
Chimica bioinorganica
Chimica bioinorganica
SOMMARIO: 1. Premesse storiche. 2. La chimica inorganica della vita. 3. I metalli negli esseri viventi. 4. Metalloproteine. 5. Metalloenzimi. 6. Metalli e acidi nucleici. 7. I metalli in medicina. 8. Metodi di indagine. 9. Prospettive della disciplina. □ Bibliografia.
1. Premesse storiche
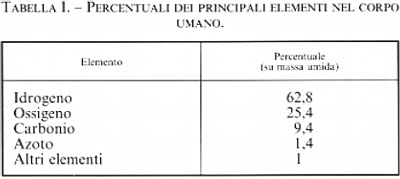
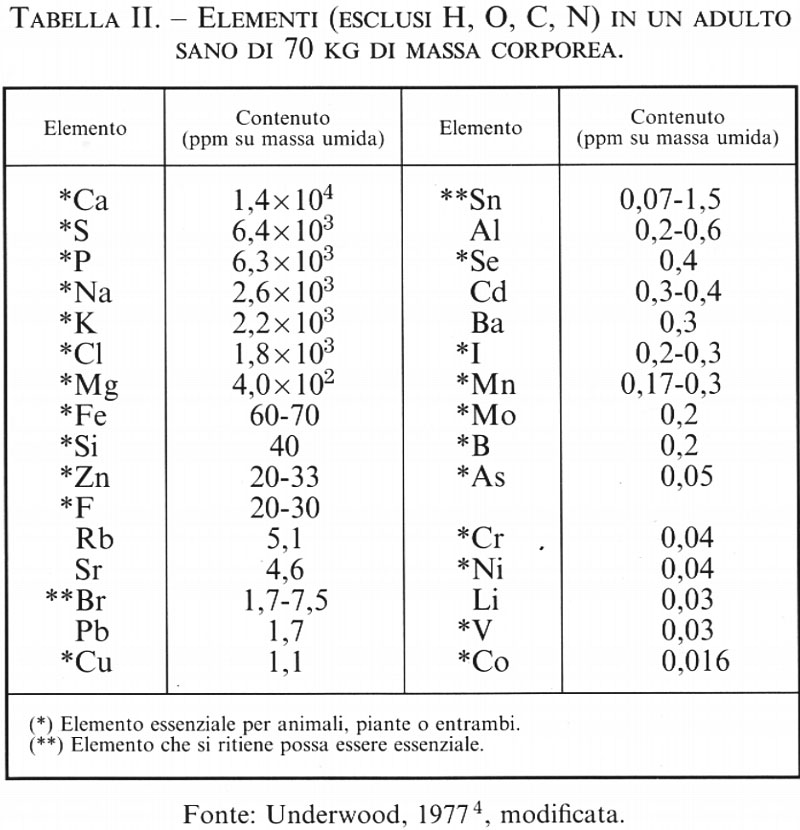
La chimica bioinorganica è quella branca della chimica che studia la funzione esercitata dagli elementi inorganici classici nella chimica degli esseri viventi, o biochimica; si tratta, come vedremo, di una funzione fondamentale, non ostante la biochimica sia storicamente legata soprattutto alla chimica organica. Il fatto che l'aggettivo ‛bioinorganica' richieda, a tutt'oggi, una giustificazione, testimonia quanto lentamente i progressi delle scienze chimiche si diffondano al di fuori della cerchia degli specialisti ed entrino a far parte della cultura generale. Giova ricordare a questo proposito che la distinzione sei-settecentesca fra ‛chimica minerale', ‛chimica vegetale' e ‛chimica animale' fu superata già ai primi dell'Ottocento, per lasciare il posto ai due regni della ‛chimica inorganica' e della ‛chimica organica'; e che la teoria della vis vitalis che avrebbe dovuto caratterizzare tutte le sostanze di quest'ultimo regno, e quindi precluderne l'accesso al chimico sintetico, veniva già fortemente scossa dalla sintesi dell'urea realizzata da Wöhler nel 1828, e definitivamente seppellita dalle sintesi di Berthelot a cavallo degli anni sessanta e settanta dello stesso secolo. Da più di un secolo, quindi, la chimica organica ha perduto ogni legame con la chimica della vita ed è rimasta solo ‛la chimica dei composti del carbonio' (cfr. A. Kekulé, Lehrbuch der organischen Chemie oder der Chemie der Kohlenstoffverbindungen, 1867). Sopravvive ed è diffusa, però, la convinzione che la chimica degli esseri viventi sia basata sulla chimica organica. Questa convinzione ha per la verità un fondamento quantitativo: infatti, carbonio, idrogeno, ossigeno e azoto - i componenti principali delle sostanze organiche - rappresentano il 99% della massa umida del corpo umano (v. tab. I); non ha però fondamento qualitativo: il restante 1% è infatti costituito in gran parte da sodio, potassio, calcio, magnesio, fosforo, zolfo e cloro, che sono elementi essenziali, cioè necessari per la vita (v. Underwood, 19774), talché la mancanza di anche uno solo di essi porta alla morte dell'organismo. Negli organismi viventi è presente in quantità ancora minori un'altra ventina di elementi, diversi dei quali ugualmente essenziali. Il contenuto dei vari elementi in un adulto sano di 70 kg di massa corporea è riportato nella tab. II.
La fig. 1 riporta la tavola periodica degli elementi, in cui sono evidenziati quelli presenti negli esseri viventi, e quelli essenziali fra questi ultimi (v. Frausto da Silva e Williams, 1991). Di molti elementi ‛inorganici' è nota l'essenzialità da più di un secolo; un discreto numero di altri elementi, specie metalli di transizione, sono però stati riconosciuti come essenziali solo da alcuni decenni. Solo dagli anni cinquanta si è cominciato a capire che l'essenzialità di molti degli elementi di transizione deriva dalla funzione catalitica che essi svolgono quando sono incorporati in matrici proteiche, e che una parte significativa delle attività catalitiche negli esseri viventi è svolta da metalloenzimi. L'esplosione degli studi in questo campo ha finalmente imposto la necessità di dare un nome alla nuova disciplina. Il nome che si è affermato è ‛chimica bioinorganica', insieme ai meno diffusi ‛biochimica inorganica' e ‛chimica inorganica biologica' (v. Bertini e altri, 1994; v. Lippard e Berg, 1994; v. Cowan, 1993; v. Hay, 1987; v. Hughes, 19812; v. Ochiai, 1977).
2. La chimica inorganica della vita
I termini ‛minerali' e ‛sostanze organiche' evocano immagini molto diverse fra loro: solidi puri e cristallini i primi, materiali impuri e amorfi le altre. Queste immagini, tuttavia, rappresentano una eredità dei secoli scorsi: le sostanze organiche, per definizione di sostanza chimica, sono anch'esse composti puri, spesso ottenibili anche allo stato cristallino. Peraltro i composti organici, così come sono organizzati nella materia vivente, difficilmente potrebbero dar luogo a strutture con la robustezza e la solidità necessarie a costituire l'esoscheletro di un mollusco, o i denti e l'endoscheletro di un mammifero. Strutture di questo tipo sono realizzate in natura con i cosiddetti ‛biominerali' (v. Crick, 1989; v. Mann e altri, 1989; v. Westbroeck e De Jong, 1983), cioè sostanze inorganiche come carbonato di calcio, silice, fosfato di calcio, che si sviluppano in aggregati cristallini analoghi a quelli dei minerali sulla crosta terrestre, ma il cui accrescimento è regolato dal metabolismo dell'essere vivente.
I principali solidi inorganici presenti nei sistemi biologici sono elencati in tab. III. Come si vede, la funzione dei biominerali non è solo strutturale. Piccoli cristalli di calcite si trovano in sospensione acquosa adagiati alla base dei canali semicircolari nel labirinto dell'orecchio interno a livello delle macule e delle creste ampollari. Il loro contatto con queste ultime fornisce informazioni al cervello sulla direzione delle linee di forza del campo gravitazionale terrestre e permette il mantenimento della posizione eretta. Cristalli di magnetite all'interno di certi batteri forniscono informazioni sulla direzione delle linee di forza del campo magnetico terrestre, e conferiscono un certo ‛senso dell'orientamento' a questi organismi non dotati di organi sensoriali sofisticati. Cristalli di idrossido di ferro vengono fatti crescere all'interno di strutture proteiche (ferritina, v. cap. 4) per immagazzinare il ferro in forma insolubile fintantoché l'elemento non deve essere utilizzato per costituire il centro attivo di metalloproteine e metalloenzimi (v. capp. 4 e 5).
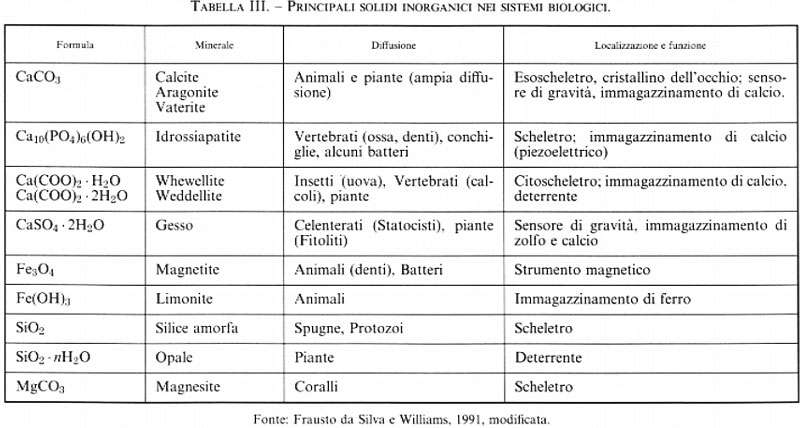
Lo studio dei biominerali è una delle aree più classiche della bioinorganica, e anzi storicamente la precede. Grazie alle moderne tecniche di diffrazione dei raggi X (v. cap. 8) è ormai possibile analizzare la morfologia dei biominerali nei minimi dettagli. Rimane invece molto da capire sui meccanismi attraverso i quali l'organismo guida e controlla l'accrescimento dei biominerali. Si tratta di meccanismi molto complicati, come si può immaginare dalla complessità delle strutture cristalline risultanti e dalla loro varietà. La comprensione del controllo metabolico della formazione dei biominerali è indispensabile per elaborare le strategie di intervento nelle malattie a essi correlate.
3. I metalli negli esseri viventi
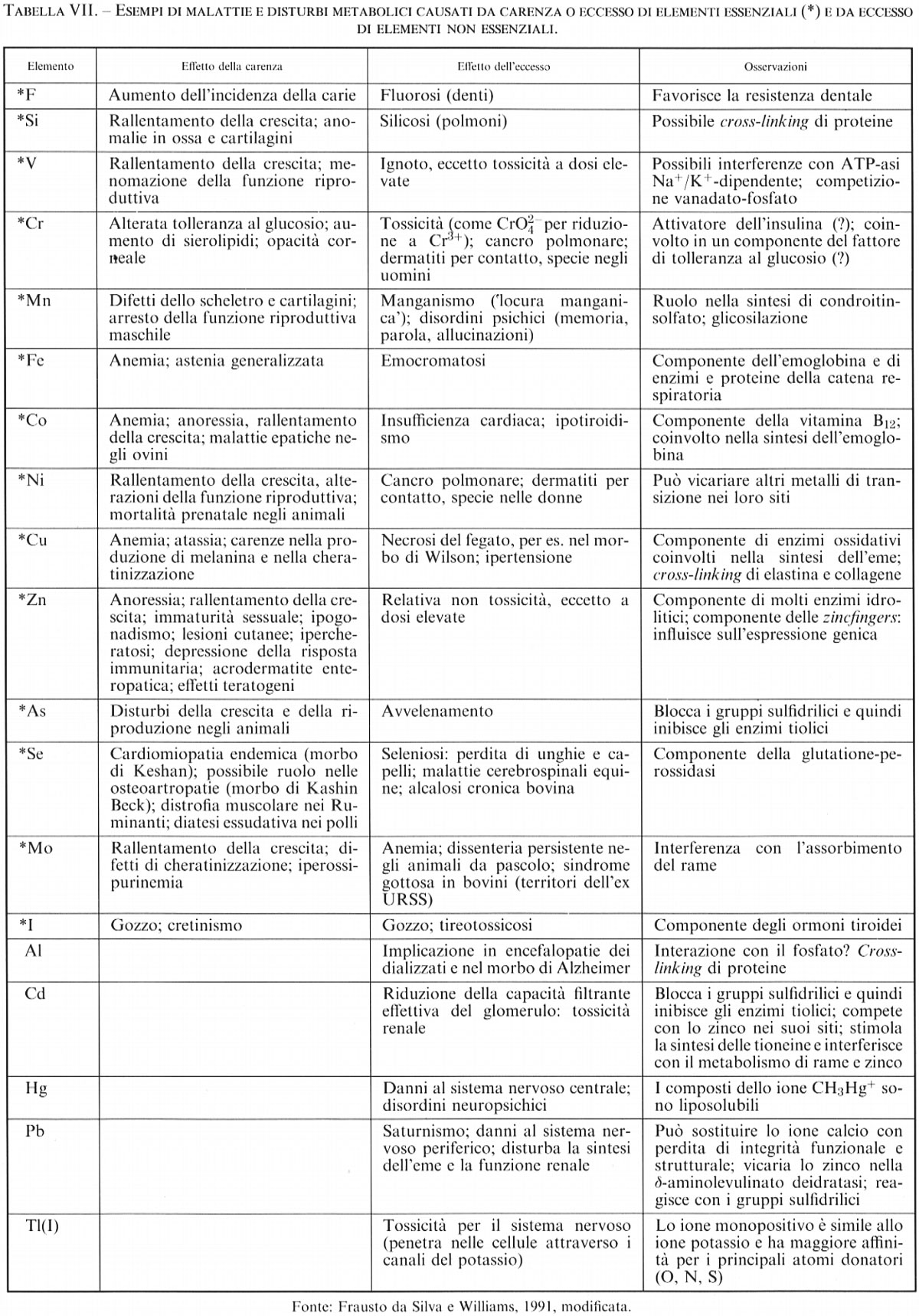
Molti degli elementi elencati nella tab. II e nella fig. 1 sono elementi metallici o semimetallici alcuni dei quali sono notoriamente tossici, come il piombo (si pensi alla progressiva messa al bando del piombo tetraetile dai combustibili per autotrazione), il rame (il solfato di rame è largamente usato in agricoltura perché tossico verso molti organismi nocivi per le piante) o l'arsenico; eppure il rame e l'arsenico sono addirittura essenziali. Questo apparente paradosso si spiega molto semplicemente con le cosiddette ‛curve di risposta alla dose' (v. Hamer e Winge, 1989). In questi grafici - nei quali si riporta in ascisse la concentrazione (o il dosaggio) dell'elemento e in ordinate la risposta dell'organismo - si indica con 100 le condizioni ottimali di salute o di crescita e con 0 la morte dell'organismo. Ad alta concentrazione la curva della risposta scende a 0 per qualsiasi elemento, cioè una concentrazione eccessiva provoca sempre la morte. Per un elemento essenziale la curva parte da 0 a concentrazione 0 (cioè anche la mancanza dell'elemento provoca la morte), mentre per un elemento non essenziale la curva parte da 100 (cioè la mancanza dell'elemento non ha influenza sulla salute e la crescita). Si capisce quindi che anche elementi non essenziali possono essere normalmente presenti nell'organismo, se la loro concentrazione è abbastanza bassa da mantenere una risposta vicina a 100, mentre gli elementi essenziali devono mantenersi entro l'intervallo di concentrazione che dà una risposta vicina a 100. Per alcuni di questi elementi l'intervallo ottimale è abbastanza ristretto. Si noti anche che la massima concentrazione tollerata di un elemento non essenziale (ad esempio, il rubidio) può essere molto più alta di quella di un elemento essenziale (ad esempio, l'arsenico).
La chimica degli elementi metallici negli esseri viventi è essenzialmente la chimica delle loro soluzioni acquose, perché l'acqua è la sostanza (inorganica) più abbondante negli esseri viventi (circa il 70% in peso nei Mammiferi). Gli elementi metallici si trovano sempre sotto forma di ioni positivi, o ‛cationi', coordinati da altri atomi che formano con essi dei legami covalenti più o meno polari. A seconda dell'elemento, gli atomi preferiti come ‛leganti' possono essere atomi di ossigeno, azoto o zolfo (v. tab. IV): se l'elemento preferisce essere coordinato da atomi di ossigeno, spesso questo si trova sotto forma di ione idrato, o ‛aquaione', cioè legato esclusivamente da molecole d'acqua; altrimenti, l'elemento preferirà interagire con quelle sostanze organiche (amminoacidi, proteine, acidi nucleici, gruppi prostetici di varia natura, ecc.) che forniscono atomi leganti più forti dell'ossigeno dell'acqua. La forza del legame di coordinazione aumenta con la carica dello ione:

M+ 〈 M2+ 〈 M3+,
e quest'ultima dipende a sua volta dal numero di ossidazione (o stato di ossidazione) preferito dall'elemento. La diversificazione dei ruoli biologici dei vari elementi metallici (v. tab. V) trae origine essenzialmente dalla preferenza dell'elemento per certi atomi donatori, dalla forza dei legami di coordinazione che si formano, e dalla possibilità per l'elemento di esistere in più stati di ossidazione. Gli ioni alcalini, come Na+ e K+, data la loro scarsa tendenza a formare composti di coordinazione stabili, sono principalmente usati come trasportatori di carica elettrica per regolare il potenziale elettrochimico attraverso le membrane cellulari (v. trasporto attraverso membrane biologiche, vol. VII). Gli ioni Mg2+ e Ca2+, che possono interagire con leganti organici formando composti discretamente stabili, sono spesso usati come attivatori, cioè per modificare temporaneamente la struttura, ad esempio, di una proteina, e per far sì che essa possa svolgere un certo compito quando richiesto. Gli ioni Mg2+ e Ca2+ possono anche avere un ruolo strutturale, possono cioè contribuire a determinare e a consolidare la struttura del composto organico (in genere della proteina) a cui sono legati; in questo caso ne diventano costituenti permanenti. Tutti gli altri metalli della tab. V si trovano come costituenti permanenti di proteine, e ne caratterizzano la funzione, come si vedrà nei prossimi due capitoli.

4. Metalloproteine
Le proteine sono fra i principali costituenti della materia vivente (v. proteine, vol. V). Esse sono composte da unità monomeriche elementari, gli amminoacidi, polimerizzati per eliminazione di una molecola d'acqua e formazione di un legame ammidico (detto ‛legame peptidico'). Gli atomi di azoto e di ossigeno del legame peptidico possono a volte interagire con gli ioni metallici, ma più frequentemente l'interazione delle proteine con gli ioni metallici è a carico delle catene laterali degli amminoacidi: molte di queste contengono gruppi di atomi particolarmente adatti a funzionare come ligandi di ioni metallici. Nella tab. VI sono riportati i più comuni gruppi leganti con le relative costanti di dissociazione acida. Infatti, gli atomi che coordinano gli ioni metallici devono usare una coppia di elettroni solitaria o ‛di non-legame', e quest'ultima può anche legare uno ione idrogeno invece di uno ione metallico. Al diminuire del pH, ovvero all'aumentare della concentrazione dello ione idrogeno, questo può competere con lo ione metallico causandone la dissociazione dalla proteina che lo lega. Più alto è il pKa , ovvero più alta è l'affinità dell'atomo legante per lo ione idrogeno, più questo effetto può essere rilevante. I gruppi più importanti come ligandi sono i gruppi carbossilato, imidazolo, fenolato e tiolato (v. Bertini e altri, 1983 e 1988; v. Bertini e Luchinat, 1985).

Le tre funzioni principali degli ioni metallici nelle metalloproteine sono quelle di mantenere la struttura di queste ultime (ruolo strutturale), di legare l'ossigeno per la funzione respiratoria (ruolo di trasporto di ossigeno), e di cambiare numero di ossidazione accettando e cedendo elettroni (ruolo di trasporto elettronico; v. Bertini e altri, 1983). I metalli con ruolo strutturale sono il magnesio, il calcio - come già accennato - e lo zinco. Gli altri due ruoli sono ricoperti quasi esclusivamente da metalli di transizione che presentano un riempimento incompleto degli orbitali d. I composti di coordinazione relativi hanno spesso la proprietà di poter variare facilmente il numero di coordinazione, vale a dire di poter legare temporaneamente un ulteriore ligando oltre a quelli forniti dalla proteina, e inoltre di poter cambiare il numero di ossidazione. Al ruolo di trasporto di ossigeno o di elettroni è sempre associato, ovviamente, anche un ruolo strutturale, perché la presenza di uno ione metallico legato alle catene laterali conferisce comunque maggiore stabilità alla struttura proteica.
La fig. 4 mostra in modo schematico i principali gruppi metallo-leganti responsabili del trasporto di ossigeno: essi si trovano rispettivamente nell'emoglobina e nella mioglobina (Vertebrati: v. Antonini e Brunori, 1971), nell'emocianina (Molluschi e Artropodi: v. Sykes, 1985) e nell'emoeritrina (Invertebrati marini: v. Wilkins e Wilkins, 1987). L'emoglobina e la mioglobina usano il gruppo prostetico eme, che a sua volta lega uno ione Fe2+, mentre l'emocianina usa una coppia di ioni Cu+ e l'emoeritrina una coppia di ioni Fe2+, in entrambi i casi coordinati da catene laterali imidazoliche dell'amminoacido istidina. Queste strutture piuttosto complicate si sono evolute per risolvere il problema di trasportare l'ossigeno, che è il responsabile di tutte le reazioni di ossidazione negli esseri viventi aerobici, senza farlo reagire prima del tempo (v. Kurtz, 1990). Gli ioni metallici (ferro o rame) sono indispensabili per questo scopo, perché possono cambiare il proprio stato di ossidazione (da + 2 a + 3 il ferro, e da + 1 a + 2 il rame) riducendo temporaneamente l'ossigeno a superossido o perossido per aumentarne le capacità leganti e quindi impedirgli di dissociarsi prima di essere giunto al luogo di utilizzo. L'ossigeno viene poi liberato all'interfaccia della membrana mitocondriale, dove viene ridotto ad acqua (v. cap. 5) ricevendo gli elettroni da una serie di proteine di trasporto che formano la cosiddetta catena respiratoria. Molte di queste proteine sono anch'esse metalloproteine.
Molte metalloproteine di trasporto elettronico sfruttano la stessa struttura ferro-eme già vista per l'emoglobina e la mioglobina, con la differenza che le due posizioni di coordinazione assiale sono entrambe occupate da ligandi della proteina stessa, e quindi il ferro non può interagire con ligandi esterni come l'ossigeno. Può però cambiare il proprio stato di ossidazione, per esempio acquistando un elettrone da una proteina per passare da Fe3+ a Fe2+ e cedendo l'elettrone a un'altra proteina in un diverso comparto biologico per tornare a Fe3+. Oltre alle emoproteine, altre tipiche metalloproteine di trasporto elettronico sono le cosiddette proteine a ferro-zolfo (v. Cammack e Sykes, 1992; v. Beinert, 1990). La fig. 5 mostra la struttura di una tipica proteina a ferro-zolfo (v. Banci e altri, 1994). La proteina lega degli atomi di ferro (quattro nell'esempio in figura) tramite i gruppi tiolati delle catene laterali dell'amminoacido cisteina; gli atomi di ferro sono ulteriormente legati da ioni solfuro (anch'essi quattro nell'esempio in figura) per formare un aggregato, o cluster (v. Berg e Holm, 1982). Il numero di ossidazione di tutto il cluster Fe4S4 può variare di una unità, permettendo alla proteina di esercitare la sua funzione di trasporto elettronico. Le proteine a ferro-zolfo si trovano in un gran numero di organismi, dalle forme di vita primordiali precedenti i Batteri (Archaea) all'uomo. Alcune recenti teorie sull'evoluzione della vita sulla Terra ipotizzano addirittura che i primi sistemi chimici dotati della capacità di autoriprodursi siano originati dall'interazione di alcune semplici sostanze organiche con la pirite (FeS2) per formare dei clusters ferro-zolfo (v. Wächtershäuser, 1994).
Si potrebbe a buon diritto definire una quarta classe di metalloproteine, cioè quella costituita dalle proteine adibite al trasporto o all'immagazzinamento degli ioni metallici stessi (v. Theil e Raymond, 1994). La disponibilità biologica (o ‛biodisponibilità') degli ioni metallici per l'organismo dipende da molti fattori, fra i quali la relativa abbondanza nell'ambiente esterno e nel cibo, ma anche la speciazione, ovvero la forma chimica sotto la quale gli ioni metallici si trovano. Ad esempio, lo ione Fe3+ è abbondante, ma è pochissimo solubile al pH fisiologico; occorre quindi una proteina (la transferrina; v. Baker e altri, 1991) che sia capace di legarlo con alta affinità per solubilizzarlo nel plasma e trasportarlo ai siti di utilizzo (sintesi dell'emoglobina, ecc.) o di accumulo (nella ferritina, v. Theil, 1990). Nei Batteri il trasporto del ferro è affidato a piccole molecole dette siderofori, anch'esse dotate di alta affinità per il ferro, sebbene non così alta come quella della transferrina. Una varietà di quest'ultima, detta lattoferrina, è presente nel latte dei Mammiferi con funzioni batteriostatiche: la sua presenza riduce drasticamente la biodisponibilità del ferro ai batteri occasionalmente presenti e ne impedisce la crescita. Il trasporto del rame è prevalentemente a carico della ceruloplasmina, mentre il trasporto dello zinco, e probabilmente l'accumulo del cadmio (che è un metallo non essenziale), è a carico delle tioneine. È stato rilevato che la biosintesi di queste ultime è stimolata nei casi di avvelenamento da cadmio o da altri metalli pesanti.
5. Metalloenzimi
L'importanza degli elementi metallici nei sistemi biologici è esemplificata ancora meglio dai metalloenzimi (v. Bertini e altri, 1983; v. Dennard e Williams, 1966). Gli enzimi sono catalizzatori biologici: un catalizzatore è, per definizione, una sostanza chimica che accelera il procedere di una reazione chimica verso l'equilibrio finale senza essere consumata nella reazione stessa. La vita non potrebbe esistere senza gli enzimi perché, alle temperature ottimali per la vita degli organismi, molte delle reazioni necessarie al metabolismo procederebbero a velocità troppo bassa. Sono noti migliaia di enzimi suddivisi, a seconda del tipo di reazione che catalizzano, in sei classi: ossidoreduttasi, transferasi, idrolasi, liasi, isomerasi e ligasi (v. catalisi enzimatica, vol. I). In molti casi, la catalisi richiede la presenza di uno ione metallico: quasi la metà degli enzimi finora scoperti sono metalloenzimi (v. enzimi, vol. II).
Un importantissimo metalloenzima della classe delle ossidoreduttasi è la citocromo-c-ossidasi (v. Malmström, 1990). Come si è già accennato, la riduzione dell'ossigeno ad acqua, che avviene alla superficie della membrana mitocondriale, è il passaggio chiave per l'utilizzazione della capacità ossidante dell'ossigeno nel metabolismo degli aerobi:
O2 + 4H+ + 4 e- → 2H2O .
Questa è la reazione finale della catena respiratoria. La riduzione della molecola di O2 ad acqua richiede quattro elettroni che, quando sono ceduti uno alla volta alla molecola, producono intermedi come il superossido, il perossido o il radicale ossidrile che sono molto tossici se liberati nei fluidi biologici. La natura si è cautelata affidando la riduzione dell'ossigeno alla citocromo-c-ossidasi, un enzima inglobato nella membrana mitocondriale; l'enzima ha un sito di attacco per la metalloproteina di trasporto elettronico (citocromo c) all'interno del mitocondrio, e un sito di attacco per l'ossigeno all'esterno. L'enzima contiene quattro centri metallici, costituiti da due gruppi ferro-eme, uno ione rame, e due ioni rame raggruppati in un cluster (v. Antholine e altri, 1992), che possono acquistare un totale di quattro elettroni da quattro molecole di citocromo c. L'enzima, quando è totalmente ridotto, può legare l'ossigeno e ridurlo completamente ad acqua cedendogli in rapida sequenza i quattro elettroni, senza liberare nessuno degli intermedi tossici.
Un'altra classe in cui i metalloenzimi sono particolarmente numerosi è quella delle idrolasi. Spesso le idrolasi contengono zinco, un metallo che non può svolgere funzioni di ossidoriduzione perché ha gli orbitali d completamente riempiti e un solo numero di ossidazione accessibile, il 2 + (v. Christianson, 1991; v. Bertini e altri, 1986; v. Bertini e Luchinat, 1994). La completezza del sottostrato d conferisce però allo zinco una grande flessibilità nel numero di coordinazione, che può passare da 4 a 5 e a 6 senza grandi variazioni di energia del sistema. Lo zinco è quindi molto adatto a legare transitoriamente il substrato che deve subire la reazione e, nel caso delle idrolasi, ad attivare per coordinazione la molecola d'acqua che deve attaccare il substrato stesso (v. Auld e altri, 1989). La fig. 8 mostra uno schema del meccanismo catalitico della carbossipeptidasi (v. Bertini e Luchinat, 1994; v. Christianson e Lipscomb, 1989): il substrato è un peptide, cioè un poliamminoacido, e l'enzima catalizza l'idrolisi del legame peptidico dell'amminoacido carbossi-terminale:
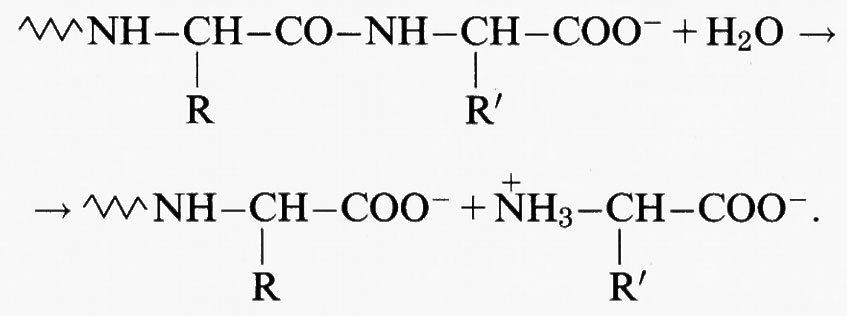
Il peptide si lega in una cavità della molecola dell'enzima, posizionandosi in modo tale che l'atomo di ossigeno della molecola d'acqua coordinata allo zinco possa formare un legame con il carbonio del legame peptidico e provocare la rottura del legame C-N. Il gruppo carbossilato della catena laterale di un amminoacido adiacente serve da accettore dello ione idrogeno della molecola d'acqua, che diventa uno ione ossidrile e risulta così attivata per l'attacco al carbonio peptidico.
Metalloenzimi fra i più interessanti e importanti per la loro funzione sono l'enzima nitrogenasi e gli enzimi coinvolti nel ‛fotosistema II'. Il primo catalizza la reazione di fissazione dell'azoto atmosferico, molecola particolarmente inerte, in alcuni batteri (detti appunto azoto-fissatori):
N2 + 10H+ + 8e- → 2NH4+ + H2.
La reazione di fissazione dell'azoto, con produzione di ammonio o ammoniaca, è fondamentale per l'immissione dell'elemento nella biosfera, ed è anche molto importante dal punto di vista industriale, perché pressoché tutti i composti chimici che contengono azoto vengono sintetizzati a partire dall'ammoniaca; tuttavia è molto difficile da realizzare proprio per l'inerzia della molecola di N2. Il meccanismo della nitrogenasi non è del tutto chiarito, ma è ormai evidente, anche grazie alla recente risoluzione della struttura ai raggi X (v. cap. 8) della parte del sistema enzimatico contenente il cosiddetto cofattore ferro-molibdeno, che gli ioni metallici sono essenziali. La fig. 9 mostra il cluster metallico ferro-molibdeno come risulta dalla risoluzione della struttura ai raggi X (v. Kim e Rees, 1992). Sembra che sia cruciale, perché la reazione proceda a velocità accettabile, che il sistema sia in grado di fornire in rapida sequenza i sei elettroni che servono per ridurre la molecola di N2 (o gli otto elettroni se la reazione coinvolge anche, come sembra, riduzione di ioni idrogeno). Riduzioni parziali dell'azoto sono molto sfavorite dal punto di vista energetico e producono composti instabili, come N2H2 o N2H4; proprio a questo è dovuta l'inerzia chimica della molecola.
Altrettanto affascinante è la sfida posta agli scienziati dalla comprensione del sistema fotosintetico, che è responsabile della produzione di ossigeno da parte delle piante e di alcuni batteri (v. fotosintesi, vol. III). La luce è trasformata in energia chimica durante il ciclo fotosintetico nei cosiddetti ‛centri di reazione', che contengono ioni magnesio legati a gruppi eme. Lo schema di uno di questi centri, che fanno sempre parte del fotosistema II, è mostrato in fig. 11 (v. Okamura e Feher, 1992). L'assorbimento di fotoni si traduce in ossidazione di un altro componente del fotosistema II, il centro di svolgimento di ossigeno. Di quest'ultimo fanno parte quattro ioni manganese (v. Renger, 1988), che sono quindi ossidati allo stato di ossidazione 3 + dall'azione indiretta (tramite i centri di reazione già visti) della radiazione solare ultravioletta. La struttura del complesso è ancora ignota, e costituisce una delle grandi sfide tuttora aperte per la chimica bioinorganica. Probabilmente gli ioni manganese passano dallo stato di ossidazione 3 + allo stato 2 +, sottraendo all'acqua i quattro elettroni necessari per la formazione di ossigeno, in una reazione che è la reazione inversa di quella catalizzata dalla citocromo-c-ossidasi:
2H2O → O2 + 4H+ + 4e-.
Si chiude così il ciclo: la radiazione solare fornisce l'energia necessaria per ossidare gli ioni manganese al numero di ossidazione 3 +; questi sono quindi in grado di ossidare l'acqua a ossigeno, processo che sarebbe sfavorito dal punto di vista energetico. L'ossigeno viene utilizzato dagli organismi aerobi per liberarsi degli elettroni che si producono nel ciclo metabolico, ed è così nuovamente ridotto ad acqua. Il ciclo metabolico produce, fra gli altri, composti organici ad alta energia come l'ATP (adenosintrifosfato). L'energia solare è così immagazzinata negli esseri viventi in una forma conveniente per essere utilizzata quando opportuno.
6. Metalli e acidi nucleici
Gli ioni metallici possono interagire direttamente con gli acidi nucleici (RNA e DNA), che sono polimeri aventi come unità monomeriche costituenti, rispettivamente, ribonucleotidi e desossiribonucleotidi, polimerizzati tramite la formazione di legami fosfodiesterei. Gli acidi nucleici assumono le ben note strutture a doppia elica, a loro volta organizzate in superstrutture in cui le doppie eliche formano anse o si avvolgono ulteriormente formando doppie eliche di doppie eliche (v. acidi nucleici, vol. I). Alcune delle interazioni dei metalli con gli acidi nucleici sono piuttosto aspecifiche, come ad esempio la stabilizzazione di alcune superstrutture da parte di ioni Na+ e Mg2+ attraverso interazioni elettrostatiche che schermano i gruppi fosfato, carichi negativamente, gli uni dagli altri. Si è però scoperto recentemente che gli ioni Mg2+ possono anche funzionare da attivatori specifici di certe attività catalitiche dell'RNA, e che gli ioni K+ sembrano stabilizzare in particolare la struttura delle unità terminali delle doppie eliche del DNA alla fine dei cromosomi (v. Lippard e Berg, 1994; v. Draper, 1992; v. Tullius, 1989).
Un'altra relazione importante fra ioni metallici e acidi nucleici è data dal ruolo che molte proteine contenenti zinco svolgono nell'esplicare le loro funzioni di attivatori o regolatori del DNA. In particolare, una classe di zinco-proteine scoperte di recente, le cosiddette zincfingers (dita di zinco), sembra utilizzare un certo numero di ioni zinco per creare delle strutture peptidiche che si protendono all'infuori della proteina, come delle dita (v. Berg, 1990), che vengono usate per intercalarsi tra le spire della doppia elica del DNA. In questo caso il metallo sembra indispensabile per costringere la catena peptidica ad assumere la struttura adatta per l'interazione con il DNA.
7. I metalli in medicina
Come si è già accennato nei capp. 1-3, in un essere umano in buona salute è presente una grande varietà di elementi chimici, essenziali e non essenziali. Le curve di risposta alla dose (v. cap. 3) forniscono informazioni preziose dal punto di vista nutrizionale (per esempio per evitare scompensi nelle diete, per sopperire alle carenze o rimediare agli eccessi di uno o più elementi - problemi endemici in certe popolazioni -, ecc.), ma rappresentano solo il primo passo verso una conoscenza approfondita del ruolo che ciascuno di questi elementi riveste nel metabolismo. Alcune informazioni qualitative sono riportate in tab. VII.
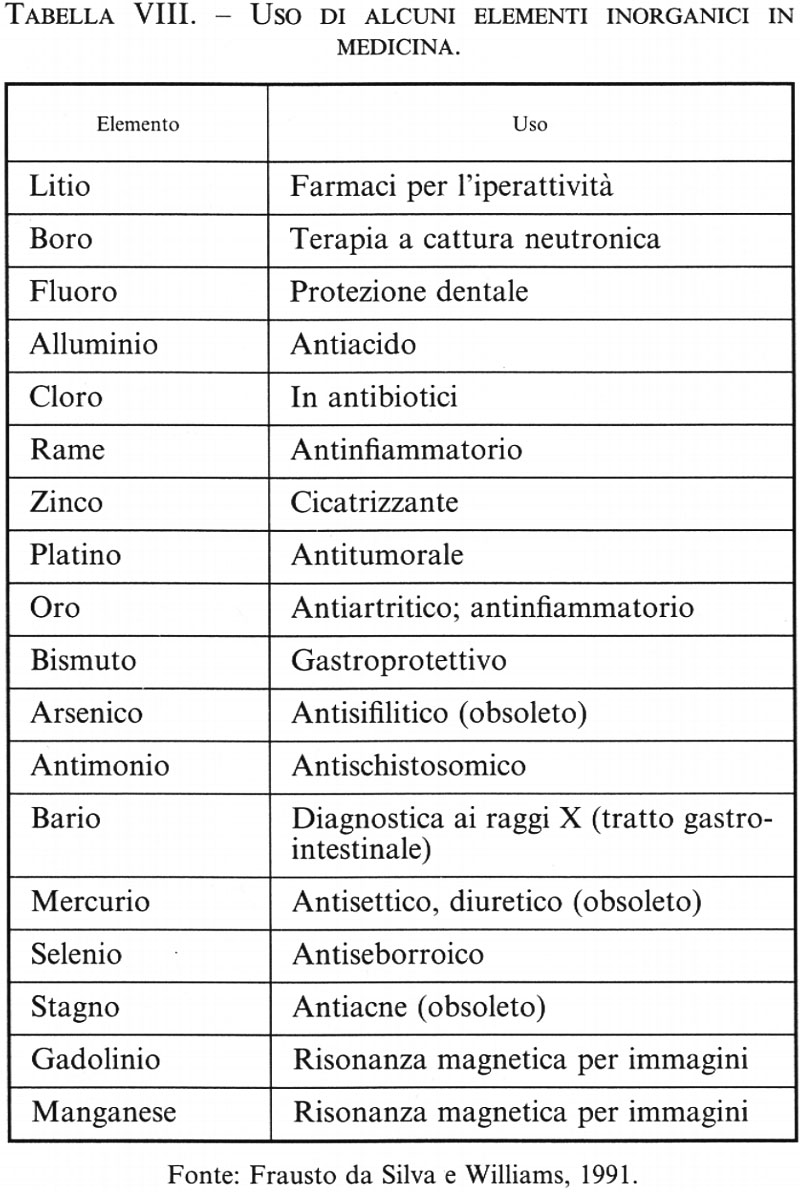
La chimica bioinorganica sta contribuendo a gettare le basi di questa conoscenza a livello molecolare. Il risultato è lo sviluppo, accanto ad alcuni farmaci classici o addirittura già usati su basi empiriche fin dai tempi antichi, di chemioterapici e diagnostici di efficacia impensabile fino a pochi anni fa. La tab. VIII riporta alcuni esempi, antichi e moderni, di uso terapeutico di elementi inorganici, e la tab. IX elenca alcuni radionuclidi usati, in dosi bassissime data l'alta sensibilità dei rivelatori di radioattività, come diagnostici specifici per vari organi. Alcuni composti di coordinazione di ioni metallici (soprattutto Gd3+ e Mn2+) sono usati come mezzi di contrasto non radioattivi nelle immagini di risonanza magnetica (v. tomografia: Tomografia a risonanza magnetica nucleare, vol. VIII; v. cap. 8); la loro selettività per i vari tessuti è conferita non dal metallo ma dai diversi tipi di ligandi a cui esso è coordinato, per esempio ligandi idrosolubili o liposolubili, ecc.

La fig. 13 mostra le strutture di due farmaci e di due diagnostici per immagini, di formulazione recente e di larghissimo uso. Il primo di essi, il cosiddetto cis-platino, merita un commento particolare. La formula è in realtà rappresentativa di una intera classe di composti, tutti caratterizzati dalla presenza di uno ione Pt2+ legato a due atomi di cloro che formano con esso un angolo di circa 90° (legame cis), e ad altri due ligandi più deboli (nel caso più comune, NH3), che dopo la somministrazione si dissociano, sostituiti da due molecole d'acqua. La specie PtCl2(OH2)2 reagisce con il DNA, che sostituisce le due molecole d'acqua con gli atomi di azoto di due residui guaninici. Sembra che la modifica strutturale nel DNA indotta da questo legame (v. Pil e Lippard, 1992) stimoli l'interazione del DNA con specifiche proteine cellulari che ne bloccano la riparazione rapida. Questo effetto inibitorio sulla capacità della cellula di riparare il DNA è, in linea di principio, dannoso per tutte le cellule, ma soprattutto per le cellule tumorali, che si riproducono molto più rapidamente di quelle sane. In alcuni tipi di tumore la terapia con cis-platino, se iniziata precocemente, si dimostra efficace in più del 90% dei casi.
8. Metodi di indagine
Il progresso delle scienze chimiche in questi ultimi decenni non è tanto dovuto al progredire delle teorie del legame chimico, che sono maturate essenzialmente nella prima metà del secolo, quanto all'incredibile sviluppo che hanno avuto i metodi di indagine da un lato (v. chimica: Analisi chimica strumentale, vol. VIII; v. molecole: Metodi di indagine strutturale, vol. IV), e l'informatica dall'altro. In entrambi i casi, tale sviluppo è principalmente legato a quello dell'elettronica, che ha permesso il rapido evolversi sia della strumentazione relativa a metodi fisici di misura e di analisi sempre più sofisticati, sia della capacità di calcolo, e quindi di rappresentazione con modelli teorici, di molecole anche complesse come proteine, enzimi o acidi nucleici. Due ulteriori fattori che hanno dato impulso alla chimica dei sistemi biologici sono stati da un lato il progresso nei prodotti e nei materiali usati per le purificazioni biochimiche (si pensi soprattutto alle resine per cromatografia), e dall'altro lo sviluppo delle tecniche di ingegneria genetica e, in generale, delle biotecnologie.
La chimica bioinorganica ha ovviamente beneficiato, insieme alla biochimica, di tutti questi progressi, anche se ciò che è risultato di importanza fondamentale è stato lo sviluppo dei metodi fisici di indagine. La presenza caratterizzante di elementi metallici nei sistemi bioinorganici richiede infatti spesso l'uso di metodi chimico-fisici e spettroscopici sofisticati; d'altro canto, questi ultimi forniscono informazioni molto dettagliate e importanti per la comprensione della struttura e della funzione delle molecole in questione. Si riassumono qui di seguito i metodi più importanti a disposizione del chimico bioinorganico, rimandando, per una descrizione più esauriente dei principî su cui si basano, a trattazioni più specializzate (v. Drago, 19922).
Diffrazione dei raggi X
I raggi X sono una forma di radiazione elettromagnetica caratterizzata da una lunghezza d'onda dell'ordine di 1 Å (10-10 m), cioè dell'ordine delle distanze di legame fra gli atomi, e sono quindi diffratti quando attraversano un materiale. Se quest'ultimo è sotto forma cristallina, cioè in una forma in cui le molecole sono ordinate in un reticolo regolare, la tecnica permette, tramite algoritmi ormai quasi completamente automatizzati, di trasformare l'informazione contenuta nella radiazione diffratta in informazione strutturale sul campione che causa la diffrazione. La diffrazione dei raggi X è usata in chimica da molti decenni, ma ha ricevuto un impulso straordinario negli anni novanta dalla disponibilità di radiazione X da sincrotrone, molto più intensa di quella delle sorgenti radioattive convenzionali, e da rivelatori della radiazione diffratta molto più sensibili (i cosiddetti area detectors). Allo stato attuale è possibile risolvere la struttura tridimensionale di una proteina in poche settimane, raggiungendo una risoluzione inferiore a 2 Å e a volte vicina a 1 Å (v. Dickerson, 1992). Molecole biologiche contenenti metalli non presentano problemi particolari. La presenza del metallo può essere anzi utile nell'analisi preliminare dei dati.
Spettroscopia XAS
La spettroscopia di assorbimento dei raggi X (XAS, X-ray Absorption Spectroscopy) utilizza la stessa sorgente di radiazione usata per le tecniche di diffrazione, ma in questo caso viene misurato l'assorbimento della radiazione in funzione della sua energia. Per questi esperimenti occorre usare una sorgente di radiazione X da sincrotrone. La spettroscopia XAS (v. Teo, 1986), attraverso l'eccitazione degli elettroni interni di un elemento metallico, fornisce informazioni sulle energie degli orbitali più esterni liberi oppure, a più alte energie, sull'energia di ionizzazione di tale elemento. In questo modo si ottengono informazioni, ad esempio, sullo stato di ossidazione di un elemento metallico in una matrice biologica, sulla sua geometria di coordinazione e anche, dall'analisi dell'interferenza con gli atomi vicini, sul tipo e sul numero degli atomi dei ligandi. Non è necessario che il campione sia allo stato cristallino e neanche allo stato solido. L'informazione è limitata alle immediate vicinanze del metallo, ma in questo ambito ristretto può essere anche molto dettagliata.
EPR
La risonanza paramagnetica elettronica (EPR, Electron Paramagnetic Resonance; v. Hoff, 1989; v. Sigel, 1987; v. Lowe, 1992) sfrutta la presenza di elettroni spaiati che spesso caratterizza i composti dei metalli di transizione. Gli elettroni sono dotati di un momento magnetico, detto di spin, che è rivelabile quando non si trovano appaiati nello stesso orbitale, ma spaiati in uno o più orbitali diversi (sostanze paramagnetiche). Se immerso in un campo magnetico, un elettrone spaiato può assumere due diversi stati di spin, a diversa energia (n elettroni spaiati possono assumere n + 1 stati di spin). Si può far passare lo ione metallico che contiene elettroni spaiati da uno stato di spin all'altro irradiandolo con una radiazione elettromagnetica a bassa energia (nella regione delle microonde, per i campi magnetici tipici di uno spettrometro EPR), e ottenere così il valore della differenza di energia fra gli stati di spin. Tale valore dipende dall'intorno chimico dello ione metallico in questione, e permette di ricavare informazioni su di esso sia che la sostanza studiata si trovi allo stato solido sia che si trovi in soluzione. Altre spettroscopie correlate all'EPR sono la doppia risonanza elettrone-nucleo (ENDOR, Electron-Nuclear DOuble Resonance; v. Hoffman, 1991; v. Sigel, 1987; v. Lowe, 1992) - che coniuga un esperimento di tipo EPR con uno di tipo NMR (vedi sotto) - e l'eco di spin elettronico (ESEEM, Electron Spin Echo Envelope Modulation; v. Sigel, 1987). Queste tecniche sono relativamente più recenti dell'EPR e stanno avendo una buona diffusione in chimica bioinorganica perché spesso forniscono informazioni ancora più dettagliate.
NMR
La risonanza magnetica nucleare (NMR, Nuclear Magnetic Resonance; v. Wüthrich, 1986; v. Bertini e Luchinat, 1986 e 1996; v. Banci e altri, 1991; v. Sigel, 1986) è forse la spettroscopia che singolarmente fornisce maggiori informazioni chimiche. Analogamente all'EPR, si basa sull'eccitazione di transizioni di spin, nucleari anziché elettroniche, prodotta dall'irradiazione con onde radio del campione immerso in un campo magnetico. La risposta del campione dipende, a seconda degli esperimenti, dall'intorno chimico di ogni singola specie nucleare (ad esempio dei nuclei di idrogeno) contenuta nel campione e dalle interazioni fra i nuclei attraverso lo spazio o attraverso legami chimici. La spettroscopia NMR è progredita nell'ultimo decennio a un punto tale che è ora possibile, analizzando al calcolatore i diversi tipi di interazione internucleare con procedure ormai discretamente automatizzate, ottenere la struttura in soluzione di piccole proteine. Sebbene tuttora meno potente delle tecniche ai raggi X per le determinazioni strutturali, la spettroscopia NMR risulta indispensabile quando non si riescano a ottenere cristalli della molecola in questione, oppure quando si sospetti che la struttura in soluzione differisca da quella allo stato cristallino: se l'attività biologica della molecola si esplica in soluzione, la struttura allo stato cristallino potrebbe fornire informazioni fuorvianti. Nel caso dell'NMR, la presenza di metalli con elettroni spaiati è stata considerata, fino alla prima metà degli anni ottanta, un fattore sfavorevole, perché l'interazione con il centro paramagnetico fa decadere più rapidamente il segnale che si genera per eccitazione degli spins nucleari, a volte impedendone la rivelazione. Si è dimostrato tuttavia che, a parte le situazioni di paramagnetismo molto pronunciato, l'interazione con gli elettroni spaiati può essere un'ulteriore fonte di informazione sulla struttura della molecola (v. Bertini e Luchinat, 1986 e 1996). Un'applicazione particolare della tecnica NMR in presenza di metalli paramagnetici è data dalla rilassometria, che è lo studio dell'effetto degli elettroni spaiati sul decadimento (rilassamento) dei segnali nucleari in funzione del campo magnetico applicato. In particolare, lo studio del rilassamento degli atomi di idrogeno dell'acqua in soluzioni di metalloproteine paramagnetiche può fornire informazioni preziose sulla presenza di molecole d'acqua coordinate al metallo (v. Banci e altri, 1991). Le tecniche di risonanza magnetica per immagini usate in medicina (v. cap. 7) sfruttano il fatto che il rilassamento dei protoni dell'acqua in un organismo è diverso a seconda del tipo di tessuto; da tale diversità, infatti, dipende il contrasto, ovvero la qualità dell'immagine. Come si è visto, l'aggiunta di composti di coordinazione contenenti ioni metallici paramagnetici, che si concentrino in particolari zone dell'organo da esaminare, permette di migliorare la qualità dell'immagine fino a rivelare particolari altrimenti invisibili.
Spettroscopia Mössbauer
Questa tecnica si basa sull'assorbimento dei raggi γ, emessi dai nuclei di un elemento sorgente in uno stato eccitato, da parte di un nucleo dello stesso elemento presente nel campione (v. Dickson e Berry, 1986). La differenza di energia fra sorgente e bersaglio fornisce informazioni sull'intorno chimico del nucleo metallico nel campione, in particolare sul suo numero di ossidazione. In chimica bioinorganica la tecnica è usata soprattutto nel caso di composti contenenti ferro, che a questo scopo vengono spesso arricchiti con l'isotopo 57Fe per migliorare la sensibilità.
Spettroscopie elettroniche e vibrazionali
Si tratta di tecniche spettroscopiche classiche, come la spettroscopia nell'infrarosso, vicino infrarosso, visibile e ultravioletto (v. Lever, 19842), che si basano sull'eccitazione tramite radiazione elettromagnetica di transizioni fra diversi stati energetici di una molecola. Sono tecniche di fondamentale importanza in tutte le branche della chimica; di particolare interesse in chimica bioinorganica sono le transizioni fra i livelli elettronici d dei metalli di transizione - spesso fra le più atte a diagnosticare il numero e la geometria di coordinazione - che si osservano nel visibile o nel vicino infrarosso. Una variante utile è la spettroscopia di dicroismo circolare, che permette in molti casi di ottenere una migliore risoluzione delle bande di assorbimento, o di dicroismo circolare magnetico (v. Johnson e altri, 1982) che, sia pur di difficile interpretazione, fornisce spesso una ‛impronta digitale' utile in certi casi per individuare l'intorno di coordinazione. Nel vicino ultravioletto spesso si trovano le cosiddette bande a trasferimento di carica, indicative dell'interazione di legame fra il metallo e i suoi leganti. Oltre alla spettroscopia XAS, già descritta, vi sono poi le spettroscopie di fotoelettroni ultravioletti (UPS, Ultraviolet Photoelectron Spectroscopy), e di fotoelettroni ai raggi X (XPS, X-ray Photoelectron Spectroscopy), meno diffuse ma relativamente promettenti. Fra le spettroscopie vibrazionali nell'infrarosso è particolarmente utile la spettroscopia Raman di risonanza, che permette di evidenziare le bande dovute alle vibrazioni dei leganti in metalloproteine tramite l'eccitazione di transizioni elettroniche del metallo (v. Drago, 19922).
Misure magnetiche
Dalle informazioni quantitative sulle proprietà magnetiche del centro metallico è possibile ottenere dati relativi a molte altre proprietà, inclusa la reattività chimica del metallo stesso, alle quali sono collegate. Il paramagnetismo di una molecola può essere misurato con tecniche già diffuse in chimica da molti decenni (magnetochimica; v. O'Connor, 1982). Quando però il centro paramagnetico è parte di una macromolecola come una proteina, la misura è resa difficile dalla presenza di un rilevante contributo di segno opposto (diamagnetismo) che si origina dagli elettroni appaiati di tutti gli atomi della macromolecola. Queste misure richiedono quindi strumenti molto sensibili; si sono diffusi recentemente strumenti che hanno queste caratteristiche, detti SQUID (Superconducting Quantum Interference Device), che hanno reso possibili misure di suscettività magnetica relativamente accurate anche su metalloproteine di medie dimensioni e di non elevato paramagnetismo.
Misure elettrochimiche
Come abbiamo visto, in molti casi la funzione dello ione metallico è legata alla sua capacità di cambiare stato di ossidazione e quindi di trasportare elettroni o catalizzare reazioni di ossidoriduzione. È quindi comprensibile che assuma particolare importanza la misura del potenziale di riduzione dello ione metallico quando è incorporato nella matrice proteica. Come nel caso delle misure magnetiche, le tecniche elettrochimiche non sono fra le più nuove nell'arsenale del chimico; tuttavia, le misure elettrochimiche su metalloproteine sono spesso complicate dalla scarsa risposta elettrica del sistema, a sua volta legata a difficoltà di interazione della proteina con la superficie dell'elettrodo (v. Hill, 1987). Questo campo di ricerca fornisce un tipico esempio di quanto le caratteristiche dei nuovi materiali (struttura cristallina degli elettrodi, metodi di pulitura della loro superficie) e la varietà di composti chimici disponibili in commercio (molecole che agiscono da promotori disponendosi a ponte fra la superficie dell'elettrodo e la metalloproteina) possano ampliare le potenzialità delle tecniche (v. Azab e altri, 1992).
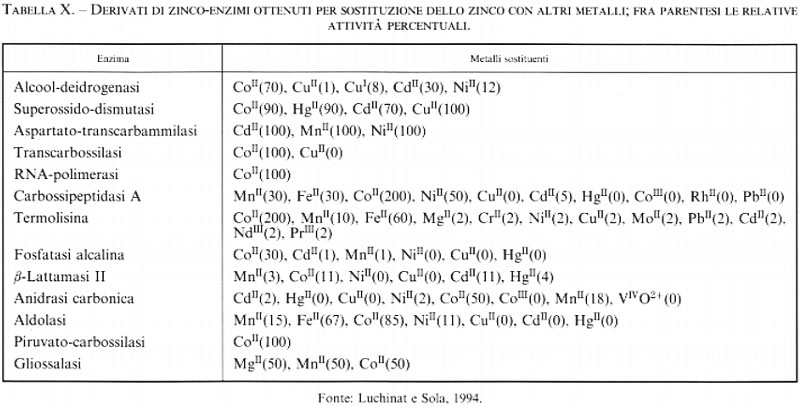
I cenni forzatamente limitati che sono stati dati in questo capitolo sui più diffusi metodi di indagine non rendono piena giustizia all'importanza che essi hanno avuto, e certamente avranno ancora in futuro, nello sviluppo della chimica bioinorganica. Si può forse aggiungere che certe tecniche spettroscopiche specifiche per particolari ioni metallici si rivelano così importanti per la caratterizzazione dei sistemi biologici che li contengono, da stimolare lo sviluppo di una vera e propria metodologia, detta di ‛sostituzione metallica', che ha lo scopo di inserire, al posto dello ione metallico naturale, uno ione metallico diverso, che sia studiabile con una certa tecnica spettroscopica (v. Bertini e Luchinat, 1983). Così ad esempio è divenuto usuale, nella ricerca bioinorganica, sostituire lo ione Zn2+ (v. Bertini e altri, 1986), cieco a quasi tutte le spettroscopie, con altri ioni bipositivi di metalli di transizione, come il cobalto (per la spettroscopia elettronica e NMR), il rame e il manganese (per l'EPR e la rilassometria), il cadmio (per l'NMR del suo isotopo 113Cd), ecc. A volte la sostituzione mantiene l'attività biologica della molecola (v. tab. X; v. Luchinat e Sola, 1994); ma anche quando questo non si verifica, le informazioni ottenute costituiscono sempre un aiuto prezioso per la comprensione del sistema.
9. Prospettive della disciplina
Occorre premettere che è difficile parlare di prospettive a lunga o anche a media scadenza per una disciplina giovane e in tumultuoso sviluppo come la chimica bioinorganica. Nell'esperienza ventennale di ricerca in questo campo da parte dell'autore non è stato infrequente veder raggiunti nell'arco di mesi traguardi fino a poco prima ritenuti raggiungibili solo in più di un decennio. Fra le principali componenti che hanno permesso lo sviluppo delle scienze biochimiche, in generale, e della bioinorganica, in particolare, abbiamo cercato (v. cap. 8) di mettere in luce l'importanza dei progressi verificatisi nel campo della strumentazione, dell'informatica, dei nuovi materiali e prodotti chimici, delle biotecnologie. Non a caso, questi campi sono oggetto, in Italia come in tutti i paesi industrializzati, di particolare attenzione e sono considerati temi strategici da parte degli organi di finanziamento della ricerca.
Per ciò che riguarda la strumentazione, si possono prevedere ulteriori progressi a breve scadenza nelle tecniche di raccolta dei dati di diffrazione di raggi X. È prevedibile che il numero annuo di risoluzioni di strutture di metalloproteine, metalloenzimi e, soprattutto, di addotti metalloproteine-DNA (v. cap. 6) aumenti in modo ancor più significativo di quanto avvenuto nel recentissimo passato. La tecnica dovrebbe anche potersi avvantaggiare della crescente automazione delle metodiche di crescita di cristalli di macromolecole biologiche. Progressi altrettanto rapidi si attendono nelle tecniche di risonanza magnetica, grazie sia alla diffusione di strumenti EPR pulsati, sia al continuo innalzamento dei campi magnetici (fino a oltre 20 tesla) per l'NMR. Piccoli produttori di apparecchi scientifici stanno già offrendo sul mercato strumenti per la rilassometria, tecnica finora ristretta a ricercatori che disponevano di strumenti ‛fatti in casa', data la crescente importanza che essa riveste come tecnica di supporto per la diagnostica medica per immagini.
Nel campo dell'informatica, le previsioni di un rapido sviluppo sono tanto facili quanto, spesso, errate per difetto. I progressi dell'informatica hanno contribuito notevolmente - ed è logico attendersi che continueranno a farlo - al crescente successo delle determinazioni strutturali sia tramite raggi X che per mezzo della risonanza magnetica nucleare. Se poi nel prossimo decennio la potenza di calcolo continuasse ad aumentare anche solo al ritmo attuale, potrebbe non essere azzardato prevedere la risoluzione di un problema centrale della biochimica: quello della previsione a tavolino dell'avvolgimento (folding) di una qualunque proteina data la sua sequenza primaria. Gli algoritmi esistenti sono già basati su buone previsioni teoriche dei potenziali per le interazioni atomo-atomo per tutti i residui amminoacidici. Mancano invece ancora, in gran parte, dei buoni potenziali per gli ioni metallici; la loro messa a punto costituirà probabilmente un campo di ricerca molto attivo nel prossimo futuro.
In relazione ai nuovi materiali e prodotti chimici, va rilevato che l'isolamento e la purificazione di sostanze biologiche da estratti cellulari hanno fatto enormi progressi grazie allo sviluppo di tecniche di laboratorio quali le tecniche cromatografiche, elettroforetiche, ecc. Queste ultime continueranno a trarre vantaggio dalle tecnologie chimiche sviluppate per la produzione di resine, comprese quelle per affinity chromatography che permettono la separazione immediata della proteina voluta. Usi molto promettenti di nuovi materiali e prodotti chimici si dovrebbero avere nel campo delle zeoliti e nel campo dei kits analitici di riconoscimento o di misura dell'attività biologica.
Non è questa la sede per illustrare l'importanza delle biotecnologie nel progresso della scienza di questi ultimi anni (v. biotecnologie, vol. VIII e biotecnologie, vol. X). Dal punto di vista della caratterizzazione strutturale e funzionale di metalloproteine e metalloenzimi, soprattutto quando si usano metodi fisici d'avanguardia, spesso il fattore limitante è dato dalla quantità di sostanza disponibile. La semplice possibilità di superesprimere una certa proteina, isolabile in piccolissime quantità da un certo organismo, in un organismo diverso, sta già cambiando radicalmente il modo di concepire la ricerca. Le tecniche del DNA ricombinante stanno progredendo e standardizzandosi così rapidamente che già ora è impensabile affrontare lo studio di una nuova metalloproteina o di un nuovo metalloenzima senza programmare in parallelo l'isolamento o la sintesi chimica del gene relativo e la sua espressione in un microrganismo compiacente. Disporre di una proteina espressa significa poi non solo poter superare il problema della quantità di sostanza, ma anche poter progettare e caratterizzare un gran numero di mutanti ottenuti tramite mutagenesi sito-specifica e, attraverso di essi, formulare ipotesi sui dettagli del meccanismo d'azione. Infine, è doveroso accennare ai sempre più numerosi programmi di ricerca che si pongono l'obiettivo ambizioso di progettare e realizzare, attraverso tecniche genetiche, enzimi artificiali (il cosiddetto de novo design). La progettazione di nuovi metalloenzimi è particolarmente promettente, perché già in natura esistono molti esempi di uso dello stesso gruppo prostetico (per esempio l'eme, come abbiamo visto) per funzioni che vanno dal trasporto elettronico al trasporto dell'ossigeno, alla sua stessa attivazione. Le differenze funzionali del gruppo eme sono principalmente da ascriversi alla natura dei leganti assiali (v. Rietjens e altri, 1996): tramite mutagenesi sito-specifica si sta già realizzando la trasformazione, ad esempio, della mioglobina o di un citocromo in una perossidasi. Si può prevedere che il de novo design diverrà uno dei maggiori temi della chimica bioinorganica dei prossimi anni.
L'importanza che la chimica bioinorganica avrà nel prossimo futuro non sarà però tanto legata al progresso delle metodiche sperimentali, quanto e soprattutto alla capacità della disciplina di porsi in posizione centrale nei riguardi dei grandi problemi del nostro tempo: la durata e la qualità della vita, la salute, l'ambiente. Per i primi, le prospettive della disciplina dovrebbero essere apparse chiare, anche se attraverso pochi cenni, già in questa rassegna. Merita invece soffermarsi sulla collocazione che la bioinorganica potrebbe avere in futuro nelle problematiche ambientali, dato il ruolo fondamentale che rivestono i microrganismi del suolo nella cosiddetta bonifica biologica (bioremediation) di siti inquinati dalla presenza di composti organici in gran parte derivati dalla petrolchimica (v. inquinamento ambientale, vol. III e inquinamento ambientale vol. VIII). La presenza di questi nuovi inquinanti sulla superficie del pianeta ha stimolato l'evoluzione di nuovi cammini (pathways) metabolici da parte dei batteri del suolo. Attraverso lo sviluppo di plasmidi, questi batteri hanno codificato enzimi che permettono, ad esempio, la diossigenazione di composti aromatici o poliaromatici condensati, o la mono-ossigenazione degli alcani, seguite da ulteriori ossigenazioni che portano, dopo vari passaggi, a intermedi del ciclo di Krebs, e quindi metabolizzabili facilmente da tutti gli organismi. Un pathway metabolico comune a molte sostanze aromatiche è riportato in fig. 14. La maggior parte di queste nuove diossigenasi e monoossigenasi sono metalloenzimi, in genere contenenti ferro, il cui meccanismo di azione deve ancora essere chiarito. In questi studi la chimica bioinorganica avrà un ruolo preminente. È anche plausibile che gli enzimi più interessanti dal punto di vista ambientale non siano ancora ‛perfetti' dal punto di vista evolutivo come la maggior parte degli enzimi del corredo genetico stabile. Si può quindi ipotizzare che la bioinorganica possa ‛aiutare' l'evoluzione dei microrganismi benefici, contribuendo a progettare plasmidi modificati che migliorino l'efficienza catalitica o lo spettro d'azione dei loro enzimi.
BIBLIOGRAFIA
Antholine, W. E., Kastrau, D. H. W., Steffens, G. C. M., Buse, G., Zumft, W. G., Kroneck, P. M. H., A comparative EPR investigation of the multicopper proteins nitrous-oxide reductase and cytochrome c oxidase, in ‟European journal of biochemistry", 1992, CCIX, pp. 875-881.
Antonini, E., Brunori, M., Hemoglobin and myoglobin in their reactions with ligands, New York 1971.
Auld, D. S., Riordan, J. F., Vallee, B. L., Probing the mechanism of carboxypeptidase A by inorganic, organic and mutagenic modifications, in Metal ions in biological systems, vol. XXV, Interrelations among metal ions, enzymes and gene expression (a cura di H. Sigel), New York 1989, pp. 359-415.
Azab, H. A., Banci, L., Borsari, M., Luchinat, C., Sola, M., Viezzoli, M. S., Redox chemistry of superoxide dismutase. Cyclic voltammetry of wild-type enzymes and mutants on functionally relevant residues, in ‟Inorganic chemistry", 1992, XXXI, pp. 4649-4655.
Baker, E. N., Anderson, B. F., Baker, H. M. e altri, Structure, function and flexibility of human lactoferrin, in ‟International journal of biological macromolecules", 1991, XIII, pp. 122-129.
Banci, L., Bertini, I., Eltis, L. D., Felli, I. C., Kastrau, D. H. W., Luchinat, C., Piccioli, M., Pierattelli, R., Smith, M., The three dimensional structure in solution of the paramagnetic high-potential iron-sulfur protein I from Ectothiorhodospira halophila through nuclear magnetic resonance, in ‟European journal of biochemistry", 1994, CCXXV, pp. 715-725.
Banci, L., Bertini, I., Luchinat, C., Nuclear and electron relaxation, Weinheim 1991.
Beinert, H., Recent developments in the field of iron-sulfur proteins, in ‟FASEB journal", 1990, IV, pp. 2483-2491.
Berg, J. M., Zinc fingers and other metal binding domains, in ‟Journal of biological chemistry", 1990, CCLXV, pp. 6513-6516.
Berg, J. M., Holm, R. H., Structures and reactions of iron-sulfur protein clusters and their synthetic analogues, in Iron-sulfur proteins (a cura di T. G. Spiro), New York 1982, pp. 1-66.
Bertini, I., Bren, K. L., Clemente, A., Fee, J. A., Gray, H. B., Luchinat, C. Malmström, B. G., Richards, J. H., Sanders, D., Slutter, C. E., The CuA center of a soluble domain from Thermus cytochrome ba3: an NMR investigation of the paramagnetic protein, in ‟Journal of the American Chemical Society", 1996, CXVIII, pp. 11658-11659.
Bertini, I., Drago, R. S., Luchinat, C. (a cura di), The coordination chemistry of metalloenzymes, Dordrecht 1983.
Bertini, I., Gray, H. B., Lippard, S. J., Valentine, J. S. (a cura di), Bioinorganic chemistry, Mill Valley, Cal., 1994.
Bertini, I., Luchinat, C., An insight on the active site of zinc enzymes through metal substitution, in Metal ions in biological systems, vol. XV, Zinc and its role in biology and nutrition (a cura di H. Sigel), New York 1983, pp. 101-156.
Bertini, I., Luchinat, C., Metal ions in biochemistry, in Structure and motion: membranes, nucleic acids and proteins (a cura di E. Clementi e altri), Boston 1985, pp. 293-328.
Bertini, I., Luchinat, C., NMR of paramagnetic molecules in biological systems, Menlo Park, Cal., 1986.
Bertini, I., Luchinat, C., The reaction pathways of zinc enzymes and related biological catalysts, in Bioinorganic chemistry (a cura di I. Bertini, H. B. Gray, S. J. Lippard e J. S. Valentine), Mill Valley, Cal., 1994, pp. 37-106.
Bertini, I., Luchinat, C., NMR of paramagnetic substances, Lousanne 1996.
Bertini, I., Luchinat, C., Maret, W., Zeppezauer, M. (a cura di), Zinc enzymes, Boston 1986.
Bertini, I., Luchinat, C., Messori, L., The role of aminoacidic residues inside the active sites of metalloproteins, in ‟Journal of pure and applied chemistry", 1988, LX, pp. 1261-1266.
Cammack, R., Sykes, A. G. (a cura di), in Iron-sulfur proteins, San Diego, Cal., 1992.
Christianson, D. W., Structural biology of zinc, in Advances in protein chemistry, vol. XLII (a cura di C. B. Anfinsen, J. T. Edsall, D. Eisenberg e F. M. Richards), San Diego, Cal., 1991, pp. 281-355.
Christianson, D. W., Lipscomb, W. N., Carboxypeptidase A, in ‟Accounts of chemical research", 1989, XXII, pp. 62-69.
Cowan, J. A., Inorganic biochemistry: an introduction, Weinheim 1993.
Crick, R. E. (a cura di), Origin, evolution and modern aspects of biomineralization in plants and animals, New York 1989.
Dennard, A. B., Williams, R. J. P., Transition metal ions as reagents in metalloenzymes, in Transition metal chemistry (a cura di R. L. Carlin), London 1966, vol. II, pp. 115 ss.
Dickerson, R. E., A little ancient history, in ‟Protein science", 1992, I, pp. 182-186.
Dickson, D. P. E., Berry, F. J. (a cura di), Mössbauer spectroscopy, New York 1986.
Drago, R. S., Physical methods for chemists, Fort Worth, Tex., 19922.
Draper, D. E., The RNA-folding problem, in ‟Accounts of chemical research", 1992, XXV, pp. 201-207.
Frausto da Silva, J. J. R., Williams, R. J., The biological chemistry of the elements: the inorganic chemistry of life, Oxford 1991.
Hamer, D. H., Winge, D. R. (a cura di), Metal-ion homeostasis: molecular biology and chemistry, New York 1989.
Harayama, S., Timmis, K. N., Catabolism of aromatic hydrocarbons by Pseudomonas, in Genetics of bacterial diversity (a cura di D. A. Hopwood e K. F. Chater), London 1989, pp. 151-174.
Hay, R. W., Bio-inorganic chemistry, Chichester 1987.
Hill, H. A. O., Bio-electrochemistry, in ‟Pure and applied chemistry", 1987, LIX, pp. 743-748.
Hoff, A. J., Advanced EPR-application in biology and biochemistry, Amsterdam 1989.
Hoffman, B. M., Electron Nuclear DOuble Resonance (ENDOR) of metalloenzymes, in ‟Accounts of chemical research", 1991, XXIV, pp. 164-170.
Hughes, M. N., The inorganic chemistry of biological processes, London 19812.
Johnson, M. K., Robinson, A. E., Thomson, A. J., Low-temperature magnetic circular dichroism studies of iron-sulfur proteins, in Iron-sulfur proteins (a cura di T. G. Spiro), New York 1982, pp. 367-406.
Kim, J., Rees, D. C., Structural models for the metal centers in the nitrogenase molybdenum-iron protein, in ‟Science", 1992, CCLVII, pp. 1677-1682.
Kurtz, D. M. Jr., Oxo- and hydroxo-bridged diiron complexes: a chemical perspective on a biological unit, in ‟Chemical reviews", 1990, XC, pp. 585-606.
Lever, A. B. P., Inorganic electronic spectroscopy, New York 19842.
Lippard, S. J., Berg, J. M., Principles of bioinorganic chemistry, Mill Valley, Cal., 1994.
Lowe, D. J., ENDOR and EPR of metalloproteins, in ‟Progress in biophysics & molecular biology", 1992, LVII, pp. 1-22.
Luchinat, C., Sola, M., Zinc enzymes, in Encyclopedia of inorganic chemistry, vol. VIII, New York 1994, pp. 4406-4434.
Malmström, B. G., Cytochrome c oxidase as a redox-linked proton pump, in ‟Chemical reviews", 1990, XC, pp. 1247-1260.
Mann, S., Webb, J., Williams, R. J. P., Biomineralisation, Weinheim 1989.
Ochiai, E. I., Bioinorganic chemistry: an introduction, Boston 1977.
O'Connor, C. J., Magnetochemistry: advances in theory and experimentation, in ‟Progress in inorganic chemistry", 1982, XXIX, pp. 203-283.
Okamura, M. Y., Feher, G., Proton transfer in reaction centers from photosynthetic bacteria, in ‟Annual review of biochemistry", 1992, LXI, pp. 861-896.
Pil, P. M., Lippard, S. J., Specific binding of chromosomal protein HMG1 to DNA damaged by the anticancer drug cisplatin, in ‟Science", 1992, CCLVI, pp. 234-237.
Renger, G., On the mechanism of photosynthetic water oxidation to dioxygen, in ‟Chemica scripta", 1988, XXVIII A, pp. 105-109.
Rietjens, I. M. C. M., Poulos, T. L., Goodin, D. B., Banci, L., Gross, Z., Weiss, R. e altri, The role of the axial ligand in heme-based catalysis, in ‟Journal of biological inorganic chemistry", 1996, I, pp. 355-383.
Sigel, H. (a cura di), Metal ions in biological systems, vol. XXI, Applications of NMR to paramagnetic species, New York 1986.
Sigel, H. (a cura di), Metal ions in biological systems, vol. XXII, ENDOR, EPR and electron spin echo for probing coordination sphere, New York 1987.
Stiefel, E. I., George, G. N., Ferrodoxins, hydrogenases and nitrogenases: metalsulfide proteins, in Bioinorganic chemistry (a cura di I. Bertini, H. B. Gray, S. J. Lippard e J. S. Valentine), Mill Valley, Cal., 1994, pp. 365-453.
Stryer, L., Biochemistry, New York 19954 (tr. it.: Biochimica, Bologna 19964).
Sykes, A. G. (a cura di), Advances in inorganic and bioinorganic mechanisms, New York 1985.
Teo, B. K., EXAFS: basic principles and data analysis, New York 1986.
Theil, E. C., The ferritin family of iron storage proteins, in ‟Advances in enzymology and related areas of molecular biology", 1990, LXIII, pp. 421-449.
Theil, E. C., Raymond, K. N., Transition metal storage, transport and biomineralization, in Bioinorganic chemistry (a cura di I. Bertini, H. B. Gray, S. J. Lippard e J. S. Valentine), Mill Valley, Cal., 1994, pp. 1-35.
Tullius, T. D. (a cura di), Metal-DNA chemistry, Washington 1989.
Underwood, E. J., Trace elements in human and animal nutrition, New York 19774.
Wächtershäuser, G., The origin of evolution in an iron-sulfur world, in ‟Journal of inorganic biochemistry", 1994, LVI, p. 59.
Westbroek, P., De Jong, E. W. (a cura di), Biomineralization and biological metal accumulation, Dordrecht 1983.
Wilkins, P. G., Wilkins, R. G., The coordination chemistry of the binuclear iron site in hemerythrin, in ‟Coordination chemistry review", 1987, LXXIX, pp. 195-214.
Wüthrich, K., NMR of proteins and nucleic acids, New York 1986.