
Clonazione: tecniche di trasferimento nucleare
Clonazione: tecniche di trasferimento nucleare
Il termine clonazione può avere numerosi e diversi significati: può indicare la propagazione di piante per talea, la produzione di copie multiple di materiale genetico, la formazione di due, tre o quattro embrioni a partire da uno solo. In questo testo useremo il termine 'clonazione' per indicare la creazione di un embrione attraverso il trasferimento del nucleo di una cellula somatica in un uovo non fecondato e privato del nucleo. Nel caso della clonazione riproduttiva, l'embrione così ottenuto viene trasferito in un organismo ricevente affinché porti a termine il proprio sviluppo. La clonazione terapeutica, per contro, si propone di produrre embrioni umani tramite il trasferimento nucleare, al fine di ottenere cellule particolari, le cellule staminali, che potrebbero rivoluzionare la medicina contemporanea e la cura di malattie finora considerate incurabili.
Durante la fecondazione naturale, dalla fusione di uno spermatozoo e un ovocita si forma un embrione unicellulare chiamato 'zigote'. Nel corso della divisione cellulare avviene una specializzazione delle cellule ('differenziamento') che permette loro di svolgere funzioni particolari: per esempio, un neurone può trasmettere impulsi elettrici o una cellula epatica può trasformare tossine in sostanze innocue. Tutte le cellule dell'organismo (con rare eccezioni) possiedono le stesse informazioni genetiche e il differenziamento avviene tramite l'attivazione o la disattivazione di geni specifici. Mentre prima si riteneva che le modificazioni genetiche che si verificano durante il differenziamento fossero fissate così rigidamente da non potere essere cambiate, la nascita di Dolly ha dimostrato che la plasticità dello sviluppo cellulare è maggiore di quanto le precedenti ricerche non avessero lasciato supporre.
Come annunciato nel 1997 dal gruppo guidato da Ian Wilmut, la pecora Dolly è stata clonata grazie al trasferimento del nucleo di una cellula della ghiandola mammaria di una pecora di razza Finn-Dorset di sei anni in un uovo non fecondato (ovocita), privato del proprio materiale genetico tramite rimozione del nucleo. Con questo procedimento sono stati ottenuti 277 embrioni, 29 dei quali sono stati trasferiti nella fase di morula (o blastocisti) in 13 pecore adattate per portarne a termine lo sviluppo. Una delle pecore è rimasta gravida e il 5 luglio 1996 ha dato alla luce un agnello, Dolly, che possedeva lo stesso patrimonio genetico della pecora di sei anni da cui era stato prelevato il nucleo della cellula mammaria.
Fino a oggi sono state clonate diverse specie animali. Il procedimento è però tuttora soggetto a diversi problemi, in particolare per lo scarso numero di nati vivi rispetto agli embrioni ottenuti per trasferimento di nucleo: vengono persi embrioni per insufficiente sviluppo e molti feti sono espulsi per aborto. Molto frequenti sono anche i casi di morte alla nascita dell'animale clonato o successivamente a essa (talvolta anche dopo vari mesi). Con altrettanta frequenza si verificano anomalie specifiche nello sviluppo, che possono interessare, per esempio, cuore e polmoni.
La clonazione riproduttiva negli animali
Tipi di cellule e ciclo cellulare
I nuclei di cellule somatiche da trasferire sono stati prelevati da molti tipi diversi di cellule adulte, tra cui cellule epatiche, muscolari e nervose, ma con maggiore frequenza sono state impiegate quelle che circondano l'ovocita (cellule del cumulo ooforo) e i fibroblasti fetali. Ciò nonostante, l'efficienza del procedimento di clonazione è ancora sotto il 10%. Inoltre, nei topi si è osservata una notevole differenza tra le cellule dei ceppi inbred (ceppi geneticamente puri, in quanto ottenuti attraverso incroci tra consanguinei) e quelle degli ibridi: l'impiego di cellule inbred, sia staminali embrionali che somatiche, riduce in modo significativo le possibilità di successo della clonazione.
Nella maggior parte delle specie, gli ovociti subiscono un arresto naturale nella cosiddetta 'metafase della seconda divisione meiotica'. È in questa fase che essi vengono generalmente utilizzati per il trasferimento nucleare. A questo stadio è molto elevato il livello della proteina-chinasi MPF (M-phase promoting factor), che promuove il passaggio dalla fase G1 (la prima fase del ciclo cellulare) alla fase M di divisione nel ciclo cellulare. Per ottenere il normale assetto cromosomico è dunque necessario tenere in considerazione la fase del ciclo cellulare in cui si trova il nucleo donatore. Poiché il trasferimento di un nucleo nell'ovocita è seguito dalla rottura della membrana nucleare che permette la liberazione dei fattori che promuovono la replicazione del DNA, al momento del trasferimento il nucleo della cellula donatrice deve trovarsi nella fase precedente la fase di replicazione del DNA. La pecora Dolly è stata creata utilizzando cellule donatrici allo stadio G0, la fase quiescente. L'arresto in G0 viene ottenuto coltivando le cellule in presenza di una scarsa quantità di siero e quindi privandole di nutrimento in modo da renderle quiescenti, con conseguente sospensione della divisione e della crescita cellulare. Nella produzione di animali clonati sono state impiegate con successo cellule sia nella fase G0 che nella fase G1. Tuttavia, studi comparativi hanno dimostrato che le probabilità di successo sono maggiori se le cellule si trovano allo stadio G0. Per il trasferimento nucleare sono stati impiegati anche nuclei nella fase G2/M. In questo caso la citocalasina B non viene inclusa nel mezzo di coltura, in quanto in fase G2/M si ottiene la rimozione di metà dei cromosomi del nucleo. L'embrione ricostruito è quindi riportato a uno stato diploide.
In alternativa, è possibile attivare l'ovocita prima del trasferimento del nucleo permettendogli di entrare nella fase S di sintesi del DNA e inducendo in tal modo bassi livelli di attività dell'enzima MPF. Poiché in questo caso l'assetto dei cromosomi viene conservato indipendentemente dal ciclo della cellula donatrice, gli ovociti preattivati vengono denominati 'riceventi universali'. Pur essendo stati utilizzati con esito positivo nella produzione di pecore clonate da cellule embrionali ovine coltivate, i riceventi universali non hanno avuto successo nella clonazione di topi da cellule somatiche; pertanto non sono considerati ottimali per la riprogrammazione di nuclei somatici e vengono usati raramente.
Il trasferimento di un nucleo donatore direttamente in uno zigote enucleato è inefficace. Ciò nonostante, in taluni casi si è riusciti a ottenere la clonazione di maiali e di topi mediante un trasferimento in serie, ossia trasferendo il nucleo dapprima in un ovocita enucleato in metafase II, quindi in uno zigote enucleato precedentemente ottenuto mediante fecondazione in vitro. Questo procedimento ha posto le basi del metodo impiegato per creare i primi maiali clonati.
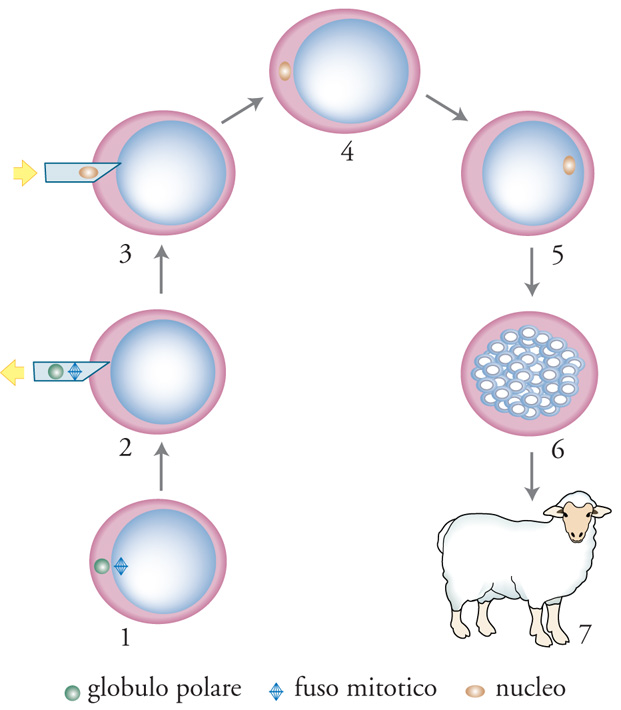
Il termine 'enucleazione' non indica la rimozione di un nucleo intatto, ma quella del primo globulo polare e del fuso mitotico. Il trasferimento del nucleo nell'ovocita enucleato si ottiene principalmente con due metodi. Nel primo, l'intera cellula donatrice viene inserita nello spazio perivitellino dell'ovocita con una micropipetta cui fa seguito l'elettrofusione (fig. 2). Questa tecnica è stata impiegata per gli ovini e i bovini. In alternativa, il nucleo viene prima prelevato dalla cellula donatrice e quindi iniettato nel citoplasma dell'ovocita per mezzo di uno speciale apparecchio piezoelettrico per microiniezione. Questa procedura è stata utilizzata per creare i primi topi clonati da cellule somatiche.
Durante la fecondazione naturale, lo stimolo che dà inizio alla divisione cellulare viene fornito dal liquido seminale, mentre nella clonazione è provocato da una scarica elettrica e/o da sostanze chimiche (quali, per es., ionomicina, ionofori del calcio, etanolo). È stato dimostrato che nei bovini e nei topi un ritardo dell'attivazione (da 15 minuti a 6 ore) dopo il trasferimento del nucleo rappresenta un evento vantaggioso ma non essenziale. Dopo l'attivazione, gli embrioni ricostruiti che sono sopravvissuti vengono generalmente coltivati in vitro per un periodo che varia da un giorno a una settimana, a seconda delle specie. Tuttavia per gli animali per i quali i protocolli per la coltura in vitro non sono ottimali, possono essere utilizzati dei riceventi 'temporanei'. In questo caso, gli embrioni ricostruiti vengono posti in agar e trasferiti nell'ovidotto per un periodo che può raggiungere i sette giorni, prima di essere rimossi e trasferiti nei riceventi definitivi per permetterne il completamento dello sviluppo.
I metodi di trasferimento del nucleo per clonare animali potranno essere impiegati in zootecnia e nella produzione di bestiame. Quando le procedure per il trasferimento del nucleo saranno diventate più efficaci ed economiche, potrà dimostrarsi utile clonare esemplari particolari, per esempio tori selezionati per la produzione di sperma, oppure mucche con una produzione eccezionale di latte o con una resistenza naturale alle malattie. Tuttavia, la clonazione riproduttiva di animali ha il suo maggior impatto in biologia e in medicina quando è associata con le biotecnologie utilizzate per ottenere organismi transgenici.
Attualmente gli animali transgenici sono ottenuti iniettando molte centinaia di copie di un determinato gene nel nucleo di embrioni unicellulari, solo pochi dei quali, peraltro, riescono a incorporare quel gene nel proprio genoma. Gli embrioni vengono poi impiantati nell'organismo ricevente per completare lo sviluppo fino alla nascita, ma solo pochi tra i nati sono transgenici e di conseguenza il procedimento ha una bassissima efficienza. In alternativa all'iniezione di geni nell'embrione è possibile modificare geneticamente le cellule in coltura e quindi impiegare le tecniche del trasferimento nucleare per creare una progenie transgenica. La combinazione di tali tecniche offre numerosi vantaggi: è possibile introdurre precisi cambiamenti genetici nel DNA della cellula donatrice (per es., modificando, aggiungendo o eliminando geni) prima che il nucleo venga trasferito e quindi verificare che nelle cellule siano intervenuti i cambiamenti desiderati prima del trasferimento all'embrione, riducendo così potenzialmente il numero degli animali necessari. Una maggiore efficienza dei metodi di trasferimento del nucleo potrebbe tradursi in un maggiore rendimento nella produzione di animali transgenici ottenuti con questo metodo.
Le applicazioni biotecnologiche per la clonazione di animali modificati geneticamente sono numerose e includono la produzione di farmaci, di organi o di tessuti animali adatti per i trapianti negli esseri umani (xenotrapianti →), nonché la creazione di modelli animali per lo studio delle malattie e la cura di alcune di esse.
Le prime pecore transgeniche create attraverso il trasferimento nucleare di cellule geneticamente modificate sono state prodotte al Roslin Institute di Edimburgo, lo stesso di Dolly, nel 1997. Ognuna di esse possedeva il gene umano per il fattore IX della coagulazione del sangue (carente nell'emofilia B), legato a un gene promotore della produzione del latte. Di conseguenza, il fattore di coagulazione viene prodotto nel latte, dal quale può essere purificato. Nel 2000 gli stessi ricercatori hanno introdotto in pecore ottenute per trasferimento nucleare il gene che codifica per l'alfa-1-antitripsina, utile per alleviare le sofferenze di pazienti affetti da enfisema e fibrosi cistica.
Oltre all'inserimento di geni umani nei genomi animali per la produzione di farmaci, anche la delezione di geni può avere applicazioni specifiche. Il topo era l'unico mammifero nel quale fosse stata praticata la delezione di geni (topi knock out), ma nel 2001 è stata clonata una pecora priva di una copia del gene per la proteina prionica (PrP). Il gene della PrP è direttamente collegato alla malattia denominata 'scrapie' nelle pecore, alla encefalopatia spongiforme nei bovini e, nell'uomo, al morbo di Creutzfeld-Jakob. Grazie all'eliminazione di ambedue le copie del gene della PrP, il bestiame clonato, privo di proteina prionica, dovrebbe essere resistente a queste malattie neurodegenerative.
La disponibilità di organi umani per il trapianto è molto limitata rispetto al numero di pazienti che necessitano di cuore, fegato, reni o polmoni nuovi. Tale carenza non diminuirà in modo significativo con le sole donazioni di organi umani, e si rende quindi necessario trovare fonti alternative. Una delle soluzioni più promettenti è lo xenotrapianto, ovvero il trasferimento in esseri umani di organi prelevati da animali; tra questi, i maiali sono la specie favorita, in quanto la loro fisiologia e le dimensioni dei loro organi sono simili a quelli dell'uomo, sono domestici e si allevano con facilità. Le scimmie sono ritenute non adatte, sia perché ragioni etiche impediscono l'uso dei Primati, sia perché farebbero aumentare il rischio di trasferire virus, trattandosi di specie relativamente vicine all'uomo. Tuttavia, l'impiego di maiali non esclude il rischio di infezioni da retrovirus endogeni (presenti all'interno del loro genoma), non solo per il paziente, ma per chiunque venga in contatto diretto o indiretto con il paziente trapiantato. Prima di procedere con gli xenotrapianti sono necessarie ulteriori ricerche sulla possibile insorgenza di infezioni da retrovirus suino, come pure test diagnostici, screening per virus, studi clinici e una valutazione globale del rischio. La marcata reazione immunologica provocata dai trapianti di tessuti suini nell'uomo rappresenta, inoltre, il maggiore impedimento a una loro applicazione clinica. Uno degli obiettivi della ricerca è quello di ottenere le modifiche genetiche necessarie a creare organi suini più compatibili con l'uomo.
In tutte le specie, con ogni tipo di cellula e indipendentemente dalla tecnica impiegata, la clonazione riproduttiva si è dimostrata una tecnica altamente inefficiente. Spesso gli embrioni vengono perduti o non riescono a impiantarsi nell'utero, i feti vengono abortiti o muoiono al momento della nascita. Tra i problemi più frequenti che insorgono durante la gestazione vi sono l'eccessivo accumulo di liquido amniotico e la crescita abnorme del feto e della placenta. Spesso il parto deve essere indotto artificialmente e possono insorgere problemi durante il travaglio e dopo la nascita. Oltre a essere sovradimensionate, le mucche e le pecore clonate presentano numerose deformità, problemi cardiovascolari e deficienze immunitarie. I problemi connessi alla gestazione e ai primi giorni di vita appaiono meno frequenti nelle capre e nei maiali clonati. Nei topi, il sovradimensionamento della prole è stato osservato nella clonazione da cellule staminali, mentre nella clonazione da cellule somatiche è la placenta a essere sovradimensionata. Inoltre, due studi sui topi condotti separatamente hanno evidenziato conseguenze abnormi della clonazione. Un confronto tra topi ottenuti rispettivamente da embrioni clonati, da fecondazione in vitro e da accoppiamenti naturali ha rivelato una significativa riduzione della longevità dei topi clonati, l'83% dei quali, a fronte del 23% dei controlli, moriva prima degli 800 giorni. La mortalità precoce dei topi clonati risultava dovuta a un basso livello di produzione di anticorpi e a varie patologie degli organi, tra cui polmoniti, necrosi del fegato e tumori. Un altro studio ha dimostrato che i topi clonati erano soggetti a obesità in età avanzata, anche se tale disfunzione non veniva trasmessa alla prole nata naturalmente da genitori clonati, il che sembrerebbe dimostrare che si tratta di un difetto di natura epigenetica.
Spermatozoi e ovociti sono cellule aploidi altamente specializzate, specificamente strutturate per fondersi al momento della fecondazione formando uno zigote unicellulare capace di portare a termine lo sviluppo. Di conseguenza, l'ovocita cui è stato tolto il nucleo deve riprogrammare il nucleo diploide della cellula somatica differenziata per trasformarlo in un nucleo embrionale funzionante. Non stupisce tanto l'inefficienza del trasferimento nucleare, quanto piuttosto il fatto che talvolta riesca a verificarsi. Le conoscenze su quanto avviene a livello molecolare negli embrioni ottenuti per fecondazione naturale sono tuttora poche e ancora più scarse sono quelle relative agli embrioni clonati, ma di recente questa area di ricerca è stata oggetto di un crescente interesse.
Affinché la riprogrammazione del nucleo abbia successo, è necessaria una riprogrammazione epigenetica del nucleo della cellula somatica (che influenza il DNA senza alterarne la sequenza nucleotidica), per permettere che la trascrizione e l'espressione dei geni avvengano in modo appropriato. Ciò può comportare cambiamenti sia nelle proteine associate al DNA, sia nella metilazione del DNA. Quest'ultimo aspetto è stato studiato nel tentativo di individuare i meccanismi molecolari responsabili dei problemi presentati da embrioni, feti e nuovi nati per clonazione, anche se altri eventi che influenzano il DNA potrebbero avere un ruolo significativo.
I processi di metilazione del DNA sono molto dinamici nel primo periodo di vita degli embrioni di mammifero, e il numero degli errori che possono essere introdotti con il trasferimento del nucleo è consistente. Gli embrioni clonati di bovini hanno profili di metilazione altamente irregolari, simili a quelli delle cellule donatrici da cui provengono e molto diversi da quelli degli embrioni normali prodotti attraverso fecondazione. Diversi studi hanno mostrato una riprogrammazione incompleta della metilazione in molti embrioni clonati: in tutti questi casi, l'ovocita non era in grado di riprogrammare adeguatamente il nucleo in modo da raggiungere il giusto livello di metilazione nell'embrione. Per contro, gli embrioni di maiale clonati non mostrano rilevanti anomalie nei profili di metilazione. Anche nei topi clonati l'analisi dei profili di metilazione tessuto-specifici ha anomalie rispetto al profilo presentato da embrioni ottenuti per fecondazione.
Tuttavia, una difettosa riprogrammazione epigenetica non impedisce il completamento dello sviluppo e anche dopo la nascita può non essere evidente nel fenotipo. Alcune ricerche hanno dimostrato che i topi clonati da cellule staminali embrionali possono sopravvivere fino all'età adulta e avere un fenotipo apparentemente normale nonostante le anomalie epigenetiche presenti nei loro genomi. Ma non tutti i meccanismi di riprogrammazione sono difettosi in tutti gli embrioni clonati; alcuni esperimenti hanno dimostrato una normale riprogrammazione relativamente alla lunghezza dei telomeri e all'inattivazione del cromosoma X.
Come ha evidenziato uno studio effettuato su embrioni bovini, un corretto sviluppo può essere ostacolato, oltre che da un'espressione impropria dei geni soggetti a imprinting (i geni che esprimono solo l'allele paterno o materno), anche dalla disregolazione dei geni non soggetti a imprinting. Di conseguenza, negli animali clonati una insufficiente riprogrammazione del genoma per quanto riguarda i geni sia soggetti che non soggetti a imprinting può essere responsabile di un carente sviluppo dell'embrione e del feto.
Cellule staminali embrionali e clonazione terapeutica
Un tipo molto specializzato di linee cellulari coltivate in vitro è costituito dalle cellule staminali embrionali (ES). Derivate da embrioni molto precoci, tali cellule sono potenzialmente in grado di rivoluzionare lo studio e la terapia di malattie degenerative e debilitanti. In condizioni di coltura appropriate, le cellule ES rimangono indifferenziate anche per lungo tempo, possono continuare a dividersi indefinitamente (e pertanto sono dette 'immortali') e, una volta trasferite nell'organismo, possono differenziarsi in qualsiasi tipo cellulare (sono, cioè, cellule 'totipotenti'). Queste caratteristiche le rendono una fonte ideale per la produzione dei diversi tipi cellulari umani destinati a sostituire cellule che si sono deteriorate o sono andate distrutte in seguito a traumi o a malattie quali il diabete, il morbo di Parkinson, o le patologie a carico del fegato, del cuore e del midollo spinale.
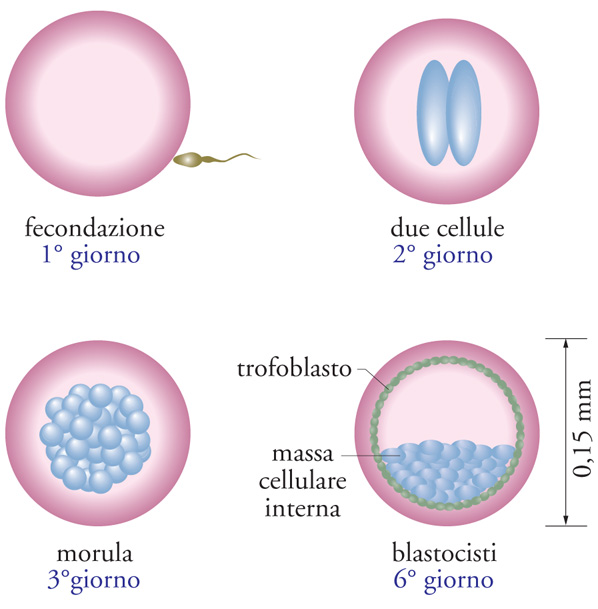
La fecondazione di un uovo da parte di uno spermatozoo dà origine a uno zigote, il quale si divide in due cellule, ciascuna delle quali si divide a sua volta dando luogo a 4, 8, 16, 32 cellule e così via, fino a formare un agglomerato sferico, chiamato morula (fig. 4); dopo questo stadio, l'embrione assume la forma di una sfera cava (blastocisti), costituita da uno strato periferico di cellule (trofoblasto), che darà origine alla placenta, e da una massa di cellule aderenti alla parete interna (ICM, Inner cell mass). È dalla ICM che vengono prelevati e poi trasferiti in colture idonee i potenziali 'precursori' delle cellule staminali, che si ottengono solo se queste cellule in coltura rimangono indifferenziate, totipotenti e immortali per lungo tempo. Le cellule ES umane sono state ottenute per la prima volta nel 1998 dal gruppo di James Thomson alla University of Wisconsin-Madison, quasi venti anni dopo la scoperta delle loro omologhe nel topo da parte di Mark J. Evans e Matthew H. Kaufman nel 1981.
Le cellule staminali embrionali umane possono essere originate da due fonti principali: embrioni umani ottenuti mediante fecondazione in vitro o mediante trasferimento nucleare. Il potenziale vantaggio di quest'ultima procedura, nota anche come 'clonazione terapeutica', risiede nel fatto che il trapianto di cellule ES ottenute trasferendo negli ovociti enucleati nuclei prelevati da cellule del paziente stesso non provoca una risposta immunitaria di rigetto. La clonazione terapeutica ha un'altra preziosa applicazione, in quanto permette la creazione di modelli cellulari in vitro di malattie. È possibile, cioè, ricostruire degli embrioni dai nuclei di un paziente affetto, per esempio, da una malattia del motoneurone, per ottenere colture di cellule ES affette dalla malattia. Cellule di questo tipo, totipotenti e in continua divisione, potrebbero rappresentare un eccellente modello della malattia per individuarne le cause genetiche, la biochimica, ecc., e fornirebbero cellule utilizzabili per saggiare e selezionare i farmaci.
La ricerca sui topi ha dimostrato che le cellule ES ‒ differenziatesi sia prima che dopo il trapianto ‒ possono integrarsi dal punto di vista funzionale e promuovere la guarigione. Per esempio, cellule staminali di topo differenziate in neuroni trapiantate in ratti con il midollo spinale danneggiato hanno restituito agli arti posteriori la capacità di sostenere pesi e migliorato le funzioni motorie. Cellule secernenti insulina derivate da cellule staminali embrionali di topo sono state trapiantate nella milza di topi diabetici provocando una normalizzazione della glicemia.
Prospettive per il futuro
Il fatto che un embrione ottenuto mediante trasferimento del nucleo di una cellula somatica possa portare a compimento lo sviluppo, dando origine a un essere vivente, costituisce un'importante dimostrazione della plasticità cellulare. Sia la terapia con cellule ES sia la clonazione per fini terapeutici aprono prospettive estremamente interessanti. La produzione di cellule ES da embrioni ottenuti per mezzo di trasferimento nucleare è appena agli inizi. La clonazione terapeutica destinata all'uomo richiede che si sviluppi un embrione ai primi stadi di vita, e gli esperimenti sul trasferimento del nucleo nella clonazione riproduttiva negli animali hanno evidenziato i limiti e i successi di questo metodo. Un'ottimizzazione delle tecniche attuali potrebbe rendere un po' più efficace il trasferimento nucleare, ma progressi significativi potranno ottenersi attraverso la comprensione degli eventi che si verificano nel corso di un'efficace riprogrammazione nucleare e cercando di aiutare quest'ultima dopo il trasferimento, per esempio con l'uso di complessi per rimodellare o con la rimozione della metilazione del DNA somatico prima o dopo il trasferimento. Il progresso delle conoscenze sui meccanismi della riprogrammazione nucleare dovrebbe portare allo sviluppo di tecniche che permettano il transdifferenziamento di un tipo di cellula somatica in un altro, senza dovere ricorrere alle cellule ES per la terapia di sostituzione cellulare.
Bibliografia
Evans, Kaufman 1981: Evans, Mark J. - Kaufman, Matthew H., Establishment in culture of pluripotential cells from mouse embryos, "Nature", 292, 1981, pp. 154-156.
Humphreys 2001: Humphreys, Davina e altri, Epigenetic instability in ES cells and cloned mice, "Science", 293, 2001, pp. 95-97.
McDonald 1999: McDonald, John W. e altri, Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord, "Nature medicine", 5, 1999, pp. 1410-1412.
Schnieke 1997: Schnieke, Angelika E. e altri, Human factor IX transgenic sheep produced by transfer of nuclei from transfected fetal fibroblasts, "Science", 278, 1997, pp. 2130-2133.
Thomson 1998: Thomson, Janus A. e altri, Embryonic stem cell lines derived from human blastocysts, "Science", 282, 1998, pp. 1145-1147.
Wilmut 1997: Wilmut, Ian e altri, Viable offspring derived from fetal and adult mammalian cells, "Nature", 385, 1997, pp. 810-813.