo in cui la convergenza di fibre afferenti su un neurone produce sommazione dei potenziali sinaptici eccitatori che, dato che si registrano attraverso la membrana postsinaptica, prendono il nome di potenziali postsinaptici eccitatori o PPSE. Dopo G un ulteriore aumento dello stimolo non dà luogo ad aumento del PPSE. In G c'è stata la stimolazione di tutte le fibre di gruppo Ia; l'aumento che si osserva nel potenziale afferente (v. fig. 5, H-J) è dovuto alla stimolazione delle fibre di gruppo Ib che non hanno azione eccitatoria monosinaptica (v. Eccles e altri, 1957). In genere esiste una relazione lineare fra la quantità dell'eccitazione monosinaptica e l'ampiezza del PPSE: ciò è illustrato nella fig. 5, K-N, dove i PPSE sono stati evocati in un motoneurone dalla convergenza di fibre afferenti Ia che si trovano in nervi di tre diversi muscoli (v. fig. 5, K, L e M), e quando la loro scarica era sincrona (N) il PPSE era esattamente la somma dei tre singoli PPSE. Secondo lo schema elettrico della fig. 8B, ci si deve aspettare una deviazione apprezzabile dalla linearità solo quando i PPSE sono molto grandi. Pertanto, si può presumere che i PPSE osservati prendano origine dalla semplice sommazione dei PPSE prodotti da ciascuna sinapsi.
Si può ritenere certo che, come il PP, il PPSE sia generato da un trasmettitore chimico, la cui identificazione non è ancora sicura. Vi sono, per esempio, caratteri peculiari, come il ritardo sinaptico visto nella fig. 5 fra l'impulso presinaptico e il PPSE, i PPSE in miniatura con proprietà quantiche illustrati nella fig. 6H e gli effetti che il variare del potenziale di membrana determina sull'ampiezza e la direzione del PPSE (v. fig. 8, A e C). Come nel caso del PP (v. articolo seguente, fig. 4B), esperimenti compiuti usando la tecnica del blocco di voltaggio mostrano che il PPSE è prodotto da correnti depolarizzanti relativamente brevi (la corrente postsinaptica eccitatoria, CPSE), che depolarizzano la membrana postsinaptica, come mostrato nell'ingrandimento della fig. 7A. La lenta fase esponenziale di recupero è dovuta al ristabilirsi del potenziale di membrana mediante la messa in funzione di una batteria di membrana (EM) (v. fig. 8B) in serie con la resistenza della membrana (RM) e in parallelo con la capacità della membrana (CM). La costante di tempo della membrana è stata misurata direttamente nel motoneurone, e nella fig. 7A è stata usata per calcolare la CPSE (linea tratteggiata) che dà il PPSE della fig. 7A. Dopo 2 o 3 ms vi è al massimo soltanto un piccolo residuo dell'azione del trasmettitore.
I PPSE evocati in un motoneurone per via monosinaptica da una scarica afferente di gruppo la (v. fig. 5, B-N) rappresentano la somma di diversi PPSE unitari, ciascuno dei quali è prodotto dai bottoni sinaptici o bottoni che prendono origine da una singola fibra Ia (v. fig. 5A). Si può riconoscere facilmente il PPSE unitario prodotto da singoli impulsi presinaptici quando vi è un bombardamento casuale del motoneurone che dà luogo a quello che si può definire ‛rumore sinaptico' (v. Brock e altri, 1952). Nella fig. 6A, per esempio, si possono distinguere due diversi tipi di PPSE unitari, ciascuno in successione ritmica indipendente, uno molto breve, l'altro prolungato. Ingrandimenti di questi due tipi di PPSE, I e II, sono mostrati in B e C, rispettivamente, sulla stessa scala di tempo del PPSE prodotto dalla scarica la in D. Il PPSE unitario in B (I) è eccezionalmente breve e sarebbe prodotto da una modesta eccitazione sinaptica vicino al soma del motoneurone, che è presumibilmente il luogo di registrazione, mentre il PPSE unitario in C (II) mostra il rallentamento del decorso temporale conseguente alla trasmissione elettrotonica da una zona sinaptica piuttosto remota su un dendrite. Il PPSE composto di D è 10 volte più grande del PPSE unitario di tipo II (9 mV anziché 0,9 mV) e ha un decorso temporale un po' più rapido. Evidentemente esso è formato dalla sommazione di molti PPSE unitari che sono per la maggior parte generati in posizioni remote sui dendriti. È stato individuato un intero spettro di PPSE unitari (v. Burke, 1967), e in effetti i tipi I e II mostrati nella fig. 6, A-C, sono esempi estremi.
La fig. 6E rappresenta una serie più complessa di PPSE unitari spontanei registrati da un motoneurone dopo sezione di tutte le radici dorsali; questi PPSE sono pertanto prodotti mediante bombardamento sinaptico degli interneuroni. Si nota un assortimento di PPSE unitari di diverso tipo: di particolare interesse sono le due brevi interpolarizzazioni marcate da punti, identificabili come potenziali unitari inibitori postsinaptici.
A rigor di termini, in analogia con i potenziali di placca, la locuzione PPSE in miniatura dovrebbe essere applicata non ai PPSE unitari della fig. 6, A, B, C, E, ma ai PPSE prodotti dall'emissione di quanti di trasmettitore da singole vescicole sinaptiche.
Tali PPSE in miniatura sono stati osservati dopo l'eliminazione di ogni trasmissione di impulsi, ottenuta immergendo il midollo spinale isolato di rana in una soluzione ad alta concentrazione di potassio (v. Katz e Miledi, 1963). Per esempio nella fig. 6F vi sono i PPSE unitari spontanei come nella fig. 6A, e in G si vedono i PPSE unitari evocati da una scarica molto piccola di impulsi presinaptici, probabilmente di un solo impulso, come nella fig. 6I; in diversi casi lo stimolo non evoca nulla. Il midollo spinale è stato poi immerso in una soluzione di Ringer con un'alta concentrazione di potassio (60 mM); la fig. 6H mostra dei PPSE unitari altrettanto grandi di quelli di E ed F che devono essere veri PPSE in miniatura. Più recentemente (v. Blankenship e Kuno, 1968) nei motoneuroni di Mammifero sono stati riconosciuti veri PPSE in miniatura, dopo che con la tetrodotossina era stata eliminata ogni trasmissione di impulsi. La fig. 6I è simile alla fig. 6G, ma si riferisce a un motoneurone di gatto normale che negli 8 tracciati superiori è stato eccitato per via monosinaptica da un singolo impulso di gruppo Ia: pertanto si tratta di veri PPSE unitari (v. Kuno, 1964). La mancanza di risposta in tre tracce e le variabili latenze dei PPSE unitari nelle altre cinque tracce ricordano la serie di PP in miniatura della fig. 6C dell'articolo seguente. Dopo aver analizzato estese serie di PPSE di tipo unitario, con la loro frequente assenza dopo lo stimolo, Kuno (v., 1964) ha concluso che la trasmissione monosinaptica sui motoneuroni avviene in stadi quantici, e che un impulso in una singola fibra afferente libera in media solo un quanto di trasmettitore. Ciò contrasta con i 100 o 200 quanti che si liberano alla giunzione neuromuscolare normale, ma non è un dato inaspettato in considerazione del fatto che i bottoni sinaptici sono di dimensioni piccole, in confronto alla terminazione nervosa motoria sulla fibra muscolare. Evidentemente, pertanto, i PPSE unitari della fig. 6, A-G e I, devono spesso essere PPSE in miniatura.
In un'eccellente ricerca sui PPSE unitari, Mendell e Henneman (v., 1971) hanno registrato con la tecnica delle medie le risposte di 6 motoneuroni a impulsi singoli nella stessa fibra afferente la (v. fig. 6J). La grande varietà dell'ampiezza e del decorso temporale di questi sei PPSE è attribuibile alle localizzazioni e alle dimensioni delle sinapsi formate da questa fibra Ia. La collocazione relativa ai tre tracciati superiori sarebbe vicina alla zona di registrazione nel soma; per i due più in basso, in particolare, le sinapsi avrebbero una localizzazione remota nei dendriti.
Nella fig. 7 è mostrato un insieme di PPSE prodotti monosinapticamente in un'ampia varietà di neuroni del sistema nervoso centrale. Vi è sempre il caratteristico breve ritardo sinaptico, quindi la graduale depolarizzazione con una fase di ascesa rapida e un più lento declino che spesso ha le caratteristiche temporali dovute alla membrana neuronale (v. fig. 7, A e B). Tuttavia in alcuni neuroni l'azione del trasmettitore dura più a lungo e si ha pertanto un declino molto prolungato, come nella fig. 7C. In questo caso si sa che il trasmettitore è acetilcolina, che pare venga inattivata solo molto lentamente dall'acetilcolinesterasi. Nella fig. 7, B e D, sono mostrati i PPSE delle cellule di origine dei tratti spinocerebellari dorsale e ventrale.
Questi PPSE, come quello delle grandi cellule piramidali della corteccia motoria (v. fig. 7E), hanno una caratteristica brusca fase di salita e un declino più lento, cosicché in 10 ms vi è poco o nessun residuo di depolarizzazione. La fig. 7F mostra che anche per i grandi potenziali unitari prodotti in una cellula cerebellare di Purkinje (v. anche fig. 21) da un singolo impulso di una fibra rampicante, il decorso temporale ha le stesse caratteristiche di brusca ascesa e di declino lento, approssimativamente esponenziale. La freccia indica i PPSE prodotti da un treno più tardivo di 1 o 2 o 3 impulsi nella stessa fibra rampicante.
La fig. 8C ci fornisce un esempio di una disposizione sinaptica molto interessante sui neuroni del nucleo rosso. Si tratta di due localizzazioni ben distinte di sinapsi eccitatorie, quelle sul soma, che provengono dai neuroni del nucleo interposito (IP), e quelle sui dendriti piuttosto lontani, che prendono origine dalle cellule piramidali della corteccia (IC). L'esistenza di questa duplice localizzazione è messa in evidenza dalle seguenti caratteristiche: il primo gruppo di sinapsi mostra PPSE con insorgenza rapida e lento declino, fortemente aumentati dall'iperpolarizzazione del neurone; il secondo mostra PPSE con una fase di ascesa molto lenta attribuibile a distorsione elettrotonica, non influenzati dall'iperpolarizzazione del neurone. I PPSE compositi della fig. 8C, pertanto, assomigliano ai PPSE unitari della fig. 6, A e B, mentre le sinapsi IP e IC corrispondono rispettivamente ai tipi I e II (v. Tsukahara e Kosaka, 1968).
c) Effetti della variazione del potenziale di membrana sui PPSE.
Nelle ricerche dirette a studiare gli effetti della variazione del potenziale di membrana sulla corrente di placca (CP) è stato mostrato (v. articolo seguente, figg. 12 e 13) che esso si invertiva a un potenziale di membrana di circa −15 mV; questo potenziale di equilibrio è stato spiegato come un compromesso fra i potenziali di equilibrio degli ioni sodio e potassio, le cui conduttanze erano notevolmente aumentate dall'azione del trasmettitore sinaptico sulle zone recettrici della membrana postsinaptica. In condizioni favorevoli è possibile provocare un cambiamento del potenziale di membrana di un motoneurone sufficiente a invertire il PPSE (v. fig. 8A). Il potenziale di equilibrio si trova a un potenziale di membrana di circa 0 mV, il che fa pensare che il rapporto tra la conduttanza del sodio e quella del potassio sia qui maggiore che non nella placca motrice, anche se la conduttanza agli ioni sodio è meno predominante di quanto è stato osservato nella sinapsi gigante del calamaro (v. articolo seguente, fig. 16C). Quando le sinapsi eccitatorie sono molto lontane sui dendriti, la variazione del potenziale di membrana nel soma causata dall'applicazione di corrente ha un effetto molto piccolo o addirittura nullo sul PPSE (v. fig. 8C, IC), che mostra il caratteristico decorso temporale lento, come nella fig. 6C. Ciò pertanto contrasta con le sinapsi IP che sono situate vicino al soma.
Si possono produrre notevoli cambiamenti nella composizione ionica dei motoneuroni, mediante l'iniezione elettroforetica attraverso un elettrodo intracellulare. Tali iniezioni di anioni o cationi possono determinare notevoli diminuzioni del potenziale di membrana; il PPSE, che peraltro subisce un corrispondente abbassamento, non ne è influenzato in altro modo (v. Eccles, The physiology..., 1964). Poiché recentemente è stato dimostrato che gli anioni non sono implicati in maniera apprezzabile nel generare i potenziali di placca (PP), è auspicabile che si facciano ulteriori esperimenti, particolarmente su preparati isolati, nei quali sia possibile cambiare gli ioni extracellulari nello stesso modo in cui è stato fatto dai Takeuchi sul PP (v. articolo seguente, figg. 12 e 13).
La fig. 8B ci fornisce lo schema elettrico di come la sinapsi eccitatoria agisce sul motoneurone di Mammifero (gatto). Tutti i valori, eccetto il valore minimo dato a RE, rappresentano la media di esperimenti condotti su molte cellule. Dal momento che il potenziale di equilibrio per le sinapsi eccitatorie è di circa 0 mV, il canale di conduttanza potrebbe essere mostrato come un semplice cortocircuito con resistenza variabile sotto il controllo del trasmittitore sinaptico. Questo schema dimostra che se si avesse una serie di piccole conduttanze cortocircuitanti in parallelo, rappresentanti azioni sinaptiche individuali, si avrebbe approssimativamente una sommazione di queste conduttanze fintantoché il cambiamento di conduttanza totale non fosse grande (20% o più della conduttanza totale attraverso la membrana). Si spiegano così le osservazioni della fig. 5, L-M.
4. Generazione dell'impulso per azione sinaptica.
a) Potenziali d'azione dei motoneuroni.
Come mostrato nella fig. 9, A-D, se il PPSE raggiunge un livello critico di soglia (B), il neurone scarica un impulso, e la latenza è tanto più breve quanto più grande è il PPSE (v. fig. 9, B-D). Nella fig. 9, B-D, ciò è stato ottenuto aumentando l'ampiezza della scarica presinaptica, cioe aumentando il numero di sinapsi attivate sui motoneuroni (v. fig. 5A). Come è prevedibile, si può fare in modo che anche un PPSE inferiore alla soglia generi un impulso mediante procedure che spostano il potenziale di membrana verso il livello critico di soglia di depolarizzazione. Per esempio nella flg. 9, F-H, lo stesso PPSE della fig. 9E è stato reso efficace dall'applicazione di una corrente di fondo depolarizzante iniziata 12 ms prima (l'intensità è riportata in ciascun tracciato in nA). Lo spostamento del potenziale di membrana è indicato a sinistra. L'impulso inizia in corrispondenza delle frecce quando il livello totale di depolarizzazione è di circa 18 mV, livello che è formato in proporzioni diverse da una depolarizzazione condizionante e dal PPSE sovrapposto. Il livello di soglia di depolarizzazione può essere ottenuto anche sovrapponendo il PPSE alla depolarizzazione prodotta da un precedente PPSE, fatto che offre una spiegazione sufficiente del fenomeno riflesso noto come facilitazione temporale: una singola scarica afferente di impulsi non riesce a evocare una scarica riflessa, ma due di tali scariche a breve intervallo sì (v. fig. 1C). Simili osservazioni sono state effettuate per impulsi generati da PPSE prodotti in un'ampia varietà di cellule nervose.
Nella fig. 9, I-L, è illustrato il livello critico di depolarizzazione utile a generare un impulso in una cellula piramidale della corteccia alla quale sono stati applicati impulsi depolarizzanti di ampiezza crescente, come mostrato nei tracciati superiori. In I la depolarizzazione non ha raggiunto la soglia. In J e K la scarica di impulsi è stata generata sempre più precocemente ma, come indicato dalle frecce, pressoché allo stesso livello di depolarizzazione di circa 13 mV.
È ormai generalmente accettato che l'azione sinaptica eccitatoria sia efficace nel generare la scarica di un impulso solo producendo un PPSE che raggiunga il livello soglia di depolarizzazione.
Quando i potenziali d'azione generati da un PPSE sono registrati a velocità abbastanza alta, si vede che presentano un'inflessione sulla fase ascendente paragonabile a quella che si ha nel potenziale d'azione generato nel motoneurone dall'invasione antidromica di un impulso dal suo assone motore. La duplice composizione del potenziale d'azione del motoneurone è particolarmente evidente quando si registra la sua derivata ottenuta elettricamente, come nei tracciati inferiori delle figg. 9, B-D ed F-H, e 10, A, B, D, E. La prima parte del potenziale d'azione, che inizia in corrispondenza della prima freccia a doppia testa nella fig. 10B, è chiamata potenziale SI, in quanto vi sono convincenti prove sperimentali che esso sia generato nel segmento iniziale del motoneurone (v. fig. 10C), che è formato dalla parte semilineare dell'assone motore e dall'eminenza assonica dalla quale prende origine l'assone (v. fig. 5A). La seconda parte del potenziale inizia quando, per le linee del flusso di corrente illustrate nella fig. 10F, il potenziale SI ha depolarizzato la membrana somatodendritica al suo livello soglia che è più alto di quello del segmento iniziale (la linea orizzontale SD nella flg. 10, A e B). Si chiama potenziale SD, perché è prodotto dal soma e dai dendriti. In altri tipi di cellule nervose i potenziali d'azione hanno spesso una duplice composizione paragonabile a quella dei potenziali SI-SD del motoneurone. Per esempio nella fig. 10, D ed E, si vedono potenziali d'azione delle cellule piramidali della corteccia che sono generati dall'invasione antidromica e dall'eccitazione sinaptica, e sono pertanto paragonabili a quelli della fig. 10, A e B. Nei tracciati inferiori le registrazioni della derivata ottenuta elettricamente mostrano la tipica configurazione SI-SD, sebbene in maniera meno evidente che per il motoneurone. La generazione di impulsi nel segmento iniziale delle cellule piramidali della corteccia è stata studiata mediante stimolazione con diversi tipi di correnti depolarizzanti - rampe, gradini, ecc. (v. Koike e altri, 1968).
La generazione di un impulso nel segmento iniziale avviene per il fatto che la soglia di depolarizzazione è molto più bassa che per la membrana somatodendritica (v. Eccles, Presynaptic inhibition..., 1964). Per esempio per i motoneuroni del gatto i valori medi sono rispettivamente di 10 e di 25 mV e per i motoneuroni della rana di 9 e di 26 mV. Si può presumere che questa bassa soglia del segmento iniziale abbia l'importante funzione di assicurare che la generazione dell'impulso avvenga per un effetto integrativo della combinazione fra le azioni sinaptiche eccitatorie e inibitorie sul neurone, poiché tutte queste azioni, per effetto della diffusione elettrotonica, convergeranno nella zona a bassa soglia di eccitazione che si trova nella regione d'origine dell'assone.
In diverse specie di neuroni non si trova traccia di composizione SI-SD del potenziale d'azione (v. Eccles, Presynaptic inhibition..., 1964). Pertanto, la zona in cui iniziano gli impulsi è incerta. Tuttavia, nelle cellule piramidali dell'ippocampo l'attivazione sinaptica del dendrite apicale, che si trova molto lontano dal soma, genera nel dendrite la scarica di un impulso che si propaga poi al soma e così all'assone. Le cellule piramidali sono disposte in un singolo strato dal quale fuoriescono ad angolo retto i dendriti apicali (v. fig. 11A); le collaterali di Schaffer (Sch) formano sinapsi eccitatorie sui dendriti in una zona definita, lontana dal soma. Queste sono le sinapsi indicate con g nella fig. 2B. È così possibile esaminare la relazione tra l'eccitazione sinaptica e la generazione di impulsi osservando i potenziali di campo extracellulari a varie profondità dalla superficie oltre il soma fino alle punte dei dendriti apicali, come nella fig. 11B. Nella regione delle sinapsi (profondità 0,7 e 0,8 mm) vi è una grande onda negativa che è il corrispettivo extracellulare dei PPSE che ivi si generano. Più vicino al soma, a 0,6 mm, si sovrappone un potenziale d'azione. Come è prevedibile, il campo del potenziale sinaptico si inverte a una profondità di 0,5-0,3 mm, ma il grande potenziale d'azione negativo aumenta, come si può vedere dalle misurazioni riportate nella fig 11C. Registrazioni più veloci prese a una profondità di 0,5-0,3 mm mostrano che il potenziale è progressivamente più tardivo, e i ritardi ci permettono di calcolare una velocità di conduzione di circa 0,4 m/s (v. Andersen e altri, 1966).
Così la fig. 11 fornisce una dimostrazione convincente che, in risposta a un'eccitazione sinaptica da parte delle collaterali di Schaffer, si genera un impulso nei dendriti proprio dalla parte somatica delle sinapsi e questo si propaga quindi verso il soma con la bassa velocità che ci si deve aspettare per dendriti piuttosto sottili. L'origine dendritica degli impulsi è stata dimostrata per altre specie di neuroni, per esempio le cellule di Purkinje del cervelletto, ed è presente anche in motoneuroni sottoposti a cambiamenti metabolici conseguenti alla sezione dei loro assoni effettuata alcuni giorni prima. È importante ricordare che su molte specie di neuroni, come quelli disposti in maniera irregolare e di dimensioni troppo esigue per la registrazione intracellulare, non è ancora stato possibile eseguire controlli sperimentali.
b) Motoneuroni tonici e motoneuroni fasici.
L'eccitazione sinaptica dei neuroni per mezzo di una singola scarica sincrona (v. figg. 5, B-N, 7 e 9, A-H) è un procedimento certo non fisiologico, ma giustificato perché ci chiarisce i meccanismi sinaptici. In condizioni fisiologiche i neuroni sarebbero depolarizzati da prolungate attivazioni sinaptiche asincrone mediante impulsi scaricati da organi recettori o da altri neuroni. In gran parte questa depolarizzazione può essere riprodotta dall'azione di una corrente depolarizzante costante applicata attraverso un elettrodo intracellulare. Molti motoneuroni (v. fig. 12A) e tutte le cellule piramidali della corteccia (v. fig. 12B) mostrano una risposta a lento adattamento, con una frequenza iniziale più alta che declina verso un livello mantenuto costante. Con questi motoneuroni cosiddetti tonici, c'è una relazione approssimativamente lineare fra intensità di corrente e frequenza di scarica per limiti di frequenze che eccedono quella prodotta da una stimolazione sinaptica prolungata. I rimanenti motoneuroni sono fasici, in quanto rispondono a una corrente costante con un rapido e notevole declino di frequenza talvolta fino alla cessazione completa della scarica.
Idealmente l'eccitazione naturale dei motoneuroni dovrebbe essere modellata da bombardamenti sinaptici asincroni controllati. Anche in tali condizioni, e mediante registrazione dai loro assoni nelle radici ventrali, i motoneuroni possono essere classificati in tonici e fasici con tutte le gradazioni intermedie, e pertanto sembra che questi diversi comportamenti non siano da attribuirsi a danno cellulare prodotto dall'elettrodo intracellulare o alla iniezione di ioni dovuta alle correnti direttamente applicate. Almeno in parte, i motoneuroni tonici sono quelli che innervano i muscoli a lenta contrazione che sono responsabili del tono posturale e del mantenimento di contrazioni costanti.
5. Azione sinaptica inibitoria.
a) Potenziali postsinaptici inibitori (PPSI).
Esiste una seconda categoria di sinapsi che si oppongono all'eccitazione e tendono a prevenire la generazione di impulsi da parte delle sinapsi eccitatorie; pertanto esse vengono chiamate sinapsi inibitorie (v. Eccles, The physiology..., e Ionic mechanism..., 1964). Il fatto che queste due modalità fondamentali di azione sinaptica regolino la generazione di impulsi da parte delle cellule nervose è ormai generalmente accettato. La via per la più semplice inibizione centrale è mostrata nella fig. 1A; si tratta della via per le azioni inibitorie sui motoneuroni illustrata nelle figg. 1D, 13, 14 e 16. Come mostrato nella fig. 13, A-C, l'attivazione di sinapsi inibitorie provoca un aumento del potenziale della membrana postsinaptica. Tale potenziale postsinaptico inibitorio, PPSI (D), è virtualmente l'immagine speculare del PPSE (E). Gli effetti delle singole sinapsi inibitorie (v. fig. 13, A-C) sulla cellula nervosa si sommano esattamente nella stessa maniera delle sinapsi eccitatorie; naturalmente l'inibizione dell'azione sinaptica eccitatoria si spiega con l'azione opposta che essa esercita sul potenziale della membrana postsinaptica (v. fig. 14, A e B).
Gli effetti prodotti sull'ampiezza e la direzione del PPSI mediante variazione del potenziale di membrana iniziale (v. fig. 13G) corrispondono esattamente ai cambiamenti che ci si dovrebbe aspettare di osservare se le correnti che generano i PPSI fossero dovute a ioni che si muovono seguendo i loro gradienti elettrochimici, dato che c'è un'inversione della corrente a circa −80 mV. Queste correnti sarebbero dovute all'aumento della permeabilità ionica della membrana subsinaptica prodotto sotto l'influenza della sostanza trasmettitrice inibitoria. Le condizioni che danno luogo alla generazione di un PPSI sono mostrate nello schema elettrico della fig. 13F: l'attivazione delle sinapsi aumenta notevolmente la conduttanza elettrica della resistenza nell'elemento di destra dello schema.
La fig. 14 è uno schema generale che illustra i cambiamenti di potenziale prodotti attraverso la membrana della cellula nervosa dall'attivazione delle sinapsi eccitatorie e inibitorie che la fig. 14C mostra distribuite sulla superficie del soma e dei dendriti. I PPSI e i PPSE di controllo sono riportati a sinistra e a destra della fig. 14A, mentre in mezzo sono riportati tracciati sovrapposti che mostrano le interazioni a tre diversi intervalli di tempo del PPSI in relazione al PPSE. È evidente che le sinapsi inibitorie contrastano efficacemente la depolarizzazione prodotta dalle sinapsi eccitatorie. In condizioni appropriate, l'azione sinaptica inibitoria può impedire la generazione di un impulso da parte della sinapsi eccitatoria, come si può vedere nella registrazione presa da un'altra cellula (v. fig. 14B). Pertanto, abbiamo qui la prova che queste sinapsi eccitatorie e inibitorie sono effettivamente in contrasto per il potenziale di membrana; se questo potenziale cade a livello critico, la cellula genera un impulso (v. fig. 14D) che può essere registrato mentre scende lungo l'assone. Pertanto, per quanto riguarda l'eccitazione e l'inibizione postsinaptica, conosciamo con chiarezza l'azione antagonista sul potenziale di membrana del motoneurone.
La fig. 15B mostra schematicamente il flusso di corrente che si determina per effetto di una sinapsi inibitoria attivata: tale flusso è diretto all'esterno attraverso la membrana subsinaptica lungo lo spazio sinaptico, e gira poi indietro per iperpolarizzare la membrana postsinaptica mediante un flusso verso l'interno sopra la sua intera superficie; è l'opposto di ciò che si verifica alla sinapsi eccitatoria (v. fig. 15A). La corrente diretta verso l'esterno attraverso la membrana subsinaptica inibitoria potrebbe essere dovuta al movimento verso l'esterno di un catione - come il potassio - o al movimento verso l'interno di un anione - come il cloro - oppure alla combinazione di movimenti di cationi e di anioni tale che vi sia un movimento netto di corrente verso l'esterno guidato da una batteria di circa −80 mV in serie con una resistenza abbastanza bassa.
La fig. 15 serve a illustrare i risultati più semplici sul PPSE e sul PPSI e sulla loro interazione. I potenziali di equilibrio approssimativi per gli ioni sodio, cloro e potassio sono indicati dalle linee orizzontali; si considera che il potenziale di equilibrio del cloro sia identico al potenziale di riposo della membrana. Nella fig. 15C a sinistra si vede che un PPSE è abbastanza grande da generare un potenziale d'azione, mentre il decorso del PPSE in assenza di potenziale d'azione è rappresentato dalla linea tratteggiata. La linea continua dello schema di destra rappresenta un PPSI iniziale, che diminuisce la depolarizzazione prodotta dalla stessa eccitazione sinaptica cosi che non è più in grado di generare un potenziale d'azione.
b) Meccanismi ionici dell'azione sinaptica inibitoria.
Nelle ricerche sperimentali sui meccanismi ionici dell'azione sinaptica inibitoria si deve variare il gradiente di concentrazione attraverso la membrana postsinaptica di una o dell'altra specie di ioni normalmente presenti; si deve inoltre impiegare una grande varietà di altri ioni, al fine di esaminare la permeabilità ionica della membrana subsinaptica. Nel caso delle sinapsi inibitorie che si trovano su cellule nervose o muscolari di Invertebrati, le ricerche sono in genere eseguite su preparati isolati. Cambiamenti della concentrazione ionica relativa attraverso la membrana postsinaptica si possono ottenere con facilità alterando la composizione ionica del mezzo esterno (v. articolo seguente, figg. 11 e 12). Questo metodo non è adatto per lo studio dei neuroni dei Mammiferi, perché questi devono essere esaminati in condizioni di normale circolazione del sangue e la contrazione del cuore sarebbe gravemente disturbata o soppressa dai notevoli cambiamenti che è necessario produrre nella composizione ionica del sangue. Invece, per alterare la composizione ionica all'interno della cellula postsinaptica è stata impiegata la procedura di iniettare ioni per via elettroforetica al di fuori dell'elettrodo intracellulare. Per esempio, le specie di anioni che possono passare attraverso la membrana inibitoria sono state identificate mediante l'iniezione dell'una o dell'altra specie dentro la cellula nervosa e osservando se l'aumento della concentrazione intracellulare modificava il potenziale postsinaptico inibitorio. Queste iniezioni vengono eseguite riempiendo i microelettrodi con sali che contengono gli anioni sotto esame. Quando il microelettrodo è inserito dentro la cellula nervosa, si può iniettare elettroforeticamente una determinata quantità dell'anione entro la cellula facendo passare una corrente appropriata attraverso il microelettrodo.
Il PPSI della fig. 16A è stato cambiato in un potenziale depolarizzante (v. fig. 16B) mediante l'aggiunta di circa 5 picoequivalenti di ioni cloro alla cellula, operazione con la quale si dovrebbe più che triplicarne la concentrazione, mentre l'iniezione di una quantità più che doppia di ioni solfato in un'altra cellula non modifica il PPSI (v. fig. 16, E ed F). Questo semplice esame ci permette di stabilire che, sotto l'azione del trasmettitore inibitorio, la membrana subsinaptica diventa momentaneamente permeabile agli ioni cloro, ma non a quelli solfato. Nella fig. 16, I-J, si vede che in due tipi di azione sinaptica inibitoria nel midollo spinale di Mammifero la membrana inibitoria era permeabile agli ioni nitrito, e che il recupero dall'effetto dell'iniezione ionica era completo in 2-3 minuti.
Va detto che la fig. 16I e la fig. 16J esemplificano due processi ben distinti di scambio ionico. Anzitutto, le aree subsinaptiche specializzate sotto l'influenza del trasmettitore inibitorio sviluppano momentaneamente una permeabilità ionica specifica di alto grado, determinando in tal modo il notevole aumento di conduttività della resistenza illustrato sulla destra della fig. 13F. Questa aumentata conduttanza ionica per 1-2 ms è responsabile dei flussi ionici che trasportano le correnti subsinaptiche inibitorie disegnate schematicamente nella fig. 15B. In secondo luogo la permeabilità ionica dell'intera membrana postsinaptica controlla la composizione ionica intracellulare del neurone ed è responsabile del suo ripristino alla normalità dopo che essa è stata alterata dall'iniezione di ioni. Nella fig. 16, I e J, occorrono poco più di 2 minuti affinché lo ione iniettato si disperda diffondendo verso l'esterno.
Prove sperimentali come quelle illustrate nella fig. 16 hanno dimostrato che 12 anioni possono passare attraverso i canali aperti dal trasmettitore inibitorio. Nella fig. 17 si vede che questi anioni permeabili (linee nere) sono caratterizzati dall'avere, allo stato idratato, piccoli diametri, mentre quelli degli anioni impermeabili sono più grandi. L'unica eccezione a questa generalizzazione è rappresentata dallo ione formiato; altrimenti la membrana inibitoria attivata è permeabile a tutti quegli anioni che nello stato idratato hanno un diametro di non più di 1,14 volte quello dello ione potassio, cioè non più di 2,85 A, che è la misura dello ione cloro idratato. I diametri degli ioni nella fig. 17 sono calcolati, dalle conducibilità equivalenti limite degli ioni, secondo la legge di Stokes, nell'ipotesi che gli ioni idratati siano sferici. È possibile che lo ione formiato idratato abbia una forma ellissoidale e pertanto riesca ad attraversare i pori della membrana che non permettono il passaggio a ioni sferici più piccoli. Simili serie di ioni permeabili e impermeabili, e perfino la permeabilità anomala dello ione formiato, sono state osservate in analoghe ricerche sulle sinapsi inibitorie dei Pesci, del rospo e della lumaca. Sarebbe davvero straordinario se il meccanismo ionico dell'inibizione centrale fosse uguale in tutto il regno animale (v. Eccles, The physiology... e Ionic mechanism..., 1964). Recentemente Eccles e altri (v., 1977) hanno visto che i PPSI molto grandi delle cellule piramidali dell'ippocampo (v. fig. 20, A, B e C) possiedono una capacità discriminativa fra gli anioni paragonabile a quella illustrata nella fig. 17.
Dato che normalmente il potassio si trova nella cellula ad alta concentrazione, l'iniezione intracellulare di ioni K+ non può produrre grandi modificazioni della sua concentrazione, anche perché l'ipertonicità determinata dal suo aumento provoca immediatamente un richiamo di acqua attraverso la membrana cellulare. Pertanto le tecniche di iniezione di ioni potassio non hanno fornito prove sicure a favore o contro il fatto che la permeabilità a tali ioni contribuisca alla genesi del PPSI. Ciò nonostante esistono prove indirette a favore del fatto che la permeabilità al potassio contribuisca in modo apprezzabile l'iperpolarizzazione del potenziale di equilibrio del potassio è di circa 20 mV maggiore di quella del potenziale di riposo (v. Eccles, 1957; v. Gustafsson, 1974); il potenziale di equilibrio nell'inibizione è pure in senso iperpolarizzante, ma meno, probabilmente soltanto di circa 6-10 mV (v. fig. 15). Queste considerazioni hanno suggerito l'idea che la membrana, sotto l'effetto di un processo inibitore, diventi permeabile sia agli ioni potassio sia a quelli cloro. L'ipotesi più semplice è che le due permeabilità siano uguali e che a determinare la permeabilità sia unicamente la grandezza degli ioni in stato idratato, siano essi cationi o anioni. Il criterio della grandezza è ovviamente sufficiente a escludere che intervengano gli ioni sodio, i quali allo stato idratato sono molto grandi, come indicato dalla linea superiore della fig. 17. Il potenziale di equilibrio di 80 mV per il PPSI si potrebbe raggiungere solamente se nella membrana soggetta a inibizione i pori fossero abbastanza piccoli da escludere praticamente in modo assoluto il passaggio degli ioni sodio; in caso contrario il flusso degli ioni sodio determinato dall'azione sinaptica produrrebbe depolarizzazione ed eccitazione. Di fatto, la differenza fondamentale tra sinapsi eccitatorie e inibitorie sta nel fatto che nel primo caso la permeabilità al sodio è grande, mentre nel secondo caso essa è trascurabile.
La fig. 18 illustra alcuni modelli di azione sinaptica inibitoria. In A e B si possono osservare due canali ionici attraverso il foglietto bimolecolare della superficie della membrana (v. Eccles, 19772, figg. 1-12). Una porta (A) viene chiusa e una (B) aperta dall'azione sterica della molecola di trasmettitore, che si è attaccata al sito del recettore. È stato stabilito che nel sistema nervoso centrale dei Mammiferi l'azione inibitoria è mediata da amminoacidi, e cioè glicina in alcune sinapsi, acido γ-amminobutirrico (GABA) in altre. Si ritiene che questi trasmettitori siano impacchettati nelle vescicole sinaptiche dei bottoni sinaptici inibitori (v. fig. 3) e vengano liberati a quanti nello spazio sinaptico. Per diffusione la molecola trasmettitrice troverebbe il suo sito sterico (v. fig. 18B) e riuscirebbe così a determinare un cambiamento conformazionale che aprirebbe la porta per circa 1 ms. In base alla teoria proposta originariamente da Coombs e altri (v., Excitatory synaptic..., 1955), gli ioni K+ e Cl- migrano per diffusione attraverso il canale ionico così aperto; ciò origina la corrente subsinaptica che genera il potenziale postsinaptico inibitorio (v. fig. 15B).
Sembra che i canali ionici che operano nell'inibizione non abbiano una specificità chimica per gli anioni: essi discriminano solamente in base alla grandezza. Per esempio è stato dimostrato che tutte le 11 specie di anioni che allo stato idratato sono più piccoli di una grandezza critica (2,9 Å) attraversano i canali indipendentemente dalle loro proprietà chimiche, mentre tutti quelli che sono più grandi sono esclusi. In questo modo la membrana, una volta attivata da un processo postsinaptico inibitorio, si comporta come un filtro avente pori di dimensione critica, come illustrato nella fig. 18C. La fig. 18D include sia un modello elettrico con i potenziali di equilibrio per ioni Cl- e K+, come nella fig. 15C, sia il modello elettrico di una membrana normale (v. Eccles, 19772, figg. 1-13B).
Lux e altri (v., 1970) e Lux (v., 1971) hanno criticato il modello della fig. 18, C e D, in base a prove con le quali è stato dimostrato che attraverso la membrana normale vi è una pompa che spinge il Cl- all'esterno della cellula. La dimostrazione principale è basata sul fatto che durante infusione endovenosa di acetato di ammonio l'attività delle sinapsi inibitorie è modificata e il potenziale di equilibrio del PPSI si avvicina al potenziale di membrana. Nello stesso tempo si vede che in seguito a iniezione intracellulare di Cl- si ha un rallentamento della velocità di declino del Cl- interno. Questi autori hanno pertanto postulato che normalmente attraverso la membrana vi sia una pompa che espelle lo ione cloro, che il potenziale di equilibrio del Cl- sia di conseguenza circa −80 mV e che la pompa sia inattivata in maniera irreversibile dagli ioni ammonio. Lux ha così potuto spiegare l'iperpolarizzazione del PPSI come dovuta solamente all'aprirsi delle porte per il Cl-, mentre gli ioni potassio non avrebbero alcun ruolo significativo. Risultati simili a quelli di Lux sono stati recentemente ottenuti da Nicoll (v., 1978) sulle risposte iperpolarizzanti indotte dal trasmettitore inibitorio (GABA) sui motoneuroni del midollo spinale isolato di rana. È stato, tuttavia, dimostrato recentemente (v. Allen e altri, 1977) che i risultati ottenuti da Lux sui motoneuroni non sono assolutamente riproducibili sui PPSI molto grandi dell'ippocampo (v. fig. 20), i quali non vengono modificati dall'ammonio. Anche per le cellule piramidali dell'ippocampo non vi sono prove dell'esistenza di una pompa che espelle lo ione cloro che dipenda dalla concentrazione interna di Cl-, come postulato da Lux; pertanto l'accoppiamento dei flussi degli ioni Cl- e K+ della fig. 18D potrebbe verificarsi in alcune importanti sinapsi inibitorie che si trovano nelle zone superiori del cervello.
Il meccanismo ionico che genera i PPSI dei motoneuroni, secondo l'ipotesi di Lux, è illustrato nella fig. 18, E ed F, in cui la batteria sul canale del Cl- (in F) indica il valore di −80 mV, che è il valore postulato per ECl, a causa della pompa che espelle Cl- verso l'esterno. La figura rappresenta il canale ionico impermeabile al K+. La fig. 18, G e H, mostra, con lo stesso tipo di rappresentazione, la modalità di azione del trasmettitore (anche in questo caso GABA) dell'inibizione presinaptica (v. anche fig. 27). Un'ulteriore descrizione verrà data in seguito. Nella fig. 18 non sono riportati i modelli di molte sinapsi inibitorie di Invertebrati, nelle quali l'apertura di canali ionici non dà luogo a cambiamenti del potenziale di riposo normale. In mancanza di un meccanismo di pompa per il Cl- questo ione si trova in equilibrio, ma l'inibizione può ugualmente aver luogo in quanto la depolarizzazione delle sinapsi eccitatorie è antagonizzata dal meccanismo ionico che tende a mantenere il potenziale di membrana al livello di riposo. È stato suggerito che, se vi sono cariche positive fisse sulle pareti del canale, gli anioni possono attraversarlo, mentre i cationi sono respinti, come è illustrato nella fig. 18, E e G.
I modelli del meccanismo dell'azione inibitoria centrale, illustrati nella fig. 18, D ed F, devono essere considerati provvisori. È necessaria ancora una importante massa di lavoro di tipo quantitativo per comprovare i vari aspetti dell'ipotesi di Lux. Vi sono molte caratteristiche anomale, particolarmente la strana efficacia dell'acetato intracellulare nel diminuire e perfino invertire i PPSI delle cellule piramidali dell'ippocampo (v. Allen e altri, 1977).
Gli ammirevoli studi di Kehoe (v., 1972) sulle cellule del ganglio pleurale di Aplysia suggeriscono livelli di complessità maggiore. I PPSI generati sia per azione sinaptica, sia dall'applicazione del trasmettitore (ACh), hanno un doppio decorso temporale. La componente rapida è dovuta a un aumento della conduttanza al Cl-, quella lenta a un aumento della conduttanza al K+: i potenziali di equilibrio dei due ioni sono rispettivamente 60 mV e −80 mV. Da questa interessante ricerca risulta che le due azioni evocate dallo stesso trasmettitore sono completamente indipendenti e che ciascuna conduttanza ionica è in ottimo accordo con le previsioni risultanti dalla composizione ionica in base all'equazione di Nernst.
In una ricerca simile Gerschenfeld e Paupardin-Tritsch (v., 1974) hanno dimostrato che in altri neuroni dei molluschi Helix e Aplysia il trasmettitore inibitorio è la serotonina, la quale genera anch'essa PPSI rapidi e lenti, dovuti a un aumento della conduttanza rispettivamente al Cl- e al K+.
6. Vie eccitatorie e inibitorie nel sistema nervoso centrale.
a) Introduzione.
Uno dei risultati delle recenti approfondite ricerche sul sistema nervoso centrale è stata la scoperta che quasi tutti i neuroni sono sottoposti ad azioni sinaptiche antagoniste eccitatorie e inibitorie. Un risultato ancora più notevole è l'ipotesi che un determinato neurone possa avere un solo tipo di azione sinaptica: se esso opera mediante sinapsi eccitatorie al livello di alcune delle sue terminali assoniche, opera in questo stesso modo in tutte le sue sinapsi e può avere un effetto inibitore solo attraverso l'interpolazione di un neurone, il quale a sua volta esercita un'azione inibitoria a livello di tutte le sinapsi formate dal suo assone (v. fig. 1A). Questa ipotesi ha portato a una formulazione più generale secondo la quale nel sistema nervoso centrale esisterebbero solo due classi di cellule nervose, quelle puramente eccitatorie e quelle puramente inibitorie. Si può aggiungere fra parentesi che probabilmente esiste una terza classe, quella delle cellule inibitorie presinaptiche; tuttavia, le prove a favore dell'esistenza di questa terza classe di cellule nervose sono meno convincenti, perché attualmente sembra dimostrato che la sostanza trasmettitrice sia l'acido γ-amminobutirrico (GABA), che è lo stesso trasmettitore che opera nella maggior parte delle sinapsi inibitorie di tipo postsinaptico, come sarà discusso nel successivo capitolo dedicato alle sostanze trasmettitrici sinaptiche. (Per una estesa trattazione delle vie inibitorie si rimanda al lavoro di Eccles, 1969).
b) Il sistema delle cellule di Renshaw.
La fig. 19 illustra un esempio, studiato molto a fondo, di semplice meccanismo postsinaptico inibitorio, disposto per operare come un sistema a retroazione (o feedback) negativa. Le collaterali assoniche dei motoneuroni evocano monosinapticamente scariche ripetitive rapide e prolungate (v. fig. 19B) dalle cellule di Renshaw mediante lo stesso trasmettitore eccitatorio (acetilcolina) che opera perifericamente alle sinapsi neuromuscolari (v. fig. 19A). Le cellule di Renshaw si trovano nella parte ventromediale delle corna anteriori del midollo spinale. I loro assoni decorrono in direzione dorsolaterale per formare sinapsi inibitorie sui motoneuroni e produrre PPSI grandi e relativamente prolungati (v. fig. 19D), i quali possono mostrare ondulazioni dovute alla scarica ritmica a circa 1.000/s della cellula di Renshaw (v. fig. 19C). La latenza del PPSI è calcolata paragonando le registrazioni intracellulari e quelle appena extracellulari; il PPSI inizia in corrispondenza della freccia in basso della fig. 19E e ha una latenza di 1,2 ms, che è appena sufficiente secondo la via illustrata in A, poiché la scarica della cellula di Renshaw può iniziare anche solo 0,65 ms dopo che la scarica antidromica è entrata nel midollo spinale, come indicato dalla freccia in alto della fig. 19E. Inoltre, la scarica ripetitiva della cellula di Renshaw è generata esclusivamente dalla semplice via eccitatoria monosinaptica attraverso le collaterali dell'assone motore (A). Non vi sono dimostrazioni dell'esistenza di vie eccitatorie polisinaptiche e pertanto la risposta ripetitiva (v. fig. 19B) è dovuta all'azione continuata del trasmettitore. Nella fig. 19A sono visibili anche diramazioni della cellula di Renshaw che si portano a contatto con un altro neurone inibitore il quale a sua volta inibisce il motoneurone. Abbiamo così dimostrato che questo neurone inibitore è quello rappresentato interpolato sulla via inibitrice Ia illustrata nella fig. 1A. Ovviamente lo schema della fig. 19A è notevolmente semplificato, come anche quello della fig. 1A. Molte delle fibre nervose e dei neuroni del tipo illustrato sono disposti in parallelo con divergenza e convergenza a ogni stazione sinaptica. (Sull'argomento v. Eccles e altri, 1954).
c) L'inibizione mediata dalle cellule a canestro dell'ippocampo.
La fig. 20 presenta alcune dimostrazioni che hanno portato a postulare che le cellule a canestro dell'ippocampo siano cellule inibitorie responsabili dei grandi e prolungati PPSI che le varie stimolazioni - commissurali, settali e locali - evocano in una cellula piramidale dell'ippocampo. Queste stesse stimolazioni generano anche grandi campi elettrici extracellulari, come illustrato nella fig. 20D in una serie presa lungo una traccia microelettrodica alle profondità indicate sulla scala, come nella fig. 11. L'aspetto più caratteristico di questi potenziali extracellulari è una grande onda positiva. Quando questa è misurata a una latenza appropriata (la linea tratteggiata della fig. 20D) al fine di minimizzare la contaminazione con potenziali negativi precedenti e posteriori, le misurazioni riportate nel diagramma rivelano un profilo di profondità con un chiaro massimo di positività a una profondità di circa 0,4 mm (v. fig. 20E), e questo si riscontra anche per gli altri due sistemi afferenti che producono grandi PPSI. Nella fig. 20F è rappresentata una cellula piramidale sulla stessa scala verticale (v. anche fig. 11A), che dimostra come il picco di positività avvenga alla profondità dei soma, i quali costituiscono l'unica zona in cui si concentrano le sinapsi delle cellule a canestro. Queste sinapsi (marcate con f nella fig. 2B), come dimostrato da Ramón y Cajal (v., 1911), sono formate dagli assoni delle cellule a canestro. Il profilo di profondità pertanto indica che dai soma originano potenti flussi di corrente, come indicato dalle frecce della fig. 20F; tale profilo è esattamente quello previsto nel caso che l'iperpolarizzazione dei PPSI sia generata dal denso aggregato di sinapsi delle cellule a canestro sui soma delle cellule piramidali. Come illustrato nella fig. 20F, le cellule a canestro sono attivate da collaterali degli assoni delle cellule piramidali; si tratta pertanto di un altro esempio di inibizione a fredback negativo. Le cellule a canestro scaricano in maniera ripetitiva come le cellule di Renshaw (v. fig. 19B), ma in genere solo con 3 o 4 scariche in risposta a stimoli come quelli della fig. 20, A-C.
d) Circuiti neuronali nel cervelletto.
Il più complesso sistema di vie centrali eccitatorie e inibitorie che sia stato finora chiarito si trova nel cervelletto (v. Eccles e altri, 1967). Le ricerche istologiche (v. Ramón y Cajal, 1911) hanno rivelato che nella corteccia cerebellare vi sono cinque principali tipi di neuroni e che questi hanno una disposizione laminare come illustrato nello schema della fig. 21, che mostra lo strato molecolare superficiale con le cellule stellate e a canestro, e il più profondo strato granulare con i granuli e le cellule di Golgi, separati dallo strato dei corpi delle cellule di Purkinje. Ricerche come quelle illustrate nella fig. 22 hanno portato a formulare un'ipotesi abbastanza completa sul modo di operare di queste diverse specie cellulari nell'elaborazione delle informazioni nella corteccia cerebellare. Nella fig. 21 si vede che l'afferenza principale alla corteccia cerebellare, quella mediata dalle fibre muscoidi, è in rapporto sinaptico con i granuli, i cui assoni risalgono allo strato molecolare, dove si biforcano per formare le fibre parallele che corrono per 1-2 mm in ambedue le direzioni. Lo schema in prospettiva della fig. 22A mostra la disposizione strettamente parallela di queste fibre (fp) durante il loro decorso lungo un folium formando angoli retti con le branche dendritiche delle cellule di Purkinje e di quelle a canestro disposte a spalliera. La fig. 21 mostra una sezione lungo un folium, e pertanto i dendriti possono essere schematizzati come una singola proiezione conica che parte dai corpi cellulari.
Nella fig. 22A mediante un elettrodo di superficie si stimolano direttamente le fibre parallele, in modo da produrre un ‛raggio' di fibre eccitate. La fig. 22, B e C, mostra la registrazione intracellulare da una cellula di Purkinje che si trova ‛sul raggio' in B e ‛fuori del raggio' in C. Nella traccia più amplificata della fig. 22B si vede che il PPSE iniziale si innalza fino al punto d'origine del potenziale d'azione, indicato dalla freccia, come nella fig. 9, B-D, ma il potenziale d'azione sembra che venga iniziato da una depolarizzazione di soli 3,5 mV circa, valore molto inferiore a quelli mostrati nelle figg. 9 e 10. Vi sono dati che indicano che questa differenza è dovuta al fatto che il potenziale d'azione è generato nei dendriti delle cellule di Purkinje (v. fig. 11), dove, naturalmente, la depolarizzazione sinaptica è molto più grande di quella registrata dopo trasmissione elettrotonica al soma. La registrazione dalla cellula di Purkinje ‛fuori raggio' (v. fig. 22C) mostra un PPSI puro, che ha un decorso temporale quasi altrettanto lento di quelli delle cellule piramidali dell'ippocampo (v. fig. 20, A-C). Molte prove dimostrano che il PPSI delle cellule di Purkinje è dovuto all'azione sinaptica inibitoria delle cellule a canestro, che mandano i loro assoni trasversalmente fino a 1 mm attraverso il folium per formare dense terminazioni sinaptiche sui soma delle cellule di Purkinje (v. fig. 22A). Questa concentrazione di sinapsi inibitorie è simile a quella delle sinapsi delle cellule a canestro sui soma delle cellule piramidali dell'ippocampo (v. fig. 20). La distribuzione trasversale dell'inibizione mediata dalle cellule a canestro fa sì che l'eccitazione molto localizzata di un gruppo di fibre muscoidi che proiettano ai granuli con conseguente formazione di un ‛raggio' di fibre parallele eccitate sia in grado di produrre un effetto con una ben precisa organizzazione. Nel raggio vi sarà pertanto una zona dominante di eccitazione sulle cellule di Purkinje con una zona di inibizione su ciascun lato (v. Eccles, 1969).
La fig. 21 mostra, marcati in nero, i neuroni che sappiamo avere azione inibitoria. Gli unici neuroni eccitatori sono i granuli. Anche i due tipi di fibre afferenti, muscoidi e rampicanti, sono eccitatori. Le cellule stellate sono simili alle cellule a canestro. Le cellule di Golgi sono eccitate principalmente dalle fibre parallele e inibiscono la sinapsi fra fibra muscoide e granulo e pertanto hanno una semplice azione a feedback negativo, a differenza dell'inibizione prodotta dalle cellule a canestro e dalle cellule stellate che è di tipo feedforward.
Le cellule di Purkinje nella fig. 21 sono anch'esse marcate in nero, perché, come illustrato nella fig. 23, esercitano un'azione inibitoria mediante le loro sinapsi sui neuroni dei nuclei del cervelletto e del nucleo di Deiters. I PPSI progressivamente più grandi del neurone di Deiters (v. fig. 23, B-E) sono stati evocati stimolando direttamente un numero sempre maggiore di cellule di Purkinje che proiettano a quel neurone, e assomigliano ai PPSI graduati della fig. 13, A-C. I tracciati più lenti della fig. 23, F-H, mostrano l'inibizione di un neurone di Deiters che scarica in maniera regolare (v. fig. 23G) con un più tardivo rimbalzo di attività (v. fig. 23, G e H). Vedremo che l'input proveniente dalle fibre muscoidi e diretto alla corteccia cerebellare genera molte complesse vie di interazione sinaptica.
Oltre alla complessa interazione sinaptica dovuta alle afferenze muscoidi vi è la potente azione eccitatoria della fibra rampicante. Come mostrato nella fig. 21, una cellula di Purkinje viene eccitata da una singola fibra rampicante che si attorciglia intorno ai dendriti formando centinaia di contatti sinaptici, così che un singolo impulso evoca un intensa depolarizzazione (v. fig. 22D) che dà luogo a una scarica ripetitiva della cellula di Purkinje (v. fig. 22E).
La fig. 23A fornisce uno schema semplificato delle linee di entrata e di uscita di un elemento della corteccia cerebellare. Si vede che la fibra rampicante (fr) fornisce una collaterale assonica a una cellula dei nuclei cerebellari o del nucleo di Deiters (cnsc) e prosegue per dare una potente eccitazione alle cellule di Purkinje. Anche la fibra muscoide (fm) fornisce una collaterale eccitatoria alle cnsc e prosegue per eccitare il granulo (g), il cui assone si biforca per formare la fibra parallela (fp) che eccita sia la cellula a canestro (cc) sia la cellula di Purkinje (cP), la quale ultima inibisce le cnsc. È da rilevare che la cellula a canestro esercita un'azione disinibitoria sulle cellule dei nuclei sottocorticali in virtù della sua azione inibitoria sulle cellule inibitorie di Purkinje.
Si può postulare che l'azione sinaptica eccitatoria sui nuclei sottocorticali esercitata dalle collaterali assoniche delle fibre muscoidi e rampicanti serva a fornire un sottofondo di eccitazione che è soggetto all'azione inibitoria delle cellule di Purkinje, che costituiscono la via d'uscita della corteccia cerebellare. Questa efferenza acquista una forma spazio-temporale imprimendo una tendenza inibitoria al sottofondo di eccitazione dei neuroni dei nuclei sottocorticali. Come risulta dagli schemi delle figg. 21 e 23A, la cellula di Purkinje costituisce l'unica linea d'uscita dalla corteccia cerebellare. Grazie ai tipi di scarica d'impulsi descritti, l'attività integrativa della corteccia cerebellare si riflette sulle cellule dei nuclei sottocorticali. Inoltre, è importante tener presente che le cellule di Purkinje sono in uno stato di incessante attività di fondo modulata dalle afferenze verso la corteccia cerebellare e che esercitano un'azione inibitoria sulle cellule dei nuclei sottocorticali. Di conseguenza queste cellule trasmettono in basso, lungo i loro assoni, un'immagine negativa del modello temporale delle scariche delle cellule di Purkinje.
Un altro aspetto della complessità dei circuiti inibitori della corteccia cerebellare è illustrato nella fig. 21. Le collaterali assoniche delle cellule di Purkinje (caP) formano sinapsi inibitorie sugli interneuroni inibitori della corteccia cerebellare (cellule a canestro, stellate esterne e di Golgi) e anche sulle cellule di Purkinje. Tale azione inibitoria non è molto potente e può non essere molto significativa nel funzionamento della corteccia cerebellare, ma è importante, perché dimostra che le cellule di Purkinje hanno la stessa azione inibitoria a livello di tutte le loro sinapsi.
e) Caratteristiche operative delle vie inibitorie.
Le figg. 19, 20 e 21 illustrano alcuni esempi di vie inibitorie che hanno caratteristiche di circuiti a feedback negativo; successivamente ne sono stati identificati altri sei (v. Eccles, 1969), le cui caratteristiche generali, comuni a tutti, sono riportate nella fig. 24A. Le collaterali assoniche di un neurone eccitatorio eccitano un neurone inibitorio che esercita un'azione sinaptica inibitoria sul primo neurone o su un neurone adiacente della stessa natura. La fig. 24B illustra il tipo alternativo di via inibitoria, quella dell'inibizione che opera con meccanismo ad azione in avanti (feedforward); in questa via le collaterali delle fibre eccitatorie a un neurone eccitano un interneurone inibitorio che certe volte fornisce inibizione in avanti non solo, parzialmente, su quei neuroni che sono eccitati da quelle fibre, ma anche, e in grado molto maggiore, su altri neuroni (v. fig. 21), altre volte a neuroni con funzione opposta (v. fig. 1A). Nel sistema nervoso centrale dei Mammiferi sono stati finora trovati in totale quindici esempi di vie con inibizione feedforward; uno degli esempi meglio conosciuti è l'azione inibitoria esercitata dalle fibre del gruppo la sui motoneuroni a funzione antagonista passando attraverso gli interneuroni (v. fig. 1A; v. Eccles, 1969).
Nonostante questa imponente lista di 24 vie inibitorie nel sistema nervoso centrale dei Mammiferi, non vi è alcun esempio di cellula nervosa ambivalente, cioè che eserciti un'azione eccitatoria mediante alcune delle sue diramazioni assoniche e un'azione inibitoria mediante altre. Originariamente era stato supposto che gli interneuroni inibitori fossero interpolati semplicemente per operare una trasformazione nella sostanza trasmettitrice sinaptica, come se - per usare una frase di Granit - fossero dispositivi a tipo commutatore ed esercitassero un'azione molto localizzata con assoni corti limitati alla sostanza grigia; questo, in effetti, era il caso per gli esempi allora conosciuti nel midollo spinale. Ma ricerche molto estese, specialmente ai livelli più alti del sistema nervoso e in particolare nel cervelletto (v. Eccles e altri, 1967), hanno fornito esempi di complesse funzioni integrative operate dai neuroni inibitori (v. figg. 21, 22 e 23), alcuni dei quali hanno assoni misurabili in centimetri (v. Wilson e altri, 1970). Un'altra importante scoperta è stata l'aver trovato che i neuroni inibitori possono inibire altri neuroni inibitori ed esercitare così un'azione eccitatoria mediante una disinibizione. Questo fatto è stato riscontrato per la prima volta da Wilson e Burgess (v., 1962) per il sistema delle cellule di Renshaw, nel quale l'attivazione di alcune cellule di Renshaw può produrre una depolarizzazione nei motoneuroni mediante l'inibizione della scarica di fondo di altri neuroni inibitori (v. fig. 19A). Si conoscono attualmente molte altre vie disinibitorie, particolarmente nel cervelletto, dove le cellule a canestro e le cellule stellate inibiscono le cellule di Purkinje che a loro volta sono inibitorie (v. figg. 21, 22 e 23; v. Eccles e altri, 1967).
f) Inibizione per mezzo di sinapsi reciproche.
Negli schemi della fig. 24 si presume che l'eccitazione sinaptica dei neuroni inibitori generi scariche di impulsi (v. fig. 19B) e che l'azione inibitoria sia la conseguenza del passaggio di tali impulsi alle sinapsi inibitorie che liberano trasmettitore. Tale processo sarà descritto in dettaglio nell'articolo seguente, nel quale si mostrerà anche come per la liberazione di trasmettitore da una terminazione presinaptica l'impulso presinaptico non è essenziale; infatti è sufficiente produrre una equivalente depolarizzazione della terminazione mediante l'applicazione di corrente (v. figg. 8, 9, 18 e 20 dell'articolo seguente). È stato ora dimostrato che in certi neuroni inibitori la liberazione di trasmettitore è prodotta in modo analogo, cioè da una diffusione elettrotonica della depolarizzazione e non dalla depolarizzazione prodotta da un impulso.
Rall e altri (v., 1966) e Nicoll (v., 1969) hanno fornito una prova convincente, su base sia fisiologica sia istologica, che nel bulbo olfattorio (v. fig. 25A) i granuli (g) inibiscono le cellule mitrali (m) mediante una particolarissima disposizione sinaptica. Per mezzo della microscopia elettronica è stato dimostrato che i dendriti secondari delle cellule mitrali (ds) hanno sinapsi sulle gemmule che emergono dalle diramazioni dei granuli nello strato plessiforme esterno (spe); queste stesse gemmule a loro volta formano sinapsi sui dendriti delle cellule mitrali, come illustrato nella ricostruzione grafica della fig. 25B. In B le vescicole sinaptiche nel dendrite della cellula mitrale (ds) mostrano che il dendrite stesso è in quel punto presinaptico rispetto ai granuli (g) con conseguente trasmissione sinaptica nella direzione della freccia superiore. La sinapsi più in basso è in direzione reciproca. L'analisi dei potenziali di campo registrati in profondità attraverso la struttura laminare del bulbo olfattorio (si veda la tecnica illustrata nelle figg. 11 e 20) rivela che le prime sinapsi sono eccitatorie; cioè i dendriti delle cellule mitrali eccitano direttamente le gemmule dei granuli. Per mezzo dell'analisi dei potenziali di campo e della registrazione intracellulare dalle cellule mitrali (v. fig. 25C; v. Nicoll, 1969) si può vedere che, dopo un ritardo di circa 3 ms, l'eccitazione delle cellule mitrali produce un PPSI nelle cellule mitrali stesse, anche in quelle inizialmente non eccitate. Pare dunque che questa inibizione avvenga in assenza di scarica di impulsi nei granuli.
Pertanto la proposta via inibitoria da cellula mitrale a cellula mitrale avviene tramite una connessione molto breve attraverso i granuli, che sono cellule inibitorie, tanto che tale processo può avvenire perfino in una singola gemmula. È stato rilevato che questa azione sinaptica reciproca può aver luogo in assenza di impulsi. Innanzi tutto, gli impulsi nei soma delle cellule mitrali depolarizzano i dendriti secondari tramite diffusione elettrotonica della depolarizzazione, in conseguenza della quale viene liberato il trasmettitore eccitatorio e si ha una depolarizzazione delle gemmule (freccia superiore della fig. 25B). In secondo luogo, questa depolarizzazione delle branche costellate di gemmule dei granuli dà luogo a liberazione del trasmettitore inibitorio per le loro sinapsi inibitorie (freccia inferiore della fig. 25B) con la conseguente iperpolarizzazione dei dendriti iniziali secondari. Il significato fisiologico di questo meccanismo sinaptico reciproco può essere compreso se si tiene presente che le branche costellate di gemmule dei granuli e i dendriti secondari delle cellule mitrali formano una fitta rete. Pertanto, le cellule mitrali possono inibirsi efficacemente l'una con l'altra attraverso le loro sinapsi reciproche.
Queste nuove scoperte indicano un'altra modalità con cui gli interneuroni inibitori possono generare l'inibizione postsinaptica. Sinapsi reciproche come quelle descritte esistono nelle cellule amacrine e orizzontali della retina (v. Dowling e Boycott, 1966), anch'esse inibitorie.
Queste scoperte fondamentali sui meccanismi sinaptici del bulbo olfattorio e della retina hanno aperto la strada per comprendere molti dati, che finora erano rimasti oscuri, sulla struttura sinaptica dei nuclei di relay delle vie afferenti alla corteccia cerebrale. In questi nuclei vi sono interneuroni ad assone corto (neuroni di tipo II di Golgi) i quali assomigliano in generale ai granuli della fig. 25, in quanto agiscono come meccanismi inibitori locali; essi formano infatti sinapsi inibitorie sui dendriti dei neuroni di relay. Come indicato nella fig. 26 per il nucleo genicolato laterale (v. Hamori e altri, 1974), tuttavia, vi sono tre differenze: prima di tutto, l'eccitazione sinaptica delle cellule di Golgi (Go) avviene nel modo consueto, cioè per mezzo di impulsi nelle fibre afferenti (ret); in secondo luogo, si riscontra spesso un'organizzazione sinaptica a triade, come illustrato nella microfotografia della fig. 26 e all'interno del cerchio tratteggiato nel diagramma sottostante; in terzo luogo, l'assone della cellula di Golgi forma sinapsi inibitorie con i dendriti della cellula di relay (rel). Il principale meccanismo d'azione consiste in questo: un impulso afferente in ret eccita sia il dendrite della cellula di relay, sia la protrusione che emerge dal dendrite della cellula di Golgi; quest'ultima a sua volta inibisce il dendrite della cellula di relay. È probabile che questa organizzazione serva a depolarizzare efficacemente la protrusione della cellula di Golgi, che conseguentemente produce un'efficace azione inibitoria. Ma il dendrite della cellula di Golgi è eccitato anche in altre zone al di fuori della triade. Sembra probabile che, come per le cellule mitrali del bulbo olfattorio, la trasmissione elettrotonica lungo il dendrite della cellula di Golgi sia sufficiente per integrare le varie depolarizzazioni e inibire così molte cellule di relay. Esiste anche un terzo meccanismo inibitorio, come avviene convenzionalmente, cioè per mezzo delle sinapsi assodendritiche mostrate dalla cellula Go a livello del soma e dei dendriti della cellula rel.
Oltre che nel nucleo genicolato laterale della scimmia (v. Hamori e altri, 1974), meccanismi d'inibizione essenzialmente simili sono stati identificati nel nucleo genicolato mediale del gatto (v. Morest, 1971), nel nucleo genicolato mediale e in quello laterale del gatto (v. Jones e Powell, 1969), nel nucleo talamico ventrobasale del gatto (v. Ralston e Herman, 1969) e nel collicolo superiore (v. Lund, 1969). Inoltre, Sloper (v., 1971) ha descritto sinapsi dendro-dendritiche che sarebbero presenti, anche se raramente, nelle lamine profonde della corteccia motoria dei Primati. Purtroppo non esistono studi complementari di elettrofisiologia che ci possano dare una valutazione quantitativa dell'efficacia inibitrice delle sinapsi dendro-dendritiche rispetto alla sinapsi classica formata dagli assoni di neuroni inibitori (v. fig. 26). Nel bulbo olfattorio le cose stanno diversamente; infatti la prima scoperta è stata fatta con tecniche di elettrofisiologia. L'argomento è stato trattato perfettamente da Shepherd (v., 1974), che fornisce anche una completa descrizione delle sinapsi dendro-dendritiche nella retina e nel talamo.
7. Inibizione presinaptica.
Un'altra modalità di controllo inibitorio nel sistema nervoso centrale è stata individuata per la prima volta da Frank e Fuortes (v., 1957), ma sono occorsi diversi anni prima che fosse capito il meccanismo dell'inibizione presinaptica e fosse riconosciuta l'importanza del suo ruolo nel controllo a feedback negativo delle vie sensitive (v. Eccles, The physiology... e Presynaptic..., 1964; v. Schmidt, 1971). Il modo di operare dell'inibizione presinaptica è illustrato sopra il neurone nella fig. 27, nella quale si vede la sinapsi inibitoria presinaptica che forma una sinapsi sulla sinapsi eccitatoria; sono state viste molte di tali sinapsi nelle fotografie prese al microscopio elettronico (v. Eccles, The physiology..., 1964; v. Schmidt, 1971). La seguente sequenza di eventi spiega l'inibizione presinaptica. Il bottone sinaptico eccitatorio viene depolarizzato dall'azione di un trasmettitore chimico (v. fig. 27C); conseguentemente in questo bottone un potenziale d'azione diminuisce d'ampiezza e la produzione della sostanza trasmettitrice eccitatoria decresce, come è stato dimostrato per le sinapsi periferiche (v. articolo seguente, fig. 17). Si postula così che l'azione inibitoria sia esercitata sulle terminazioni presinaptiche eccitatorie e sia invece nulla sulla membrana postsinaptica: pertanto, è chiamata inibizione presinaptica. L'inibizione presinaptica è esercitata sulle terminazioni sinaptiche centrali di tutti i tipi di grandi fibre afferenti che sono stati esaminati finora. Alla prima stazione sinaptica l'inibizione presinaptica è in genere di potenza maggiore dell'inibizione postsinaptica. Tutte le grandi fibre afferenti esercitano un'azione inibitoria presinaptica sulle terminazioni centrali delle fibre afferenti, ma la distribuzione dipende in larga misura dalle rispettive modalità delle fibre afferenti (v. Eccles, Presynaptic..., 1964; v. Schmidt, 1971). L'inibizione presinaptica può essere messa in evidenza da vari procedimenti sperimentali. Una prova diretta consiste nel registrare dalla fibra eccitatoria tramite un elettrodo intracellulare e nell'osservare direttamente la depolarizzazione, come nei tracciati della fig. 27C, di cui quelli superiori mostrano i potenziali all'interno delle fibre che si osservano quando si applicano 1, 2 o 4 stimoli al nervo afferente condizionante. Un altro metodo consiste nel registrare intracellularmente da un neurone che sia stato eccitato sinapticamente e mostrare che il potenziale postsinaptico eccitatorio è ridotto, per la diminuita produzione di trasmettitore eccitatorio: nella fig. 27A, la depressione del PPSE ritarda o previene la generazione della scarica di impulsi, che è invece presente nel controllo (CON) in corrispondenza della freccia; nella fig. 27B, dopo un tetano condizionante si vede che il PPSE è notevolmente ridotto rispetto al controllo, ma non è modificato nel suo decorso temporale. Un metodo molto semplice per valutare l'inibizione presinaptica consiste nel determinare la diminuzione della scarica di impulsi evocati da un'afferenza eccitatoria di saggio, come è illustrato nella fig. 27D: la prima risposta è il potenziale di popolazione di controllo. Questo esperimento è simile a quello usato per valutare l'inibizione postsinaptica nella fig. 1D e non è discriminativo. Tutte queste prove sono state usate sistematicamente. Un altro modo di dimostrare l'inibizione presinaptica, che tecnicamente è il più semplice, consiste nel saggiare l'eccitabilità delle fibre eccitatorie sottoposte all'inibizione presinaptica, cioè delle fibre afferenti primarie, come la fibra eccitatoria presinaptica nella fig. 27: infatti, quando sono depolarizzate da un'azione inibitoria presinaptica diventano più eccitabili (v. fig. 27C). Tuttavia, questo metodo usato da solo non è attendibile, perché l'eccitabilità può essere aumentata da altri meccanismi depolarizzanti, come per esempio un aumento della concentrazione extracellulare di potassio.
Frank e Fuortes (v., 1957) nel loro primo rapporto sull'inibizione presinaptica affermarono che il neurone postsinaptico non mostrava cambiamenti nelle sue proprietà elettnche, come il potenziale di membrana o l'eccitabilità. Questo fatto ha ricevuto un'importante conferma da parte di Eide e altri (v., 1968), come è illustrato nella fig. 27, E-G, i cui tracciati inferiori mostrano il PPSE monosinaptico di controllo (E) e la sua diminuzione (F e G) per effetto dell'inibizione presinaptica a due intervalli di saggio; i tracciati superiori mostrano che la membrana postsinaptica quando è saggiata con un breve impulso applicato intracellularmente non mostra alterazioni. Inoltre, durante l'inibizione presinaptica non si rilevano cambiamenti nel decorso temporale del PPSE (v. fig. 27B; v. Eccles, Presynaptic..., 1964; v. Eide e altri, 1968); pertanto tutte le più fini prove sperimentali concordano con l'ipotesi che l'inibizione presinaptica sia davvero esclusivamente presinaptica (v. Eccles, The physiology... e Presynaptic..., 1964; v. Eide e altri, 1968; v. Schmidt, 1971). Non sembra pertanto necessario prendere in ulteriore considerazione la proposta alternativa che essa sia dovuta a sinapsi postsinaptiche inibitorie che agiscono così remotamente sui dendriti del neurone postsinaptico, da non essere possibile rilevare alcun effetto registrando dal soma con un elettrodo intracellulare.
Un ulteriore motivo di confusione nasce dal tentativo di spiegare l'effetto inibitore presinaptico sulla base dell'accumulo extracellulare di potassio, che ovviamente si verifica come conseguenza di un'intensa attività neuronale. Tuttavia, Nicoll (v., 1976) ha dimostrato che l'inibizione presinaptica ha un decorso temporale del tutto diverso da quello dell'accumulo di potassio, il quale in ogni caso genera una depolarizzazione molto più piccola.
Esistono ormai molti dati che concordano sul fatto che il trasmettitore dell'inibizione presinaptica sia il GABA. Questo mediatore depolarizza le terminazioni presinaptiche delle fibre afferenti primarie, con un meccanismo simile a quello delle terminazioni presinaptiche inibitorie. Ambedue le azioni (sia quella dovuta al GABA, sia quella dovuta alle terminazioni presinaptiche) vengono depresse selettivamente dalla bicucullina e dalla picrotossina. Il GABA depolarizza le terminazioni presinaptiche o le cellule dei gangli delle radici dorsali, soprattutto aprendo i canali per il Cl- (v. Nicoll, 1975 e 1976; v. Nishi e altri, 1974; v. Gallagher e altri, 1975), come indicato nella fig. 18, G e H. Il potenziale d'equilibrio per questo meccanismo ionico è a circa −30 mV, il che indica che nelle fibre presinaptiche vi è una pompa che spinge il Cl- all'interno e di conseguenza ECl Si trova a circa 40 mV nella direzione della depolarizzazione rispetto al potenziale di riposo. La concentrazione intracellulare di Cl- deve pertanto raggiungere valori di 50 mM. Nella rana si raggiungono simili valori di ECl perfino in cellule così periferiche come quelle dei gangli delle radici dorsali, il che è in accordo col fatto che il GABA apre i canali per il Cl- sia nell'inibizione presinaptica che in quella postsinaptica. Infine con tecniche di immunoistochimica è stata dimostrata nelle terminazioni presinaptiche la presenza di decarbossilasi dell'acido glutammico (GAD), che è l'enzima che sintetizza il GABA.
Ciò nonostante la distribuzione dell'inibizione presinaptica segue una precisa organizzazione o schema; tale schema dipende dalla classe o modalità delle fibre afferenti sulle quali si esercita l'inibizione presinaptica: le fibre afferenti cutanee sono le più fortemente influenzate da questa azione (v. Eccles, Presynaptic..., 1964; v. Schmidt, 1971).
Un altro dato generale è che l'inibizione presinaptica è molto più efficace al livello afferente primario che a livelli più alti del sistema nervoso. Tuttavia, è stato dimostrato che ha una significativa influenza inibitoria anche sulle vie che passano attraverso il talamo e il corpo genicolato laterale. Non ci sono per ora dati in favore dell'esistenza di inibizione presinaptica ai livelli più alti del cervello dei Mammiferi - le cortecce (neocorteccia e archicorteccia) cerebellare e cerebrale.
8. Sostanze trasmettitrici sinaptiche.
a) Trasmettitore eccitatorio.
In molte specie di neuroni del midollo spinale il PPSE o il PPSI prodotti da una scarica presinaptica sincrona possono raggiungere il massimo in poco più di 1 ms e avere una durata totale di 10-20 ms (v. figg. 5, 6 e 13). È stato dimostrato (v. fig. 6, A e B) che il flusso di corrente ionica attribuibile all'azione del trasmettitore cessa praticamente entro 2 ms, e che il successivo declino esponenziale tardivo, la cui velocità dipende dalla sua costante di tempo elettrica (v. Eccles, The physiology..., 1964), è da attribuirsi al recupero passivo della membrana del neurone. Tuttavia in altre sinapsi spinali, specialmente quelle formate dalle collaterali assoniche del motoneurone sulle cellule di Renshaw, normalmente il trasmettitore continua ad agire fino a 3050 ms (v. figg. 6C e 19B) e perfino per più di un secondo quando si previene la distruzione enzimatica del trasmettitore (acetilcolina, ACh) con una sostanza anticolinesterasica come l'eserina (v. Eccles, The physiology..., 1964; v. fig. 23D). A livelli soprasegmentali del sistema nervoso, i PPSE e i PPSI prodotti da singole scariche sincrone sono più lunghi di quelli delle figg. 5 e 13 (v. i PPSI delle flgg. 20, A-C, 22C e 23, B-H). Questa lunga durata deve essere attribuita, per gran parte, all'azione prolungata del trasmettitore e non alla più lunga costante di tempo elettrica della membrana postsinaptica. Salvo il caso del trasmettitore acetilcolina, non è stato possibile scoprire alcun enzima in grado di distruggere efficacemente i trasmettitori liberati nello spazio sinaptico. Probabilmente le azioni prolungate del trasmettitore sinaptico sono dovute alla lenta diffusione del trasmettitore al di fuori dello spazio sinaptico e dentro gli stretti canali fra i neuroni e la glia.
L'unico trasmettitore eccitatorio che è stato dimostrato in maniera sicura nel sistema nervoso centrale dei Mammiferi è l'acetilcolina, la quale media l'azione eccitatoria delle collaterali assoniche del motoneurone sulle cellule di Renshaw (v. fig. 19A; v. Eccles, The physiology..., 1964; v. Curtis, Central synaptic..., 1969). Meno convincenti sono le prove che l'acetilcolina sia il trasmettitore in molte parti del cervellò, specialmente nel talamo e nella corteccia cerebrale (v. Curtis, Central synaptic..., 1969), ma i recettori per l'ACh che si trovano su queste cellule si comportano in maniera molto diversa, sia fisiologicamente sia farmacologicamente, dai recettori per l'ACh che si trovano nelle cellule di Renshaw. Probabilmente la maggior parte delle azioni del trasmettitore eccitatorio sono mediate dagli amminoacidi a carattere acido, L-glutammico o L-aspartico. L'applicazione elettroforetica di queste sostanze ai neuroni determina risposte molto somiglianti all'azione sinaptica eccitatoria, ma purtroppo finché non si saranno trovate sostanze bloccanti specifiche delle sinapsi eccitatorie nel sistema nervoso centrale non sarà possibile eseguire esperimenti definitivi (v. Curtis, Central synaptic..., 1969).
b) Trasmettitori inibitori.
In contrasto con i deludenti risultati dell'intensa ricerca di sostanze trasmettitrici eccitatorie nel sistema nervoso centrale, si sono ottenuti grandi successi nell'identificazione dei trasmettitori inibitori, in larga misura dovuti al ruolo decisivo di sostanze bioccanti specifiche, la stricnina e i suoi analoghi da una parte, la bicucullina dall'altra. Conviene considerare in primo luogo le sinapsi inibitorie che sono bloccate dalla stricnina, perché su questo argomento vi è accordo sostanziale (v. Curtis, The pharmacology..., 1969).
Nella fig. 28, A e B, si vedono PPSI disinaptici e PPSI monosinaptici di un motoneurone (v. figg. 5 e 13), mentre nella fig. 28, C e D, si vede quale risultato si ottiene invariabilmente con la somministrazione di stricnina: una notevole depressione del PPSI, mentre il PPSE rimane inalterato (v. Eccles, The physiology..., 1964; v. Curtis, The pharmacology..., 1969). La fig. 28, E ed F, mostra che l'applicazione elettroforetica sia di glicina sia di GABA aumenta il potenziale di membrana di un motoneurone. Questo effetto sul neurone assomiglia in ogni aspetto al PPSI prodotto da un'azione sinaptica inibitoria. Per esempio, in tutti gli esperimenti erano simili i potenziali di equilibrio (v. fig. 130), i notevoli aumenti della conduttanza di membrana per gli stessi ioni (v. fig. 17) e le diminuzioni dell'eccitabilità del neurone. Inoltre, la fig. 28, E ed F, mostra che le iperpolarizzazioni dovute sia alla glicina sia al GABA sono rapidamente reversibili al cessare dell'applicazione elettroforetica.
Tuttavia l'applicazione di stricnina permette di fare una netta distinzione fra i due effetti: infatti le iperpolarizzazioni prodotte dalla glicina sono completamente soppresse (v. fig. 280), mentre quelle prodotte da GABA rimangono inalterate (v. fig. 28H). Specificità, velocità e reversibilità della depressione stricninica sia del PPSI sia dell'iperpolarizzazione indotta dalla glicina (v. fig. 28I) sostengono validamente il postulato che la glicina sia il trasmettitore inibitorio, particolarmente per il fatto che nella sostanza grigia del midollo spinale, che corrisponde ovviamente alla localizzazione delle sinapsi inibitorie bloccate dalla stricnina, vi è una concentrazione relativamente alta di glicina (v. Werman e altri, 1968; v. Curtis e altri, 1968). Si postula anche che la stricnina agisca con un meccanismo di inibizione competitiva dell'azione della glicina a livello delle sinapsi inibitorie, proprio come è stato postulato per la D-tubocurarina e l'acetilcolina a livello delle sinapsi neuromuscolari (v. articolo seguente). Sperimentalmente è stato osservato che tutte le azioni inibitorie postsinaptiche nel midollo spinale possono essere riprodotte dalla glicina, anche rispetto all'antagonismo con la stricnina; è per questa ragione che si è postulato che la glicina sia il mediatore chimico dell'inibizione postsinaptica nel midollo spinale.
Anche a livelli sopraspinali si sono ottenute molte dimostrazioni sperimentali che sia la glicina sia il GABA iperpolarizzano i neuroni in un modo simile a quello delle sinapsi inibitorie. L'azione della glicina è depressa in maniera reversibile dalla stricnina proprio come nel midollo spinale, ma la stricnina non ha alcun effetto sulla trasmissione sinaptica inibitoria; si è pertanto concluso che la glicina non è il trasmettitore (v. Curtis, The pharmacology..., 1969). Recentemente la situazione è stata rivoluzionata dalla scoperta che una sostanza convulsivante, la bicucullina, non solo deprime selettivamente l'azione sinaptica inibitoria ai livelli sopraspinali, ma deprime anche selettivamente l'azione depressiva del GABA su tutti i neuroni (v. Curtis e altri, 1970). La bicucullina, pertanto, rappresenta a livello sopraspinale ciò che è la stricnina a livello spinale. Si sono saggiati molti neuroni sia spinali sia sopraspinali come mostrato nella fig. 29, A e B, dove l'azione depressiva della glicina è depressa in maniera selettiva e reversibile dalla stricnina (B) e quella del GABA dalla bicucullina (A). La bicucullina è stata saggiata in molte zone inibitorie a livello sopraspinale e ovunque deprime l'azione sinaptica inibitoria, come pure l'iperpolarizzazione e la depressione da GABA. Pertanto si può postulare che il GABA sia il trasmettitore a livello delle sinapsi inibitorie sopraspinali, come quelle illustrate nelle figg. 20, 21, 22 e 23, e anche delle sinapsi inibitorie del bialbo olfattorio (v. fig. 25).
Nella fig. 30 sono rappresentati schematicamente due neuroni, un tipico neurone spinale e un tipico neurone sopraspinale (rispettivamente A e B). Si vede che ambedue hanno zone recettrici per la glicina e il GABA; le zone della glicina sono occupate competitivamente dalla stricnina, quelle del GABA dalla bicucullina (v. anche fig. 29). La differenza è che l'azione sinaptica inibitoria si esplica sulle zone sensibili alla glicina nel neurone spinale (A) e in quelle sensibili al GABA nel neurone sopraspinale (B).
La fig. 29 mette in luce uno dei problemi più stimolanti nell'ambito della sinaptogenesi. Sembra che a tutti i livelli del sistema nervoso centrale i neuroni possiedano in relativa abbondanza zone recettrici sia per la glicina sia per il GABA. A livello spinale i neuroni inibitori producono e liberano glicina e le terminazioni assoniche trovano e formano sinapsi sulle zone appropriate sensibili alla glicina. Inversamente, a livello sopraspinale i neuroni inibitori metabolizzano GABA e formano sinapsi sulle appropriate zone sensibili al GABA.
Sembra che vi sia un'eccezione al livello spinale: l'azione inibitoria presinaptica sarebbe mediata dal GABA, perché viene bloccata dalla bicucullina mentre non è influenzata dalla stricnina (v. Curtis e altri, 1971). Questo fatto solleva il problema di come il GABA possa agire da trasmettitore causando una depolarizzazione postsinaptica a livello delle sinapsi inibitorie presinaptiche (v. fig. 26C), mentre altrove provoca una iperpolarizzazione aprendo i canali del cloro e del potassio (v. flg. 18). Il potenziale di equilibrio della depolarizzazione inibitoria presinaptica non e noto, ma probabilmente non dista più di 30 mV dal potenziale di riposo. Un'apertura piuttosto modesta dei canali del sodio causerebbe proprio un simile potenziale di equilibrio.
9. Attivazione sinaptica ripetitiva.
a) Potenziamento di frequenza delle sinapsi.
La fig. 31, A e B, ci fornisce esempi di registrazioni intracellulari di PPSE ottenute da neuroni che sono stati attivati monosinapticamente a varie frequenze. Il risultato sorprendente è che diverse sinapsi, persino sulla stessa cellula, rispondono in modo molto variabile alla stimolazione ripetitiva rapida. Per esempio, i PPSE generati dalle sinapsi nella fig. 31A aumentano in ampiezza con l'aumentare la frequenza da 1/s a 7, a 10, e cosi via fino a raddoppiare a frequenze di 100/s. Al contrario, sullo stesso neurone i PPSE generati da altre fibre presinaptiche declinano alle frequenze più alte (v. fig. 31B).
Evidentemente queste altissime frequenze possono determinare un superlavoro dei meccanismi a trasmissione chimica (come sarà discusso nell'articolo seguente, figg. 13 e 14). Per assicurare un'efficace attivazione ad alta frequenza sono necessari meccanismi assai complessi. Le vescicole sinaptiche devono migrare fino alla ‛linea di fuoco' che fronteggia lo spazio sinaptico e quindi essere svuotate e nuovamente riempite del loro contenuto. Nel frattempo esse devono essere rimpiazzate da altre vescicole, che forse devono essere fabbricate. Eppure, tutto ciò può accadere a frequenze di 100/s in modo che sia piccola o nulla la diminuzione della quantità di trasmettitore liberato a ogni singolo impulso successivo, come nella fig. 31B, o che vi sia addirittura un notevole potenziamento, come nella fig. 31A. Nella fig. 31C si vede un altro esempio di questo efficace processo per la via monosinaptica sui motoneuroni che è illustrata nella fig. 5.
Evidentemente nella fig. 31A l'intensa attività aumenta notevolmente la potenza sinaptica. In passato si riteneva che la sinapsi fosse la sede del processo di affaticamento (v. Sherrington, 1906), ma i vecchi esperimenti che sembrava dimostrassero l'affaticamento sinaptico possono ora essere interpretati sulla base di una depressione inibitoria dell'azione sinaptica che non era stata riconosciuta come tale. Non è facile determinare un affaticamenio della sinapsi inibitoria, che può perfino potenziare la sua efficacia di trasmissione in seguito a intensa attivazione, come nella fig. 31A.
La fig. 31D mostra che le sinapsi formate dai neuroni cerebrali sono potenziate dalla stimolazione ad alta frequenza in modo molto più intenso delle sinapsi spinali (v. Landgren e altri, 1962). Le registrazioni intracellulari della fig. 31, C e D, sono state ottenute dallo stesso motoneurone del midollo spinale cervicale del babbuino. Nella serie inferiore della fig. 31D la corteccia motoria è stimolata a 200/s e i potenziali postsinaptici eccitatori (PPSE) sono generati per via monosinaptica da scariche d'impulsi discendenti attraverso il tratto piramidale. Nella serie della fig. 31C lo stesso motoneurone è eccitato monosinapticamente da sinapsi che provengono dai recettori da stiramento periferici del muscolo (v. figg. lA e 5). Ambedue gli inputs monosinaptici a questo motoneurone sono segnalati dalle registrazioni extracellulari al di sopra dei PPSE intracellulari e mostrano che praticamente le scariche afferenti sono rimaste costanti. I PPSE successivi prodotti dalle fibre afferenti muscolari (v. fig. 31C) assomigliano a quelli della fig. 31B e rispondono a frequenze di 200/s senza che si noti un apprezzabile cambiamento della loro ampiezza. Ma i potenziali eccitatori monosinaptici evocati dalla stimolazione piramidale mostrano un comportamento del tutto diverso (v. fig. 31D). Il primo potenziale ha all'incirca la stessa ampiezza di quello evocato dal nervo afferente muscolare, ma nelle risposte successive aumenta enormemente, di un fattore 10, e tende a stabilizzarsi a questo livello grandemente potenziato. Ciò mostra che le sinapsi formate dalle cellule del tratto piramidale possiedono un meccanismo straordinariamente efficiente per aumentare la loro efficacia quando sono sottoposte a un superlavoro causato dal potenziamento ad alta frequenza. La fig. 31E mostra un potenziamento di frequenza molto efficace ottenuto in un neurone della corteccia visiva. I PPSE evocati per via monosinaptica sono potenziati fino al doppio del loro valore di controllo a 60/s (osservazioni inedite di Toyama; sul potenziamento di frequenza delle sinapsi v. anche Eccles, Presynaptic..., 1964).
b) Potenziamento post-tetanico delle sinapsi.
Oltre a questo potenziamento di frequenza che si osserva durante la stimolazione ripetitiva, vi è un altro processo di potenziamento che segue l'attivazione ad alta frequenza delle sinapsi e continua spesso per lungo tempo. Fin dalla scoperta di questo fenomeno di potenziamento post-tetanico (v. Lloyd, 1949) si è ritenuto che esso potesse offrire un esempio di processo di apprendimento nelle sinapsi (v. Eccles e McIntyre, 1953; v. Curtis ed Eccles, 1960; v. Kandel e Spencer, 1968). Nella fig. 32, A e B, è mostrata una stimolazione ripetitiva delle afferenze monosinaptiche al motoneurone in esame. Le aree tratteggiate indicano la durata di questa stimolazione a 640/s (5 secondi in A e 30 secondi in B). I tracciati dei PPSE degli inserti fanno vedere che dopo il tetano vi è stato un grande potenziamento il cui decorso - aumento e declino - è mostrato dai punti del diagramma; il decorso temporale è decisamente più lento con un tetano più lungo. Tuttavia, entro 4 minuti nel caso del tetano più breve ed entro 6 minuti in quello del più lungo, il potenziamento è scomparso completamente. Nell'esperimento illustrato nella fig. 32C (v. Spencer e Wigdor, 1965) si è avuta una tetanizzazione più lunga (500/s per 20 mm) e il potenziamento ha avuto un declino estremamente lento, tanto che era possibile riscontrarlo anche dopo due ore; comunque, anche in questo caso, alla fine scomparve.
Si potrebbe concludere che in queste sinapsi che sono responsabili dell'attivazione monosinaptica di motoneuroni non è possibile dimostrare alcun aumento di funzione indefinitamente prolungato conseguente a un eccesso di uso e che pertanto le nostre ricerche non hanno fornito informazioni significative sull'ipotesi che l'apprendimento sia dovuto a un eccesso di attivazione sinaptica e al conseguente aumento dell'efficacia sinaptica.
D'altra parte, secondo alcune osservazioni effettuate da Bliss e Lømo, le sinapsi della via entorinale sui granuli della fascia dentata nell'ippocampo sono fortemente potenziate da una tetanizzazione condizionante e tale potenziamento può essere sommato e quindi rimane per molte ore in forma completamente sviluppata (v. fig. 33). Di fatto non si è notato alcun declino durante l'intero periodo dell'esperimento. Le prime cinque frecce nella fig. 33 segnalano brevi tetani condizionanti (20/s per 15s), ciascuno dei quali è seguito da un potenziamento che cresce a gradini fino all'enorme potenziamento dopo l'ultimo tetano. I cerchi bianchi mostrano le risposte di controllo dalla parte non condizionata, che non mostra potenziamento. Tuttavia, questa parte è stata stimolata in corrispondenza delle ultime quattro frecce della serie, inducendo un potenziamento piccolo e prolungato. Questo esperimento di Bliss e Lømo è senz'altro molto incoraggiante per quanti sostengono la teoria dell'uso sinaptico' dell'apprendimento, perché esso mostra che in una parte del sistema nervoso centrale, che si ritiene avere un ruolo importante nei processi della memoria, vi sono sinapsi che possiedono un meccanismo altamente sviluppato per un potenziamento di durata estremamente lunga conseguente a un'eccessiva stimolazione. Questo esperimento, inoltre, evidenzia la necessità di realizzare un numero molto maggiore di esperimenti ben ideati, a livelli più alti del sistema nervoso.
Bibliografia.
Akert, K., Moor, H., Pfenninger, K., Sandri, C., Contributions of new impregnation methods and freeze-etching to the problems of synaptic fine structure, in Progress in brain research, vol. XXXI (a cura di K. Akert e P. G. Waser), Amsterdam 1969, pp. 223-240.
Akert, K., Sandri, C., Identification of the active synaptic region by means of histochemical and freeze-etching techniques, in Excitatory synaptic mechanisms (a cura di P. Andersen e J. K. S. Jansen), Oslo 1969, pp. 27-41.
Allen, G. I., Eccles, J. C., Nicoll, R. A., Oshima, T., Rubia, F. J., The ionic mechanisms concerned in generating the IPSPs of hippocampal pyramidal cells, in ‟Proceedings of the Royal Society" (Series B), 1977, CXCVIII, pp. 363-384.
Andersen, P., Eccles, J. C., Loyning, Y., Location of postsynaptic inhibitory synapses on hippocampal pyramids, in ‟Journal of neurophysiology", 1964, XXVII, pp. 592-607.
Andersen, P., Eccles, J. C., Loyning, Y., Pathway of postsynaptic inhibition in the hippocampus, in ‟Journal of neurophysiology", 1964, XXVII, pp. 608-619.
Andersen, P., Holmqvist, B., Voorhoeve, P. E., Excitatory synapses on hippocampal apical dendrites activated by entorhinal stimulation, in ‟Acta physiologica scandinavica", 1966, LXVI, pp. 461-472.
Araki, T., Ito, M., Oscarsson, O., Anion permeability of the synaptic and non synaptic motoneurone membrane, in ‟Journal of physiology", 1961, CLIX, pp. 410-435.
Blankenship, J. E., Kuno, M., Analysis of spontaneous subthreshold activity in spinal motoneurons of the cat, in ‟Journal of neurophysiology", 1968, XXXI, pp. 195-209.
Bodian, D., Electron microscopy: two major synaptic types on spinal motoneurons, in ‟Science", 1966, CLI, pp. 1093-1094.
Brock, L. G., Coombs, J. S., Eccles, J. C., The recording of potentials from motoneurones with an intracellular electrode, in ‟Journal of physiology", 1952, CXVII, pp. 431-460.
Burke, R. E., Composite nature of the monosynaptic excitatory postsynaptic potential, in ‟Journal of neurophysiology", 1967, XXX, pp. 1114-1137.
Coombs, J. S., Curtis, D. R., Eccles, J. C., The generation of impulses in motoneurones, in ‟Journal of physiology", 1957, CXXXIX, pp. 232-249.
Coombs, J. S., Eccles, J. C., Fatt, P., The specific ionic conductances and the ionic movements across the motoneuronal membrane that produce the inhibitory post-synaptic potential, in ‟Journal of physiology", 1955, CXXX, pp. 326-373.
Coombs, J. S., Eccles, J. C., Fatt, P., Excitatory synaptic action in motoneurones, in ‟Journal of physiology", 1955, CXXX, pp. 374-395.
Creed, R. S., Denny-Brown, D., Eccles, J. C., Liddel, E. G. T., Sherrington, Ch. S., Reflex activity in the spinal cord, London 1932.
Curtis, D. R., Central synaptic transmitters, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper, A.A. Ward e A. Pope), Boston 1969, pp. 105-129.
Curtis, D. R., The pharmacology of spinal postsynaptic inhibition, in Progress in brain research, vol. XXXI (a cura di K. Akert e P.G. Waser), Amsterdam 1969, pp. 171-189.
Curtis, D. R., Duggan, A.W., Felix, D., Johnston, G. A. R., GABA, bicuculline and central inhibition, in ‟Nature", 1970, CCXXVI, pp. 1222-1224.
Curtis, D. R., Duggan, A. W., Johnston, G. A. R., The specificity of strychnine as a glycine antagonist in the mammalian spinal cord, in ‟Experimental brain research", 1971, XII, pp. 547-565.
Curtis, D. R., Eccles, J. C., The time courses of excitatory and inhibitory synaptic actions, in ‟Journal of physiology", 1959, CXLV, pp. 529-546.
Curtis, D. R., Eccles, J. C., Synaptic action during and after repetitive stimulation, in ‟Journal of physiology", 1960, CL, pp. 374-398.
Curtis, D. R., Hösli, L., Johnston, G. A. R., A pharmacological study of the depression of spinal neurones by glycine and related amino acids, in ‟Experimental brain research", 1968, VI, pp. 1-18.
Dowling, J. E., Boycott, B. B., Neural connections of the retina: fine structure of the inner plexiform layer, in ‟Cold Spring Harbor symposia on quantitative biology", 1966, XXX, pp. 393-402.
Eccles, J. C., The physiology of nerve cells, Baltimore 1957.
Eccles, J. C., Excitatory and inhibitory synaptic action, in ‟Annals of the New York Academy of Sciences", 1959, LXXXI, pp. 247-264.
Eccles, J.C., Ionic mechanism of postsynaptic inhibition, in Les prix Nobel 1963, Stockholm 1964, pp. 261-283.
Eccles, J. C., Presynaptic inhibition in the spinal cord, in Progress in brain research, vol. XII (a cura di J. C. Eccles e J. P. Schadé), Amsterdam 1964, pp. 65-91.
Eccles, J. C., The physiology of synapses, Berlin 1964.
Eccles, J. C., The inhibitory pathways of the central nervous system. Sherrington lecture, Liverpool 1969.
Eccles, J. C., The understanding of the brain, New York 19772.
Eccles, J. C., Eccles, R. M., Iggo, A., Lundberg, A., Electrophysiological investigations on Renshaw cells, in ‟Journal of physiology", 1961, CLIX, pp. 461-478.
Eccles, J. C., Eccles, R. M., Lundberg, A., Synaptic actions on motoneurones in relation to the two components of the group I muscle afferent volley, in ‟Journal of physiology", 1957, CXXXVI, pp. 527-546.
Eccles, J. C., Fatt, P., Koketsu, K., Cholinergic and inhibitory synapses in a pathway from motor-axon collaterals to motoneurones, in ‟Journal of physiology", 1954, CXXVI, pp. 524-562.
Eccles, J. C., Hubbard, J. I., Oscarssson, O., Intracellular recording from cells of the ventral spino-cerebellar tract, in ‟Journal of physiology", 1961, CLVIII, pp. 486-516.
Eccles, J. C., Ito, M., Szentágothai, J., The cerebellum as a neuronal machine, Berlin 1967.
Eccles, J. C., Llinás, R., Sasaki, K., Intracellularly recorded responses of the cerebellar Purkinje cells, in ‟Experimental brain research", 1966, I, pp. 161-183.
Eccles, J. C., Llinás, R., Sasaki, K., The excitatory synaptic action of climbing fibres on the Purkinje cells of the cerebellum, in ‟Journal of physiology", 1966, CLXXXII, pp. 268-296.
Eccles, J. C., McIntyre, A. K., The effects of disuse and of activity on mammalian spinal reflexes, in ‟Journal of physiology", 1953, CCXXI, pp. 492-516.
Eccles, J. C., Nicoll, R. ., Oshima, T., Rubia, F. J., The anionic permeability of the inhibitory postsynaptic membrane of hippocampal pyramidal cells, in ‟Proceedings of the Royal Society" (Series B), 1977, CXCVIII, pp. 345-361.
Eide, E., Jurna, I., Lundberg, A., Conductance measurements from motoneurons during presynaptic inhibition, in Structure and function of inhibitory neuronal mechanisms (a cura di C. von Euler, S. Skoglund e U. Söderberg), Oxford 1968, pp. 215-219.
Frank, K., Fuortes, M. G. F., Presynaptic and postsynaptic inhibition of monosynaptic reflexes, in ‟Federation proceedings", 1957, XVI, pp. 39-40.
Gallagher, J. F., Higashi, H., Nishi, S., The ionic requirements for the production of the GABA depolarization at cat primary afferent neurones, in ‟Federation proceedings", 1975, XXXIV, p. 418.
Gerschenfeld, H. M., Paupardin-Tritsch, D., Ionic mechanisms and receptor properties underlying the responses of molluscan neurones to 5-hydroxytryptamine, in ‟Journal of physiology", 1974, CCXLIII, pp. 427-456.
Granit, R., Kernell, D., Shortess, G. K., Quantitative aspects of repetitive firing of mammalian motoneurones, as caused by injected currents, in ‟Journal of physiology", 1963, CLXVIII, pp. 911-931.
Gray, E. G., Problems of interpreting the fine structure of Vertebrate and Invertebrate synapses, in ‟International review of general and experimental zoology", 1966, II, pp. 139-170.
Gray, E. G., Electron microscopy of excitatory and ihibitory synapses: a brief review, in Progress in brain research, vol. XXXI (a cura di K. Akert e P.G. Waser), Amsterdam 1969, pp. 141-155.
Gustaffson, D., Afterhyperpolarization and the control of repetitive firing in spinal neurones of the cat, in ‟Acta physiologica scandinavica", 1974, CCCCXVI, suppl., pp. 1-47.
Hamlyn, L. H., An electron microscope study of pyramidal neurons in the Ammon's Horn of the rabbit, in ‟Journal of anatomy", 1962, XCVII, pp. 189-201.
Hamori, J., Pasik, P., Pasik, T., Szentágothai, J., ‛Triadic' synaptic arrangements and their possible significance in the lateral geniculate nucleus of the monkey, in ‟Brain research", 1974, LXXX, pp. 379-393.
Ito, M., Kostyuk, P. G., Oshima, T., Further study on anion permeability of inhibitory post-synaptic membrane of cat motoneurones, in ‟Journal of physiology", 1962, CLXIV, pp. 150-156.
Ito, M., Yoshida, M., The origin of cerebellar-induced inhibition of Deiters neurones. I. Monosynaptic initiation of the inhibitory postsynaptic potentials, in ‟Experimental brain research", 1966, II, pp. 330-349.
Jones, E. G., Powell, T. P. S., Electron microscopy of synaptic glomeruli in the thalamic relay nuclei of the cat, in ‟Proceedings of the Royal Society" (Series B), 1969, CLXXII, pp. 153-171.
Kandel, E. R., Spencer, W. A., Cellular neurophysiological approaches in the study of learning, in ‟Physiological reviews", 1968, XLVIII, pp. 65-134.
Katz, B., Miledi, R., A study of spontaneous miniature potentials in spinal motoneurones, in ‟Journal of physiology", 1963, CLXVIII, pp. 389-422.
Kehoe, J., Ionic mechanisms of a two-component cholinergic inhibition in Aplysia neurones, in ‟Journal of physiology", 1972, CCXXV, pp. 85-114.
Koike, H., Okada, Y., Oshima, T., Takahashi, K., Accommodative behavior of cat pyramidal tract cells investigated with intracellular injection of currents, in ‟Experimental brain research", 1968, V, pp. 173-188.
Kuno, M., Quantal components of excitatory synaptic potentials in spinal motoneurones, in ‟Journal of physiology", 1964, CLXXV, pp. 81-99.
Kuno, M., Transmitter action and multiple discharge, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper, A. A. Ward e A. Pope), Boston 1969, pp. 130-135.
Landgren, S., Phillips, C. G., Porter, R., Cortical fields of origin of the monosynaptic pyramidal pathways to some alpha motoneurones of the baboon's hand and forearm, in ‟Journal of physiology", 1962, CLXI, pp. 112-125.
Lloyd, D. P. C., Post-tetanic potentiation of response in monosynaptic reflex pathways of the spinal cord, in ‟Journal of general physiology", 1949, XXXIII, pp. 147-170.
Lund, R. D., Synaptic patterns of the superficial layers of the superior colliculus of the rat, in ‟Journal of comparative neurology", 1969, CXXXV, pp. 179-208.
Lux, H. D., Ammonium and chloride extrusion: hyperpolarizing synaptic inhibition in spinal motoneurones, in ‟Science", 1971, CLXXIII, pp. 555-/57.
Lux, H. D., Loracher, C., Neher, E., Action of ammonium on post-synaptic inhibition of cat spinal motoneurones, in ‟Experimental brain research", 1970, XI, pp. 431-447.
Mendell, L. M., Henneman, E., Terminals of single Ia fibers: location, density and distribution within a pool of 300 homonymous motoneurons, in ‟Journal of neurophysiology", 1971, XXXIV, pp. 171-187.
Morest, D. K., Dendro-dendritic synapses of cells that have axons: the fine structure of the Golgi type II cell int he medial geniculate body of the cat, in ‟Anatomische und entwicklungsgeschichtliche Monographien", 1971, CXXXIII, pp. 216-246.
Nicoll, R.A., Inhibitory mechanisms in the rabbit olfactory bulb: dendrodendritic mechanisms, in ‟Brain research", 1969, XIV, pp. 157-172.
Nicoll, R. A., The action of presumed blockers of chloride transport on synaptic and amino acid responses in the frog spinal cord, in ‟Neuroscience abstracts", 1975, I, p. 377.
Nicoll, R. A., Are dorsal root potentials generated by accumulation of extracellular potassium?, in ‟Neuroscience abstracts", 1976, II, p. 980.
Nicoll, R. A., The blockade of GABA mediated responses in the frog spinal cord by ammonium ions and furosemide, in ‟Journal of physiology", 1978, CCLXXXIII, pp. 121-132.
Nishi, S., Minota, S., Karczmar, A. G., Primary afferent neurones: the ionic mechanisms of GABA mediated depolarization, in ‟Neuropharmacology", 1974, XIII, pp. 215-219.
Oshima, T., Studies of pyramidal tract cells, in Basic mechanisms of the epilepsies (a cura di H. H. Jasper, A. A. Ward e A. Pope), Boston 1969, pp. 253-261.
Rall, W., Shepherd, G. M., Reese, T. S., Brightman, M. W., Dendrodendritic synaptic pathway for inhibition in the olfactory bulb, in ‟Experimental neurology", 1966, XIV, pp. 44-56.
Ralston, H. J., Herman, M. M., The fine structure of neurones and synapses in the ventrobsal thalamus of the cat, in ‟Brain research", 1969, XIV, pp. 77-97.
Ramón y Cajal, S., Histologie du système nerveux de l'homme et des Vertébrés, vol. I, Paris 1909, vol. II, Paris 1911.
Schmidt, R. F., Presynaptic inhibition in the Vertebrate central nervous system, in ‟Ergebnisse der Physiologie", 1971, LXIII, pp. 21-108.
Shepherd, G. M., The synaptic organization of the brain, New York 1974.
Sherrington, Ch. S., The integrative action of the nervous system, New Haven 1906.
Sherrington, Ch. S., Some functional problems attaching to convergence, in ‟Proceedings of the Royal Society" (Series B), 1929, CV, pp. 332-362.
Sloper, J. J., Dendro-dendritic synapses in the Primate motor cortex, in ‟Brain research", 1971, XXXIV, pp. 186-192.
Spencer, W. A., Wigdor, R., Ultra-late PTP of monosynaptic reflex responses in cat, in ‟Physiologist", 1965, VIII, p. 278.
Tsukahara, N., Kosaka, K., The mode of cerebral excitation of red nucleus neurones, in ‟Experimental brain research", 1968, V, pp. 102-117.
Uchizono, K., Synaptic organization of the Purkinje cells in the cerebellum of the cat, in ‟Experimental brain research", 1967, IV, pp. 97-113.
Werman, R., Davidoff, R. A., Aprison, M. H., Inhibitory action of glycine on spinal neurons in the cat, in ‟Journal of neurophysiology", 1968, XXXI, pp. 81-95.
Whittaker, V. P., Gray, E. G., The synapse: biology and morphology, in ‟British medical bulletin", 1962, XVIII, pp. 223-228.
Wilson, V. J., Burgess, P. R., Disinhibition in the cat spinal cord, in ‟Journal of neurophysiology", 1962, XXV, pp. 392-404.
Wilson, V. J., Yoshida, M., Schor, R. H., Supraspinal monosynaptic excitation and inhibition of thoracic back motoneurons, in ‟Experimental brain research", 1970, XI, pp. 282-295.
Fisiologia della sinapsi periferica
SOMMARIO: 1. Trasmissione neuromuscolare: a) introduzione storica; b) struttura della sinapsi neuromuscolare; c) l'azione del trasmettitore neuromuscolare, acetilcolina; d) proprietà farmacologiche della sinapsi neuromuscolare; e) natura quantica dell'azione del trasmettitore neuromuscolare; f) fattori che controllano l'emissione di quanti dalla terminazione nervosa; g) ruolo essenziale del calcio nella liberazione dei quanti; h) quadro dei probabili eventi che hanno luogo nella trasmissione neuromuscolare; i) azione molecolare dell'acetilcolina (A Ch); l) eventi postsinaptici nella trasmissione neuromuscolare; m) attivazione sinaptica ripetitiva; n) fattori che causano l'interruzione della trasmissione neuromuscolare. □ 2. La sinapsi gigante del ganglio stellato del calamaro: a) generalità; b) generazione di potenziali postsinaptici; c) meccanismi ionici di generazione di potenziali postsinaptici. □ 3. I canali ionici attraverso le membrane presinaptica e postsinaptica di una sinapsi a trasmissione chimica. □ 4. Altre sinapsi periferiche a trasmissione chimica. □ Bibliografia.
1. Trasmissione neuromuscolare.
a) Introduzione storica.
Sappiamo (v. elettrofisiologia) che la trasmissione lungo una fibra nervosa avviene tramite brevi impulsi ‛tutto-o-nulla' e che tale propagazione di impulsi dipende da due fattori principali: anzitutto dalle proprietà della fibra nervosa, che, al pari di un cavo, ha un nucleo conduttore avvolto da una membrana relativamente resistente; in secondo luogo dal grande e momentaneo aumento della conduttanza al sodio di questa membrana quando il suo potenziale di riposo viene improvvisamente diminuito. Di conseguenza la debolissima trasmissione lungo la fibra nervosa, basata sulle proprietà di cavo, è amplificata a ogni segmento con un meccanismo autorigenerativo. Il carattere tutto-o-nulla della propagazione è dovuto all'amplificazione che ogni segmento del nervo dà al segnale attenuato, trasmesso dalla regione attiva adiacente, come avviene in un cavo.
In contrasto con questa propagazione di impulsi lungo la fibra nervosa, che avviene senza attenuazione, era da lungo tempo sorta la convinzione che qualche speciale meccanismo fosse responsabile della trasmissione da un elemento conduttore a quello successivo. Il concetto di neurone venne sviluppato dai neuroanatomisti W. His, A. Forel e S. Ramón y Cajal, nell'ultima parte del XIX secolo. Partendo da tale scoperta Ch. S. Sherrington giunse al concetto di sinapsi, cioè di strutture di trasmissione specializzate, tramite le quali un neurone ne influenza un altro (v. Eccles, 1964, cap. 1). Molto tempo prima Cl. Bernard aveva messo in evidenza che la regione di contatto tra nervo e muscolo era bloccata dal curaro, ed E. Du Bois Reymond aveva in effetti ipotizzato che la trasmissione neuromuscolare fosse in parte mediata chimicamente. Ma tra la fine del secolo scorso e l'inizio di quello attuale il concetto di ‛messaggero chimico' responsabile della cosiddetta trasmissione umorale, ovvero di ormone come sostanza specifica che viene trasportata dal sangue e che agisce lentamente per periodi prolungati, era solo ai primordi.
Frattanto, nelle prime tre decadi di questo secolo era stato dimostrato che la trasmissione attraverso la sinapsi a livello della giunzione neuromuscolare era molto veloce, con un ritardo al massimo di 1 ms tra l'impulso nervoso e quello muscolare. Questo contrasto tra la lentezza della trasmissione chimica mediata dagli ormoni e la trasmissione estremamente rapida e breve a livello delle giunzioni neuromuscolari e delle sinapsi, ha fatto ritenere che la trasmissione chimica fosse di gran lunga troppo lenta. Sorse pertanto l'idea che la trasmissione rapida fosse mediata elettricamente, cioè che le correnti elettriche responsabili della trasmissione lungo la fibra nervosa fossero responsabili anche dell'eccitazione della cellula nervosa o della fibra muscolare attraverso la giunzione sinaptica. Questa ipotesi suscitò grandi perplessità a causa delle notevoli diversità delle proprietà elettriche esistenti tra la fibra nervosa e quella muscolare. Tuttavia a ciò non fu data molta importanza, a causa delle scarse conoscenze sui rispettivi parametri elettrici.
Nel frattempo O. Loewi (v., 1921 e 1933) fece una scoperta, poi sviluppata da lui stesso e dai suoi colleghi negli anni venti. Per la prima volta fu dimostrato sperimentalmente che impulsi nervosi operano nella sinapsi mediante trasmissione chimica, anche se si tratta di sinapsi molto specializzate tra nervi e muscolo cardiaco. Era da tempo noto che, nei Vertebrati, gli impulsi nel nervo vago inibiscono il cuore. Quando Loewi inibì in tal modo il cuore di rana, osservò che il liquido di perfusione proveniente da questo cuore aveva un'azione inibitrice analoga, in quanto rallentava l'attività di un altro cuore di rana. Loewi postulò che la stimolazione del vago avesse agito tramite la mediazione di una sostanza inibitrice (Vagusstoff) liberata dalle terminazioni del nervo, la cui diffusione esercitava un'azione inibitrice sulle fibre del muscolo cardiaco. Per la prima volta si ebbe la prova di una trasmissione chimica attraverso una sinapsi periferica. La trasmissione chimica fu dimostrata in maniera simile da Loewi anche per l'azione di accelerazione sul cuore di rana da parte di impulsi del nervo simpatico (v. farmacologia).
Fu a questo punto che vennero collegati alla teoria della trasmissione chimica verso il cuore precedenti suggerimenti di H. H. Dale e T. R. Elliott e cioè che l'acetilcolina e l'adrenalina fossero sostanze molto potenti capaci di riprodurre rispettivamente l'azione vagale inibitoria e quella simpatica eccitatoria sul cuore. Loewi dimostrò che l'atropina bloccava l'azione inibitoria non solo del vago e dell'acetilcolina, ma anche quella del Vagusstoff e che tutt'e tre queste azioni erano similmente potenziate dall'eserina, inibitore della colinesterasi, enzima che demolisce specificamente per idrolisi l'acetilcolina (v. Eccles, 1964, cap. 1).
Queste trasmissioni chimiche erano molto lente, con tempi di azione misurati in decimi di secondo; pertanto, l'ipotesi di Dale, secondo la quale la velocissima trasmissione neuromuscolare era mediata da acetilcolina, fu per molti anni contrastata. Le ricerche pionieristiche di Dale (v., 1937 e 1938) e dei suoi collaboratori furono confermate e approfondite da Kuffier (v., 1948 e 1949) e particolarmente da Katz (v., 1962, 1966 e 1969) e dai suoi collaboratori negli anni cinquanta e sessanta. Si può oggi asserire che le ricerche sulla trasmissione neuromuscolare sono tra le più interessanti e raffinate nel campo della neurobiologia. Come si vedrà, le osservazioni al microscopio elettronico sulla sinapsi neuromuscolare corrispondono perfettamente a numerose scoperte biofisiche. Possiamo affermare che la trasmissione chimica è un meccanismo fisiologico più complesso di quanto si era creduto negli anni trenta.
L'ipotesi chimica fu avanzata originariamente da Dale e dai suoi collaboratori sulla base di esperimenti che mostravano che, stimolando le fibre di un nervo motore, venivano liberate nel liquido di perfusione del muscolo piccole quantità di acetilcolina (ACh), e che l'iniezione intrarteriosa di quantità molto piccole di ACh evocava impulsi nelle fibre muscolari, causando così la loro contrazione. Fu inoltre messo in evidenza che sostanze curarosimili bloccavano la trasmissione neuromuscolare non deprimendo la liberazione di ACh, ma la risposta del muscolo conseguente alla iniezione di ACh. L'azione di sostanze inibitrici della colinesterasi fornì un'ulteriore prova a sostegno di questa teoria: infatti si poteva prevedere che tali sostanze avrebbero prolungato e intensificato l'azione dell'ACh e in effetti ciò fu osservato. L'effetto eccitatorio di una singola scarica nervosa veniva prolungato fino a produrre un breve tetano decrescente del muscolo, mentre nei muscoli curarizzati si restaurava la trasmissione neuromuscolare. I dettagli di questo lavoro furono discussi in rassegne di quell'epoca (v. Eccles, 1936 e 1937; v. Brown, 1937; v. Dale, 1937 e 1938).
b) Struttura della sinapsi neuromuscolare.
La fig. lA mostra schematicamente un motoneurone del midollo spinale con il suo assone diretto in basso verso le fibre muscolari che esso innerva. I muscoli di un arto del gatto, per esempio, sono innervati da alcune centinaia di motoneuroni tramite le loro grandi fibre motrici (v. fig. 1B). Di solito l'assone motore si dirama innervando in un determinato muscolo alcune centinaia di fibre muscolari, non solo le 5 mostrate nella figura, e di norma una fibra muscolare riceve una sola terminazione nervosa. Così un motoneurone contribuisce al movimento scaricando impulsi lungo il suo assone e facendo quindi contrarre il suo gruppo di fibre muscolari. L'insieme del motoneurone con le sue fibre muscolari è chiamato unità motrice. Questa è una vera componente unitaria di movimento, poiché di solito il potenziale d'azione che percorre il motoneurone genera un potenziale d'azione con conseguente contrazione in tutte le fibre muscolari innervate. La fig. 1C mostra le tensioni della contrazione isometrica prodotte dalla tetanizzazione di un'unità motrice per un'ampia gamma di frequenze. Nel gatto la tensione di contrazione prodotta dalla rapida attivazione ripetitiva di una unità motrice varia da 50 a 100 grammi-peso nei grandi muscoli estensori, e da 10 a 20 nei muscoli più piccoli (v. Devanandan e altri, 1965).
La microscopia ottica ha rivelato in qual modo la fibra nervosa termina, cioè mediante una struttura caratteristicamente ramificata, in stretta apposizione con la fibra muscolare, che forma la cosiddetta placca motrice (v. fig. 2A). Tuttavia, il microscopio elettronico, con il suo più alto potere risolutivo, è essenziale per mettere in evidenza molti dei caratteri strutturali utili a spiegare i risultati di elaborate ricerche di fisiologia. Spesso i fisiologi sono costretti a postulare strutture che potrebbero essere rivelate solo se il potere di risoluzione della microscopia elettronica venisse migliorato di un altro ordine di grandezza (v. figg. 10A e 21).
La fig. 2B è una microfotografia che mostra una piccola sezione della regione di congiunzione tra la terminazione nervosa e la fibra muscolare. La parte terminale del nervo è piena di numerose vescicole sferiche di circa 500 Å di diametro; queste tendono a raccogliersi in gruppi sulla superficie della terminazione nervosa che si trova di fronte alla fibra muscolare al di là della fessura sinaptica, indicata dalle frecce su ambedue i lati. La fessura con la sua fascia centrale scura è larga circa 400 Å ed è la sede della trasmissione chimica. I due caratteri strutturali, vescicole (vescicole sinaptiche) e fessura sinaptica, sono componenti essenziali di ogni sinapsi a trasmissione chimica. In sostanza la trasmissione chimica implica vari processi (v. fig. 10, A e B): a) la sintesi della sostanza trasmettitrice (in questo caso acetilcolina) e il suo impacchettamento nelle vescicole sinaptiche; b) l'azione dell'impulso nervoso che determina la liberazione del contenuto di alcune vescicole nella fessura sinaptica; c) la diffusione attraverso la fessura della sostanza trasmettitrice che si lega in punti recettori specifici sulla membrana postsinaptica; d) l'apertura momentanea di vie ioniche specifiche attraverso la membrana postsinaptica. Vi sono molti altri processi, alcuni dei quali saranno descritti più avanti.
c) L'azione del trasmettitore neuromuscolare, acetilcolina.
La trasmissione normale di un impulso nervoso in una fibra muscolare è illustrata dai tre tracciati della fig. 3B (v. Fatt e Katz, 1951), mentre nella fig. 3A è stata disegnata schematicamente la situazione sperimentale. Il nervo viene stimolato e si registra l'effetto sulla fibra muscolare mediante un elettrodo intracellulare, posto dapprima in corrispondenza della placca motrice e quindi a 1 mm e 2 mm di distanza. Il progressivo ritardo del potenziale d'azione muscolare è dovuto al tempo di conduzione lungo la fibra muscolare. Il potenziale d'azione registrato a 2 mm ha una configurazione tipica, ma presso la placca motrice mostra un'inflessione iniziale in fase ascendente, la sommità è ridotta in altezza e mostra una nuova inflessione in fase di declino. Tracce di questa distorsione possono essere rilevate nella registrazione ottenuta a 1 mm.
Questa distorsione osservabile in prossimità della placca motrice può essere spiegata dall'analisi dei potenziali registrati in modo simile, quando si blocca con curaro la trasmissione neuromuscolare (v. fig. 3C). La diminuzione in ampiezza e il rallentamento del decorso temporale, particolarmente nella fase ascendente, sono spiegati in maniera soddisfacente dal fatto che la depolarizzazione della membrana muscolare è indotta nella regione della placca motrice e di li si propaga lungo la fibra muscolare come lungo un cavo. Il trasmettitore chimico deve agire sulla membrana nella regione della placca motrice della fibra muscolare e determinare un potente flusso di corrente verso l'interno (depolarizzante). La depolarizzazione così prodotta (il potenziale di placca, PP) produce sia l'inflessione iniziale sia quella più tardiva sul potenziale d'azione registrato in corrispondenza della placca motrice nella fig. 3B.
La riduzione in ampiezza del potenziale d'azione è dovuta al fatto che la corrente depolarizzante nella placca motrice attivata è alimentata da una batteria di membrana che è solo di circa 80 mV nella fig. 3B, e pertanto si oppone alla più grande depolarizzazione - in eccesso di 120 mV - della sommità del potenziale d'azione.
La graduale azione depressiva del curaro sulla trasmissione neuromuscolare è bene illustrata dalla serie della fig. 3E, una registrazione effettuata da Kuffler (v., 1942) da una singola giunzione neuromuscolare, isolata per dissezione (v. fig. 3D), un decennio prima dell'epoca delle registrazioni intracellulari dal muscolo. Con tale tecnica extracellulare si registrano le correnti elettriche attraverso la membrana muscolare. Con un'accurata localizzazione dell'elettrodo registrante in corrispondenza della placca motrice (v. fig. 3D) la corrente di placca costituisce la principale componente della fase ascendente, mentre la corrente generata dall'impulso muscolare inizia in corrispondenza dell'inflessione verso l'alto, presso la sommità. Via via che la curarizzazione diviene più efficace si riducono sia la pendenza sia l'ampiezza della corrente di placca. Come conseguenza l'impulso muscolare inizia sempre più tardi e infine viene meno in una maniera tutto-o-nulla, lasciando come residuo la corrente di placca.
Come vedremo più avanti, il curaro agisce impedendo all'acetilcolina di depolarizzare efficacemente il muscolo in corrispondenza della placca motrice. Se si elimina il potenziale d'azione muscolare rimane il potenziale di placca (PP), come mostra il tracciato più in basso della fig. 3E. Nella fig. 3C si vedono alcuni tracciati sovrapposti di potenziali di placca, registrati per via intracellulare come nella fig. 3A, in corrispondenza della placca motrice (0) e a distanze di 1, 2, 3 e 4 mm da essa. Il potenziale di placca decresce progressivamente con la distanza e presenta un decorso temporale più lento. Esso si propaga lungo la fibra muscolare come lungo un cavo dimezzandosi all'incirca ogni millimetro, e pertanto si riduce moltissimo passando da o a 1, 2, 3 e 4 mm. Le ‛proprietà di cavo' della fibra muscolare sono molto peggiori di quelle della fibra nervosa.
d) Proprietà farmacologiche della sinapsi neuromuscolare.
Abbiamo già parlato di come si è arrivati a stabilire che il trasmettitore chimico della sinapsi neuromuscolare è l'acetilcolina (ACh). Il lavoro compiuto da Dale e dai suoi collaboratori può sembrare un po' rozzo, se giudicato con i mezzi che abbiamo oggi a disposizione, tuttavia, a quell'epoca, fu una scoperta straordinaria.
Un notevole miglioramento fu determinato dalla tecnica di iniettare acetilcolina elettroforeticamente attraverso una sottile micropipetta che poteva essere posta molto vicino a una placca motrice. La registrazione veniva effettuata tramite un elettrodo intracellulare in un preparato curarizzato, in modo da registrare solamente il potenziale di placca, in modo simile a quello della fig. 3C, al punto 0. Krnjević e Miledi (v., 1958) hanno dimostrato che l'applicazione elettroforetica di 1,5 × 10-14 g di ACh, iniettata molto vicino alla placca motrice, riproduceva un potenziale molto simile a quello prodotto da un impulso nervoso; si calcola infatti che quest'ultimo liberi 1,5 × 10-15 g di ACh in corrispondenza della placca. La discordanza di un fattore 10 è buona se si pensa che l'iniezione può essere fatta solo in corrispondenza di un lato della fessura sinaptica, mentre l'acetilcolina liberata dall'impulso nervoso arriva a contatto diretto e agisce in tutta la superficie della placca (v. fig. 2).
Una tecnica ancor più raffinata è stata in seguito messa a punto da McMahan, Kuffler e dai loro collaboratori (v. Peper e McMahan, 1972). Il disegno della fig. 4A è tratto da una microfotografia di una fibra muscolare vivente identificata attraverso le striature trasversali; sulla sua superficie sono visibili due fibre nervose terminali che la percorrono longitudinalmente. Questa bella riproduzione è dovuta a una tecnica speciale detta microscopia a interferenza di Nomarski, che permette di studiare il tessuto vivente a forte ingrandimento e con immagini ottiche molto nitide. Il fatto che le due terminazioni nervose siano disposte parallelamente e che finiscano alla stessa altezza rappresenta una notevole facilitazione per il ricercatore.
Al di sotto della fig. 4B si può vedere la fibra muscolare in sezione, con le due terminazioni situate in piccoli solchi, a e d. Nella fibra muscolare è inserito un elettrodo di registrazione intracellulare che si trova al di fuori dell'area rappresentata nella fig. 4A, ma a una distanza che è piuttosto piccola rispetto alla costante di spazio della fibra muscolare, in modo tale da registrare i cambiamenti di potenziale attraverso la membrana con distorsioni minime.
Iniezioni elettroforetiche di ACh venivano effettuate tramite la micropipetta, E, inserita con precisione nel punto c sulla membrana muscolare fra le due terminazioni nervose. La corrente di iniezione era di 3,2 × 10-8 A, applicata per la durata di 1 ms, e i punti di iniezione sono segnati con cerchi neri lungo la linea che congiunge le due terminazioni nervose. Quando il punto di applicazione era in corrispondenza della terminazione nervosa (a nella fig. 4A), l'ACh evocava una grande depolarizzazione (a nella fig. 4, B e C). La stessa iniezione a soli 5 μm di distanza (b in A) evocava una depolarizzazione più piccola (b in B e C), mentre quando l'iniezione era fatta a metà strada fra le due terminazioni (c in A), l'ACh aveva scarso effetto sul potenziale di membrana (c in B e C). I risultati di altre iniezioni sono riportati nella fig. 4B, e mostrano quanto precisa e delicata sia questa tecnica; al di sotto della fig. 4B sono riportate sulla stessa scala le due terminazioni.
Che cosa mostra questo esperimento? Esso dimostra, in modo veramente chiaro, che soltanto la parte di membrana muscolare che si trova sotto le fibre nervose è sensibile all'ACh. Il resto è meno sensibile di almeno due ordini di grandezza. Gli effetti ottenuti in altre zone sono in massima parte dovuti alla diffusione dell'ACh verso i siti sensibili che si trovano in corrispondenza dei contatti con le terminazioni nervose. Lo stesso decremento nella risposta osservato da a a c si osserva anche quando si allontana la punta dell'elettrodo, spostandola verticalmente dall'area della fibra nervosa. Si può pertanto concludere che la zona di sensibilità all'ACh si trova localizzata esattamente in corrispondenza del solco dove ha luogo il contatto con la fibra nervosa.
Un esperimento ancora più raffinato è stato eseguito più recentemente da Kuffler e Yoshikami (v. i contributi del 1975) sulle sinapsi delle fibre muscolari di serpente. In tale esperimento essi eliminavano il tessuto fibroso mediante pretrattamento con collagenasi, in modo da separare la terminazione nervosa con tutto l'apparato presinaptico dalla zona postsinaptica che rimane come un ‛cratere' direttamente accessibile all'applicazione elettroforetica di ACh (v. fig. 11D). Si è potuto così dimostrare che l'alta sensibilità all'ACh è localizzata esattamente nel cratere. Dettagli quantitativi saranno esposti in seguito.
e) Natura quantica dell'azione del trasmettitore neuromuscolare.
La fig. 5 illustra una delle proprietà più importanti di tutte le sinapsi a trasmissione chimica, cioè la natura quantica della loro azione, che è stata scoperta da Katz e dai suoi collaboratori (v. Fatt e Katz, 1952; v. Katz, 1958 e 1962). L'ACh è liberata in pacchetti relativamente grandi di 10.000 o più molecole. Questa liberazione di quanti procede spontaneamente in maniera lenta, come si può osservare quando l'elettrodo intracellulare è posto in prossimità della placca. Per esempio, nella fig. 5A vi è una successione di piccoli potenziali che in ogni aspetto rassomigliano a potenziali di placca miniaturizzati, da cui la loro definizione di potenziali di placca in miniatura (PP in miniatura); il tracciato inferiore è la risposta a un impulso nervoso ad amplificazione molto più bassa e rassomiglia molto al primo potenziale d'azione della fig. 3B. Al contrario, nella fig. 5B a 2 mm di distanza dalla placca non c'è quasi traccia dei potenziali in miniatura, e il potenziale d'azione del tracciato più basso rassomiglia all'ultimo della fig. 3B.
Nei tracciati più in basso della fig. 5C i PP in miniatura si producono a una frequenza molto più bassa che in A; il tracciato superiore rappresenta il potenziale prodotto da una breve emissione elettroforetica di ACh al tempo dell'impulso (ACh). I tracciati in D sono stati registrati immediatamente dopo quelli di C, ma il preparato era sotto l'azione dell'anticolinesterasico edrofonio iniettato per elettroforesi al momento dell'impulso E. Si noti che la depolarizzazione prodotta dall'iniezione di ACh risulta aumentata e prolungata e che aumentati e prolungati sono anche i PP in miniatura. In conseguenza di indagini molto approfondite e accurate si può ormai asserire che i PP in miniatura presentano sotto ogni aspetto il comportamento previsto per la liberazione spontanea di quanti di ACh dalla terminazione nervosa. Per esempio essi vengono aumentati e prolungati da tutti gli anticolinesterasici (v. fig. 5D) e diminuiti da tutti gli agenti curarizzanti nel medesimo modo che il PP.
Subito dopo la scoperta dei PP in miniatura sono state individuate nelle terminazioni nervose le vescicole sinaptiche (v. fig. 2B). La correlazione fu ovvia e attraente. Dopo quasi vent'anni di ricerche sembra ora fuor di dubbio che le vescicole sinaptiche sono pacchetti preformati di quanti della sostanza trasmettitrice e che in condizioni di riposo le vescicole scaricano il loro contenuto nella fessura sinaptica a intervalli relativamente poco frequenti, producendo così i PP in miniatura. Un'ulteriore scoperta degna di nota è quella, effettuata da Katz e collaboratori (v. Fatt e Katz, 1952; v. Katz, 1958, 1962 e 1966), che la liberazione spontanea di quanti avviene a tempi del tutto casuali.
Se si ammette che il trasmettitore sia racchiuso nelle vescicole sinaptiche, ci si deve aspettare che la sua liberazione da parte di un impulso nervoso sia di natura quantica. Tuttavia, normalmente il PP ha un'ampiezza che indica che esso è composto da molti quanti, cosicché non è stato possibile individuare una composizione in quanti.
Si può ridurre a qualsiasi grandezza il PP diminuendo il calcio nella soluzione del bagno e aggiungendo magnesio. Per esempio, nei 18 tracciati della fig. 6A l'impulso nervoso ha evocato un piccolo PP solo 4 volte, in coincidenza della seconda linea tratteggiata. Tre di questi PP sono uguali ai due PP in miniatura spontanei che si vedono più tardivamente nei due tracciati, mentre il quarto è chiaramente doppio. La fig. 6B mostra un'altra serie di tracciati con una depressione della trasmissione neuromuscolare meno pronunciata; in due il PP era assente, in due aveva l'ampiezza dei PP in miniatura e in tre un'ampiezza doppia. Tali osservazioni dimostrano che quando l'ampiezza dei PP è sufficientemente ridotta, essi rivelano una composizione quantica corrispondente a quella dei PP in miniatura.
Le serie di doppi tracciati della fig. 6C sono state registrate da un elettrodo extracellulare in vicinanza della regione della placca, quando la composizione quantica del PP veniva ridotta come conseguenza di una diminuzione della concentrazione di calcio. La registrazione extracellulare è molto più localizzata di quella intracellulare, eppure si può vedere che vi è un'ampia variazione di latenze dei PP quantici. Invece non c'è variazione di latenza nel potenziale bifasico iniziale prodotto da un impulso nervoso. La variazione di latenza del PP è illustrata nell'istogramma della fig. 6D ottenuto da numerosi tracciati, alcuni dei quali sono mostrati in C. Si può quindi concludere che la liberazione di quanti di trasmettitore da parte di un impulso nervoso varia entro considerevoli limiti di latenza. Una spiegazione di questa variabilità sarà data più avanti.
L'analisi statistica ottenuta applicando il teorema di Poisson (v. Katz, 1958 e 1962) dà una precisa conferma di questa composizione quantica dei PP, e mostra che i quanti sono identici a quelli che sono liberati casualmente e che producono i PP in miniatura. In effetti, ricerche dettagliate effettuate da Katz e collaboratori hanno mostrato che l'emissione di quanti di ACh concorda con le previsioni fatte sulla base del teorema di Poisson per la liberazione spontanea di un piccolo numero di quanti (v. Katz, 1966 e 1969).
È ormai stato stabilito che un impulso nervoso genera un PP attraverso la liberazione di un certo numero di quanti di ACh. Nascono ora due problemi quantitativi. Anzitutto, qual è il numero di quanti liberati in corrispondenza della placca in presenza di una quantità normale di calcio? Si calcola fra 100 e 300 (v. Hubbard, 1973). In secondo luogo, quante molecole vi sono in un quanto di ACh? Le stime variano entro ampi limiti (v. Hubbard, 1970 e 1973), ma, come descritto sotto, Kuffler e Yoshikami (v., The distribution..., 1975) hanno calcolato che il limite massimo è di 10.000 molecole, valore che corrisponderebbe a una soluzione moderatamente ipertonica.
In principio si pensava che i pacchetti di quanti fossero molto uniformi in grandezza, ma ora sembra che i più grandi possano essere il doppio dei più piccoli.
f) Fattori che controllano l'emissione di quanti dalla terminazione nervosa.
Se vogliamo esaminare più in dettaglio il modo in cui un impulso nervoso provoca la liberazione di quanti dalla terminazione nervosa, è importante stabilire se l'impulso nervoso arriva davvero nella terminazione nervosa o se, invece, la propagazione cessi immediatamente prima di questa e vi sia pertanto solo una propagazione elettrotonica della depolarizzazione dovuta al fatto che la terminazione si comporta come un cavo. Esperimenti come quelli illustrati nella fig. 7 (v. Katz e Miledi, Propagation of..., 1965) dimostrano che normalmente l'impulso viaggia lungo le sottili fibre nervose terminali della giunzione neuromuscolare della rana e che questa propagazione è essenziale per la sua efficacia. Nella fig. 7 gli elettrodi registranti si trovano, in A, su una fibra nervosa terminale vicino alla sua origine (a) e alla sua estremità (b). In B (prossimale) il potenziale d'azione del nervo ha la tipica configurazione trifasica (positivo-negativo-positivo) che si ha quando un impulso si propaga vicino a un elettrodo registrante di superficie, mentre in B (distale) esso è solo positivo-negativo, come ci si deve aspettare per un impulso che si propaga alla fine di una fibra. La registrazione più veloce di C permette di calcolare la velocità di conduzione di questa fibra terminale, che risulta essere di circa 0,4 m/s.
Il diagramma inserito sopra la fig. 7D mostra le condizioni sperimentali di un ulteriore esperimento di prova fatto da Katz e Miledi (v., The effect..., 1968). Si stimola il nervo (N) e si pongono tre elettrodi lungo una sottile fibra nervosa terminale. Una localizzazione accurata della liberazione dei quanti è assicurata dall'impiego di un rapporto Ca/Mg molto sfavorevole al fine di abolire ogni emissione di quanti tranne che nella zona immediatamente vicina alle micropipette registranti (CaCl2), con le quali si procede a una liberazione controllata di Ca2+. In tali condizioni l'impulso nervoso ha evocato un PP (v. fig. 7D) in entrambe le zone di registrazione, prossimale (1) e distale (2). Quindi dalla pipetta di mezzo viene emessa tetrodotossina (TTX), che agisce bloccando il nervo e determina una mancanza totale del PP (v. fig. 7E) nell'elettrodo distale (2), mentre in quello prossimale non si sono avuti cambiamenti. In F si vede che, dopo che il blocco da TTX era terminato, il PP distale è ricomparso. Questo esperimento mostra che il blocco dell'impulso a meno di 100 μm dall'elettrodo distale ha causato la completa soppressione del PP. Con le attuali tecniche possiamo solo stabilire che affinché la trasmissione dell'impulso nervoso sia efficace è necessario che questo si propaghi fino a una distanza di almeno 50 μm dalle zone di liberazione del mediatore.
È importante ora correggere un possibile equivoco. Sebbene normalmente sia necessaria la propagazione dell'impulso nelle vicinanze del punto di trasmissione per causare la liberazione del trasmettitore dai punti terminali presinaptici, la fig. 8 mostra che l'impulso, di per sé, non ha alcuna azione specifica. La trasmissione dell'impulso nervoso è stata bloccata da TTX; in questo esperimento uno degli elettrodi per il passaggio di corrente era collocato nel punto in cui il nervo entra nel muscolo, e l'altro sempre sul nervo ma in un punto più lontano. Quando si fanno passare forti correnti depolarizzanti nel primo punto, la registrazione per via intracellulare rivela sulla membrana del muscolo grandi depolarizzazioni che ricordano i PP. Correnti più deboli si mostrano prive di effetto (0,93 μA) o meno efficaci (1,85 μA). Evidentemente la depolarizzazione applicata si diffonde elettrotonicamente come in un cavo, depolarizzando le parti terminali presinaptiche e provocando così la liberazione di trasmettitore, esattamente come accadrebbe per un impulso nervoso. Si può concludere che è la depolarizzazione dell'impulso nervoso che fa scattare il meccanismo di liberazione del trasmettitore e non una qualche altra influenza, quale, per esempio, il flusso di sodio verso l'interno durante la fase di ascesa dell'impulso nervoso. Anzi, gli ioni sodio non sono affatto implicati nella liberazione del trasmettitore, che può avvenire efficacemente anche in un mezzo privo di sodio, quale la paraffina liquida. D'altra parte, esiste una quantità di prove sperimentali che dimostrano come il calcio extracellulare sia essenziale per la liberazione del trasmettitore a livello di qualunque tipo di sinapsi chimica. Negli esperimenti illustrati nelle figg. 6 e 7 si è tratto vantaggio dal fatto che la liberazione del trasmettitore può essere molto limitata o perfino abolita eliminando il calcio extracellulare. La fig. 9 illustra un'indagine più analitica su questo ruolo chiave del calcio.
g) Ruolo essenziale del calcio nella liberazione dei quanti.
Le condizioni sperimentali della fig. 9A sono mostrate sotto le tre serie di tracce sovrapposte evocate dalla stimolazione del nervo. La micropipetta che registra extracellularmente è stata riempita con una soluzione 0,5 M di CaCl2, ma la diffusione verso l'esterno di ioni Ca2+ è stata prevenuta con un voltaggio negativo (v. fig. 9A, tracciato superiore). L'impulso nervoso era completamente privo di effetto, poiché si avevano le stesse condizioni molto sfavorevoli per la liberazione del trasmettitore, cioè un mezzo privo di Ca2+ e contenente Mg2 0,84 mM. Nel secondo e terzo tracciato la polarizzazione negativa della micropipetta è stata ridotta in due stadi, in modo da permettere una certa diffusione di Ca2+ verso l'esterno, e da innalzare il livello di Ca2+ in vicinanza del punto di registrazione, tanto da consentire una certa liberazione di quanti da parte dell'impulso nervoso. Questa efficacia scompariva non appena si ripristinava il voltaggio frenante iniziale cui corrispondeva nuovamente la risposta zero illustrata nel primo tracciato. In B sono mostrate le medie di 600 tracciati delle corrispondenti registrazioni di A. È interessante notare che il potenziale dell'impulso nervoso trifasico rimane inalterato con l'aumento del calcio.
Il ruolo essenziale del calcio extracellulare per la liberazione del trasmettitore può essere dimostrato anche in assenza di impulsi nervosi. Le condizioni sperimentali sono illustrate sotto la fig. 9C. L'eliminazione di impulsi nervosi è stata ottenuta con TTX, e due micropipette sono sistemate l'una vicino all'altra su una delle fibre nervose che terminano sulla fibra muscolare dalla quale si registra intracellularmente.
La soluzione in cui si trova il preparato è priva di calcio e ha una concentrazione 1,7 mM in Mg2+. Brevi applicazioni di correnti depolarizzanti mediante l'elettrodo D non provocano liberazione del trasmettitore (v. fig. 9C, tracciato superiore). Quindi il voltaggio negativo frenante sull'elettrodo contenente Ca2+ è stato ridotto per permettere al Ca2+ di diffondere intorno alla terminazione nervosa, per cui lo stesso impulso depolarizzante in questo caso rilasciava il trasmettitore, come dimostrano gli ampi PP del secondo tracciato. Il terzo tracciato dimostra lo stesso fenomeno a una velocità di scansione molto inferiore. Gli impulsi depolarizzanti sono indicati dai punti sotto il tracciato, e la polarizzazione sulla pipetta contenente Ca2+ è stata applicata tra le due frecce superiori. Si nota l'immediata comparsa di PP a ogni impulso, sebbene vi sia una considerevole variazione nel loro contenuto di quanti. Ripristinando il voltaggio limitante, ancora una volta i PP scompaiono.
Le figg. 9 D, E ed F illustrano ricerche sperimentali sulla velocità di azione del Ca2+ liberato per elettroforesi. Le condizioni sperimentali sono identiche a quelle di C, come dimostrato nel diagramma tra C ed F. I tracciati inferiori di D, E ed F mostrano il momento in cui venivano applicati gli impulsi attraverso le micropipette, marcate nel diagramma con D e Ca. In nessuna delle cinque registrazioni di D vi è liberazione di trasmettitore, allo stesso modo che nel tracciato superiore di C. Tuttavia la situazione in E differisce da quella in D, poiché il calcio veniva liberato da un impulso molto breve applicato attraverso la micropipetta Ca, immediatamente prima dell'impulso depolarizzante, D. Anche quando la liberazione di Ca2+ avviene soltanto 2 o 3 ms prima, l'impulso depolarizzante provoca la liberazione di alcuni quanti che producono PP in ogni traccia. Tuttavia si vede che le latenze della liberazione di quanti variano entro ampi limiti, e un impulso può indurre la liberazione di due quanti con un notevole intervallo di tempo, come nel tracciato superiore di E. Data la grande variabilità delle latenze dei PP in E, è sorprendente riscontrare che se l'impulso di Ca2+ avveniva immediatamente dopo l'impulso di D, l'iniezione di Ca2+ era completamente priva di effetto (v. fig. 9F). Evidentemente l'impulso D è efficace soltanto se agisce in presenza di Ca2+ extracellulare. L'ampia variazione della latenza deve essere attribuita a qualche avvenimento che si verifica dopo che l'impulso D ha utilizzato il Ca2+ per la liberazione di quanti.
h) Quadro dei probabili eventi che hanno luogo nella trasmissione neuromuscolare.
La fig. 10 illustra schematicamente gli elementi essenziali che si ipotizza entrino in gioco nella trasmissione a livello della sinapsi neuromuscolare. Nella fig. 10A si vede la terminazione nervosa con le vescicole sinaptiche, delle quali una è in stretta vicinanza della membrana presinaptica e una sta scaricando il suo contenuto (il quanto di molecole di ACh) dentro la fessura sinaptica; a sinistra, una vescicola collassata che sta ricostruendo la sua riserva di ACh. L'enzima colinacetiltransferasi sembra trovarsi nel citoplasma, e utilizza, per la formazione di acetilcolina, colina e acido acetico (v. fig. 10B). Quando la membrana presinaptica è depolarizzata al di sopra di un livello critico, si aprono i canali che permettono agli ioni Ca2+, o alternativamente ad alcuni complessi carichi del Ca2+ (CaX+), di raggiungere la parte più interna della membrana, nella quale il calcio può provocare qualche variazione, permettendo alle vescicole di scaricare il loro contenuto. Poiché il rilascio di quanti di acetilcolina è approssimativamente proporzionale alla quarta potenza della concentrazione esterna di Ca2+, si postula che siano necessari 4 ioni Ca2+ per scaricare un quanto (v. Dodge e Rahamimoff, 1967). Nella fig. 10A le molecole del trasmettitore diffondono attraverso la fessura sinaptica per attaccarsi ai siti del recettore sulla membrana postsinaptica. La fig. 10B mostra che in questo modo si produce una depolarizzazione (il PP) che, se sufficientemente ampia, evoca la scarica di un impulso lungo la fibra muscolare e così provoca una contrazione. Sulla membrana postsinaptica c'è anche acetilcolinesterasi (AChE) che idrolizza l'ACh; la colina così formata è riassorbita in sede presinaptica per essere usata ancora nella sintesi di ACh da parte dell'enzima colinacetiltransferasi nel citoplasma presinaptico. Si è postulato che l'ACh così prodotta sia concentrata nelle vescicole da un meccanismo a pompa localizzato sulla parete della vescicola. La fig. 10B illustra la doppia funzione dell'AChE: fa terminare l'azione depolarizzante dell'ACh e fornisce colina libera per la sintesi presinaptica di ACh.
i) Azione molecolare dell'acetilcolina (ACh).
Finora l'azione dell'ACh è stata studiata depolarizzando la membrana o tramite applicazione elettroforetica di un elevato numero di molecole di ACh - circa 20 milioni nella fig. 4 - o tramite emissione quantica di alcune migliaia di molecole, come nelle figg. 5 e 6. Recentemente, Katz e Miledi (v., 1972 e 1973) hanno compiuto un formidabile passo avanti in queste ricerche, riuscendo addirittura a identificare l'ampiezza e il decorso temporale delle depolarizzazioni prodotte da singole molecole di ACh. L'identificazione è stata ottenuta con l'analisi di frequenza spettrale del rumore elettrico evocato attraverso la membrana postsinaptica dall'applicazione di ACh (v. fig. 11, A e B). L'analisi del rumore effettuata da un calcolatore rivela che esso e costituito da un insieme casuale di piccolissimi componenti, ciascuno dei quali ha un'ampiezza di circa 0,3 μV e una durata di circa 1 ms. Questo rumore è almeno 1.000 volte più piccolo del ‛rumore biologico' scoperto da Fatt e Katz (v., 1952) ed è dovuto ai potenziali in miniatura. Attraverso metodologie sperimentali raffinatissime e controlli rigorosi, è stato possibile stabilire che questi piccoli potenziali sono dovuti a singole molecole di ACh, le quali transitoriamente (per circa 1 ms) si attaccano ai recettori della membrana postsinaptica e aprono i canali ionici, probabilmente attraverso un cambiamento di breve durata nella conformazione della struttura proteica del poro. Siccome il potenziale in miniatura del muscolo di rana è circa 1.500 volte più grande, è probabile che in un quanto (o in una vescicola sinaptica) ci siano circa 1.500 molecole di ACh. Tuttavia questo dovrebbe rappresentare un numero minimo, perché una considerevole frazione - almeno il 30% - delle molecole del quanto è idrolizzata dalla colinesterasi prima di poter raggiungere il recettore, come è indicato dall'interazione tra ACh-AChE nella fig. 10B.
Questo interessante aspetto, cioè il numero quantico, è stato studiato in maniera diversa da Kuffler e Yoshikami (v. i contributi del 1975). Questi autori irroravano con getti di ACh, applicati elettroforeticamente per la durata di 1 ms, i crateri messi a nudo nella giunzione neuromuscolare di serpente. Quando l'iniezione veniva effettuata con la pipetta in posizione molto vicina alla membrana, si otteneva una depolarizzazione che aveva una notevole somiglianza con il potenziale in miniatura, come illustrato nella fig. 11C.
Per calcolare il numero di molecole di ACh effettivamente iniettate elettroforeticamente, essi hanno utilizzato un metodo di determinazione biologico molto raffinato, arrivando a stabilire un numero quantico di 10.000. Tuttavia, gli stessi ricercatori considerano questo numero superiore a quello reale, perché il luogo di iniezione sopra il cratere non può trovarsi nella medesima posizione ottimale nella quale si trova invece la terminazione nervosa. Un possibile valore di 5.000 molecole per il serpente non sarebbe molto diverso dal minimo di 1.500 molecole calcolato da Katz e Miledi nella rana. È interessante notare che 5.000 molecole di ACh in una vescicola sferica di 450 Å di diametro darebbero una concentrazione pressoché isosmolare (v. Hubbard, 1973). Sulla base di un numero quantico di 5.000 e di un potenziale in miniatura di ampiezza media (1,5 mV), si può calcolare che una molecola di ACh provoca l'apertura di un canale ionico attraverso il quale passano 6.000 cationi univalenti in 1 ms. Katz e Miledi hanno calcolato per la rana un valore molto più alto, cioè 50.000, ottenuto, però, sulla base di un numero quantico molto basso, cioè 1.000.
Ho riportato in dettaglio questi lavori recenti perché bene illustrano il progresso dalla neurobiologia alla neurobiologia molecolare. Sono trascorsi solo 40 anni da quando Dale e i suoi collaboratori prospettarono la sensazionale ipotesi che la trasmissione neuromuscolare avvenisse tramite la secrezione di ACh da parte della terminazione nervosa. Oltre alle scoperte testé descritte, questa ricerca a livello molecolare ha portato a uno studio particolareggiato dei tempi di apertura dei canali ionici, degli analoghi dell'ACh, degli effetti della temperatura, degli agenti di blocco e degli anticolinesterasici. Non è stato sorprendente trovare che il curaro e la prostigmina, per esempio, non hanno effetto sull'azione unitaria dell'ACh. I recettori non occupati dal curaro si comportano con le molecole di ACh in maniera del tutto normale.
l) Eventi postsinaptici nella trasmissione neuromuscolare.
È ora il caso di considerare le conduttanze ioniche che concorrono a determinare la corrente di placca, che a sua volta dà origine al PP. Le uniche specie di ioni che si trovano in quantità tali da poter partecipare in modo significativo a tale conduttanza sono sodio, potassio e cloro. Al fine di registrare la corrente di placca (CP) è stata impiegata la tecnica del blocco di voltaggio (v. fig. 12A; v. Takeuchi e Takeuchi, 1960). Il potenziale di membrana veniva cambiato dal passaggio di un impulso a corrente continua attraverso di esso, mentre i potenziali di membrana reali sono riportati in ogni tracciato.
Com'è indicato dalla linea che congiunge i circoletti bianchi nella fig. 12B, la CP è in relazione lineare col potenziale di membrana, e l'estrapolazione mostra che esso è zero a un potenziale di membrana di −15 mV. La riduzione del sodio esterno di circa il 70% ha spostato verso destra la linea (circoletti neri), mentre il potenziale di equilibrio per la CP è passato da −15 a −32 mV. Si può ottenere una diminuzione del potenziale di equilibrio per il sodio anche aumentando il sodio intracellulare, il che provoca il medesimo spostamento del potenziale di equilibrio per la CP. Questi risultati indicano che gli ioni sodio trasportano la CP in parte considerevole. In modo analogo nella fig. 12C la riduzione della concentrazione extracellulare di K+ ha spostato il potenziale d'equilibrio per la CP di circa 28 mV in direzione depolarizzante. Questo effetto indica pertanto che ioni K+ trasportano una considerevole parte della CP.
Al contrario di quanto avviene in questi esperimenti con cationi, una variazione della concentrazione del cloro esterno o persino la sua eliminazione, come illustrato nella fig. 13A, non causa un apprezzabile cambiamento nel potenziale d'equilibrio della CP; infatti le due linee incrociano la linea di base nello stesso punto. Le linee hanno diversa pendenza, perché dopo la prima serie la concentrazione di tubocurarina è diminuita. Lo schema della fig. 13B mostra i canali per gli ioni Na+ e K+ attraverso la membrana subsinaptica con i loro rispettivi potenziali di equilibrio di +50 e −99 mV. L'ACh fa aumentare notevolmente la conduttanza ionica in questi due canali. Vi sono prove che nella regione della placca motrice si trovino due distinti canali ionici per il sodio e il potassio, cioè che i due canali della fig. 13B siano separati. La loro conduttanza relativa può essere modificata (entro ampi limiti) e la procaina cambia il decorso temporale della conduttanza del canale del potassio (v. Maeno, 1966; v. Gage e Armstrong, 1968). Tuttavia i pareri su questa supposta indipendenza sono ancora discordi (v. Hubbard, 1973; v. Krnjević, 1974). Si può calcolare che i canali ionici aperti dal trasmettitore abbiano una conduttanza molto alta; infatti i valori possono raggiungere, in maniera transitoria, 10-10 Ω-1 per i cambiamenti di conduttanza indotti da una singola molecola di ACh, e 5 × 10-8 Ω-1 nel caso di un quanto che produce un potenziale in miniatura (v. Gage e McBurney, 1972).
m) Attivazione sinaptica ripetitiva.
Quando la sinapsi neuromuscolare curarizzata di Anfibio viene attivata in maniera ripetitiva, si osserva una fase iniziale di notevole potenziamento che gradualmente declina verso una depressione costante dopo 15 o 20 risposte (v. fig. 14A). Quando il contenuto di quanti del PP viene fortemente ridotto per effetto di una bassa concentrazione di Ca2+ e/o alta di Mg2+, il potenziamento risulta molto aumentato e prolungato indefinitamente (v. fig. 14B; v. del Castillo e Katz, The membrane..., 1954; v. Braun e altri, 1966). La registrazione intracellulare nella zona della placca mostra che il potenziamento è interamente dovuto all'aumento del numero di quanti emessi da ciascun impulso. Il decorso temporale del declino di questo potenziamento è illustrato nella fig. 14C dopo l'applicazione di un breve tetano condizionante di 5 impulsi. La media di queste tracce mostra il rapido instaurarsi del potenziamento, mentre il suo successivo decorso temporale è provato da un singolo stimolo più tardivo a intervalli progressivamente più lunghi. È sorprendente constatare che il potenziamento risulta assai al di sopra di quello che si ha al quinto PP, nel caso che lo stimolo di prova avvenga a intervalli più brevi della durata di un ciclo, mentre successivamente il potenziamento declina rapidamente. Questo declino, tuttavia, raggiunge un livello costante di basso potenziamento che persiste per almeno 500 ms (v. Mallart e Martin, 1967).
Durante l'attivazione ripetitiva della sinapsi neuromuscolare di Mammifero sotto curaro, i PP successivi declinano progressivamente in ampiezza fino a raggiungere un livello costante di depressione dopo 10 o più impulsi (v. fig. 15A; v. Liley e North, 1953; v. Hubbard, 1963; v. Hubbard e Willis, 1962). Questa depressione è stata osservata per il PP evocato da una seconda stimolazione del nervo (v. Eccles e altri, 1941) e persisteva per vari secondi. La depressione non può essere attribuita a desensibilizzazione delle zone recettrici, poiché l'applicazione elettroforetica di ACh nella fig. 15D è altrettanto efficace che nel controllo (E) anche nel momento in cui si riscontra un'ampia depressione del PP di saggio (B). In C il PP di saggio a 1,9 s mostra un recupero quasi completo (v. Otsuka e altri, 1962). Quando la trasmissione neuromuscolare era depressa per effetto di una presenza bassa di Ca2+ e/o alta di Mg+2, il preparato di Mammifero si comportava come quello di Anfibio nel senso che la stimolazione ripetitiva produceva un potenziamento progressivo (v. fig. 15F): una risposta di saggio successiva, come nella fig. 15, F-I, mostra un declino a zero in circa 150 ms (v. Hubbard, 1963). È probabile che anche in questo caso il declino avvenga in due stadi, come dimostrato nel caso del preparato di Anfibio.
Katz e Miledi (v., The role..., 1968) hanno prospettato l'ipotesi che il breve e intenso potenziamento sia dovuto al calcio residuo entrato nella parte presinaptica terminale durante gli impulsi precedenti (v. fig. 10A). Basandoci sull'ipotesi che siano necessari 4 ioni Ca2+ per il rilascio di un quanto di trasmettitore (v. Dodge e Rahamimoff, 1967), l'effetto del calcio residuo doveva essere assai potente; e infatti, si è trovato che la facilitazione era assai più grande quando la concentrazione di Ca2+ era aumentata al momento dell'arrivo del primo impulso (v. fig. 9E). Inoltre, quando gli impulsi nervosi venivano eliminati da TTX si verificava il previsto aumento nell'emissione di quanti, allorché l'impulso depolarizzante durava più a lungo.
Se, come sembra probabile, il breve potenziamento iniziale è dovuto al calcio residuo sulla faccia più interna della membrana della parte terminale presinaptica, resta ancora da spiegare il potenziamento che dura per centinaia di millisecondi, e la ancor più lunga depressione. Alcuni ricercatori hanno avanzato l'ipotesi che nella zona terminale presinaptica debba trovarsi un qualche meccanismo biologico per la ricostituzione di quanti emessi in risposta a un impulso nervoso. Tale postulata mobilitazione di quanti entro la ‛zona di attivazione' sarebbe responsabile del prolungato potenziamento. Altrimenti alcuni dati indicano che l'impulso nervoso aumenta in grandezza durante questa fase ritardata di potenziamento (v. Hubbard e Schmidt, 1963; v. Hubbard, 1970) e questo sarebbe certo un fattore che contribuisce al potenziamento.
È importante notare che la depressione è assai più evidente nel preparato curarizzato (v. figg. 14A e 15, A e B), poiché in queste condizioni il primo impulso nervoso produce una piena emissione di quanti, mentre i PP più piccoli possono essere attribuiti al fatto che il curaro blocca i recettori postsinaptici. In tal modo la deplezione di trasmettitore diviene un fattore dominante durante la stimolazione ripetitiva, il che contrasta con il caso in cui il blocco neuromuscolare sia provocato dalla notevole diminuzione nell'emissione di quanti dovuta a una concentrazione bassa di Ca2+ e/o alta di Mg2+. In tali condizioni la depressione è trascurabile o assente durante e dopo la stimolazione ripetitiva (v. figg. 14B e 15F).
n) Fattori che causano l'interruzione della trasmissione neuromuscolare.
In condizioni normali il numero di quanti di ACh emessi è di molte volte più grande del numero minimo necessario per evocare un potenziale di placca capace di generare un impulso muscolare e quindi la contrazione del muscolo (v. fig. 10B). Pertanto, con tale margine di sicurezza la scarica ripetitiva degli impulsi dei motoneuroni, alla frequenza normale relativamente bassa di 10-30/s, continua a essere trasmessa al muscolo producendo una contrazione continua o tetano, come nel caso della singola unità motrice della fig. 1C. Se un intero muscolo si contrae fortemente, si determinerà una mancanza di circolazione nella massa muscolare in tensione, e questo interferirà con la capacità di mantenere per lunghi periodi forti contrazioni. Fortunatamente quasi tutti i movimenti naturali sono ritmici e richiedono solo brevi risposte tetaniche, per cui i dispositivi fisiologici della circolazione e il fattore di sicurezza della trasmissione sono adeguati per un quasi infinito prolungarsi di tali movimenti come nel camminare, correre, nuotare e anche respirare.
Nelle forme di myasthenia gravis (m.g.) la ricostituzione dei quanti di trasmettitore durante la stimolazione ripetitiva è inadeguata, per cui si verifica una progressiva mancanza di trasmissione neuromuscolare con conseguente mancanza di contrazione. Una cura efficace consiste nel dare un anticolinesterasico, come la prostigmina, che inibisce l'AChE e pertanto conserva l'ACh liberata nella fessura sinaptica (v. fig. 10A). (Dopo la stesura di questo articolo si è scoperto che la causa principale del blocco della trasmissione neuromuscolare nella m.g. è dovuta a una diminuzione dei siti dei recettori sulla placca motrice. Di regola questi siti subiscono un processo di degradazione e di ricostituzione, ma nella m.g. la frequenza può aumentare fino a 2 o 3 volte senza un ricambio compensativo. Si ammette in tal modo che la m.g. sia causata da una reazione autoimmune, ma tale scoperta non ha ancora portato a un trattamento efficace della malattia). Anche un blocco parziale della trasmissione provocato dal curaro o da simili agenti di blocco neuromuscolare può essere contrastato da un trattamento con anticolinesterasici. Questa scoperta è di grande importanza pratica: infatti il blocco neuromuscolare è utilizzato dagli anestesisti per assicurare il rilassamento muscolare richiesto dai chirurghi in molti interventi operatori. Gli anticolinesterasici ripristinano rapidamente la situazione normale dopo tale trattamento. La tossina di Clostridium botulinum blocca la trasmissione neuromuscolare prevenendo efficacemente la liberazione di quanti dalle terminazioni nervose, come è dimostrato sia dalla soppressione della liberazione spontanea che dà luogo ai PP in miniatura, sia da quella del potenziale di placca evocato da un impulso nervoso.
2. La sinapsi gigante del ganglio stellato del calamaro.
a) Generalità.
Ulteriori informazioni sui fenomeni fisiologici che si verificano nella trasmissione chimica della sinapsi sono state tratte da esperimenti sulla sinapsi gigante del ganglio stellato del calamaro. Il vantaggio peculiare di questo preparato sta nel fatto che la fibra presinaptica è così grande che si possono inserire microelettrodi sia al suo interno sia entro la fibra postsinaptica, come è illustrato nella fig. 16A. La sinapsi è veramente di dimensioni gigantesche; infatti il contatto tra una fibra presinaptica e una postsinaptica è lungo fino a 1 mm, e ogni fibra ha un diametro di centinaia di micron. Tuttavia, non c'è un contatto sinaptico continuo su questa intera area, ma una moltitudine di piccoli contatti, come indicato nella fig. 16A. Ci sono molte prove che indicano che si tratta di una sinapsi a trasmissione chimica: il caratteristico ritardo sinaptico tra la risposta presinaptica e quella postsinaptica; nessuna trasmissione elettrica diretta di un potenziale elettrotonico in alcuna direzione (v. Hagiwara e Tasaki, 1958); le funzioni reciprocamente antagoniste di ioni calcio e magnesio, proprio come avviene per la trasmissione neuromuscolare (v. Takeuchi e Takeuchi, 1962); la presenza di minuti potenziali sinaptici simili ai PP in miniatura, ma assai più piccoli (v. Miledi, 1967). Il trasmettitore non è stato identificato con certezza; sono stati forniti buoni argomenti a favore del fatto che si tratti di glutammato o di una sostanza molto simile, ma uno studio più recente di Miledi (v., 1969) ha fatto sorgere serie difficoltà, anche se forse non insormontabili. Fortunatamente questa incertezza non riguarda la nostra presente indagine sui meccanismi ionici responsabili della generazione dei potenziali postsinaptici.
b) Generazione di potenziali postsinaptici.
Nella fig. 16B la stimolazione diretta della fibra presinaptica evoca un potenziale d'azione, e la fibra postsinaptica viene a sua volta eccitata a vari intervalli da questo. Le tracce sovrapposte mostrano che il potenziale d'azione generato dallo stimolo postsinaptico ha esattamente la stessa ampiezza del potenziale d'azione generato per via sinaptica - rappresentato dal tracciato più marcato formato dalle varie tracce sovrapposte - quando lo stimolo postsinaptico segue la risposta presinaptica e conseguentemente non la modifica per effetto della refrattarietà. D'altra parte, quando lo stimolo presinaptico segue quello postsinaptico, la trasmissione sinaptica avviene in una fibra postsinaptica in stato di refrattarietà, e quindi esso provoca i PPSE sovrapposti senza i potenziali d'azione, salvo uno piccolo e tardivo al più lungo intervallo di prova. Tali osservazioni, descritte da Gage e Moore (v., 1969), differiscono da quelle della fig. 3B condotte sulla giunzione neuromuscolare, poiché il potenziale d'azione evocato sinapticamente non aveva mostrato alcun segno di riduzione per l'azione di cortocircuito delle aree della membrana postsinaptica attivate sinapticamente.
L'ampiezza della corrente eccitatoria postsinaptica (CEPS) veniva misurata con un meccanismo di blocco di voltaggio per un'ampia gamma di potenziali di membrana postsinaptici, ed espressa in forma di diagramma nella fig. 16C (v. anche fig. 12). Il diagramma lineare estrapolato mostra che il potenziale d'equilibrio per la CEPS è di circa +55 mV. In questa indagine di Gage e Moore il valore medio per 9 sinapsi è stato di +45 mV. Ricerche su un'altra specie di calamaro hanno fornito potenziali d'equilibrio compresi tra +13 e +40 mV (Miledi), valori che sono anch'essi assai piu vicini al potenziale d'equilibrio del sodio (circa +50 mV) che non il potenziale d'equilibrio del potenziale di placca. Sembra sicuro poter concludere che, nella membrana postsinaptica del calamaro, il canale di conduttanza del sodio predomina sul canale del potassio assai più che nella placca motrice. Tale scoperta offre un ulteriore sostegno all'ipotesi che queste due conduttanze ioniche operino attraverso canali indipendenti.
La grande dimensione della fibra presinaptica fa sì che il ganglio stellato del calamaro sia ideale per ricerche precise (v. Takeuchi e Takeuchi, 1962; v. Miledi e Slater, 1966) sul rapporto tra depolarizzazione presinaptica e liberazione del trasmettitore da essa indotta. Nella fig. 17 le tracce superiori e inferiori di ciascun esempio sono registrazioni intracellulari dalle componenti rispettivamente presinaptica e postsinaptica della sinapsi gigante. L'esempio segnato con O mostra le risposte di controllo ed evidenzia molto bene il ritardo sinaptico. Negli esempi segnati con −3, −7, −9 la fibra presinaptica è stata depolarizzata e negli esempi segnati con +3, +5, +6, +7, +8 iperpolarizzata da prolungate correnti con queste intensità relative. Come previsto, il potenziale d'azione presinaptico veniva depresso dalla depolarizzazione, e si osservava corrispondentemente una diminuzione della produzione del trasmettitore che veniva quasi soppressa dalla depolarizzazione più intensa (−9). L'iperpolarizzazione aveva l'effetto opposto, e a +8 la produzione del trasmettitore era talmente aumentata che il potenziale postsinaptico eccitatorio generava un potenziale d'azione. Evidentemente è necessaria un'ampia depolarizzazione presinaptica per determinare una pur minima liberazione di trasmettitore, e nel caso di depolarizzazioni maggiori la curva che esprime la relazione fra l'ampiezza dell'impulso presinaptico e la sua efficacia nel liberare il trasmettitore è molto ripida. È importante ricordare che durante questa indagine la capacità di risposta al trasmettitore della membrana postsinaptica era inalterata.
Un'indagine sperimentale meglio controllata sulla depolarizzazione presinaptica e sulla produzione di trasmettitore è illustrata nella fig. 18. Entrambi gli impulsi, presinaptico e postsinaptico, sono stati eliminati da TTX, e il diagramma nella parte superiore in G mostra che nella fibra presinaptica erano stati inseriti due elettrodi, uno (a) per portare brevi correnti depolarizzanti, l'altro (b) per registrare. Si osserva anche un elettrodo per la registrazione (c) nella fibra postsinaptica. Tutti questi elettrodi, e in particolare (b), venivano posti il più vicino possibile alla sinapsi, indicata dall'ispessimento della linea. A-F rappresentano 6 esempi con corrente presinaptica progressivamente maggiore, riportata nella traccia più in alto. La traccia più bassa in ciascun esempio fornisce il potenziale di membrana presinaptico, che diminuisce bruscamente con le più ampie depolarizzazioni anche durante il passaggio della corrente. Questo effetto può essere attribuito all'instaurarsi di una notevole conduttanza per il potassio attraverso la membrana, e viene chiamato ‛rettificazione ritardata'. La più piccola depolarizzazione presinaptica (v. fig. 18A) non determina alcuna produzione di trasmettitore, come dimostra l'assenza di un PPSE. Con successivi aumenti della depolarizzazione presinaptica (v. fig. 18, B-F) si verifica un aumento assai sensibile nella produzione del trasmettitore (v. anche fig. 17). Le misure riportate nel diagramma G mostrano che è stata necessaria una depolarizzazione presinaptica di circa 40 mV per provocare anche la più piccola produzione di trasmettitore; per depolarizzazioni da 60 fino a 100 mV la curva saliva rapidamente. In H si osserva una curva simile presa da un altro esperimento. Katz e Miledi (v., 1966 e A study..., 1967) riferiscono che il livello di soglia di depolarizzazione varia da 25 a 40 mV in diversi esperimenti.
Un confronto tra le figg. 17 e 18 mostra che la depolarizzazione passiva delle parti terminali presinaptiche è altrettanto efficace quanto il potenziale d'azione nel produrre liberazione di trasmettitore. Nelle ricerche sulla trasmissione neuromuscolare (v. figg. 7, 8 e 9) si era notato che, quando gli impulsi nervosi erano eliminati da TTX, una depolarizzazione prodotta localmente riusciva a indurre una produzione di trasmettitore. Ma un confronto quantitativo, quale quello delle figg. 17 e 18, è possibile solo quando la registrazione intracellulare della terminazione presinaptica dà una precisa misura delle rispettive depolarizzazioni.
c) Meccanismi ionici di generazione di potenziali postsinaptici.
Nella fig. 19A la trasmissione sinaptica è stata quasi completamente bloccata in una soluzione a basso contenuto di Ca2+. Essa veniva ripristinata nella fig. 19B dall'iniezione elettroforetica di una piccola quantità di calcio attraverso un microelettrodo vicino alla sinapsi (v. anche figg. 7 e 9). In tali condizioni la liberazione di 10-14-10-13 grammoioni Ca2+ in un secondo può essere sufficiente per ripristinare la trasmissione.
La fig. 19C mostra il decorso temporale del ripristino della trasmissione, durante un'applicazione elettroforetica costante di ioni Ca2+ (tra la prima e la seconda freccia). I cerchietti bianchi mostrano i PPSE intracellulari (traccecampioni inferiori di A e B) e i cerchietti neri le correnti postsinaptiche registrate per via extracellulare (tracce-campioni superiori di A e B). Evidentemente il calcio extracellulare è altrettanto necessario per la trasmissione sinaptica del ganglio stellato del calamaro quanto per la trasmissione neuromuscolare (v. figg. 6, 7 e 9).
Katz e Miledi (v., A study..., 1967; v. fig. 20) hanno impiegato un'ampia gamma di depolarizzazioni presinaptiche per analizzare il rapporto fra il trasporto verso l'interno di ioni Ca2+ e la liberazione di trasmettitore. La tetrodotossina (TTX) applicata esternamente eliminava la trasmissione dell'impulso e gli ioni tetraetilammonio (TEA) iniettati per elettroforesi nella fibra presinaptica eliminavano il meccanismo di trasporto del potassio; tale meccanismo viene attivato dalla depolarizzazione e produce una rettificazione ritardata che fa diminuire rapidamente la depolarizzazione presinaptica durante i brevi impulsi depolarizzanti della fig. 18.
Nella fig. 20, A-F, si verificava un progressivo aumento dell'impulso di corrente depolarizzante (tracciato superiore) applicata attraverso un elettrodo intracellulare presinaptico. A causa dell'eliminazione dei canali di conduttanza al potassio, le correnti producevano un potenziale presinaptico abbastanza costante (tracciato inferiore) che in F era spostato di 200 mV dal potenziale di riposo. Il PPSE registrato nel tracciato intermedio dà un'indicazione della liberazione di trasmettitore sinaptico. In A si aveva un breve picco e in B un picco iniziale più largo con declino ritardato; in C il notevole mutamento di potenziale determinava una considerevole liberazione di trasmettitore per tutta la durata dell'impulso; in D un cambiamento ancor più notevole era associato a una minore liberazione di trasmettitore, ma v'era un breve picco di liberazione alla fine dell'impulso: questi effetti aumentavano in E; infine in F non v'era quasi liberazione durante il cambiamento di potenziale assai notevole di 200 mV, ma un ‛effetto di chiusura' molto grande. Evidentemente, come suggerito da Katz e Miledi (v., A study..., 1967), sono coinvolti 2 fattori: 1) la depolarizzazione di membrana apre i canali del calcio allo stesso modo in cui apre i canali per il potassio e il sodio nel caso della fibra nervosa e della fibra muscolare (Hodgkin e Huxley), e anche in questo caso la soglia si ha a circa 30-40 mV di depolarizzazione (v. fig. 18); tanto più grande è la depolarizzazione, tanto più efficace è l'apertura dei canali del calcio, come mostrato nella fig. 18; 2) tuttavia il gradiente elettrochimico per gli ioni Ca2+ diviene progressivamente inferiore man mano che cresce la positività interna e in tal modo limita l'entrata di Ca2+ e la conseguente liberazione di trasmettitore. Infine, a circa 200 mV, cioè a un potenziale di membrana inverso di circa +130 mV, si raggiunge presumibilmente il potenziale di equilibrio per il Ca2+, cosicché, praticamente, non v'è passaggio di Ca2+ attraverso i suoi canali largamente aperti. Tuttavia l'effetto di chiusura (D-F) è prevedibile se il potenziale di membrana cade più rapidamente di quanto si chiudano i canali del Ca, cosicché momentaneamente gli ioni Ca2+ possono passare attraverso i canali sotto un favorevole gradiente elettrochimico. Si deve ricordare che in realtà non esiste una misura precisa della concentrazione di ioni Ca2+ intracellulari liberi negli assoni del calamaro, ma è noto che essa è estremamente bassa (v. Hodgkin e Keynes, 1957; v. Baker e altri, 1970) e pertanto è possibile che il potenziale di equilibrio arrivi a +130 mV.
Nella fig. 20, G-I, è stata applicata la stessa grande corrente depolarizzante come in F, ma di durata più breve. Un effetto di chiusura virtualmente identico segue entro 1 ms dall'impulso di corrente, proprio come in F. Pertanto l'effetto di chiusura è chiaramente in relazione alla terminazione dell'impulso. Evidentemente la chiusura ritardata dei canali del calcio giustifica gli effetti di chiusura mostrati nella fig. 20, G-I, proprio nello stesso modo che nella fig. 20, D-F.
3. I canali ionici attraverso le membrane presinaptica e postsinaptica di una sinapsi a trasmissione chimica.
La fig. 21 offre una rappresentazione diagrammatica dei vari canali ionici attraverso la membrana postsinaptica della giunzione neuromuscolare e della sinapsi gigante del calamaro, in cui vi sono canali NaT e KT separati per le correnti postsinaptiche (v. anche fig. 13B). Nella fibra nervosa presinaptica sono mostrati anche i canali per il Na+ e il K+ il cui ingresso viene aperto dalla depolarizzazione, e che sono responsabili dell'aumento e della caduta del potenziale d'azione. Essi sono contrassegnati NaD e KD per indicare il loro rapporto con la depolarizzazione. Sebbene selettivamente permeabile agli ioni sodio, il canale NaD può servire anche per il passaggio di Li+ (v. Hodgkin, 1964) e, in qualche misura, per K+, Rb+ e Cs+ (v. Chandier e Meeves, 1965). Gli ioni ammonio e alcuni sostituti possono passare attraverso il canale NaT (per una bibliografia sull'argomento, v. Eccles, 1964, p. 52). Il canale CaD è anche permeabile a Sr2+ e Ba2+, ma meno efficacemente (v. Dodge e altri, 1969). È importante notare che la TTX blocca selettivamente il canale NaD, ma non ha alcun effetto sui canali KD e CaD (v. Katz e Miledi, 1969), mentre il TEA intracellulare blocca solo il canale KD. Il canale CaD si distingue per il fatto che è bloccato efficacemente dalla presenza di Mn2+ e Mg2+ all'esterno (v. Katz e Miledi, 1969). Quando i canali NaD e KD sono bloccati rispettivamente da TTX e TEA, il canale CaD diviene così attivo da mostrare anche una risposta depolarizzante di tipo rigenerativo (v. Katz e Miledi, 1969).
Con un metodo estremamente raffinato di fluorescenza per rilevare la presenza di calcio, Baker e altri (v., 1970) hanno mostrato che la depolarizzazione di membrana è associata a un'entrata di Ca2+ in due stadi, uno con quasi lo stesso decorso temporale dell'Na+, e l'altro, più lento nell'inizio, che continua durante la depolarizzazione. Il primo è bloccato da TTX e può essere attribuito a un modesto trasporto di Ca2+ attraverso il canale NaD; l'altro è resistente alla TTX ed è bloccato da Mn2+ e Mg2+, e corrisponde in maniera precisa al canale CaD della fig. 21.
Si può calcolare che anche se sono necessari 4 ioni Ca2+ per la liberazione di ogni quanto (v. sopra), la corrente del calcio verso l'interno durante una tale risposta rigenerativa provoca un'entrata di Ca2+ 10.000 volte superiore a quella necessaria per la liberazione del trasmettitore. Katz e Miledi (v., 1969) suggeriscono che questa grande quantità in eccesso possa essere richiesta per provocare la necessaria azione quadrupla in ciascuna sede. Si può suggerire che la considerevole variabilità nei ritardi con cui avviene la liberazione di quanti nella sinapsi (v. figg. 6 e 9E) sia dovuta al tempo impiegato dagli ioni Ca2+ a raggiungere una vescicola in posizione di scaricare il suo quanto. Anche con tale grande sovrabbondanza di ioni Ca2+, questo fattore potrebbe essere coinvolto soprattutto nello stabilire la durata del ritardo sinaptico.
La fig. 21 illustra due caratteristiche dei cinque tipi di canali ionici rappresentati: in primo luogo le specificità relative dei canali stessi, come simbolizzato dai vari ioni principalmente interessati in condizioni normali. Presumibilmente si tratta di specificità a base chimica, e la scoperta di questi meccanismi chimici sarebbe una grande conquista per la scienza. Le diverse proprietà dei vari tipi fanno presumere che ognuno di essi abbia una composizione chimica unica.
La seconda caratteristica riguarda gli ingressi che controllano questi canali, che vengono aperti sotto depolarizzazione nel caso dei canali NaD e KD e per azione del trasmettitore chimico (cioè acetilcolina nella sinapsi neuromuscolare e probabilmente glutammato nella sinapsi del calamaro), nel caso dei canali NaT e KT . Un'ulteriore differenza consiste nel fatto che i canali NaD e KD sono bloccati rispettivamente da TTX extracellulare e TEA intracellulare che lasciano invece inalterati i canali NaT e KT, aperti dall'azione del trasmettitore nelle sinapsi neuromuscolari e in quelle del calamaro. Per questa ragione i simboli all'interno dei canali della fig. 21 sono diversi, come pure i contrassegni NaT e KT , così da evidenziare i diversi meccanismi chimici.
La fig. 21B illustra ulteriormente una caratteristica speciale della membrana presinaptica, cioè la presenza di canali specifici per il calcio, CaD , azionati dalla depolarizzazione di membrana. L'entrata di ioni Ca2+ lungo tali canali è implicata soprattutto nella liberazione di trasmettitore dentro la fessura sinaptica (v. figg. 7, 9, 19 e 20). In accordo con i dati di Katz e Miledi (v., Propagation of.., 1965; v. fig. 7, A-C), nella fig. 21, A e B, i canali NaD e KD si trovano lungo la parte terminale del nervo, ma non nella componente subsinaptica della membrana postsinaptica, nella quale vi sono esclusivamente i canali NaT e KT, ma certo si trovano almeno in stretta vicinanza. È interessante notare che i canali CaD vengono aperti approssimativamente agli stessi livelli di depolarizzazione (v. figg. 17, 18 e 20) dei canali NaD e KD, che sono associati al potenziale d'azione. Nella fig. 21B, i canali CaD sono rappresentati solo sulla membrana presinaptica e non sono bloccati da TTX: ciò è in contrasto con quanto osservato per i canali CaD di alcune membrane eccitabili (v. Baker e altri, 1970).
I simboli che rappresentano i vari tipi di canali con le loro porte nella fig. 21 B non devono essere presi troppo alla lettera, anche se è stata avanzata l'ipotesi che ogni tipo abbia una composizione chimica peculiare. Inoltre, vi sono le porte che controllano tali canali e che possono essere aperte da depolarizzazione (NaD, KD e CaD) o dall'azione del trasmettitore sinaptico o da qualche sostanza simile (NaT e KT).
La sinapsi neuromuscolare e la sinapsi gigante del calamaro esemplificano molto bene il meccanismo della trasmissione chimica, e sembrano molto simili negli elementi essenziali, sebbene differiscano riguardo alle sostanze trasmettitrici e alle relazioni dimensionali; esse differiscono anche nei rapporti di conduttanza degli ioni per i canali NaT e KT della membrana postsinaptica. Un rapporto di 1,3 è stato determinato dai Takeuchi per la sinapsi neuromuscolare (v. figg. 12 e 13), ma nella sinapsi del calamaro il rapporto deve essere superiore a 5 (v. fig. 16). Nel diagramma della fig. 21B questo potrebbe essere simbolizzato dal numero relativo di canali NaT e KT. Invece del rapporto di circa 1,3 per la sinapsi neuromuscolare, per la sinapsi del calamaro potrebbe esserci, per esempio, un rapporto di 5.
4. Altre sinapsi periferiche a trasmissione chimica.
Nei Vertebrati sono state studiate molte sinapsi periferiche a trasmissione chimica, ma non sono stati scoperti nuovi principî di funzionamento. Per ora tutte le ricerche hanno mostrato che tutte hanno vescicole sinaptiche e che la trasmissione è quantica. È stata riscontrata la presenza di potenziali sinaptici in miniatura e si ritiene che la trasmissione, nella risposta alla scarica del nervo, sia quantica, secondo il teorema di Poisson, sebbene nel caso di molti tipi di sinapsi manchino ricerche in proposito. Il ruolo essenziale del Ca2+ per la liberazione di trasmettitore sembra una proprietà sinaptica generale, come anche il ruolo antagonista del Mg2+. Ulteriori progressi nella comprensione della trasmissione sinaptica in un sistema di gangli perifeci di Vertebrati dipendono dalla scoperta di un preparato di una singola cellula nervosa che possa essere studiato in isolamento e in condizioni controllate, in modo da poter osservare le sinapsi direttamente nel preparato vivente e sottoporle a ricerche sperimentali durante le quali si registra dalla cellula mediante un microelettrodo intracellulare. Tutti questi requisiti sono stati soddisfatti in ricerche sulle cellule gangliari del setto di cuore di rana (v. McMahan e Kuffler, 1969; v. Dennis e altri, 1971). Un rapido resoconto di questo elegante lavoro completerà la nostra breve rassegna sulla trasmissione sinaptica periferica.
Nel piccolo inserto della fig. 22A è rappresentata una cellula gangliare (CG), con il suo nucleo e il suo nucleolo, il cui assone si ramifica distribuendosi alle 2 fibre del muscolo cardiaco (M). La fibra vagale (V) si avvicina alla cellula formando una spirale intorno all'assone e termina in una serie di sinapsi, mostrate come espansioni scure, sulla superficie del corpo della cellula. Non vi sono dendriti. Nella fig. 22A tale cellula del ganglio è vista a forte ingrandimento in sezione ottica mediante la tecnica a interferenza di Nomarski. La microfotografia della fig. 22B mostra, con lo stesso ingrandimento, la maggior parte della cellula gangliare; il nucleo e il nucleolo servono come punti di riferimento. Infine, la fig. 22C è un forte ingrandimento della microfotografia della fig. 22B; si vede con chiarezza un bottone sinaptico pieno di vescicole e con la fessura sinaptica parzialmente differenziata come un sito attivo colorato più scuro. Questa sinapsi può essere identificata non solo in B, ma anche in A come la struttura mal definita entro il riquadro. In tal modo Kuffler e i suoi collaboratori hanno potuto individuare per la prima volta le sinapsi nella cellula vivente (A). Un ulteriore sviluppo di questa tecnica ha permesso di colorare con blu di metilene o con ossido di zinco e osmio le sinapsi, in modo da renderle più facilmente visibili allo stato vivente.
Le figg. 23 e 24 illustrano gli eleganti esperimenti che possono essere eseguiti quando si vedono direttamente le sinapsi. Questi esperimenti ricordano quelli sulla sinapsi neuromuscolare, ma forniscono una dimostrazione ancor più evidente della modalità con cui l'acetilcolina applicata elettroforeticamente può riprodurre l'eccitazione dovuta a una sinapsi colinergica. Nella prima colonna della fig. 23 sono riportate le risposte prodotte dall'applicazione di 3 diverse quantità di acetilcolina. I tracciati superiori sono le risposte intracellulari, quelli inferiori gli impulsi di corrente che espellono ACh. In C, l'ACh era insufficiente a generare una scarica come in B. Con una quantità ancora maggiore si verificava in A la stessa scarica, ma seguita da una grande depolarizzazione. La seconda colonna riporta le risposte a un'attivazione del nervo. D mostra la risposta in condizioni di riposo e il potenziale d'azione con una successiva ampia depolarizzazione, come quello che si osserva dopo la più grande applicazione elettroforetica (A). Con la stimolazione ripetitiva del nervo si osserva un progressivo declino della risposta, come è evidente in E, e alla fine (F) l'impulso viene a mancare. Appare chiara la stretta corrispondenza tra le due serie. La risposta evocata elettroforeticamente concorda con quella evocata dalla stimolazione del nervo, quando si applica una quantità di acetilcolina di circa 10 volte maggiore.
La fig. 24 fornisce un'eccellente illustrazione delle potenzialità di questo preparato. Tutti i tracciati sono registrazioni intracellulari, e quelli più in basso ci danno, a un potenziale di membrana normale, le risposte evocate da un'applicazione elettroforetica di ACh e da una stimolazione del nervo, ridotta come nella fig. 23F così da dare solo un PPSE. Nel resto della serie il potenziale di membrana è stato cambiato tramite il passaggio verso l'esterno di una corrente costante di intensità progressivamente crescente. Sulle ordinate è riportato il potenziale di membrana. Si può osservare che i potenziali prodotti dall'applicazione di ACh, sia elettroforetica sia nervosa, sono stati dapprima ridotti, poi aboliti, quindi invertiti. Vi è un notevole parallelismo a tutti i livelli di potenziale di membrana e, in tutt'e due i casi, l'inversione è a −2 mV.
Bibliografia.
Baker, P.F., Hodgkin, A.L., Ridgway, E.P., Two phases of calcium entry during the action potential in giant axons of Loligo, in ‟Journal of physiology", 1970, CCVIII, pp. 80P-82P.
Birks, R., Huxley, H. E., Katz, B., The fine structure of the neuromuscular junction of the frog, in ‟Journal of physiology", 1960, CL, pp. 134-144.
Braun, M., Schmidt, R. F., Zimmermann, M., Facilitation at the frog neuromuscular junction during and after repetitive stimulation, in ‟Pflügers Archiv", 1966, CCLXXXVII, pp. 41-55.
Brooks, V. B., An intracellular study of the action of repetitive nerve volleys and of Botulinum toxin on miniature end-plate potentials, in ‟Journal of physiology", 1956, CXXXIV, pp. 264-277.
Brown, G. L., Transmission at nerve endings by acetylcholine, in ‟Physiological reviews", 1937, XVII, pp. 485-513.
Castillo, J. del, Katz, B., Quantal components of the end-plate potential, in ‟Journal of physiology", 1954, CXXIV, pp. 560-573.
Castillo, J. del, Katz, B., The membrane change produced by neuromuscular transmitter, in ‟Journal of physiology", 1954, CXXV, pp. 546-565.
Castillo, J. del, Katz, B., On the localization of acetylcholine receptors, in ‟Journal of physiology", 1955, CXXVIII, pp. 157-181.
Chandler, W. K., Meeves, H., Voltage clamp experiments on internally perfused giant axons, in ‟Journal of physiology", 1965, CLXXX, pp. 788-820.
Couteaux, R., Morphological and cytochemical observations on the postsynaptic membrane at motor end-plates and ganglionic synapses, in ‟Experimental cell research", 1958, V, pp. 294-322.
Dale, H. H., Transmission of nervous effects by acetylcholine, in ‟Harvey lectures", 1937, XXXII, pp. 229-245.
Dale, H. H., Acetylcholine as a chemical transmitter substance of the effects of nerve impulses, in ‟Mount Sinai journal of medicine", 1938, IV, pp. 401-429.
Dennis, M. J., Harris, A. J., Kuffler, S. W., Synaptic transmission and its duplication by focally applied acetylcholine in parasympathetic neurons in the heart of the frog, in ‟Proceedings of the Royal Society" (Series B), 1971, CLXXVII, pp. 509-539.
Devanandan, M. S., Eccles, R. M., Westerman, R. A., Single motor units of mammalian muscle, in ‟Journal of physiology", 1965, CLXXVIII, pp. 359-367.
Dodge, F. A., Miledi, R., Rahamimoff, R., Strontium and quantal release of transmitter at the neuromuscular junction, in ‟Journal of physiology", 1969, CC, pp. 267-283.
Dodge, F. A., Rahamimoff, R., Co-operative action of calcium ions in transmitter release at the neuromuscular junction, in ‟Journal of physiology", 1967, CXCIII, pp. 419-432.
Eccles, J. C., Synaptic and neuromuscular transmission, in ‟Ergebnisse der Physiologie", 1936, XXXVIII, pp. 339-444.
Eccles, J. C., Synaptic and neuro-muscular transmission, in ‟Physiological reviews", 1937, XVII, pp. 538-555.
Eccles, J. C., The physiology of synapses, Berlin 1964.
Eccles, J. C., The ionic mechanisms of excitatory and inhibitory synaptic action, in ‟Annals of the New York Academy of Sciences", 1966, CXXXVII, pp. 473-495.
Eccles, J. C., Katz, B., Kuffler, S. W., Nature of the ‛end-plate potential' in curarized muscle, in ‟Journal of neurophysiology", 1941, IV, pp. 362-387.
Eccles, J. C., MacFarlane, W. V., Actions of anticholinesterases on end-plate potential of frog muscle, in ‟Journal of neurophysiology", 1949, XII, pp. 59-80.
Eccles, J. C., Sherrington, Ch. S., Numbers and contraction values of individual motor-units examined in some muscles of the limb, in ‟Proceedings of the Royal Society" (Series B), 1930, CVI, pp. 326-357.
Fatt, P., Katz, B., An analysis of the end-plate potential recorded with an intra-cellular electrode, in ‟Journal of physiology", 1951, CXV, pp. 320-370.
Fatt, P., Katz, B., Spontaneous subthreshold activity at motor nerve endings, in ‟Journal of physiology", 1952, CXVII, pp. 109-128.
Gage, P. W., Armstrong, C. M., Miniature end-plate currents in voltage clamped muscle fibres, in ‟Nature", 1968, CCXVIII, pp. 363-365.
Gage, P. W., McBurney, R. N., Miniature end-plate currents and potentials generated by quanta of acetylcholine in glycerol-treated toad sartorius fibres, in ‟Journal of physiology", 1972, CCXXVI, pp. 79-94.
Gage, P. W., Morre, J. W., Synaptic current at the squid giant synapse, in ‟Science", 1969, CLX, pp. 510-512.
Hagiwara, S., Tasaki, I., A study of the mechanism of impulse transmission across the giant synapse of the squid, in ‟Journal of physiology", 1958, CXLIII, pp. 114-137.
Hodgkin, A. L., The conduction of the nervous impulse, Liverpool 1964.
Hodgkin, A. L., Keynes, R. D., Movements of labelled calcium in squid giant axons, in ‟Journal of physiology", 1957, CXXXVIII, pp. 253-281.
Hubbard, J. I., Repetitive stimulation at the mammalian neuromuscular junction, and mobilization of transmitter, in ‟Journal of physiology", 1963, CLXIX, pp. 641-662.
Hubbard, J. I., Mechanisms of transmitter release, in ‟Progress in biophysics and molecular biology", 1970, XXI, pp. 33-124.
Hubbard, J. I., Microphysiology of Vertebrate neuromuscular transmission, in ‟Physiological reviews", 1973, LIII, pp. 674-723.
Hubbard, J. I., Schmidt, R. F., An electrophysiological investigation of mammalian motor nerve terminals, in ‟Journal of physiology", 1963, CLXVI, pp. 145-167.
Hubbard, J. I., Willis, W. D., Hyperpolarization of mammalian motor nerve terminals, in ‟Journal of physiology", 1962, CLXIII, pp. 115-137.
Katz, B., Microphysiology of the neuro-muscular junction. A physiological ‛quantum of action' at the myoneuronal junction, in ‟Bulletin of the Johns Hopkns Hospital", 1958, CII, pp. 275-295.
Katz, B., The transmission of impulses from nerve to muscle, and the subcellular unit of synaptic action, in ‟Proceedings of the Royal Society" (Series B), 1962, CLV, pp. 455-479.
Katz, B., Nerve, muscle and synapse, New York 1966.
Katz, B., The release of neural transmitter substances, Liverpool 1969.
Katz, B., Miledi, R., Propagation of electric activity in motor nerve terminals, in ‟Proceedings of the Royal Society" (Series B), 1965, CLXI, pp. 439-468.
Katz, B., Miledi, R., Measurement of synaptic delay and time course of acetylcholine release at neuromuscular junction, in ‟Proceedings of the Royal Society" (Series B), 1965, CLXI, pp. 469-481.
Katz, B., Miledi, R., The effect of calcium on acetylcholine release from motor nerve terminals, in ‟Proceedings of the Royal Society" (Series B), 1965, CLXI, pp. 482-489.
Katz, B., Miledi, R., Input-output relation of a single synapse, in ‟Nature", 1966, CCXII, pp. 1242-1245.
Katz, B., Miledi, R., A study of synaptic transmission in the absence of nerve impulses, in ‟Journal of physiology", 1967, CXCII, pp. 407-436.
Katz, B., Miledi, R., The timing of calcium action during neuromuscular transmission, in ‟Journal of physiology", 1967, CLXXXIX, pp. 535-544.
Katz, B., Miledi, R., Tetrodotoxin and neuromuscular transmission, in ‟Proceedings of the Royal Society" (Series B), 1967, CLXVII, pp. 8-22.
Katz, B., Miledi, R., The effect of local blockage of motor nerve terminals, in ‟Journal of physiology", 1968, CXCIX, pp. 729-741.
Katz, B., Miledi, R., The role of calcium in neuromuscular facilitation, in ‟Journal of physiology", 1968, CXCV, pp. 481-492.
Katz, B., Miledi, R., Tetrodotoxin-resistanc electric activity in presynaptic terminals, in ‟Journal of physiology", 1969, CCIII, pp. 459-487.
Katz, B., Miledi, R., The statistical nature of acetylcholine potential and its molecular components, in ‟Journal of physiology", 1972, CCXXIV, pp. 665-699.
Katz, B., Miledi, R., The binding of acetylcholine to receptors and its removal from the synaptic cleft, in ‟Journal of physiology", 1973, CCXXXI, pp. 549-574.
Krnjević, K., Chemical nature of synaptic transmission in Vertebrates, in ‟Physiological reviews", 1974, LIV, pp. 418-540.
Krnjević, K., Miledi, R., Acetylcholine in mammalian neuromuscular transmission, in ‟Nature", 1958, CLXXXII, pp. 805-806.
Kuffler, S. W., Electric potential changes at an isolated nerve-muscle junction, in ‟Journal of neurophysiology", 1942, V, pp. 18-26.
Kuffler, S. W., Specific excitability of the endplate region in normal and denervated muscles, in ‟Journal of neurophysiology", 1943, VI, pp. 99-110.
Kuffler, S. W., Physiology of neuro-muscular junctions: electrical aspects, in ‟Federation proceedings", 1948, VII, pp. 437-446.
Kuffler, S. W., Transmitter mechanism at the nerve-muscle junction, in ‟Archives des sciences physiologiques", 1949, III, pp. 585-601.
Kuffler, S. W., Yoshikami, D., The distribution of acetylcholine sensitivity at the postsynaptic membrane of vertebral skeletal twitch muscles: iontohoretic mapping in the micron range, in ‟Journal of physiology", 1975, CCXLIV, pp. 703-730.
Kuffler, S. W., Yoshikami, D., The number of transmitter molecules in a quantum: an estimate from iontophoretic application of acetylcholine at the neuromuscular synapse, in ‟Journal of physiology", 1975, CCLI, pp. 465-482.
Liley, A. W., The quantal components of the mammalian end-plate potential, in ‟Journal of physiology", 1956, CXXXIII, pp. 571-587.
Liley, A.W., North, K. A. K., An electrical investigation of effects of repetitive stimulation on mammalian neuromuscular junction, in ‟Journal of neurophysiology", 1953, XVI, pp. 509-527.
Loewi, O., Über humorale Übertragbarkeit der Herznervenwirkung, in ‟Pflügers Archiv", 1921, CLXXXIX, pp. 239-242; CXCIII, pp. 201-213.
Loewi, O., Problems connected with the principle of humoral transmission of nervous impulses, in ‟Proceedings of the Royal Society" (Series B), 1933, CXVIII, pp. 299-316.
McMahan, U. J., Kuffler, S. W., Viewing cholinergic synaptic areas on living nerve cells of the frog's heart, in Excitatory synaptic mechanisms (a cura di P. ndersen e J. K. S. Jansen), Oslo 1969, pp. 57-66.
Maeno, T., Analysis of sodium and potassium conductances in the procaine end-plate potential, in ‟Journal of physiology", 1966, CLXXXIII, pp. 592-606.
Mallart, A., Martin, A. R., An analysis of facilitation o transmitter release at the neuromuscular junction of the frog, in ‟Journal of physiology", 1967, CXCIII, pp. 679-694.
Miledi, R., Spontaneous synaptic potentials and quantal release of transmitter in the stellate ganglion of the squid, in ‟Journal of physiology", 1967, CXCII, pp. 379-406.
Miledi, R., Transmitter action in the giant synapse of the squid, in ‟Nature", 1969, CCXXIII, pp. 1284-1286.
Miledi, R., Slater, C. R., The action of calcium on neuronal synapses in the squid, in ‟Journal of physiology", 1966, CLXXXIV, pp. 473-498.
Otsuka, M., Endo, M., Nonomura, Y., Presynaptic nature of neuromuscular depression, in ‟Japanese journal of physiology", 1962, XII, pp. 573-584.
Peper, K., McMahan, U. J., Distribution of acetylcholine receptors in the vicinity of nerve terminals on skeletal muscle of the frog, in ‟Proceedings of the Royal Society" (Series B), 1972, CLXXXI, pp. 431-440.
Takeuchi, A., Takeuchi, N., Active phase of frog's end-plate potential, in ‟Journal of neurophysiology", 1959, XXII, pp. 395-411.
Takeuchi, A., Takeuchi, N., On the permeability of the end-plate membrane during the action of transmitter, in ‟Journal of physiology", 1960, CLIV, pp. 52-67.
Takeuchi, A., Takeuchi, N., Electrical changes in pre- and post-synaptic axons of the giant synapse of Loligo, in ‟Journal of general physiology", 1962, XLV, pp. 1181-1193.
Farmacologia della sinapsi neuromuscolare
SOMMARIO: 1. Introduzione: a) sostanze attive sulla sinapsi neuromuscolare; b) il periodo iniziale. □ 2. Il periodo moderno: a) prima fase; b) trasmissione chimica; c) progressi nella chimica dei farmaci; d) progressi nel dosaggio biologico; e) l'impiego clinico della tubocurarina; f) lo sviluppo di curari sintetici; g) gli agenti bloccanti per depolarizzazione. □ 3. Meccanismi di blocco neuromuscolare: a) blocco competitivo; b) blocco per depolarizzazione; c) effetti presinaptici; d) il problema del blocco duplice e fenomeni relativi; e) la miastenia grave e altri stati anormali della funzione neuromuscolare; f) farmacogenetica del sussametonio (succinilcolina). □ 4. Antagonismo del blocco neuromuscolare. □ 5. La sinapsi neuromuscolare per gli studi sul recettore: a) tipi di sinapsi neuromuscolare e specificità di azione; b) relazioni fra struttura e funzione; c) studi funzionali sul recettore; d) studi di permeabilità al livello della giunzione neuromuscolare; e) studi biochimici sul recettore. □ 6. I più recenti sviluppi. □ Bibliografia.
1. Introduzione.
a) Sostanze attive sulla sinapsi neuromuscolare.
I veleni naturali hanno da sempre colpito la fantasia dell'uomo, e il magico fascino che essi esercitano diviene tanto più profondo nel caso di un veleno come il curaro, capace di paralizzare il movimento senza interferire con il pensiero e la sensibilità. Questa è la ragione del posto che il curaro si è conquistato nei romanzi d'avventure, in cui la vittima si abbatte esanime anche se solo scalfita da una freccia avvelenata, ed è la ragione anche del ruolo più serio che ha avuto contribuendo a formare quel timore reverenziale nei confronti del fisiologo, che nell'ultimo secolo favorì lo sviluppo dei movimenti contro la vivisezione. Ma questa stessa selettiva attività paralizzante ha anche creato il campo di studio della trasmissione neuromuscolare, ha contribuito a rivoluzionare la pratica anestesiologica e rappresenta uno degli approcci più fecondi per lo studio delle basi molecolari dell'azione farmacologica.
b) Il periodo iniziale.
La storia della scoperta del veleno e del suo uso, e delle piante da cui deriva, è trattata nelle rassegne di Mclntyre (v., 1947) e di Bovet e altri (v., 1959). È ancora interessante la lettura del resoconto di Waterton, del 1825, di come fu dimostrato per la prima volta che un asino, dopo somministrazione di una dose naturalmente letale di curaro, poteva essere salvato dalla respirazione artificiale - prova della selettività dell'azione del curaro e anticipazione di quanto quotidianamente avviene nelle moderne camere operatorie.
‟Un'asina ricevette il veleno wourali nella spalla e dopo dieci minuti morì. Le fu allora praticata un'incisione nella trachea e per questo tramite i polmoni le vennero regolarmente insufflati con mantici per due ore. L'animazione sospesa, allora, ritornò: l'animale sollevò la testa e guardò intorno. Ma non appena l'insufflazione fu interrotta, l'animale ripiombò nella morte apparente. La respirazione artificiale fu immediatamente ripresa, e proseguita senza interruzione per altre due ore. Questo salvò l'asina dalla morte definitiva; essa si rialzò e camminò, e non sembrò nè agitata nè sofferente. La ferita attraverso cui era stato introdotto il veleno si rimarginò senza difficoltà. Il suo stato generale, tuttavia, fu compromesso così seriamente che per lungo tempo si dubitò che essa potesse mai riprendersi del tutto. Essa apparve denutrita e malaticcia per più di un anno, ma cominciò a migliorare la primavera seguente, e verso la metà dell'estate era diventata grassa e vivace.
Il lettore compassionevole si rallegri apprendendo che il conte Percy, commosso dalla sua disgrazia, la mandò da Londra a Walton Hall, vicino a Wakefield, dove tuttora vive con il nome di Wouralia. Wouralia sarà protetta dal maltempo invernale, e in estate si nutrirà sul pascolo migliore. Non sarà gravata di pesi e finirà i suoi giorni in pace (C. Waterton, Wanderings in South America, London 1825).
Ugualmente fertili furono gli studi sul curaro di Claude Bernard (v., 1857). La sua opera, nel complesso, non portò al quadro semplice e chiaro presentato di solito - probabilmente perché la preparazione da lui usata conteneva, come prevedibile, molte altre sostanze con attività diverse da quella specifica del curaro. Pure, è rimasta classica la sua dimostrazione che in presenza di curaro il muscolo striato mantiene la sua eccitabilità elettrica, e che le fibre sensitive, il midollo spinale e le fibre motrici continuano a condurre. Meno spesso citata, ma ugualmente importante, fu la sua osservazione che il curaro paralizza l'effetto della stimolazione vagale sul cuore - probabilmente la prima dimostrazione di blocco gangliare, e una delle prime indicazioni, destinata ad aspettare lungo tempo la completa verifica, che le sinapsi gangliare e neuromuscolare hanno basi biochimiche comuni.
Non molto più tardi si aggiungevano a queste prime dimostrazioni dell'azione selettiva sulla trasmissione sinaptica i notevoli studi di Crum Brown e Fraser (v., 1869), che mostrarono come varie ammine con azioni diverse, quali la morfina, l'atropina e la stricnina, perdevano, se rese quaternarie con ioduro di metile, le loro attività originarie e acquistavano un'unica azione comune a tutte - quella curarizzante. Il fatto che composti contenenti azoto quaternario fossero curaro-simili rappresentò il primo esempio, e forse ancora il migliore, di correlazione fra struttura chimica e azione farmacologica. Nel corso di questa ricerca, gli stessi autori descrissero saggi biologici dell'azione curarica, compreso il metodo della caduta della testa nel coniglio, e sulla base della loro teoria realizzarono il primo curaro sintetico, il tetrametilammonio.
A questo primo periodo appartengono anche i pregevoli studi di J. N. Langley (v., 1907-1909) sull'antagonismo fra nicotina e curaro al sito delle terminazioni nervose motrici nel muscolo di rana. Partendo dall'osservazione dell'effetto stimolante della nicotina nella regione della placca terminale, che persisteva anche dopo denervazione, Langley avanzò l'idea di una ‛sostanza recettiva' capace di reagire con i farmaci secondo le normali leggi delle reazioni chimiche. Questo concetto fu poi sviluppato da P. Ehrlich (v., 1914) nella sua nuova concezione del recettore come base della chemioterapia. Ehrlich stesso affermò ‟Furono, in particolare, le brillanti ricerche di Langley sugli effetti degli alcaloidi a cancellare i miei dubbi e a farmi apparire probabile l'esistenza dei chemorecettori". Analogamente, Langley caratterizzò qualitativamente le proprietà principali dell'antagonismo dei farmaci nella regione della placca motrice. È interessante ricordare che A. V. Hill (v., 1910), tentando, sulla scorta delle ricerche sulla nicotina, di descrivere in termini quantitativi le contratture che l'azione della nicotina causava nel muscolo di rana, fu il primo a formulare matematicamente l'equazione - ora di uso generale - dell'azione di massa per l'interazione farmaco-agonista. Il tentativo fu infruttuoso per una ragione tuttora valida: e cioè che non è possibile trarre conclusioni sull'interazione farmaco-recettore solo in base alla risposta meccanica a una sostanza stimolante, perché la risposta registrata rappresenta il risultato finale di una catena di eventi di cui l'interazione farmaco-recettore è solo il primo. Fu necessario attendere fino a che J. H. Gaddum (v., 1937) formulò le equazioni appropriate per l'interazione tra sostanza stimolante e antagonista, prima che la strada indicata da Hill portasse al successo.
Infine, bisogna ricordare che Brodie già nel 1811 aveva suggerito l'uso terapeutico del curaro nella cura del tetano, dopo aver dimostrato che la respirazione artificiale resuscitava gli animali curarizzati (v. Mclntyre, 1947). Il farmaco fu sperimentato nel tetano umano nel 1858, e non molto tempo dopo per l'epilessia, la corea e la rabbia. Maggior impulso si ebbe dopo la dimostrazione, fornita da F. Bremer e J. Titeca nel 1927, dell'aumentato effetto rilassante del curaro su muscoli stimolati ripetutamente e del suo effetto sulla rigidità da decerebrazione, e dopo che R. West (v., 1935), studiando la tetania paratireopriva, ebbe osservato che alcune preparazioni di curaro avevano un'azione che permetteva il rilassamento selettivo di stati spastici.
2. Il periodo moderno.
a) Prima fase.
Intorno agli anni trenta, quindi, lo stato delle conoscenze era il seguente: 1) si sapeva che il curaro aveva un'azione selettiva su una sostanza recettiva postsinaptica nella sinapsi neuromuscolare; 2) era stata identificata la principale caratteristica della struttura chimica delle sostanze curaro-simili, quella di essere composti con azoto quaternario (composti ‛onio'); 3) il curaro era stato di fatto sperimentato in varie circostanze cliniche. Eppure esso era ancora considerato come una curiosità. Il passaggio alla fase moderna è dato da tre sviluppi: la formulazione di una teoria della trasmissione chimica; i progressi nella chimica dei farmaci; i progressi nei dosaggi biologici.
b) Trasmissione chimica.
Per quanto fosse nota da lungo tempo la capacità del curaro di provocare il blocco neuromuscolare, il reale significato di questo fenomeno non fu compreso per molti anni a causa della teoria dell'isocronismo di L. Lapicque (1908), secondo la quale tale effetto del farmaco era dovuto alla sua interferenza con la relazione fra le cronassie della terminazione nervosa e della fibra muscolare. Poiché tale teoria si fondava sulle proprietà ‛elettriche' dei tessuti eccitabili, si perdeva di vista la reale chiave interpretativa che era invece basata sulla caratteristica struttura ‛chimica' del curaro. Inoltre, per quanto questa teoria fosse stata pesantemente criticata, specialmente da W. A. H. Rushton (v., 1933), progressi razionali nell'uso del curaro poterono verificarsi solo quando si sviluppò una nuova e valida spiegazione dell'azione del farmaco, cioè la teoria della trasmissione chimica; anzi, il curaro ebbe un ruolo importante nel far sì che questa teoria si affermasse. La prova più importante fu che una sostanza mediatrice era liberata in seguito alla stimolazione di un nervo motore (v. fig. 1), che essa si identificava con l'acetilcolina, e che l'acetilcolina poteva riprodurre i fenomeni della trasmissione nervosa. Fu tuttavia importante anche mostrare che quelle procedure capaci di modificare la trasmissione trovavano una spiegazione razionale. Così le azioni degli anticolinesterasici e del curaro costituirono parte importante di queste ricerche. Si dimostrò da un lato che le colinesterasi prolungano l'azione dell'acetilcolina, dando in tal modo origine a un effetto prolungato del mediatore sulla placca terminale e quindi a una scarica ripetitiva dello stimolo, ‛un tetano breve e decrescente' in risposta a una singola eccitazione; dall'altro che il curaro diminuisce la risposta all'iniezione di acetilcolina ma non interferisce con la liberazione del mediatore (v. fig. 2). Il parallelismo fra questi effetti farmacologici e i reali effetti sulla trasmissione dimostrò che il fenomeno della liberazione di acetilcolina non era una semplice evenienza, ma l'evento necessario anche per il normale processo di trasmissione. L'ulteriore analisi della trasmissione neuromuscolare condotta con metodi elettrofisiologici, resa possibile dalla rivoluzione nelle conoscenze sulla fisiologia della membrana e dalla scoperta della liberazione quantica del mediatore (v. sinapsi: Fisiologia della sinapsi periferica; v. neurone e impulso nervoso), ha ampiamente confermato la teoria della trasmissione chimica in tutti questi aspetti.
L'identificazione del mediatore con l'acetilcolina ebbe anche un altro significato: poiché l'acetilcolina è essa stessa un composto con azoto quaternario, in cui l'azoto è sostituito con gruppi metilici, si ipotizzò immediatamente che si sarebbe potuto interferire nel processo della trasmissione chimicamente per mezzo di sostanze che, contenendo azoto quaternario, specie se metilato, potessero imitare il mediatore naturale e competere con esso. Il fatto che la tubocurarina fosse una sostanza di questo tipo, come pure le generalizzazioni fatte da Crum Brown e Fraser 60 anni prima, si rivelarono allora nel loro pieno significato.
c) Progressi nella chimica dei farmaci.
Una delle difficoltà nello studio farmacologico del curaro era stata la mancanza di una sostanza chimica pura e ben caratterizzata con cui eseguire ricerche fisiologiche e farmacologiche. Un importante passo avanti fu quello compiuto da H. King (v., 1935), il quale stabilì le principali caratteristiche della struttura della tubocurarina (v. fig. 3). La sua caratterizzazione in forma cristallina pura costituì la prima purificazione di un composto curaro-simile ad alta e specifica attività, e stimolò una quantità enorme di lavoro. Una difficoltà immediata era data dalla scarsa disponibilità di tubocurarina (così detta perché conservata in tubi fatti con canne di bambù). Questo fattore limitante fu eliminato quando O. Wintersteiner e J. D. Dutcher (v., 1943) scoprirono un arbusto amazzonico, Chondrodendron tomentosum, da cui poteva essere isolata la tubocurarina. La produzione di tubocurarina divenne allora un fatto commerciale, e l'antico metodo di rifornirsi dagli Indios del Brasile di veleno da frecce raccolto in vasi, zucche vuote e canne fu soppiantato da un rifornimento in larga scala (con trasporto in barattoli e altri contenitori più moderni). Venne anche stimolata la sintesi chimica, e più avanti discuteremo alcuni degli sviluppi in questo senso.
Il valore del risultato ottenuto da King in un campo ancora molto confuso è di poco sminuito dalla recente scoperta del suo errore nel considerare la (+)-tubocurarina un composto bis-quaternario. In realtà, uno dei due atomi di azoto della molecola è terziario (v. Everett e altri, 1970). L'errore fu scoperto nel tentativo di sintetizzare tubocurarina marcata con il 14C, e fu probabilmente originato, nel corso del lavoro di King, dall'uso di derivati metilati. A ogni modo King determinò correttamente la struttura di base e la posizione dei gruppi fenolico e metossilico.
Importante, da un punto di vista più generale, fu il lavoro di Ing e collaboratori (v. Ing, 1936) sui composti quaternari, che ne mise in rilievo la natura ionica (paragonabile a quella dei cationi metallici) e l'importanza della sostituzione metilica. Esso mostrò anche che la classe generale dei composti ‛onici' - formati da fosforo, arsenico e zolfo, oltre che da azoto - in cui un atomo può formare un altro legame accettando una carica positiva, possedeva nel suo insieme un'azione curaro-simile.
d) Progressi nel dosaggio biologico.
Ben presto si presentò un'altra difficoltà, prima che una sostanza chimica pura e standardizzata fosse disponibile per esperimenti e impieghi clinici. Infatti si scoprì che gli estratti di tubocurarina non erano facilmente purificabili e che, fra l'altro, potevano contenere un etere dimetilico con attività farmacologica considerevolmente maggiore, attività che variava a seconda della specie su cui si conduceva l'esperimento. Le farmacopee dovettero perciò ideare dei procedimenti di controllo in modo da garantire che i campioni di tubocurarina non fossero contaminati da quantità significative del composto più attivo. Questa esigenza dette origine a numerose procedure di dosaggio biologico, che vennero anche usate per verificare l'attività di preparazioni meno pure. Fra questi metodi ricordiamo la preparazione del diaframma isolato di ratto in vitro (v. Bùlbring, 1946) e il metodo in vivo di Holaday della caduta della testa del coniglio (v. Varney e altri, 1949), in cui la sostanza è infusa nella vena auricolare di un coniglio finché l'indebolimento dei muscoli del collo provoca il rilassamento del collo stesso (è interessante notare che Crum Brown e Fraser avevano già descritto il fenomeno 70 anni prima). Con queste procedure si riuscì a stabilire l'attività biologica dei composti entro limiti di errore accettabili ai fini clinici.
Si deve aggiungere che, appena fu scoperto che l'attività relativa delle sostanze curarizzanti variava nelle diverse specie, divenne importante provare queste sostanze nell'uomo. Per molte di esse il primo controllo fu fatto in soggetti anestetizzati sottoposti a operazione chirurgica, ma spesso i ricercatori sperimentarono la sostanza su se stessi. Fra questi studi ricordiamo quelli di Prescott e altri (v., 1946) sulla tubocurarina, di Marsh e altri (v., 1948) su tubocurarina e alcuni analoghi, di Organe e altri (v., 1949) sul decametonio e di Unna e altri (v., 1950) sulla tubocurarina, il suo etere metilico e il decametonio. A questo piccolo gruppo di autocurarizzati si deve aggiungere il professor E. A. Pask, che si fece anestetizzare, curarizzare e gettare in acqua, per collaudare una veste galleggiante atta a sostenere un uomo privo di coscienza. Il più eminente soggetto di questa serie è, in ogni modo, S. M. Smith (v. Smith e altri, 1947), che assunse una dose di tubocurarina tre volte superiore a quella sufficiente a produrre paralisi, per verificare l'assenza di effetti centrali della sostanza.
e) L'impiego clinico della tubocurarina.
A prima vista l'uso di una sostanza in grado di provocare paralisi respiratoria per facilitare le operazioni chirurgiche sembrò paradossale; vale quindi la pena di soffermarsi un momento su questo argomento. Una delle difficoltà che il chirurgo deve affrontare, specie nelle operazioni addominali, è quella di ottenere un'esposizione soddisfacente del campo operatorio. Se i muscoli addominali sono contratti, c'è una forte tendenza del contenuto intestinale a erniare attraverso l'apertura operata nell'addome, e anche se questo non succede, il tono della parete addominale, accresciuto dai riflessi nocicettivi conseguenti agli stimoli operatori, può rendere molto difficile l'accesso. Conseguentemente gli anestesisti erano giunti alla pratica di anestesie piuttosto profonde per i grandi interventi sull'addome, come quelli sulla cistifellea, e questo comportava i rischi degli effetti indesiderabili dell'anestesia profonda combinati con quelli della depressione della respirazione. Spesso ne conseguivano collasso polmonare e broncopolmonite. Col curaro, invece, poteva essere usata un'anestesia molto più leggera (e meno tossica), ottenendo al contempo un maggiore rilassamento muscolare. Questa tecnica richiedeva che gli anestesisti sviluppassero, divenendone esperti, metodi di valida ventilazione artificiale: di fatto, essa rappresentò un efficace stimolo a sviluppare tali metodi. L'effetto di rilassamento addominale può essere veramente imponente, e con la perdita del tono dei muscoli laterali dell'addome l'intero contenuto addominale sembra offrirsi esposto all'accesso del chirurgo. La tecnica fu usata per la prima volta da Griffith e Johnson (v., 1942) e quasi contemporaneamente da Cullen (v., 1943) e si affermò rapidamente, divenendo una normale pratica anestesiologica.
Circa nello stesso periodo, il curaro fu introdotto nell'uso clinico anche in un ambito totalmente differente, e cioè per diminuire la violenza delle convulsioni durante l'elettroshock-terapia nelle malattie mentali. L'elettroshock si era gradualmente affermato come il metodo migliore per indurre queste convulsioni terapeutiche, la cui violenza, tuttavia, provocava un considerevole danno, particolarmente a causa della contrazione dei retti addominali, che dava luogo a fratture da compressione della colonna vertebrale. Inoltre, le energiche contrazioni muscolari potevano provocare un aumento notevole della pressione venosa, con conseguente pericolo di emorragie. Seguendo un suggerimento di MacIntyre, Bennett (v., 1941) mostrò che le convulsioni potevano essere sostanzialmente attenuate dalla tubocurarina, senza interferire con il loro effetto terapeutico (come era da aspettarsi, dato che da tempo era noto, e ripetutamente era stato confermato, che la tubocurarina quasi non passava nel liquido cefalorachidiano o nella sostanza cerebrale, cosicché non avrebbe dovuto interferire con i fenomeni neurofisiologici ed elettrici che vi hanno luogo). La tubocurarina non è il rilassante ideale a questo scopo, poiché, mentre le convulsioni durano solo per un breve periodo, la sua azione può durare per mezz'ora o più, e pertanto permane il rischio del collasso respiratorio anche quando non è più necessaria l'azione del farmaco. Comunque questa pratica è divenuta normale in clinica psichiatrica, ed è stata notevolmente facilitata dall'uso di rilassanti ad azione più rapida.
f) Lo sviluppo di curari sintetici.
La determinazione della struttura della tubocurarina da parte di King indirizzò Bovet e i suoi collaboratori verso un affascinante studio degli analoghi sintetici (v. Bovet, 1951). In breve, il metodo di Bovet consistette nello staccare pezzi successivi della molecola. La fig. 3 mostra alcune delle sostanze da lui sperimentate in successione e la progressiva semplificazione della struttura. Queste ricerche illustrano bene una delle caratteristiche della farmacologia di questo campo e cioè che se una sostanza è correlata con l'acetilcolina per un aspetto, per esempio la sua capacità di competere con questa a livello della sinapsi neuromuscolare, probabilmente possiede anche altre azioni correlate all'acetilcolina. La ricerca deve perciò essere sempre accompagnata da indagini sull'attività muscarinica, atropinica, anticolinesterasica e ganglioplegica della sostanza, oltre che sulla sua attività in assoluto. Deve essere ricercata anche una possibile attività sulla liberazione di istamina, che talvolta è presente. La sostanza scelta in definitiva per gli studi clinici fu, più per ragioni di specificità di azione che di potenza, la gallammina (Flaxedil). Questa sostanza si rivelò un efficace sostituto della tubocurarina, meno attivo, ma anche più specifico, poiché non possedeva gli effetti ganglioplegici e di liberazione d'istamina dell'alcaloide naturale. Essa aveva anche un'interessante proprietà peculiare, quella di paralizzare l'effetto del vago sul ritmo cardiaco (v. Riker e Wescoe, 1951) producendo una tachicardia specifica, ma non sembrava possedere altre azioni atropino-simili. Le ricerche di Bovet sono state le prime di una serie che ha permesso di individuare un'amplissima gamma di composti curaro-simili, che sono stati estesamente trattati in articoli più o meno recenti (v. Bovet e altri, 1959; v. Cavallito e Gray, 1960).
g) Gli agenti bloccanti per depolarizzazione.
Un fattore nuovo fu apportato dall'avvento del decametonio. Questa sostanza (v. fig. 3), che può essere considerata quasi come il caso limite della semplificazione di Bovet della molecola della tubocurarina, fu scoperta nel 1948 da due gruppi di ricercatori, indipendentemente l'uno dall'altro. Paton e Zaimis (v., 1949), usando metodi di saggio su gatto, coniglio, ratto e topo, trovarono che essa era il membro più attivo della serie omologa di composti metonici, nell'ambito di una ricerca iniziata al fine di studiare i rapporti fra struttura e attività di composti dibasici che determinavano la liberazione di istamina. Barlow e Ing (v., 1949), usando i metodi del diaframma di ratto e della caduta della testa del coniglio, trovarono anch'essi che era il membro più attivo di varie serie bisquaternarie, in cui erano stati studiati diversi nuclei azotati e semplici gruppi bis-trimetilammonici separati da una catena alifatica. Queste ricerche mostrarono che il decametonio, oltre a essere il più semplice di tutti i composti dibasici quaternari curarizzanti, era anche estremamente specifico (v. fig. 4): a parte una certa affinità per la colinesterasi, del resto trascurabile alle dosi paralizzanti, esso non ha virtualmente alcuna azione su altri sistemi, se si escludono le conseguenze derivanti dalla sua azione di blocco neuromuscolare.
Il decametonio si rivelò interessante e stimolante anche da un altro punto di vista, cioè in rapporto alla sua azione di blocco neuromuscolare, che all'inizio apparve contraddittoria (v. Paton e Zaimis, 1952). Il blocco neuromuscolare soddisfaceva tutti i criteri classici, vale a dire non interferiva con la conduzione nervosa e con la liberazione di acetilcolina, non aboliva l'eccitabilità del muscolo alla stimolazione diretta e antagonizzava gli effetti dell'iniezione di acetilcolina. Eppure per molti altri aspetti sembrava l'antitesi della tubocurarina. Esso provocava contrazioni transitorie non coordinate delle fibre muscolari (le cosiddette fascicolazioni) all'inizio della sua azione; la sensibilità a esso di specie differenti variava in maniera inversa rispetto a quella della tubocurarina; i muscoli bianchi erano più sensibili di quelli rossi, effetto opposto a quello della tubocurarina, ed era assente il caratteristico aumento del blocco con la stimolazione tetanica, messo in evidenza da Bremer nei suoi studi con il curaro (v. fig. 5); l'anestesia tendeva ad antagonizzare il suo effetto, mentre potenziava quello della tubocurarina; nel gatto gli anticolinesterasici erano inefficaci contro di esso, mentre erano eccellenti antagonisti della tubocurarina. Il fatto forse più difficile da spiegare era che, se veniva paragonata l'attività farmacologica della tubocurarina e del decametonio sullo stesso animale (gatto), si trovava che il decametonio diveniva meno attivo quando era somministrato subito dopo la ripresa dell'animale da una dose precedente di tubocurarina. Inoltre, si vide che il decametonio faceva contrarre i muscoli degli Anfibi, degli Uccelli (v. fig. 6) e quelli denervati dei Mammiferi, e che la tubocurarina antagonizzava questo effetto. In breve, il decametonio sembrava comportarsi, sotto molti punti di vista, più come un mediatore naturale che come un inibitore. Si poteva infatti ottenere uno spasmo del muscolo tibiale del gatto mediante iniezione endoarteriosa di decametonio, allo stesso modo che con l'acetilcolina.
Questa relazione fra agente bloccante e mediatore naturale divenne ancora più evidente con l'avvento della succinilcolina (sussametonio). Questa molecola, che può essere rappresentata come due molecole di acetilcolina legate mediante i gruppi acetilici terminali, fu scoperta da tre gruppi diversi, Bovet e collaboratori a Roma nel 1949, E. T. de Beer e i suoi colleghi della Burroughs Wellcome (v. Phillips, 1949) e J. Walker (v., 1950) del National Institute for Medical Research. Questa sostanza si affermò rapidamente nella pratica clinica, in quanto provvista di azione miorilassante di breve durata, dovuta al fatto di essere un doppio estere dell'acido succinico, attaccabile dalla colinesterasi aspecifica del sangue. In linea di principio si poteva perciò produrre, con una infusione o con iniezioni ripetute del farmaco, un grado determinato di rilassamento muscolare, che scompariva rapidamente appena se ne sospendeva la somministrazione. Il sussametonio si rivelò molto simile al decametonio anche per altre proprietà, mentre la principale differenza era quella di avere un'azione più breve e prolungabile da parte degli inibitori della colinesterasi, che normalmente hanno scarso effetto sull'azione del decametonio. Come nel caso del decametonio, sorse il problema del perché una sostanza così simile al mediatore naturale fosse capace di produrre blocco neuromuscolare.
3. Meccanismi di blocco neuromuscolare.
La scoperta di fattori che agivano evidentemente in modi diversi portò a un considerevole numero di studi sul meccanismo con cui essi producevano il loro effetto. Mentre inizialmente sembrava che ci fosse una distinzione abbastanza semplice fra blocco tubocurarino-simile o competitivo, da una parte, e blocco decametonio-simile o depolarizzante, dall'altra, divenne poi chiaro che la situazione era assai più complicata. Discuteremo perciò, qui di seguito, i vari processi che possono contribuire al blocco della trasmissione neuromuscolare (v. anche Cookson e Paton, 1969).
a) Blocco competitivo.
1. Caratteristiche generali. - L'idea, implicita nei primi studi sul curaro in rapporto alla trasmissione chimica, che tubocurarina e farmaci analoghi semplicemente competano con il mediatore naturale per i siti recettori sulla membrana postsinaptica era stata ampiamente confermata dagli studi successivi. Questo processo obbedirebbe a quelle che sono ora le ben note regole dell'antagonismo farmacologico, descritte inizialmente da J. H. Gaddum (v., 1937). Van Maanen, Jenkinson e altri effettuarono ricerche sulla capacità della tubocurarina di antagonizzare l'acetilcolina e stimolanti simili, e trovarono che il loro schema di antagonismo aveva le caratteristiche tipiche della competizione, e cioè: a) si ottenevano curve dose-risposta parallele in assenza e in presenza dell'antagonista; b) fu dimostrato che il rapporto di dose (vale a dire di quanto deve essere aumentata la dose di agonista in presenza dell'antagonista per ottenere lo stesso effetto fisiologico) è proporzionale alla concentrazione dell'antagonista. L'antagonismo fra tubocurarina e acetilcolina a livello della placca motrice si collocò così accanto ad altri antagonismi, come quelli fra atropina e acetilcolina o fra mepirammina e istamina nel muscolo liscio, o quello fra propranololo e isoprenalina nel cuore.
Un punto che ha dovuto esser chiarito è emerso prendendo in considerazione la cinetica dell'azione del curaro. Le equazioni dell'antagonismo farmacologico, così come erano state formulate originariamente, sembra che ammettano un completo equilibrio fra stimolante e antagonista. Pure, è assai poco probabile che un tale equilibrio si possa stabilire in condizioni di trasmissione normale, quando l'agonista è presente soltanto per un periodo di pochi millisecondi (la durata del potenziale di placca) o meno. Al principio pareva che vi fossero prove di una relativa stabilità del complesso sostanza attiva-antagonista. Per esempio, è facile riscontrare che l'effetto di un antagonista su una preparazione isolata neuromuscolare in vitro persiste per un tempo notevole dopo che la sostanza è allontanata dal liquido di perfusione. V'è, in effetti, una regola generale, valida per agenti bloccanti competitivi di vario tipo, ben illustrata nel caso della sinapsi neuromuscolare, e cioè che intensa attività e azione prolungata vanno di pari passo, per quanto si possano avere eccezioni dovute a degradazione metabolica o captazione selettiva. Questo è proprio quanto ci si aspetta dal punto di vista cinetico (v. Paton, 1961); dato che l'affinità dipende dal rapporto delle costanti cinetiche di associazione e di dissociazione del complesso sostanza-recettore, più lenta è la velocità di dissociazione (e perciò più persistente l'effetto), tanto più intensa sarà l'attività. Si era anche sperato di ottenere dai dati cinetici sull'insorgenza e la scomparsa dell'antagonismo una qualche stima delle costanti cinetiche di associazione e dissociazione della sostanza rispetto al recettore, ma è chiaro che barriere di diffusione nel muscolo complicano assai seriamente la situazione (v. Waud, 1968).
Su questa base occorre invece supporre che, mentre l'antagonista può arrivare a occupare i recettori con una concentrazione regolata dall'equilibrio, si verifica solo una lieve alterazione del suo legame quando viene applicato l'agonista. Questo processo, però, non produrrebbe il comportamento classico che si osserva nell'antagonismo farmacologico se non a condizione che la sostanza stimolante stessa debba occupare solo una frazione relativamente piccola dei recettori per produrre i suoi caratteristici effetti. Questo introduce il concetto dei ‛recettori di riserva', che fu sviluppato da Stephenson (v., 1956) e Nickerson (v., 1956) ed è ben dimostrato per le risposte del muscolo liscio. Esso è implicito nella definizione di Stephenson di ‛efficacia' di una sostanza stimolante: egli attribuì un'efficacia pari a 1 a sostanze che producevano una risposta del 50% con occupazione dei recettori del 100%, cosicché l'efficacia è il reciproco dell'occupazione al 50% dell'effetto massimo. Nel blocco competitivo, perciò, la situazione sarebbe quella di equilibrazione graduale dei recettori con la sostanza antagonista mentre questa diffonde dai capillari ai siti recettori, e di reazione rapida del mediatore (ad alta efficacia) con l'insieme dei recettori liberi durante l'attività neuromotoria; l'effetto risultante dipende dalla quantità di mediatore, dalle dimensioni dell'insieme di recettori e dall'efficienza dell'accoppiamento della risposta della membrana a un dato grado di attivazione del recettore.
2. Il fattore di sicurezza nella trasmissione neuromuscolare. - Questo quadro del blocco neuromuscolare competitivo fornisce un modello operativo soddisfacente. Gli anticolinesterasici, determinando la conservazione del mediatore, rendono l'azione del mediatore maggiore e più lunga, e permettono una più intensa attivazione dell'insieme dei recettori. La stimolazione ripetuta provoca un approfondimento del blocco, in quanto gli stimoli successivi riducono l'emissione di acetilcolina. I processi che aumentano la produzione del mediatore (come l'incremento ‛dopo' stimolazione ripetuta) o la diminuiscono (interferendo con la sintesi di acetilcolina) causano rispettivamente resistenza o sensibilità al blocco. Bisogna però usare cautela nell'interpretazione, perché il processo di trasmissione dispone di notevoli margini di sicurezza. A regimi elevati di eccitazione, viene liberato solo un terzo di mediatore rispetto ai bassi regimi, pur mantenendosi una trasmissione normale; ne consegue che, almeno a bassi livelli di attività, vi è un fattore di sicurezza di circà tre volte. Studi farmacologici (v. Paton e Waud, 1967) hanno mostrato che bisogna antagonizzare il mediatore circa 3-5 volte per cominciare a produrre blocco neuromuscolare e fino a 20 volte per paralizzare le sinapsi più resistenti. Il blocco neuromuscolare osservato in pratica è perciò solo un rivelatore incompleto di quanto avviene a livello delle placca terminale (v. fig. 7). Prima che appaia alcun segno di blocco, si deve immaginare che vi sia un'occupazione graduale dei recettori fino a ridurre il fattore di sicurezza al valore di soglia, dopo di che si instaura progressivamente il blocco. Se la curarizzazione è spinta oltre la paralisi completa, c'è un'ulteriore ma non osservabile occlusione dei recettori. Infine, anche dopo l'apparente recupero completo, ci sarà ancora una considerevole occupazione di recettori, che praticamente può essere rivelata con facilità dall'accresciuta sensibilità a un'ulteriore dose di agente bloccante.
b) Blocco per depolarizzazione.
1. Ineccitabilità elettrica. - È stato già ricordato che decametonio e succinilcolina mostrano di condividere molte proprietà con lo stesso mediatore; ci si chiese allora come queste sostanze riuscissero a provocare il blocco. Analizzando la questione nel gatto in vivo (v. Burns e Paton, 1951; v. Zaimis, 1951), fu osservato che queste sostanze depolarizzavano la placca motrice nello stesso modo dell'acetilcolina e producevano nel muscolo denervato un'analoga contrattura elettricamente silente. La differenza fondamentale era che mentre il mediatore naturale ha un effetto transitorio, la modificazione del potenziale di membrana provocata dagli agenti bloccanti depolarizzanti durava per minuti o più (v. fig. 8). La depolarizzazione della placca spiega, ovviamente, la fascicolazione iniziale dei muscoli, ma fu anche osservato che questa depolarizzazione determinava un cambiamento nelle proprietà elettriche della regione della placca, rendendola elettricamente meno eccitabile (v. fig. 9). A distanza dalla placca la fibra manteneva la sua normale eccitabilità, cosicché si generava un nuovo stato in cui i potenziali di placca venivano a trovarsi nel mezzo di una regione con relativa ineccitabilità elettrica. La placca veniva depolarizzata solo parzialmente e rimaneva sensibile allo stimolo chimico, cosicché i potenziali di placca potevano essere ancora registrati, anzi quelli registrati alla soglia di propagazione erano maggiori del normale, allo scopo di superare questa barriera di eccitabilità. Inoltre, c'erano notevoli analogie fra i cambiamenti prodotti nel muscolo di gatto dal decametonio o dalla succinilcolina, da una parte, e quelli osservati al semplice passaggio di corrente attraverso un catodo nella regione di placca, dall'altra. L'analogia era accresciuta dall'osservazione che il blocco veniva ridotto dal passaggio di corrente attraverso un anodo nella placca depolarizzata, trattamento questo che, come è noto, intensifica la paralisi da curaro. Questo tipo di blocco è perciò paragonabile alla ben nota ‛depressione catodica' di altri tessuti, dovuta in gran parte all'inattivazione della conduttanza del sodio prodotta dalla depolarizzazione. Ne risulterebbe un'interferenza con l'instaurarsi di un impulso propagato che spiegherebbe l'ineccitabilità della regione della placca, perché per propagarsi i potenziali di placca devono essere maggiori del normale, e il modo in cui la placca forma una barriera al passaggio di un impulso direttamente eccitato da un lato all'altro della regione della placca. Essa spiega anche perché non persistano le fascicolazioni iniziali: si può supporre che con una placca durevolmente depolarizzata verrebbe prodotto un centro di eccitazione continua, che porterebbe i muscoli a uno spasmo tetanico prolungato, ma se la regione nelle vicinanze della placca divenisse una zona di depressione catodica, la partenza di impulsi propagati sarebbe bruscamente interrotta.
Questa spiegazione dell'azione bloccante del decametonio, partendo dalla capacità di questa sostanza di imitare l'azione del mediatore e di farlo in maniera persistente, rende anche conto della notevole relazione inversa e del mutuo antagonismo fra blocco competitivo e depolarizzante. Rende conto inoltre della grande analogia fra il blocco prodotto da decametonio o sussametonio e quello che si osserva in presenza di anticolinesterasici. Bacq e Brown (v., 1937) avevano già molto tempo prima dimostrato che queste sostanze potenziavano contratture singole del muscolo di gatto eccitate indirettamente, ma che la stimolazione ripetitiva portava a un grado di blocco neuromuscolare tanto più alto quanto maggiori erano la frequenza e la durata della stimolazione. Ciò potrebbe ora essere interpretato come la conseguenza naturale di una depolarizzazione persistente prodotta dall'accumulo del mediatore non più distrutto con sufficiente rapidità dalla colinesterasi; un simile modello di depolarizzazione è stato realmente riscontrato.
Tuttavia, quando il meccanismo fu studiato in una serie più ampia di muscoli e in altri animali, e in particolare quando furono usati metodi in vitro, apparve subito chiaro che la relazione fra depolarizzazione sviluppata ed entità del blocco non era affatto semplice, e la semplice antitesi fra depolarizzazione e blocco competitivo divenne molto sfumata. Fu anche notato che ripetute somministrazioni di una sostanza depolarizzante potevano portare a un mutamento del carattere del blocco, che diveniva più tubocurarino-simile e più sensibile agli anticolinesterasici, con tetano decrescente e maggiore effetto sulla muscolatura rossa. Per di più fu subito notato che la depolarizzazione stessa tendeva a essere massima quando la sostanza era somministrata per la prima volta, e che la risposta diventava successivamente alquanto minore. Questo sollevò la questione di un terzo meccanismo in base al quale la regione della placca diveniva meno sensibile sia al mediatore sia a sostanze analoghe, somministrati in maniera continua.
2. Desensibilizzazione. - Il caso più evidente di desensibilizzazione è quello osservato nel muscolo di rana isolato in vitro (v. Thesleff, 1955). Se questo tessuto è esposto a decametonio, succinilcolina, nicotina, o alla stessa acetilcolina in presenza di anticolinesterasici, la depolarizzazione della placca diminuisce gradualmente e in circa 15 minuti il potenziale può tornare al valore normale. Nei muscoli di Mammifero in vitro si osserva un fenomeno analogo ma più modesto, e senza ritorno al potenziale normale di membrana. In vivo, l'effetto è meno evidente, ma anche in questo caso, per esempio durante una somministrazione continuata di succinilcolina, si può vedere svanire la depolarizzazione di placca. Non si sa bene fino a che punto questo sia dovuto a una riduzione della depolarizzazione di placca, poiché ora si sa anche che le sostanze depolarizzanti liberano potassio dai muscoli, e che questo, elevando il livello del potassio plasmatico, può ridurre il potenziale di membrana sotto l'elettrodo di riferimento. Ma è chiaro che la desensibilizzazione rappresenta un meccanismo importante che ricorda il blocco competitivo, in quanto la sensibilità del recettore è diminuita, ma si differenzia da esso sia perché si produce soltanto dopo una precedente depolarizzazione, sia perché è intensificato (invece che antagonizzato) da sostanze depolarizzanti. Il suo ruolo nello sviluppo della teoria del recettore sarà discusso più avanti.
c) Effetti presinaptici.
1. Metabolismo della colina. - La trasmissione neuromuscolare è molto vigorosa e per lungo tempo fu presa in poca considerazione la possibilità di un esaurimento delle scorte di mediatore, a parte il transitorio calo della sua produzione durante il tetano. L'emicolinio (HC3) diede il primo contributo a una visione più completa. Esso fu scoperto da Schueler (v., 1955) in una serie di emiacetali, e fu inizialmente presentato come un composto capace di produrre a basse dosi la morte per blocco neuromuscolare (azione, questa, antagonizzata dalla colina) insieme con una caratteristica paralisi secretoria e midriasi, quasi combinasse le proprietà della tubocurarina e dell'atropina. Questo suggerì a F. C. MacIntosh che esso potesse essere un inibitore della colina-acetilasi. Egli e i suoi collaboratori trovarono, tuttavia, che il farmaco, inattivo da questo punto di vista, era un potente inibitore competitivo del trasporto della colina, sia nelle terminazioni dei nervi colinergici sia nel tubulo renale di pollo. Studi successivi hanno mostrato un'analoga efficacia contro il trasporto della colina in assoni, eritrociti e sinaptosomi. L'emicolinio fornì per la prima volta un mezzo per la deplezione (e quindi determinazione) delle scorte utilizzabili di acetilcolina, e Birks e MacIntosh (v., 1961) lo usarono nei loro classici studi sul metabolismo dell'acetilcolina nel ganglio del sistema autonomo. È stato dimostrato che la sua azione è condivisa specialmente da molecole contenenti i gruppi della colina, come l'HC3 o la trietilcolina, e da numerosi altri composti quaternari fra cui i comuni rilassanti. Il blocco neuromuscolare che produce è caratteristico: si sviluppa lentamente, più rapidamente se la velocità di stimolazione è maggiore, ed è lentissimo in assenza di attività nervosa motoria. Il blocco è antagonizzato alquanto dagli anticolinesterasici e intensificato dalla tubocurarina. Gli effetti dell'acetilcolina non sono antagonizzati; la cessazione della stimolazione porta a parziale recupero della trasmissione e i potenziali di placca in miniatura diminuiscono gradualmente di intensità. Tutti questi effetti possono essere attribuiti a un graduale esaurimento delle riserve di acetilcolina nella terminazione nervosa, in quanto si impedisce che questa capti la colina necessaria a mantenere la sintesi dell'acetilcolina.
MacIntosh ha anche osservato che i livelli plasmatici di colina possono essere abbassati da operazioni chirurgiche. Di conseguenza, nella pratica clinica, c'è una possibilità reale che questo fenomeno, insieme con la presenza in circolo di miorilassanti o altri composti quaternari, produca deplezione colinica e debolezza muscolare.
2. Altre sostanze che influenzano la produzione di acetilcolina. - Il calcio è necessario per la produzione di ACh, mentre il magnesio la deprime; la riduzione del rapporto Ca2+/Mg2+ del liquido di perfusione è un metodo comunemente usato per ridurre il livello dei quanti di acetilcolina negli studi di elettrofisiologia. Il meccanismo col quale la tossina botulinica diminuisce la produzione di ACh è ignoto, dato che non interferisce con la conduzione nervosa né riduce i quanti di acetilcolina. Le catecolammine possono aumentarne la produzione, mediante un effetto α che contribuisce ad antagonizzare il curaro; ma poiché esse possono esercitare sulla membrana postsinaptica anche un effetto β, che causa iperpolarizzazione e aumento del blocco curarico, possono essere ottenuti risultati assai disparati. La tetrodotossina paralizza la conduzione nervosa, ma non riduce la produzione di acetilcolina in risposta a depolarizzazione della terminazione nervosa. Questi effetti hanno soprattutto un significato sperimentale o tossicologico, ma quelli delle catecolammine possono avere un interesse più ampio.
3. Stimolazione e blocco delle terminazioni nervose presinaptiche. - Masland e Wigton nel 1940 aprirono un campo difficile e controverso quando osservarono che la stimolazione di un nervo motorio o l'iniezione di acetilcolina in un muscolo di gatto trattato con neostigmina poteva provocare una scarica antidromica nelle fibre nervose motrici. Questo fu il primo dato di quella che è ora una ampia serie di risultati sperimentali indicanti che un notevole numero di sostanze, in particolare composti quaternari etilati e anticolinesterasici, sono capaci di produrre tali effetti. Inoltre è stato osservato che talvolta questa scarica antidromica può essere soppressa da dosi molto basse di tubocurarina o analoghi antagonisti.
Basandosi su scoperte di questo tipo Riker e altri (v. Standaert e Riker, 1967) misero in dubbio i principi su cui poggiava la teoria classica della trasmissione chimica; essi prospettarono la possibilità che l'azione primaria dell'acetilcolina fosse quella di eccitare la terminazione nervosa invece dei recettori postsinaptici, e che l'azione primaria del curaro si esercitasse a questo livello. Viene così proposto, per esempio, che la contrattura del muscolo di Mammifero prodotta da iniezione di acetilcolina in arteria al punto di entrata nel muscolo stesso sia dovuta all'eccitazione delle terminazioni nervose con successiva trasmissione al muscolo in altro modo. Si mette in dubbio la prova della liberazione di acetilcolina e dell'incapacità del curaro di impedirla. Il fatto che dopo denervazione del muscolo l'acetilcolina e sostanze analoghe, come pure gli agenti bloccanti competitivi, mantengano i loro effetti caratteristici, viene confutato mettendo in evidenza l'alterazione delle proprietà e della distribuzione dei recettori che segue, come ora si sa, la denervazione. Il problema che ne risulta, cioè di come, se l'evidenza della trasmissione colinergica si riferisce soltanto a processi che interessano la terminazione nervosa, l'eccitazione si trasmetta alle fibre muscolari, è lasciato senza risposta. La teoria è interessante in quanto sposta tutta la farmacologia attuale delle sinapsi verso la terminazione nervosa.
La questione è controversa, e chi scrive può solo esporre le sue personali conclusioni. La prima è che le prove fornite non sono sufficienti a infirmare la teoria classica. La dimostrazione della liberazione di acetilcolina; le prove elettrofisiologiche della risposta postsinaptica, durante la trasmissione neuromuscolare, a quanti di un mediatore che assomiglia in ogni aspetto all'acetilcolina; le risposte del muscolo denervato; l'omologia della sinapsi neuromuscolare con quella, per esempio, fra il plesso di Auerbach e il muscolo longitudinale dell'ileo di cavia, nella quale i fattori pre- e postsinaptici possono essere distinti inequivocabilmente (v. Paton e Zar, 1968); il fatto che l'acetilcolina, anche se può essere capace di eccitare la terminazione nervosa motrice al primo nodo di Ranvier, o almeno di abbassarne la soglia di eccitazione, non aumenta, anzi diminuisce, la produzione di mediatore (v. Hubbard, 1965); questi e altri fatti rendono il concetto tradizionale molto più convincente. In secondo luogo, i dati sperimentali mettono in risalto la situazione farmacologica assai complicata che può riscontrarsi a livello della giunzione neuromuscolare se, per esempio, sono presenti sostanze che inibiscono la colinesterasi, depolarizzano la placca motrice, inducono - come in particolare i composti quaternari etilati - scariche ripetitive nei nervi dopo eccitazione singola, o fanno tutte queste cose insieme (v. Blaber e Bowman, 1963). Se c'è un'azione anticolinesterasica, il mediatore normale, ora in grado di uscire dalla fessura sinaptica, può verosimilmente raggiungere siti d'azione altrimenti irraggiungibili. Se la placca terminale è depolarizzata dalla persistenza del mediatore o dall'uso di una sostanza direttamente depolarizzante, allora, oltre tutto, la terminazione nervosa si troverà in un campo elettrico, e il contenuto potassico circostante sarà più elevato. I derivati etilati saranno così capaci di trasformare risposte singole in risposte multiple sia nel nervo sia nel muscolo. Infine, può aver luogo una trasmissione efaptica di potenziali d'azione dal muscolo al nervo. Sfortunatamente, nella maggior parte degli esperimenti in questo campo si registrano soltanto la tensione di contrattura muscolare e la scarica della radice ventrale, e non i cambiamenti del potenziale di membrana a livello della placca motrice.
Comunque, lasciando da parte le questioni fondamentali, è evidente che possono aver luogo effetti presinaptici antidromici, il cui significato non è ancora chiaro. Essi possono spiegare, almeno in parte, una risposta alle sostanze depolarizzanti, e cioè le fascicolazioni, che, potendo essere assai potenti, richiedono la contrazione coordinata di almeno un'unità motrice. È difficile che singole fibre muscolari possano riprodurre questo fenomeno, che è ben diverso dalla fibrillazione da denervazione, mentre non si può escludere una scarica antidromica provocante un riflesso assonico che interessi intere unità motrici, con un possibile riverbero all'interno di una unità.
d) Il problema del blocco duplice e fenomeni relativi.
Con queste nozioni fondamentali sui possibili meccanismi di blocco (competitivo, di depolarizzazione con ineccitabilità, di depolarizzazione con desensibilizzazione, presinaptico) possiamo ora considerare alcune situazioni complesse. Prima di tutto, il ‛blocco duplice'. Questo termine fu introdotto da Zaimis (v., 1953) per descrivere la situazione riscontrata nel caso in cui il carattere generale di un blocco neuromuscolare indotto da una sostanza cambiava da quello normalmente associato a sostanze depolarizzanti (fascicolazione iniziale, interessamento preferenziale del tibiale nel gatto, tetano sostenuto, resistenza agli anticolinesterasici, paralisi spastica nel pulcino) a quello associato a sostanze competitive (mancanza di effetti iniziali da stimolazione, interessamento primario del soleo nel gatto, tetano decrescente, sensibilità agli anticolinesterasici, paralisi flaccida nel pulcino). Talvolta, come quando il decametonio era somministrato a scimmie o lepri, il cambiamento si instaurava fra dosi successive; talvolta, come nel caso del tridecametonio, si manifestava durante l'azione di una dose singola. Zaimis formulò l'ipotesi che la sostanza cambiasse, in queste circostanze, il suo controllo sul recettore.
Inoltre, un fenomeno non dissimile fu descritto da Churchill-Davidson e Richardson (v., 1953), i quali osservarono che, somministrando decametonio a un paziente miastenico, si stabiliva un iniziale antagonismo alla miastenia (come era logico aspettarsi da una sostanza depolarizzante), seguito però da un periodo di accentuata paralisi di tipo curaro-simile.
In terzo luogo, durante studi in vitro di preparazioni nervo-muscolo esposte a decametonio o a succinilcolina, fu notato che il blocco poteva mostrare due fasi (v. Jenden e altri, 1951). La prima fase si sviluppava rapidamente e aveva alcune caratteristiche del blocco depolarizzante; quindi poteva scomparire, malgrado la presenza continuata della sostanza, e dar luogo a una seconda fase, più lenta e più curaro-simile, che durava, approfondendosi a poco a poco, finché la sostanza non era lavata via.
In quarto luogo può essere menzionato il blocco sinaptico in un'altra struttura, e cioè l'azione della nicotina sul ganglio cervicale superiore, paragonato da Zaimis al ‛blocco duplice'. Nel gatto, in vivo, la nicotina dapprima depolarizza il ganglio, bloccando la trasmissione di segnali ma provocando anche scariche ganglio-cellula; quindi la depolarizzazione svanisce, la scarica diminuisce, ma la trasmissione rimane paralizzata, e si ha antagonismo a dosi ulteriori di nicotina (v. Paton e Perry, 1953).
I dati sperimentali non permettono di interpretare in maniera soddisfacente questi fenomeni. Diversi meccanismi possono essere proposti. In primo luogo, come è stato suggerito per la nicotina in rapporto alla teoria cinetica dell'azione farmacologica, ci potrebbe essere un'occupazione graduale e crescente da parte del farmaco, cosicché la componente competitiva della sua azione diventerebbe via via predominante. Oppure potrebbe esserci una desensibilizzazione crescente, che riduce sia la depolarizzazione, sia l'effetto del mediatore. Oppure ancora, in conseguenza della depolarizzazione, potrebbero verificarsi nella regione della placca alterazioni ioniche (accumulo di sodio e perdita di potassio) in grado di cambiare il valore del fattore di sicurezza. Infine, continue o ripetute esposizioni potrebbero determinare effetti presinaptici, che ridurrebbero il fattore di sicurezza della trasmissione e così faciliterebbero la componente competitiva del blocco. Discriminare fra queste possibilità è assai complicato, e quasi impossibile nella pratica clinica, in quanto richiederebbe la valutazione del mediatore prodotto, della sensibilità chimica e dell'eccitabilità elettrica della placca terminale in tutte le condizioni considerate.
e) La miastenia grave e altri stati anormali della funzione neuromuscolare.
La miastenia grave è una condizione di debolezza muscolare episodica caratterizzata da un'accentuatissima ‛affaticabilità' del muscolo, nota da lungo tempo. Tuttavia solo nel 1934 essa venne comunemente accettata come una malattia neuromuscolare, e fu allora che Mary Walker, sulla base dell'analogia con il blocco da curaro, provò l'anticolinesterasico fisostigmina come agente terapeutico. La sua scoperta che questo farmaco poteva rendere reversibile la debolezza miastenica fu la base per la prima cura efficace, che è ancor oggi la più valida. I pazienti sono anche anormalmente sensibili alla tubocurarina; questo fece pensare che nel loro circolo vi fosse una sostanza curaro-simile, ma una prova certa di tale supposizione non è stata mai ottenuta e la causa della riduzione del fattore di sicurezza è ancora oscura. Non c'è aumento di attività della colinesterasi della placca. Studi con microelettrodi di muscoli miastenici relativamente non compromessi hanno rivelato potenziali di placca in miniatura di dimensioni molto ridotte. Poiché in questi studi la sensibilità della placca ai farmaci depolarizzanti non risultava alterata, si deve ammettere una riduzione del contenuto in acetilcolina dei singoli quanti. Fu ipotizzato anche un difetto di sintesi di acetilcolina, visto che in muscoli miastenici ‛compromessi' la stimolazione tetanica è seguita da un'accentuazione del blocco, come succede con l'emicolinio, invece che dalla facilitazione post-tetanica che si verifica dopo tubocurarina. D'altra parte, questo effetto presinaptico non spiega la relativa resistenza al decametonio di certi muscoli miastenici. Alterazioni istologiche sono state osservate nelle placche miasteniche; è frequente l'associazione della miastenia con un timoma. Esistono dati in favore di una reazione autoimmune verso il muscolo striato. La miastenia grave non è una malattia comune, ma stati miastenici sono presenti in affezioni maligne, particolarmente neoplasie polmonari.
f) Farmacogenetica del sussametonio (succinilcolina).
Nella pratica clinica vennero presto individuati pazienti in cui il sussametonio non aveva un'azione breve come nei soggetti normali, e in cui fu scoperta una carenza plasmatica di colinesterasi aspecifica. La sintesi dell'enzima sembra sia controllata da un gene singolo, ed esistono tre principali gruppi di individui facilmente distinguibili (v. Kalow, 1967): i normali, omozigoti per il gene normale; gli eterozigoti, circa il 3-4% della popolazione, con esterasi plasmatica più bassa che nel normale, in cui l'azione del sussametonio può essere un po' più prolungata; gli omozigoti per il gene difettivo, circa uno ogni 2-3.000, in cui una dose normale di sussametonio produce un effetto molto prolungato. Sono stati elaborati metodi enzimologici per l'identificazione di questi individui e sono state scoperte altre varianti genetiche.
Un altro aspetto genetico si rivela quando un farmaco depolarizzante è somministrato a individui affetti da certe miopatie. Nella distrofia miotonica, o miotonia congenita, il sussametonio causa uno spasmo che, una volta prodotto, resiste a ogni agente bloccante neuromuscolare, perché si genera al di là della placca motrice. L'unico rimedio efficace è il chinino, che è usato normalmente in questa malattia. Un altro caso è la rabdomiolisi idiopatica ricorrente, in cui fascicolazioni muscolari possono portare a mioglobinuria grave e blocco renale.
Queste alterazioni genetiche nell'uomo trovano corrispondenze negli animali. Vi sono variazioni notevoli dell'esterasi plasmatica in specie differenti. Per esempio, il gatto e la scimmia Papas hanno valori bassi, il macaco valori alti. Inoltre, si sa che esistono anche nei tessuti esterasi implicate nell'idrolisi. Nello studio generale dei rilassanti ad azione breve è divenuto importante tenere presenti queste differenze di specie e stabilire correlazioni con l'uomo.
4. Antagonismo del blocco neuromuscolare.
Poiché il blocco neuromuscolare può determinare la morte per paralisi dei muscoli respiratori, ci si è dedicati in particolare allo studio dei modi per antagonizzarlo, sia in caso di paralisi indotta durante operazioni chirurgiche o terapie convulsivanti, sia nella cura di stati miastenici. Gli obiettivi generali e i metodi usati derivano direttamente dai meccanismi di blocco già discussi.
1. Antagonisti del blocco competitivo. - Sono quelli più comunemente usati, e sono efficaci contro ogni agente bloccante competitivo non idrolizzato dalla colinesterasi. La neostigmina ha sostituito la fisostigmina per due ragioni principali. Anzitutto, è essa stessa un composto quaternario, cosicché è esclusa dal sistema nervoso centrale e perciò non ha certi effetti centrali indesiderabili prodotti dalla fisostigmina. Secondariamente, essa ha un leggero effetto depolarizzante sulla placca, in virtù dei suoi gruppi azotati metilati, e conseguentemente esplica un'azione anticurarica maggiore di quella anticolinesterasica con i suoi effetti indesiderabili su bronchi, cuore o secrezioni. Inoltre la sua attività e durata d'azione sono quelle utili. È stata esaminata un'ampia serie di altri anticolinesterasici, azotati o fosforati, tipo il diisopropilfiuorofosfato (DFP), ma senza trovare composti sostanzialmente superiori.
Un composto interessante è l'edrofonio, che può essere descritto come una neostigmina ad azione breve. Nel lavoro di ricerca esso è usato in modo particolare quando è richiesta un'inibizione transitoria. È utile anche nella cura di crisi colinergiche, che possono insorgere in decorsi postoperatori o nella miastenia quando, per caso o per errore, sia stata somministrata una dose alta di anticolinesterasico: poiché anche questo è in grado di produrre un blocco depolarizzante, può non essere chiara la natura della debolezza muscolare. La situazione può essere risolta dalla somministrazione di una dose test di edrofonio: un aggravamento significa eccesso di terapia antiesterasica, un miglioramento indica che questa deve essere mantenuta. L'aggravamento è comunque di breve durata.
Deve essere tenuto presente che gli anticolinesterasici hanno azione limitata. Una volta che l'enzima è completamente inibito, essi non possono più far nulla per antagonizzare il blocco competitivo, anzi possono cominciare a mostrare una loro propria azione di blocco neuromuscolare. Certi agenti bloccanti (per es. il benzochinonio) hanno essi stessi attività antiesterasica - una sorta di autoantagonismo interno - e di conseguenza la neostigmina ha un campo d'azione più limitato. In genere, l'agente competitivo per eccellenza dovrebbe essere completamente privo di attività antiesterasica.
Altre procedure, come l'uso di catecolammine o di potassio per antagonizzare il curaro, non hanno applicazione pratica. In teoria, dovrebbe bastare una piccola dose di un farmaco depolarizzante, ma l'equilibrio è così delicato e il pericolo di produrre un complicato blocco misto così grande, che l'uso pratico ne è escluso.
2. Antagonismo al blocco depolarizzante. - Nei primi esperimenti sul decametonio, i membri inferiori della serie, pentametonio ed esametonio, si rivelarono antagonisti, e il primo fu riconosciuto attivo sull'uomo. Questi farmaci, però, sono anche potenti agenti ganglioplegici e non hanno trovato applicazione pratica. La loro azione è paragonabile a quella di una piccola dose di agente bloccante competitivo, e rappresenta un altro aspetto del reciproco antagonismo fra le due specie di farmaci; ma, ancora una volta, l'equilibrio è troppo delicato per essere utilizzabile. Non c'è normalmente bisogno di antagonizzare il sussametonio, data la brevità della sua azione. Sono state approntate preparazioni di colinesterasi per uso clinico in casi di carenza enzimatica, ma non si sono mostrate sufficientemente attive e hanno potere antigenico. Sempre più spesso, col miglioramento delle tecniche di ventilazione controllata e della terapia di sostegno, si è riconosciuto che la migliore strategia è quella di astenersi dal somministrare altri farmaci e di permettere la ripresa normale delle placche motrici.
3. Legame dei farmaci. - Un'osservazione di grande interesse fu quella che l'inoculazione del colorante rosso Congo poteva esercitare antagonismo nei confronti della tubocurarina e di altri rilassanti (v. Kensler, 1949). Il meccanismo è dovuto alla capacità della grossa molecola acida del colorante di formare un complesso inattivo con la molecola basica del rilassante, come nel caso dell'uso della protammina per esercitare antagonismo verso l'eparina. Per quanto non sia risultato utile in clinica, questo effetto rimane una forma interessante di antagonismo, che ha il vantaggio di inattivare completamente il farmaco in oggetto, invece di contrastarlo con qualche altra azione.
4. Tetraetilammonio, germina e uso della scarica ripetitiva. - Abbiamo già menzionato che sali d'ammonio quaternari etilati, come il cloruro di tetraetilammonio, TEA, possono produrre scariche ripetitive di strutture eccitabili. Questa proprietà è alla base della capacità a prima vista sconcertante del TEA di esercitare antagonismo nei confronti sia del blocco da tubocurarina sia di quello da decametonio. Si è poi visto che il TEA fa aumentare la liberazione di acetilcolina nei gangli autonomi. Gli alcaloidi della veratrina hanno un effetto analogo, ma il loro meccanismo d'azione è differente, e mentre la veratrina è fortemente ipotensiva ed emetica e perciò non utilizzabile in clinica, il diacetato di germina è se mai ipertensivo e possiede un'azione emetica molto ridotta. Si è osservato che il diacetato di germina produce scariche ripetitive nel muscolo denervato o completamente curarizzato, esercita antagonismo verso la tubocurarina e il blocco parziale da depolarizzazione, produce le stesse scariche nel muscolo dell'uomo e aumenta la forza muscolare nella miastenia. Sembra che il suo effetto sia in gran parte muscolare, piuttosto che presinaptico. Un'azione di questo tipo richiede la presenza di sufficiente trasmissione da generare almeno alcuni potenziali d'azione, cosicché il blocco totale non può essere rimosso. D'altra parte, è un'azione abbastanza generale, indipendente da attività anticolinesterasica, che può anzi essere rafforzata. La germina è ancora scarsamente disponibile per un uso generale.
5. La sinapsi neuromuscolare per gli studi sul recettore.
Finora la discussione è stata focalizzata sugli effetti del blocco neuromuscolare più significativi dal punto di vista fisiologico e clinico. Ma fin dai classici studi di Langley, il curaro e le sostanze analoghe sono state usate per studiare il recettore farmacologico: questo capitolo tratterà tale aspetto.
a) Tipi di sinapsi neuromuscolare e specificità di azione.
Per prima cosa sembra opportuno analizzare e comparare tutte quelle strutture che possono essere considerate sinapsi neuromuscolari, cioè giunzioni fra nervi motori e muscolo striato. È noto da tempo che la sensibilità al curaro varia da muscolo a muscolo: i muscoli cranici e faringei e i piccoli muscoli delle mani sono particolarmente sensibili, il diaframma piuttosto resistente; in uno stesso arto muscoli rossi e muscoli bianchi hanno diverse sensibilità, e perfino in uno stesso muscolo alcune sinapsi sono più sensibili di altre. Ci sono notevoli variazioni fra specie e specie, e si è visto prima come la sensibilità sia diversa a seconda che l'agente sia competitivo o depolarizzante. Anche l'età influenza la sensibilità: il neonato è relativamente sensibile alla tubocurarina e resistente alla succinilcolina. Non c'è spiegazione soddisfacente per tutte queste differenze; ma alla luce di quello che sappiamo oggi su come l'innervazione di un muscolo controlla le sue proprietà fisiologiche, siamo probabilmente in presenza di influenze regolatrici protratte nel tempo, che controllano sia il risultato morfologico sia l'espressione o la repressione di capacità di sintesi e di liberazione del mediatore, la sensibilità chemocettiva e la risposta rigenerativa.
Bisogna anche distinguere fra muscoli strutturati in modo tale da essere attivati da potenziali d'azione propagati e quelle fibre con innervazione multipla (tipo retto addominale della rana) o muscoli denervati in cui l'area chemocettiva copre gran parte della membrana, attivando la contrazione senza attività propagata (v. Kuffler e Vaughan Williams, 1953). La facilità con cui la conseguente, sostenuta risposta meccanica dovuta all'applicazione di un farmaco depolarizzante può essere registrata ha fatto comunemente usare per l'analisi muscoli di questo tipo, come il retto addominale, il muscolo dorsale di sanguisuga, il muscolo di Uccello, o il muscolo denervato di Mammifero.
Dobbiamo ricordare due altri analoghi neuromuscolari. Il primo esempio è il terminale motorio della fibra γ nel fuso neuromuscolare (v. Matthews, 1972). Anatomicamente questo appare come una giunzione neuromuscolare colinergica specializzata; la parte sottostante al fuso è contrattile e causa una tensione del fuso stesso e quindi un aumento della scarica sensitiva. Un aumento di questo tipo si osserva dopo iniezione di sussametonio, decametonio e acetilcolina; la scarica è potenziata dalla fisostigmina (eserina) e bloccata dalla tubocurarina. In particolare si è visto che il sussametonio attiva le terminazioni delle fibre γ al livello dell'area del sacco nucleare del fuso che media le risposte γ dinamiche, e che queste terminazioni sono più sensibili di quelle statiche e alquanto più resistenti alla gallammina e alla tubocurarina delle sinapsi neuromuscolari ordinarie. Sarebbe davvero interessante, in vista di un impostazione nuova del problema delle malattie spastiche, poter trovare un'azione selettiva sulla placca terminale γ, ma finora non si ha indicazione di differenze sufficientemente sostanziali di sensibilità.
Il secondo esempio è fornito dagli organi elettrici di vari animali, in cui l'apparato contrattile del muscolo striato è represso e rimane una massa concentrata di placche disposte in modo da generare un potenziale elettrico. Gli organi elettrici sono riccamente innervati e contengono livelli altissimi di colinesterasi specifica. I nervi liberano acetilcolina e il tessuto è capace di sintetizzarla facilmente (v. Feldberg e Fessard, 1942). La placca terminale è attivata da acetilcolina, decametonio, sussametonio e carbacolo, mentre tubocurarina e farmaci analoghi sono antagonisti. Come vedremo più avanti, questi organi forniscono un sistema molto promettente per studiare biochimicamente il recettore dell'acetilcolina.
Malgrado le differenze quantitative individuali fra tutte queste giunzioni neuromuscolari, esse hanno caratteri generali comuni per quanto riguarda la risposta farmacologica, e si distinguono dalle sinapsi muscariniche (eccitate tipicamente dall'acetil-β-metilcolina e antagonizzate da scopolammina e atropina) e dalle sinapsi gangliari (stimolate da dosi ‛basse' di nicotina e bloccate da dosi ‛basse' di esametonio).
b) Relazioni fra struttura e funzione.
1. Generalità. - Agenti di blocco neuromuscolare sono stati oramai sintetizzati a centinaia e alcune caratteristiche generali sono chiare, anche se rimangono molti punti oscuri (v. Cavallito e Gray, 1960; v. Cavallito, 1967; v. Bovet, 1951; v. Paton e Waud, 1962).
I. Rimane stabilito, come proposto da Crum Brown e Fraser, che i composti azotati quaternari sono la fonte migliore di agenti di blocco neuromuscolare. Anche alcune ammine sono attive; le più importanti fra queste sono le β-eritroidine, che sono state sperimentate in clinica, ma si sono rivelate troppo fortemente ipotensive. In ogni modo i composti idonei sono tutti quaternari, forse perché, a parte il fatto che il mediatore stesso è un composto quaternario, tali sostanze entrano nelle cellule con difficoltà e hanno perciò la specificità e la mancanza di effetti cumulativi propri di una distribuzione extracellulare nei liquidi corporei.
II. In linea generale è anche assodato che composti con due gruppi quaternari danno la migliore riuscita. L'osservazione dell'elevata attività del decametonio, e dell'attività notevolmente inferiore dei sali di metonio con catena polimetilenica più lunga o più corta, suggerì l'ipotesi che una separazione di 10 atomi di carbonio (o il suo equivalente) fra gli atomi carichi potesse essere la distanza critica. È chiaro che con strutture più complicate questo può anche non essere vero. Sono stati trovati composti molto attivi contenenti nella molecola fino a 20 atomi di carbonio, con elementi carbonilici nella catena, e altri con solo 6-8 atomi, con altre strutture fra i gruppi carichi. Ciò nondimeno, un'elevata attività è frequentemente associata a una catena di collegamento di 9-11 atomi di carbonio. L'attività non è necessariamente connessa al fatto che ambedue i gruppi azotati siano quaternari, ma essa decresce nettamente se uno è amminico. Un ulteriore aumento del numero dei gruppi quaternari in genere riduce l'attività.
III. Prima di procedere oltre è necessario fare una distinzione fra agenti competitivi e depolarizzanti. Nel blocco competitivo, un'elevata attività è di solito associata con un azoto metilato, per quanto esistano composti validi interamente etilsostituiti (per es. la gallammina), o con altri sostituenti di grosse dimensioni. Analogamente non c'è una specifica esigenza di struttura per il resto della molecola, se si esclude il fatto che dovrebbe essere ragionevolmente lipofilo, il che in generale significa di grosse dimensioni. Ciò corrisponde alla classificazione di Bovet di queste sostanze come ‛pachicurari'.
IV. Per i composti depolarizzanti, invece, l'azoto metilato è virtualmente obbligatorio, e la struttura intermedia non deve avere grossi gruppi lipofili, come nelle catene lineari del decametonio, del sussametonio o della serie biscarbamminoica. Questo corrisponde ai ‛leptocurari' di Bovet.
V. L'importanza dell'equilibrio fra lipofilia e idrofilia si manifesta in altri modi. È particolarmente significativo che, nella serie tetraichilica, il tetrametilammonio depolarizzi facilmente, il tetraetilammonio molto debolmente, mentre i derivati tetralchilici superiori non depolarizzano affatto. Analogamente, nella serie alchiltrimetilammonica l'ottil-TMA depolarizza, il nonil-TMA è un agonista parziale e il decil-TMA è un agente di blocco competitivo. In questo caso il modo di azione cambia completamente quando non più di due −CH2− sono aggiunti a una catena già lunga otto atomi di carbonio (v. Paton e Waud, 1962). L'interpretazione di alcuni di questi fatti è abbastanza chiara. È prevedibile che, affinché una sostanza assomigli tanto all'acetilcolina da poter depolarizzare la placca terminale, debba contenere un azoto metilato legato a una catena lineare di qualche tipo. È ugualmente ragionevole, anche se più speculativo, postulare che l'elevata attività dei bisquaternari rifletta una disposizione di gruppi reattivi nella regione del recettore. Per esempio, non è necessario supporre che il decametonio reagisca con i suoi due poli ai siti anionici di due differenti gruppi recettori, anche se la simmetria della molecola lo suggerisce. Potrebbe ugualmente darsi che in vicinanza del principale sito anionico ci siano ulteriori siti carichi negativamente (che normalmente non prendono parte all'azione dell'acetilcolina), e che l'interazione con questi favorisca l'associazione farmaco-recettore. È stata anche suggerita l'ipotesi che il ruolo della seconda carica positiva nella molecola possa essere non tanto quello di un legame addizionale, quanto quello di allontanare da certe regioni critiche cationi interferenti. La distanza reale di interazione è incerta, non solo perché incerti sono i tipi di interazioni molecolari, ma anche perché non si conosce la reale conformazione assunta dalla molecola nel legame col recettore, che potrebbe non essere necessariamente la stessa che si riscontra nel cristallo o in soluzione. Il pancuronio, miorilassante steroideo, è un agente competitivo ad alta attività, la cui struttura rigida presumibilmente determina la distanza interquaternaria quando si associa al recettore (8 atomi di carbonio); tuttavia non è stato possibile ottenere una serie di strutture ‛rigide' sufficiente a dimostrare che questa è la distanza ottimale. Tutto sommato, sembra plausibile predire che quando la struttura del recettore sarà caratterizzata mostrerà un secondo sito anionico (forse un'altra area recettrice, forse no), distante dal sito anionico primario circa 10-15 Å, che potrà essere situato o meno su di un'area distinta.
Il ruolo dell'equilibrio fra idrofilia e lipofilia è più controverso. È facile identificare fattori che possono predisporre il sito recettore a una maggiore affinità, e così coadiuvare le forze di legame ionico fornite dai gruppi quaternari. Uno scheletro idrofobo può contribuire con forze di Van der Waals e di legame idrofobico, specie se l'adattamento alla superficie complementare del recettore è corretto. Del pari, la mancanza di attività può essere dovuta a impedimento sterico. Una parte idrofoba potrebbe anche alterare radicalmente lo stato delle molecole d'acqua sia al polo quaternario sia al sito recettore, provocando una riduzione della costante dielettrica vicino al gruppo quaternario, con conseguente diminuzione di affinità per l'acqua e movimento della molecola fuori della fase dispersa in direzione dell'interfacie del recettore. Inoltre potrebbe essere cambiato lo stato stesso dell'acqua nella regione del recettore (le membrane cellulari contengono una notevole quantità d'acqua e hanno una considerevole permeabilità all'acqua). Considerazioni analoghe possono essere valide anche nel caso di sostituzioni sul polo quaternario stesso, dove una perdita di affinità a causa di dispersione di carica o aumento di distanza dal punto d'aggancio più vicino potrebbe essere compensata da un'affinità maggiore determinata da legami di Van der Waals o idrofobici. Questo aumento di affinità potrebbe dar luogo alla descritta associazione di elevata attività e persistenza di azione. Tuttavia bisogna essere cauti nell'interpretazione dei dati, poiché può accadere che aumenti anche l'affinità verso altri punti di legame, e ciò può portare a riduzione d'attività, convogliando il farmaco verso siti inefficaci, o può produrre un prolungamento dell'azione, ritardando l'eliminazione.
Tuttavia queste considerazioni non spiegano il modo in cui molte forze molecolari che dovrebbero, teoricamente, aumentare l'affinità, riducano anche l'attività depolarizzante, o la trasformino in attività competitiva. L'esempio più chiaro, già menzionato, è quello in cui l'aggiunta di due gruppi metilenici all'ottiltrimetilammonio, che farebbe prevedere un aumento d'affinità di circa 6-9 volte, ben lungi dall'aumentare l'attività depolarizzante, di fatto l'abolisce. Questo e molti altri esempi di questo tipo possono essere spiegati facilmente sulla base della teoria cinetica dell'azione farmacologica, secondo la quale per l'eccitazione è necessario un turnover della molecola agonista, cosicché ogni processo che favorisce il legame, e ritarda pertanto la dissociazione dell'agonista dal recettore, nello stesso tempo diminuisce l'azione depolarizzante e aumenta quella competitiva.
2. Esigenze cliniche e rilassanti ad azione breve. - Riguardo ai farmaci utili in clinica, un successo considerevole è stato ottenuto in tre dei quattro obiettivi principali. L'anestesista ha adesso a disposizione soddisfacenti farmaci di tipo paralizzante ad azione sia lunga sia breve (per es. decametonio, carbolonio e sussametonio), e molti farmaci competitivi a media o lunga durata. Fra questi farmaci competitivi ci sono composti la cui azione è stata migliorata eliminando le azioni istamino-liberatrice e ganglioplegica della tubocurarina, e alcuni hanno attività elevatissima (10-30 μg/kg). Incidentalmente, è interessante che la C-tossiferina I, alcaloide naturale isolato dal calabassa-curaro, ha non solo una notevole attività, ma la mantiene in una larga gamma di specie (v. Paton e Perry, 1951); l'aver estratto un veleno così versatile è un altro contributo alla chimica applicata da parte degli Indios del Sudamerica! Rimane però l'esigenza di un farmaco del tipo della tubocurarina che abbia un'azione transitoria. L'uso di un legame estere per rendere il farmaco labile nell'organismo non è promettente, dato che serve per renderlo antagonizzabile con un anticolinesterasico. Fra i molti agenti bloccanti provati, ci sono molecole abbastanza complesse, alcune delle quali con un nucleo steroideo rigido, che hanno azione breve in molti animali, ma poi si rivelano ad azione molto più prolungata nell'uomo e nella scimmia. In questi casi la brevità d'azione è associata alla captazione da parte dei tessuti. Un farmaco competitivo ad azione breve è diventato sempre più desiderabile da quando sono stati osservati gli svantaggi del sussametonio, e cioè la stimolazione muscolare con conseguente dolore, la risposta anormale negli stati miopatici e l'eccessivo aumento del potassio plasmatico negli ustionati. Rimane perciò l'esigenza di trovare nuovi metodi per abbreviare la durata d'azione.
3. Legami a proteine e altri costituenti di tessuti. Ogni specie di legame di un farmaco attivo è interessante, in quanto può far luce sulle forze implicate a livello del recettore. Proprietà interessanti sono rivelate dal legame dei miorilassanti a proteine. È noto da tempo che in caso di malattie epatiche bisogna usare dosi più alte di tubocurarina. Ora si sa che questo deriva dal forte legame della tubocurarina a una γ-globulina, che aumenta nelle malattie epatiche. In generale sembra che questo legame sia favorito da gruppi -OH fenolici e ostacolato da gruppi -OH alcolici, e che gruppi allilici e di alcuni altri tipi cooperino al legame con l'ovoalbumina (v. Stovner e altri, 1972). Ora che la struttura delle proteine si sta definendo sempre meglio, studi più accurati di queste interazioni potrebbero dare ottimi risultati.
Un secondo tipo generale di legame è quello con le macromolecole acide. Uno studio di Chagas (v., 1962) sul destino della dimetiltubocurarina marcata con 14C ha sorprendentemente mostrato che, dopo somministrazione di una dose sostanziale, il tessuto che ne contiene quantità più elevate più di 20 volte del muscolo - è la cartilagine. Ora si sa che la tubocurarina si lega anche alle ribonucleoproteine (v. Namba e Grob, 1967) e che senza dubbio esistono numerose altre molecole capaci di legarla.
Questi fatti devono essere tenuti in considerazione non solo nell'interpretare la durata di azione di un farmaco e nel calcolare il bilancio del suo destino nell'organismo, ma anche nel tentare l'isolamento dei recettori per mezzo di una marcatura specifica.
c) Studi funzionali sul recettore.
In un capitolo precedente, il blocco competitivo è stato discusso nei termini dell'analisi dell'antagonismo farmacologico fatta da Gaddum (v., 1937), la quale poggia su un modello semplice espresso dalle seguenti reazioni reversibili:
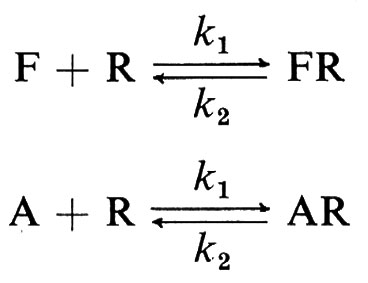
dove F è un farmaco stimolante che forma con il recettore R un complesso attivo FR, e A è un antagonista che si combina col recettore per dare un complesso inattivo. Da ciò si può desumere il parallelismo delle curve bg dose-risposta in presenza e in assenza dell'antagonista e la relazione fra (rapporto di dose) −1 e concentrazione dell'antagonista ÷ costante di dissociazione dell'antagonista (v. fig. 10). Questa teoria va bene per molti antagonisti; il suo successo, fra l'altro, prova che una molecola di farmaco (stimolante o antagonista) si combina con una molecola di recettore e che i vari recettori sono indipendenti fra di loro. Sulla base di questo schema fu avanzata l'ipotesi, nella teoria cinetica, che la stimolazione non dipendesse dalla quantità di FR, ma dalla sua velocità di formazione; questo permette di spiegare perché i fenomeni di stimolo sono spesso più pronunciati all'inizio dell'azione dell'agonista, quando il pool di recettori è ancora non occluso, e di considerare la differenza fra agonisti e antagonisti come dovuta alla forza del legame e alla costante di velocità della dissociazione.
Schemi ancora più elaborati sono stati proposti da studi sulla desensibilizzazione. Katz e Thesleff (v., 1957) trovarono, in particolare, che la velocità con cui si produce la desensibilizzazione può essere più bassa di quella con cui essa scompare. Questa osservazione non è compatibile con lo schema precedente. Perciò questi autori presero in considerazione altre formulazioni, fra cui il seguente schema generale:

in cui durante l'attivazione il recettore è trasformato dalla forma R in una forma non reattiva R′.
Una variante di questo schema fu usata da Rang e Ritter (v., On the mechanism... e The relationship..., 1970) per interpretare le loro osservazioni su un altro aspetto della desensibilizzazione. Essi mostrarono, sul muscolo di Uccello e di sanguisuga, che in seguito all'azione di uno stimolante cresceva l'affinità del recettore per certi antagonisti, specie quelli fortemente lipofili. L'effetto si mostrava inizialmente come una relativa inefficienza di questi antagonisti, a meno che il tessuto non fosse stato stimolato ripetutamente, o appena prima della loro applicazione o per lungo tempo in loro presenza. Questo fu una prova diretta che certe variazioni si producevano nel recettore in conseguenza dell'azione dell'agonista; esse furono denominate ‛effetto metafilico'. Quest'effetto si adattava bene allo schema precedente, ponendo come irreversibili i passaggi (2) e (4), e poté essere correlato quantitativamente alla desensibilizzazione.
Bisogna ricordare che è stato preso in considerazione anche un altro modello di recettore, basato sulla teoria allosterica. Si è avanzata l'ipotesi (v. Karlin, 1967) che il recettore possa esistere in due conformazioni, inattiva e attiva, e che stimolanti e antagonisti, avendo maggiore affinità per le forme attive e inattive rispettivamente, spostino in maniera appropriata l'equilibrio fra le due forme. Per quanto il fondamento teorico di questo schema sia piuttosto differente dal precedente, il comportamento funzionale predetto è simile; per il momento non si è riusciti a verificare sperimentalmente quale delle due alternative fosse corretta.
Conoscenze ulteriori sulla funzione del recettore sono state apportate dai recenti studi di Katz e Miledi (v., 1972 e 1973), che hanno mostrato come l'aumento di ‛rumore di fondo' della membrana nella placca motrice durante l'azione dell'acetilcolina possa essere analizzato in termini di singoli impulsi elementari di corrente prodotti dalle molecole di acetilcolina. Da questa analisi gli autori deducono che ogni interazione determina una depolarizzazione transitoria di 0,3 μV, vale a dire un aumento in conduttanza di circa 10-10 Ω-1 che dura circa 1 ms, con un trasferimento netto di carica di circa 5 • 104 ioni. Il raffreddamento prolunga il tempo di vita dell'evento elementare; il curaro non lo influenza; il decametonio, il carbacolo e l'acetilcolina producono impulsi più brevi (0,12-0,35 ms), la suberilcolina più lunghi (1,65 ms).
d) Studi di permeabilità al livello della giunzione neuromuscolare.
La variazione di conduttanza della placca motrice causata dall'acetilcolina e agonisti similari permette un aumento del flusso degli ioni sodio e potassio. Essa è differente dalle variazioni di conduttanza che avvengono durante l'attività propagata. Non è un fenomeno ‛tutto-o-nulla', ma di intensità dipendente dalla quantità del farmaco; l'aumento della conduttanza del potassio non è differito; la tetrodotossina non antagonizza l'aumento della conduttanza del sodio; il fenomeno può essere prodotto solo chimicamente, non elettricamente. Questa variazione è all'origine della liberazione di potassio negli spazi tessutali e nel plasma, che produce nell'uomo innalzamenti di concentrazione clinicamente significativi dopo somministrazione di sussametonio. È stato inoltre osservato da Creese e collaboratori (v. Creese ed England, 1970; v. Creese e MacLagan, 1970) che la placca diventa permeabile anche al decametonio marcato e ad altri ioni quaternari, che questo aumento di permeabilità non è dovuto semplicemente alla depolarizzazione locale indotta dalla sostanza, ma avviene anche in muscoli depolarizzati dal potassio, e che quest'ultima situazione è bloccata dalla tubocurarina. Il decametonio sembra penetrare nella fibra ed espandersi lateralmente con una diffusione molto rallentata, persistendo nella fibra per molti giorni.
Queste osservazioni spiegano alcuni dati di Waser (v., 1967). Egli aveva mostrato con l'autoradiografia che la placca motrice del topo era capace di captare l'agente bloccante competitivo tossiferina, con una distribuzione nettamente localizzata, e che a saturazione conteneva circa 5 • 106 molecole per placca. Col decametonio, invece, egli trovò una captazione oltre 100 volte maggiore, circa 1-2 • 109 molecole per placca, localizzate molto più diffusamente, senza segni di saturazione (v. fig. 11). Sembra perciò chiaro che una delle conseguenze della somministrazione di una sostanza depolarizzante è la sua captazione al livello della placca e anche, forse, la permeazione di questa ad altri cationi presenti, di dimensioni relativamente grandi. L'importanza di ciò per la funzione neuromuscolare in un arco di tempo lungo deve ancora essere indagata, per quanto non vi sia alcun dato che indichi che, a tempi brevi, la presenza di una piccola quantità di cationi estranei nella placca abbia qualche effetto.
e) Studi biochimici sul recettore.
A mano a mano che il concetto del recettore farmacologico diveniva più credibile e che si veniva affermando il successo delle moderne tecniche biochimiche di isolamento e caratterizzazione di vari tipi di macromolecole, sono stati fatti ripetuti tentativi per isolare e studiare il recettore dell'acetilcolina. Questi studi sono stati preceduti e accompagnati da approcci meno diretti, che hanno prodotto due risultati utili. Il primo è che appare esserci un ponte disolfuro di importanza critica nella regione del recettore, la cui riduzione con ditiotreitolo altera le proprietà del recettore, e in particolare converte l'esametonio da agente bloccante del muscolo striato o dell'elettroplacca a stimolante (v. Karlin e Winnik, 1968). Il secondo è che, sulla base di argomentazioni chimiche, è stato proposto (v. Cavallito, 1967) che gruppi polifosforici possano rappresentare il sito anionico; ed è interessante che lo ione uranile, che ha un'alta affinità per il fosfato, si sia rivelato un agente di blocco competitivo.
Il maggiore interesse, però, risiede nell'approccio diretto. I primi tentativi fallirono, poiché quello che si era ritenuto fosse la combinazione del farmaco con il recettore si rivelò essere un legame aspecifico da parte di altri costituenti del tessuto, quali il condroitinsolfato o le ribonucleoproteine. Ma questi sforzi resero molto più chiaro quello che bisognava fare: e cioè che, per verificare che avveniva una vera e propria combinazione col recettore, era necessario mostrare che la costante di associazione per il farmaco usato, misurata a forza ionica fisiologica, corrispondeva a quella ottenuta da studi funzionali, e che il legame era inibito competitivamente da altri agenti bloccanti a una concentrazione in accordo con le loro note proprietà funzionali. Due progressi tecnici si rivelarono utili. Era stato trovato, studiando la captazione di atropina da parte del muscolo liscio (v. Paton e Rang, 1965), che, benché a livello cellulare tale captazione fosse considerevole, la quantità totale legata specificamente era molto piccola (circa 100 pmoli per grammo). Questo fatto mise in luce la necessità di un tessuto il più ricco possibile in recettori, e tale tessuto fu trovato nell'organo elettrico. Il secondo punto fu che, malgrado non sia troppo difficile dimostrare la captazione di agenti competitivi reversibili da parte di un tessuto, la reversibilità stessa significa che il farmaco combinato può dissociarsi ed essere perduto durante i procedimenti di purificazione o di analisi; era assolutamente necessario trovare un agente specifico di blocco neuromuscolare che fosse irreversibile. Un agente di questo tipo fu trovato nei veleni di serpente, la α-bungarotossina di Bungarus multicinctus e la α-tossina di Naja nigricollis. È interessante notare che la natura ha avuto più successo dell'uomo nella sintesi chimica di un agente bloccante irreversibile.
Miledi, Molinoff e Potter (v., 1971) hanno trattato con bungarotossina marcata con 131I porzioni di organo elettrico di Torpedo e sono poi riusciti, frazionando il tessuto, a isolare un componente proteico dalla frazione delle membrane, distinguibile biochimicamente dalla colinesterasi, e un enzima marcante, purificato fino a circa 200 nanomoli di siti di recettore per grammo di proteina. Changeux e altri (v., 1971) hanno usato frammenti di membrana di Electroplax electricus, solubilizzati con desossicolato, combinati con decametonio marcato con 14C; la bungarotossina e la tossina di Naja nigricollis servirono a identificare la frazione specifica del recettore combinata col decametonio (che si dimostrò anche legato alla colinesterasi; v. fig. 12). Fu ottenuta un'attività di 100 nanomoli di recettore per grammo di proteina. La macromolecola interessata si dimostrò termolabile, digeribile dall'enzima proteolitico pronasi e di peso molecolare superiore a 50.000. De Robertis e altri (v., 1971) usarono tecniche alquanto differenti, isolando un proteolipide in condizioni relativamente non polari in cui non poterono essere compiuti adeguati studi di legame; anche in questo caso si è ottenuto un materiale purificato capace di legare l'acetilcolina, l'esametonio e la tubocurarina.
Da questi e altri studi appare chiaro che è stato compiuto un progresso notevole. Si può notare che in ogni caso il materiale legante si comporta solo approssimativamente come ci si aspetterebbe dagli esperimenti con cellule intatte, ed è chiaro che il recettore può avere una struttura complessa, incapace di resistere a procedimenti di frazionamento. È probabile che dovremo fronteggiare problemi ancora più complessi per caratterizzare completamente ‛l'anatomia' del recettore.
6. I più recenti sviluppi.
In anni recenti c'è stato un enorme sviluppo degli studi sulla trasmissione colinergica, ampiamente descritti in numerose rassegne (v. Goldberg e Hanin, 1976; v. Jenden, 1977; v. Rang, 1975; v. Cohen e Changeux, 1975; v. MacIntosh e Collier, 1976; v. Cheney e Costa, 1977; v. Heidmänn e Changeux, 1978). La reale portata di questi studi ai fini della comprensione del blocco neuromuscolare e del suo migliore impiego terapeutico deve ancora essere valutata. Pertanto sembra più utile in quest'ultimo capitolo descrivere brevemente alcuni di questi studi piuttosto che tentare di metterli direttamente in relazione al blocco neuromuscolare.
1. Il recettore dell'acetilcolina. - L'organo elettrico di Torpedo si è rivelato una ricca fonte di tale recettore, una proteina che, nella sua forma purificata, appare al microscopio elettronico come un disco costituito di 5-6 subunità disposte intorno a un'apertura centrale; sono state riscontrate strutture simili raggruppate in una disposizione esagonale in membrane ottenute dalla regione della placca terminale sottoposte a criodecapaggio. Secondo dati recenti sembra che il recettore contenga cinque subunità costituite da quattro peptidi di peso molecolare 40.000, 50.000, 60.000 e 65.000, nelle proporzioni di 2:1:1:1. Da quanto risulta in base alle analisi biochimiche sembra che, geneticamente, la divergenza da un gene ancestrale comune sia avvenuta in un precoce stadio evolutivo del recettore (v. Raftery e altri, 1980). Gli studi sui lisati reticolocitari hanno permesso di identificare un mRNA che regola la sintesi dei peptidi del recettore, di peso molecolare un po' più basso, il che potrebbe esser dovuto alla mancanza di glicosilazione (v. Mendez e altri, 1980).
Uno dei principali modi per isolare il recettore è tramite la α-bungarotossina, che riesce a legarsi quasi irreversibilmente col recettore dell'acetilcolina. Esiste tuttavia anche un altro importante sito del recettore: infatti il canale di conduttanza associato lega selettivamente la istrionicotossina (un alcaloide che deriva dalla secrezione della pelle di una rana colombiana, Dendrobates histrionicus); ciò permette di effettuare degli studi differenziali (v. Eldefrawi e altri, 1980).
Un bellissimo e importante lavoro è stato quello di Neher e Sakmann (v., 1976), i quali hanno scoperto un metodo di registrazione dai singoli canali di conduttanza nelle aree del recettore dell'acetilcolina. Ciò è stato possibile usando fibre muscolari trattate con collagenasi in modo da eliminare il tessuto connettivo e la membrana basale, e quindi fissando contro la membrana una micropipetta con una punta levigata contenente carbacolo. Si è così scoperto che i cambiamenti di conduttanza provocati consistevano in brevi ‛onde quadrate', quantiche, con andamento temporale casuale, e che il canale di conduttanza era simile a quello che Katz e Miledi avevano per primi individuato in base all'analisi del ‛rumore'. Questi studi indicano che la velocità di apertura dei canali è una funzione della concentrazione della sostanza, e quella di chiusura è una funzione del potenziale di membrana (v. Sheridan e Lester, 1977; v. Adam, 1974). Tutt'e due i metodi hanno portato a concludere che vi sono numerose sostanze che riescono a bloccare i canali di conduttanza quando sono aperti; si spiegano in tal modo quelle forme in cui il blocco si intensifica a seconda dell'intensità dell'attività dell'agonista. Altri lavori sulla sensibilità al voltaggio dei cambiamenti di conduttanza e sull'efficacia di svariati agenti bloccanti hanno condotto a una conclusione simile, e cioè che esistono dei siti di legame all'interno dei canali di conduttanza che possono essere raggiunti solo quando i canali sono aperti (v. Neher e Stembacli, 1978; v. Colquhoun e altri, 1979). L'intensità del blocco diviene una funzione del potenziale di membrana, il che determina la forza che guida la sostanza all'interno del canale. Una conseguenza importante, anche se sconcertante, è che non si possono più considerare universalmente applicabili le semplici correlazioni dell'antagonismo competitivo, in base alle quali si ritiene che le molecole competano per il sito del recettore indipendentemente da altri eventi a livello della membrana. Dal momento che l'azione bloccante della tubocurarina è di per sé sensibile al voltaggio, bisogna supporre che il suo recettore si trovi sufficientemente all'interno del campo del potenziale di membrana perché l'apparente affinità della sostanza per il complesso del recettore possa essere alterata dai cambiamenti di tale potenziale. È certo che, a livello di un'altra sinapsi colinergica, la cellula gangliare, questo lavoro ha mostrato che la tubocurarina, l'esametonio e il decametonio si comportavano da agenti di blocco dei canali piuttosto che da antagonisti competitivi in grado di legarsi a un complesso canale chiuso-recettore (v. Ascher e altri, 1979). Benché queste scoperte rendano necessario rivedere gran parte del lavoro passato, devono essere considerate rispondenti a una situazione molto più realistica, e cioè quella dell'interazione con le subunità peptidiche in un campo elettrico, e non quella degli artificiali diagrammi della precedente teoria.
Un ultimo importante sviluppo nello studio del recettore è quello della biogenesi, sintesi, diffusione, ricambio e degradazione del recettore. È probabile che i cambiamenti nella dinamica della popolazione del recettore - siano essi dovuti a trattamento farmacologico o a variazioni dell'attività nervosa, o rientrino in un processo morboso - risulteranno essere un campo di ricerca ampio e fertile (v. Fambrough, 1979; v. Edwards, 1979; v. Lewitt e altri, 1980).
2. Metabolismo dell'acetilcolina. - La scoperta, effettuata nell'ambito di studi fisiologici, della liberazione quantica dell'acetilcolina e l'individuazione ultrastrutturale delle vescicole della terminazione nervosa colinergica, che sono risultate ricche di acetilcolina, hanno portato alla tacita accettazione del fatto che la liberazione del trasmettitore avvenga normalmente per esocitosi, accettazione resa più plausibile dalla presenza di colinesterasi nel sol citoplasmatico. Numerosi lavori mostrano adesso che l'ACh può essere presente, in forma integra, nel sol citoplasmatico, diffusa al suo interno, ed essere liberata (v. Koike e Nagata, 1979; v. Johnson e Pilar, 1980). Sembra inoltre che alle riserve extravescicolari sia dovuta gran parte della liberazione ‛spontanea' a riposo, mentre le vescicole prenderebbero parte alla liberazione calcio-dipendente (v. Zimmerman, 1979; v. Boksa e Collier, 1980; v. Carroll e Aspry, 1980).
Il problema fondamentale di come la sintesi dell'acetilcolina sia regolata in modo da mantenere delle riserve adeguate nonostante le diverse velocità di liberazione è tuttora insoluto. La velocità di sintesi non sembra controllata da un'inibizione della sua produzione all'interno della terminazione nervosa o dall'entrata del sodio. È chiaro che la colina che si ottiene per idrolisi di acetilcolina di recente liberazione viene usata preferenzialmente per la sintesi; ma la velocità di sintesi non è controllata dalla disponibilità o dalla captazione di colina (v. Collier e Ilson, 1977; v. Kessler e Marchbanks, 1979). È probabile che sia collegata alla captazione di calcio o che sia la conseguenza di una depolarizzazione della terminazione nervosa. Il ruolo del calcio nella liberazione di trasmettitore è discusso altrove (v. sinapsi: Fisiologia della sinapsi periferica e Fisiologia della sinapsi centrale) e sembra appurato che la facilitazione post-tetanica sia legata alla captazione di Ca2+. È interessante il dato ottenuto da Schmidt e altri (v., 1980), i quali hanno scoperto che il contenuto in calcio delle vescicole sinaptiche dell'organo elettrico di Torpedo aumenta in conseguenza della stimolazione. Il recente interesse nei confronti delle sostanze che controllano l'entrata del calcio nelle cellule può avere ripercussioni sull'uso degli agenti di blocco neuromuscolare.
3. Myasthenia gravis. - L'aver definitivamente accertato che la myasthenia gravis è una malattia autoimmune in cui il pool del recettore a livello della placca motrice terminale viene ridotto dagli anticorpi in circolazione, rappresenta un notevole progresso che spiega in maniera elegante l'analogia con la curarizzazione e l'efficacia del trattamento con anticolinesterasici. Un quadro simile può essere prodotto nei Mammiferi tramite immunizzazione con la proteina che costituisce il recettore dell'acetilcolina, isolata dall'organo elettrico di Torpedo, il che dimostra come il recettore si sia mantenuto uguale attraverso l'evoluzione. Un simposio della New York Academy of Sciences (v. Grob, 1976) dà notizia di molti degli sviluppi che sono conseguiti a questa scoperta; uno dei più importanti è una migliore comprensione delle varie forme di miopatie. Dal punto di vista della terapia, una sostituzione di plasma unita a trattamento immunosoppressivo promette notevoli successi (v. Behan e altri, 1979).
4. Agenti di blocco neuromuscolare ad azione rapida. - Possiamo da ultimo menzionare il fatto che la lunga ricerca per trovare altri agenti di blocco ad azione rapida non dipendente dall'idrolisi operata dalla colinesterasi del plasma comincia a dare risultati. Per quanto da un punto di vista clinico il sussametonio abbia grande successo, la prolungata paralisi che esso induce in pazienti con deficienza di colinesterasi dovuta a difetti genetici, la presenza di reazioni abnormi in alcune miopatie, la tendenza a provocare dolori muscolari postoperatori e la mancanza di anti-esterasici come antidoti in caso di bisogno, hanno determinato la necessità di un agente di blocco non depolarizzante. A questo riguardo sembra molto promettente l'atracurio, un agente di blocco neuromuscolare competitivo la cui attività viene interrotta prevalentemente da una riorganizzazione intramolecolare (v. Hughes e altri, 1980).
Bibliografia.
Adam, P. R., Kinetics of agonist conductance changes during hyperpolarization at frog endplates, in ‟British journal of pharmacology", 1974, LIII, pp. 308-310.
Albuquerque, E. X., Smith, C. M., Stimulation of muscle spindle afferents by DMPP, in ‟Journal of pharmacology and experimental therapeutics", 1965, CXLIX, pp. 320-328.
Ascher, P., large, W. A., Rang, H. P., Studies on the mechanism of action of acetylcholine antagonists on rat parasympathetic ganglion cells, in ‟Journal of physiology", 1979, CCXCV, pp. 139-170.
Bacq, Z. M., Brown, G. L., Pharmacological experiments on mammalian voluntary muscles, in relation to the theory of chemical transmission, in ‟Journal of physiology", 1937, LXXXIX, pp. 45-60.
Barlow, R. B., Ing, H. R., Curare-like action of polymethylene bisquaternary ammonium salts, in ‟British journal of pharmacology and chemotherapy", 1949, III, pp. 298-304.
Behan, P. O., Shakir, R. A., Simpson, J. A., Burnett, A. K., Allan, T. L., Haase, G., Plasma-exchange combined with immunosuppressive therapy in myasthenia gravis, in ‟Lancet", 1979, II, pp. 438-440.
Bennett, A. E., Curare: a preventive of traumatic complication in convulsive shock therapy, in ‟American journal of psychiatry", 1941, XCVII, pp. 1040-1060.
Bernard, Cl., Leçons sur les effets des substances toxiques, Paris 1857.
Birks, R., MacIntosh, F. C., Acetylcholine metabolism of a sympathetic ganglion, in ‟Canadian journal of biochemistry and physiology", 1961, XXXIX, pp. 787-827.
Blaber, L. C., Bowman, W. C., Studies on the repetitive discharges evoked in motor nerve and skeletal muscle after injection of anticholinesterase drugs, in ‟British journal of pharmacology and chemotherapy", 1963, XX, pp. 326-344.
Boksa, P., Collier, B., Spontaneous and evoked release of acetylcholine and a cholinergic false transmitter from brain slices: comparison to true and false transmitter in subcellular stores, in ‟Neuroscience", 1980, V, pp. 1517-1532.
Bovet, D., Some aspects of the relationship betweenchemical constitution and curare-like activity, in ‟Annals of the New York Academy of Sciences", 1951, LIV, pp. 407-432.
Bovet, D., Bovet-Nitti, F., Marini-Bettolo, G. B. (a cura di), Curare and curare-like agents, Amsterdam 1959.
Bremer, F., Titeca, J., Atonie curarique et inhibition de Wedensky, in ‟Archives internationales de physiologie", 1935, XLII, pp. 223-250.
Bülbring, E., Isolated phrenic nerve diaphragm preparation of rat, in ‟British journal of pharmacology and chemotherapy", 1946, I, pp. 38-61.
Burns, B. D., Paton, W. D. m., Depolarization of the motor end-plate by decamethonium and acetylcholine, in ‟Journal of physiology", 1951, CXV, pp. 41-73.
Carroll, P. T., Aspry, J.-A.M., Subcellular origin of cholinergic transmitter release from mouse brain, in ‟Science", 1980, CCX, pp. 641-642.
Cavallito, C. J., Bonding characteristics of acetylcholine stimulants and antagonists and cholinergic receptor, in ‟Annals of the New York Academy of Sciences", 1967, CXLIV, pp. 900-911.
Cavallito, C. J., Gray, A. P., Chemical nature and pharmacological actions of quaternary ammonium salts, in ‟Progress in drug research", 1960, II, pp. 136-226.
Chagas, C., The fate of curare during curarization, in Curare and curare-like agents (a cura di . V. S. de Reuck), London 1962, pp. 2-10.
Changeux, J.-P., Meunire, J.-C., Huchet, M., Studies on the cholinergic receptor protein of Electrophorus electricus, in ‟Molecular pharmacology", 1971, VII, pp. 538-553.
Cheney, D. L., Costa, E., Pharmacological implications of brain acetylcholine turnover measurements in rat brain nuclei, in ‟Annual review of pharmacology", 1977, XVII, pp. 369-386.
Churchill-Davidson, H. C., Richardson, A. T., Neuromuscular transmission in myasthenia gravis, in ‟Journal of physiology", 1953, CXXII, pp. 252-263.
Cohen, J. B., Boyd, N. D., Conformational transitions of the membrane-bound cholinergic receptor, in Catalysis in chemistry and biochemistry (a cura di B. Pottman e D. Ginsburg), 1979, pp. 293-304.
Cohen, J. B., Changeux, J.-P., The cholinergic receptor protein in its membrane environment, in ‟Annual review of pharmacology", 1975, XV, pp. 83-103.
Collier, B., Ilson, D., The effect of preganglionic nerve stimulation on the accumulation of certain analogues of choline by a sympathetic ganglion, in ‟Journal of physiology", 1977, CCLXIV, pp. 489-501.
Colquhoun, D., Dreyer, F., Sheridan, R. E., The action of tubocurarine at the frog neuromuscular junction, in ‟Journal of physiology", 1979, CCXCIII, pp. 247-284.
Cookson, J.C., Paton, W. D. M., Mechanisms of neuromuscular block, in ‟Anaesthesia", 1969, XXIV, pp. 395-416.
Creese, R., England, J. M., Decamethonium in depolarized muscle and the effects of tubocurarine, in ‟Journal of physiology", 1970, CCX, pp. 345-361.
Creese, R., MacLagan, J., Entry of decamethonium in rat muscle studied by autoradiography, in ‟Journal of physiology", 1970, CCX, pp. 363-386.
Crum Brown, A., Fraser, T. R., On the connection between chemical constitution and physiological action. I. On the physiological action of the salts of the ammonium bases derived from Strychnia, Brucia, Thebaia, Codeia, Morphia, and Nicotia, in ‟Transactions of the Royal Society of Edinburgh", 1869, XXV, pp. 1-53.
Crum Brown, A., Fraser, T. R., On the connection between chemical constitution and physiological action. II. On the physiological action of the ammonium bases derived from Atropia and Conia, in ‟Transactions of the Royal Society of Edinburgh", 1869, XXV, pp. 693-739.
Cullen, S. C., The use of curare for improvement of abdominal muscle relaxation during inhalation anaesthesia, in ‟Surgery", 1943, XIV, pp. 261-266.
De Robertis, E., Lunt, G. S., La Torre, J. L., Multile binding sites for acetylcholine in a proteolipid from electric tissue, in ‟Molecular pharmacology", 1971, VII, pp. 97-103.
Edwards, C., The effects of innervation on the properties of acetylcholine receptors in muscle, in ‟Neuroscience", 1979, IV, pp. 565-584.
Ehrlich, P., Chemotherapy (1914), in The collected papers of Pau Ehrlich, vol. III, London 1960, pp. 505-518.
Eldefrawi, M. E., Aronstam, R. S., Bakry, N. B., Eldefrawi, A. T., Albuquerque, E. X., Activation, inactivation and dseensitization of acetylcholine receptor channel complex detetcted by binding of perhydrohistrionicotoxin, in ‟Proceedings of the National Academy of Sciences, 1980, LXXVII, pp. 2309-2313.
Everett, A. J., Lowe, L. A., Wilkinson, S., Revision of the structure of (+) tubocurarine, in ‟Journal of the Chemical Society. Chemical communications", 1970, pp. 1020-1021.
Fambrough, D. M., Control of acetylcholine receptors in skeletal muscle, in ‟Physiological reviews", 1979, LIX, pp. 165-227.
Feldberg, W., Fessard, A., The cholinergic nature of the nerves to the electric organ of the Torpedo (Torpedo marmorata), in ‟Journal of physiology", 1942, CI, pp. 200-216.
Gaddum, J. H., The quantitative effects of antagonistic drugs, in ‟Journal of physiology", 1937, LXXXIX, pp. 7-9P.
Goldberg, A. M., Hanin, I. (a cura di), Biology of cholinergic function, New York 1976.
Granit, R., Skoglund, S., Thesleff, S., Activation of muscle spindles by succinylcholine and decamethonium. The effects of curare, in ‟Acta physiologica scandinavica", 1953, XXVIII, pp. 134-151.
Griffith, H. R., Johnson, G. E., The use of curare in general anaesthesia, in ‟Anesthesiology", 1942, III, pp. 418-420.
Grob, D. (a cura di), Myasthenia gravis, in ‟Annals of the New York Academy of Sciences", 1976, CCLXXIV.
Heidmann, T., Changeux, J.-P., Structural and functional properties of the acetylcholine receptor protein in its purified and membrane bound states, in ‟Annual review of biochemistry", 1978, XLVII, pp. 317-357.
Hill, A. V., The mode of action of nicotine and curare determined by the form of the contraction curve and the method oftemperature coefficients, in ‟Journal of physiology", 1910, XXXIX, pp. 361-373.
Hubbard, J. I., The origin and significance of antidromic activity in motor nerves, in Studies in physiology (a cura di D. R. Curtis e A. K. McIntyre), Berlin 1965, pp. 85-92.
Hughes, R., Al Azawi, A., Hunt, T. M., Payne, J. P., Further studies with atracurium, in ‟British journalof anaesthesia", 1980, LII, pp. 956-957P.
Ing, H. R., The curariform action of onium salts, in ‟Physiological reviews", 1936, XVI, pp. 527-544.
Jenden, D. J., Cholinergic mechanisms and psychopharmacology, New York 1977.
Jenden, D. J., Kamijo, K., Taylor, D.R., The action of decamethonium (C10) on the isolated rabbit lumbrical muscle, in ‟Journal of pharmacology and experimental therapeutics", 1951, CIII, pp. 348-349.
Johnson, D. A., Pilar, G., The release of acetylcholine from post-ganglionic cell bodies in response to depolarization, in ‟Journal of physiology", 1980, CCIC, pp. 605-619.
Kalow, W., Pharmacogenetics and the predictability of drug responses, in Drug responses in man (a cura di G. Wolstenholme e R. Porter), London 1967, pp. 220-239.
Karlin, A., On the application of ‛a plausible model' of allosteric proteins to the receptor for acetylcholine, in ‟Journal of theoretical biology", 1967, XVI, pp. 306-320.
Karlin, A., Winnik, M., Reduction and specific alkylation of the receptor for acetylcholine, in ‟Proceedings of the National Academy of Sciences", 1968, LX, pp. 668-674.
Katz, B., Miledi, R., The statistical nature of the acetylcholine potential and its molecular components, in ‟Journal of physiology", 1973, CCXXX, pp. 707-717.
Katz, B., Miledi, R., The characteristics of ‛end-plate noise' produced by different depolarizing drugs, in ‟Journal of physiology", 1973, CCXXX, pp. 707-717.
Katz, B., Thesleff, S., A study of the ‛desensitization' produced by acetylcholine at the motor end-plate, in ‟Journal of physiology", 1957, CXXXVIII, pp. 63-80.
Kellgren, J. H., McGowan, A. J., Wood, D. R., The effect of d-tubocurarine chloride on sensation in man, in ‟British medical journal", 1946, II, pp. 898-899.
Kensler, C. J., The antagonism of curare by Congo red and related compounds, in ‟Journal of pharmacology and experimental therapeutics", 1949, XCV, pp. 28-44.
Kessler, P. D., Marchbanks, R. M., Choline transport is not coupled to acetylcholine synthesis, in ‟Nature", 1979, CCLXXIX, pp. 542-544.
King, H., Curare alkaloids. I. Tubocurarine, in ‟Journal of the Chemical Society", 1935, pp. 1381-1389.
Koike, H., Nagata, Y., Intra-axonal diffusion of [3H] acetylcholine and [3H] aminobutyric acid in a neurone of Aplysia, in ‟Journal of physiology", 1979, CCXCV, pp. 397-417.
Kuffler, S. W., Vaughan Williams, E. M., Small-nerve junctional potentials. The distribution of small motor nerves to frog skeletal muscle, and the membrane chararcteristics of the fibres they innervate, in ‟Journal of physiology", 1953, CXXI, pp. 289-317.
Kuffler, S. W., Vaughan Williams, E. M., Properties of the ‛slow' skeletal muscle fibres of the frog, in ‟Journal of physiology", 1953, CXXI, pp. 318-340.
Langley, J. N., On the contractin of muscle, chiefly in relation to the presence of ‛receptive' substances. I., in ‟Journal of physiology", 1907, XXXVI, pp. 347-384.
Langley, J. N., On the contraction of muscle, chiefly in relation to the presence of ‛receptive' substances. II., in ‟Journal of physiology", 1908, XXXVII, pp. 165-212.
Langley, J. N., On the contraction of muscle, chiefly in relation to the presence of ‛receptive' substances. III. The reaction of frog's muscle to nicotine after denervation, in ‟Journal of physiology", 1908, XXXVII, pp. 285-300.
Langley, J. N., On the contraction of muscle, chiefly in relation to the presence of ‛receptive' substances. IV. The effect of curari and of some other substances on the nicotine response of the sartorius and gastrocnemius muscles of the frog, in ‟Journal of physiology", 1909, XXXIX, pp. 235-295.
Lapicque, L., Lapicque, M., Variations de l'excitabilité du muscle dans la curarisation, in ‟Comptes rendus des séances de la Société de Biologie", 1906, LX, pp. 991-996.
Lewitt, T. A., Loring, R. M., Salpeter, M. M., Neuronal control of acetylcholine receptor turnover rate at a vertebrate neuromuscular junction, in ‟Science", 1980, CCX, pp. 550-551.
MacIntosh, F. C., Synthesis and storage of acetylcholine in nervous tissue, in ‟Canadian journal of biochemistry and physiology", 1963, XLI, pp. 2555-2571.
MacIntosh, F. C., Collier, C., Neurochemistry and cholinergic terminals, in Handbook of experimental pharmacology, vol. XLII, Neuromuscular junction (a cura di E. Zaimis), Berlin 1976, pp. 99-228.
McIntyre, A. R., Curare: its history, nature and clinical use, Chicago 1947.
Marsh, D. F., Herring, D. A., Sleeth, C. K., The curariform activity of N-methyloxyacanthine, in ‟Journal of pharmacology and experimental therapeutics", 1948, XCV, pp. 100-105.
Marsh, D. F., Sleeth, C. K., Tucker, E. B., The curariform activity of d-N-methyl-chondrodendrine and d-O-methyl-N-chondrodendrine, in ‟Journal of pharmacology and experimental therapeutics", 1948, XCIII, pp. 109-113.
Matthews, P. B. C., Mammalian muscle receptors and their central actions, London 1972.
Mendez, B., Valenzuela, P., Martial, J. A., Baxter, J. D., Cell-free synthesis of acetylcholine receptor polypeptides, in ‟Science", 1980, CCIX, pp. 695-697.
Miledi, R., Molinoff, P., Potter, L., Isolation of the cholinergic receptor protein of Torpedo electric tissue, in ‟Nature", 1971, CCXXIX, pp. 554-557.
Namba, T., Grob, D., Cholinergic receptors in skeletal muscle: isolation and properties of muscle ribonucleoprotein with affinity for d-tubocurarine and acetylcholine, and binding activities of the subneural apparatus of motor end plates with divalent metal ions, in ‟Annals of the New York Academy of Sciences", 1967, CXLIV, pp. 772-800.
Neher, E., Sakmann, B., Single-channel currents recorded from membrane of denervated frog muscle fibres, in ‟Nature", 1976, CCLX, pp. 799-801.
Neher, E., Steinbach, J. H., Local anaesthetics transiently block currents through single acetylcholine-receptor channels, in ‟Journal of physiology", 1978, CCLXXVII, pp. 153-176.
Nickerson, M., Receptor occupancy and tissue response, in ‟Nature", 1956, CLXXVIII, pp. 697-698.
Organe, G., Paton, W. D. M., Zaimis, E. J., Preliminary trials of bistrimethylammonium decane and pentane diiodide (C10 and C5) in man, in ‟Lancet", 1949, I, pp. 21-23.
Paton, W. D. M., A theory of drug action based on the rate of drug-receptor combination, in ‟Proceedings of the Royal Society" (Series B), 1961, CLIV, pp. 21-69.
Paton, W. D. M., Perry, W. L. M., The pharmacology of the toxiferines, in ‟British journal of pharmacology and chemotherapy", 1951, VI, pp. 299-310.
Paton, W. D. M., Perry, W. L. M., The relationship between depolarization and block in the cat's superior cervical ganglion, in ‟Journal of physiology", 1953, CXIX, pp. 43-57.
Paton, W. D. M., Rang, H. P., The uptake of atropine and related drugs by intestinal smooth muscle of the guinea-pig in relation to acetylcholine receptors, in ‟Proceedings of the Royal Society" (Series B), 1965, CLXIII, pp. 1-44.
Paton, W. D. M., Waud, D. R., Drug-receptor interactions at the neuromuscular junction, in Curare and curare-like agents (a cura di A. V.S de Reuck), London 1962, pp. 34-54.
Paton, W. D. M., Waud, D. R., The margin of safety of neuromuscular transmission, in ‟Journal of physiology", 1967, CXCI, pp. 59-90.
Paton, W. D. M., Zaimis, E. J., The pharmacological actions of polymethylene bistrimethylammonium salts, in ‟British journal of pharmacology and chemotherapy", 1949, IV, pp. 381-400.
Paton, W. D. M., Zaimis, E. J., The action of D-tubocurarine and of decamethonium on respiratory and other muscles in the cat, in ‟Journal of physiology", 1951, CXII, pp. 311-331.
Paton, W. D. M., Zaimis, E. J., The methonium compounds, in ‟Pharmacological reviews", 1952, IV, pp. 219-243.
Paton, W. D. M., Zar, M. A., The origin of acetylcholine released from guinea-pig intestine and longitudinal muscle strips, in ‟Journal of physiology", 1968, CXCIV, pp. 13-33.
Phillips, A. P., Synthetic curare substitutes from aliphatic dicarboxylic acid aminoethyl esters, in ‟Journal of the American Chemical Society", 1949, LXXI, p. 3264.
Prescott, F., Organe, G., Rowbotham, S., Tubocurarine chloride as an adjunct to anaesthesia, in ‟Lancet", 1946, II, pp. 80-84.
Raftery, M. A., Hunkapiller, M. W., Stracher, C. D., Hood, L. E., Acetylcholine receptor: complex of homologous subunits, in ‟Science", 1980, CCVIII, pp. 1454-1457.
Rang, H. P., Acetylcholine receptors, in ‟Quarterly review of biophysics", 1975, VII, pp. 283-399.
Rang, H. P., Ritter, J. M., On the mechanism of desensitization at cholinergic receptors, in ‟Molecular pharmacology", 1970, VI, pp. 357-382.
Rang, H. P., Ritter, J. M., The relationship between desensitization and the metaphilic effect at cholinergic receptors, in ‟Molecular pharmacology", 1970, VI, pp. 383-390.
Riker, W. F., Wescoe, W. C., The pharmacology of Flaxedil, with observations on certain analogues, in ‟Annals of the New York Academy of Sciences", 1951, LIV, pp. 373-392.
Rushton, W.A.H., Lapicque's theory of curarization, in ‟Journal of physiology", 1933, LXXVII, pp. 337-364.
Schmidt, R., Zimmerman, H., Whittaker, V. P., Metal ion content of cholinergic synaptic vesicles isolated from the electric organ of Torpedo: effect of stimulation-induced transmitter release, in ‟Neuroscience", 1980, V, pp. 625-638.
Schueler, F. W., A new group of paralyzants. I. The ‛hemicholiniums', in ‟Journal of pharmacology and experimental therapeutics", 1955, CXV, pp. 127-143.
Sheridan, R. E., Lester, H. A., Rates and equilibria at the acetylcholine receptor of Electrophorus electroplaques, in ‟Journal of general physiology", 1977, LXX, pp. 187-219.
Smith, S. M., Brown, H. O., Toman, J. E. P., Goodman, L.S., Lack of cerebral effects of d-tubocurarine, in ‟Anesthesiology", 1947, VIII, pp. 1-14.
Standaert, F. G., Riker, W. F., The consequences of cholinergic drug actions on motor nerve terminals, in ‟Annals of the New York Academy of Sciences, 1967, CXLIV, pp. 517-533.
Stephenson, R. P., A modification of the receptor theory, in ‟British journal of pharmacology", 1956, XI, pp. 379-393.
Stovner, J., Theodorsen, L., Bjelke, E., Sensitivity to dimethyltubocurarine and toxiferine with special reference to serum proteins, in ‟British journal of anaesthesia", 1972, XLIV, pp. 374-379.
Thesleff, S., The mode of neuromuscular block caused by acetylcholine, nicotine, decamethonium and succinylcholine, in ‟Acta physiologica scandinavica", 1955, XXXIV, pp. 218-231.
Unna, K. R., Pelikan, E. W., MacFarlane, D. W., Cazort, R. J., Sadove, M. S., Nelson, J. T., Drucker, P., Evaluation of curarizing drugs in man. I. Potency, duration of action and effects on vital capacity of d-tubocurarine, dimehtyl-d-tubocurarine and decamethylene-bis (trimethylammonium bromide), in ‟Journal of pharmacology and experimental therapeutics", 1950, XCVIII, pp. 318-329.
Van Maanem, E. F., The antagonism between acetylcholine and the curare alkaloids, d-tubocurarine, δ-curarine I, δ-toxiferine-II and β-erythroidine in the rectus abdominis of the frog, in ‟Journal of pharmacology and experimental therapeutics", 1950, IC, pp. 255-264.
Varney, R. F., Linegar, C.R., Holaday, H. A., The assay of curare by the rabbit ‛head-drop' method, in ‟Journal of pharmacology and experimental therapeutics", 1949, XCVII, pp. 72-83.
Walker, J., Some new curarizing substances, in ‟Journal of the Chemical Society", 1950, pp. 193-197.
Walker, M. B., Treatment of myasthenia gravis with physostigmine, in ‟Lancet", 1934, I, pp. 1200-1201.
Waser, P.G., Receptor localization by autoradiographic technics, in ‟Annals of the New York Academy of Sciences", 1967, CXLIV, pp. 737-755.
Waud, D. R., Pharmacological receptors, in ‟Pharmacological reviews", 1968, XX, pp. 49-88.
West, R., Pharmacology and therapeutics of curare and its constituents, in ‟Proceedings of the Royal Society of Medicine", 1935, XXVIII, pp. 565-578.
Wintersteiner, O., Dutcher, J. D., Curare alkaloids from Chondrodendron tomentosum, in ‟Science", 1943, XCVII, pp. 467-470.
Zaimis, E. J., The action of decamethonium on normal and denervated mammalian muscle, in ‟Journal of physiology", 1951, CXII, pp. 176-190.
Zaimis, E. J., Motor end-plate differences as a determining factor in the mode of action of neuromuscular blocking substances, in ‟Journal of physiology", 1953, CXXII, pp. 238-251.
Zimmerman, H., Vesicle recycling and transmitter release, in ‟Neuroscience", 1979, IV, pp. 1773-1804.