
Immunologia clinica e immunopatologia
Immunologia clinica e immunopatologia
sommario: 1. Introduzione: a) generalità; b) i concetti fondamentali dell'immunologia: note storiche. 2. Immunodeficienze primitive: a) classificazione e diagnostica; b) agammaglobulinemia congenita legata al sesso; c) difetto selettivo di IgA; d) immunodeficienza comune variabile; e) immunodeficienza grave combinata; f) sindrome di Wiskott-Aldrich; g) sindrome della terza e quarta tasca branchiale (DiGeorge's syndrome); h) atassia-teleangectasia. 3. La sindrome da immunodeficienza acquisita: a) introduzione; b) l'HIV: un particolare retrovirus; c) il danno immunologico e la patologia cellulare; d) l'anatomia patologica del linfonodo; e) diagnosi di laboratorio; f) le classificazioni e l'inquadramento nosologico; g) le conseguenze cliniche dell'infezione da HIV; h) i problemi epidemiologici; i) la prevenzione; l) la terapia. 4. Ruolo della risposta immunitaria nel controllo della crescita neoplastica: a) la cellula tumorale; b) modalità con le quali il sistema immunitario può contenere l'espansione neoplastica; c) meccanismi attraverso i quali la cellula tumorale elude la sorveglianza immunologica. 5. Il concetto di monitoraggio immunologico. □ 6. Prospettive della terapia immunologica: a) immunosoppressione; b) immunostimolazione. □ Bibliografia.
1. Introduzione
a) Generalità
Disciplina minore fino a 30 anni or sono, l'immunologia ha visto il suo campo di interessi estendersi continuamente, integrandosi con numerosi altri settori di ricerca e fornendo alla medicina nuovi strumenti di diagnosi e terapia. Negli ultimi 15 anni si sono realizzati progressi a livello di ricerca (con l'introduzione di nuove tecnologie in immunologia di base), a livello diagnostico (per la disponibilità di sofisticate e precise metodiche di indagine), a livello farmacologico e terapeutico (sintesi di nuove molecole con funzione immunomodulatrice e sviluppo dei cosiddetti biological response modifiers). Nel settore oncologico, soprattutto nel corso degli anni settanta, sono fioriti numerosi studi e sono state pubblicate numerose ricerche sul significato della teoria della sorveglianza immunologica nel controllo della crescita neoplastica. Qualche anno fa questo aspetto particolare è entrato in fase di revisione critica da molti punti di vista, soprattutto per l'incertezza e la delusione derivate dal fallimento di alcuni protocolli terapeutici basati sul criterio dell'immunoattivazione. Recentemente la scoperta delle interleuchine (IL-1, 2, 3, 4) e alcuni risultati preliminari ottenuti nel campo delle neoplasie in fase avanzata hanno riaperto nuove prospettive terapeutiche.
Tra i settori che indubbiamente hanno ricevuto il maggiore impulso c'è l'immunologia clinica, disciplina ormai autonoma, che spazia dai procedimenti diagnostici alla terapia. È appunto su questo settore che verte il presente articolo, in cui saranno trattati, con particolare riguardo, gli argomenti di più recente sviluppo.
b) I concetti fondamentali dell'immunologia: note storiche
L'antica osservazione che i soggetti guariti da una malattia infettiva raramente contraggono un secondo attacco della stessa è all'origine della disciplina immunologica. Attualmente l'immunologia, sia come area di ricerca sia come strumento di diagnosi e terapia, è una scienza in rapida maturazione che ha raggiunto un buon livello di acquisizioni teoriche su un'ampia serie di problemi correlati al rapporto tra organismo e ambiente. Naturalmente il progresso è stato lento e ha in gran parte seguito l'evolversi delle conoscenze nel settore delle malattie infettive. Soltanto da alcuni decenni il corpo dei concetti e delle informazioni raccolto ha permesso lo sviluppo indipendente e autonomo della ‛scienza della risposta immunitaria'.
È noto che già i Cinesi, nei secoli scorsi, usavano praticare la vaccinazione contro il vaiolo inoculando per via nasale polveri secche di crosticine prelevate da pazienti affetti dalla malattia, e sono noti i diversi tentativi che la specie umana ha fatto nel tempo per ridurre la gravità dei numerosi ‛fenomeni' contagiosi. Dobbiamo arrivare comunque alla storica data del 1798, quando E. Jenner pubblicò i suoi dati sull'inoculo di pus del vaiolo bovino in bambini (osservando l'instaurarsi di una condizione di immunità verso la terribile malattia), per avere un punto di riferimento cronologico preciso. Una prima ‛sistematizzazione' delle conoscenze si ebbe circa un secolo dopo, quando L. Pasteur intuì ed estese i principi generali che costituiscono le fondamenta per l'elaborazione dei vaccini.
Tra la fine del XIX secolo e i primi due decenni del Novecento la complessità dei meccanismi immunitari viene gradualmente svelata, con la dimostrazione dell'esistenza degli anticorpi (si ricordano gli studi di E. A. von Behring e di S. Kitasato, del 1890, sul significato della neutralizzazione antitossica), l'identificazione del complemento (J.-J. Bordet, 1895), l'applicazione di nuove metodiche come la precipitazione e l'agglutinazione. Alla ‛teoria umorale' dell'immunità si affiancava, nel contempo, l'approccio ‛cellulare', grazie ai lavori di E. Metchnikoff sulla fagocitosi. P. Ehrlich conia, nei primi anni del XX secolo, il termine horror autotoxicus e sviluppa le prime teorie sulla natura dei recettori di membrana. Sono tempi fecondi: C. Portier e Ch. R. Richet dimostrano l'esistenza dell'anafilassi (1902), K. Landsteiner focalizza le conoscenze sulla sieroematologia e identifica i gruppi sanguigni ABO; gli studi di H. H. Dale, sul finire del primo decennio, introducono nuove conoscenze sul significato dei mediatori chimici e costituiscono la premessa per l'analisi dei fenomeni immunopatologici. Negli anni trenta viene approfondita l'indagine sulle modalità di interazione fra antigene e anticorpo e M. Heidelberger e J. R. Marrack sviluppano la teoria ‛reticolare' per spiegare alcuni fenomeni riguardanti i legami fra molecole. In questo periodo nasce l'immunochimica, grazie all'applicazione di principi semplici e di tecniche a ottima risoluzione (elettroforesi, immunodiffusione, immunoelettroforesi). È anche del 1930 l'elaborazione, da parte di Haurowitz e Breinl, della ‛teoria istruttiva' (template theory of antibody formation), secondo la quale la molecola anticorpale si modella sul substrato costituito dall'antigene. Negli anni successivi, fino allo scoppio della seconda guerra mondiale, hanno un grande incremento gli studi applicativi sui vaccini e prende corpo l'insieme delle conoscenze fondamentali sul complesso maggiore di istocompatibilità (MHC: major histocompatibility complex).
Ma dobbiamo arrivare al termine degli anni cinquanta per vedere il tramonto delle teorie istruttive e l'imporsi della teoria della ‛selezione clonale' (v. Burnet, 1959), che porta a sintesi compiuta le precedenti osservazioni di Owen (1940-1945) sul chimerismo e di Medawar (1953) sulla tolleranza immunitaria. Alla base della teoria della selezione clonale sta l'ipotesi che un antigene ‛scelga' il proprio recettore nell'ambito delle potenziali cellule immunocompetenti, attivando una risposta altamente specifica nei propri confronti. Ne deriva che il sistema immunitario è geneticamente programmato con la formazione di milioni di linfociti, sensibili, ciascuno, al proprio antigene. Nossal dimostra sperimentalmente l'assunto, evidenziando come una sola specie di anticorpo venga prodotta da una singola cellula (v. Nossal e Lederberg, 1958; v. Nossal, 1987). L'approccio ‛darwiniano' di Burnet (che nel 1960 ha ricevuto il premio Nobel insieme a P. B. Medawar per la scoperta della tolleranza immunitaria) chiude un'epoca e ne apre una seconda.
Gli anni sessanta rappresentano qualcosa di nuovo rispetto al passato e segnano l'inizio di una tappa più avanzata dell'immunologia cellulare, ora al suo apice. Nel decennio precedente erano venuti contributi essenziali dai lavori di Porter ed Edelman sulla struttura e sull'eterogeneità delle immunoglobuline. Già nel 1961 Miller e Good identificano il ruolo del timo nella differenziazione linfocitaria e i successivi approfondimenti sul significato dello switch isotipico durante la sintesi di un'immunoglobulina modificano l'interpretazione di alcune fasi essenziali della genesi di catene proteiche. Nella seconda metà degli anni sessanta diventa chiaro come la competenza della risposta immunitaria derivi da una collaborazione fra i linfociti maturati nel timo (T) e quelli provenienti dal midollo osseo (B). Essenziale a questo riguardo il contributo di Claman (1966).
Ci avviciniamo rapidamente ai nostri giorni. Negli anni settanta Jerne postula la teoria del network idiotipico (v. Jerne, 1974), ipotizzando un complesso sistema di regolazione interna fondato sulla mediazione tra recettori di membrana e anticorpi con ‛idiotipi' complementari. Nasce l'immagine di un ‛sistema' immunitario costantemente attivato da segnali umorali e cellulari, positivi e negativi, modulati in funzione della qualità della risposta finale che deve essere prodotta. Nel 1973-1975 vengono definite le classi di antigene codificate dal complesso maggiore di istocompatibilità e viene approfondito il ruolo del rapporto fra aptene e carrier nell'ambito dei meccanismi di stimolo e soppressione. Le conoscenze acquisite nel corso di questi anni sul ruolo dei geni della risposta immunitaria (Mc Devitt e Benaceraff) vengono ampliate dagli studi sulla restrizione genetica delle interazioni cellulari di Zinkernagel e Doherty (v., 1979), i quali forniscono un contributo fondamentale all'analisi della struttura recettoriale dei linfociti T. Un'ulteriore accelerazione del progresso in atto viene dall'applicazione delle tecniche di biologia molecolare all'analisi immunologica, con i lavori di Kohler e Mustein sugli anticorpi monoclonali e le tecniche di ibridazione (v. Mustein, 1986). Il DNA ricombinante e l'uso di sonde genetiche forniscono il metodo di studio per l'analisi della diversità anticorpale durante la sintesi citoplasmatica (v. Tonegawa, 1983; v. biotecnologie, suppl.).
E ora consideriamo lo ‛stato dell'arte' ai nostri giorni, tenendo presente che, al riguardo, si può soltanto tentare una descrizione sommaria del modello che è stato costruito, pronti a trasformarlo dove non sono chiari i passaggi e le connessioni, pronti a cancellarne i punti che si dimostreranno sbagliati, disposti a modificare la prospettiva in cui lavorare sia nella ricerca che nell'applicazione clinica.
Il sistema immunitario funziona secondo un criterio di organizzazione clonale e varie modalità di cooperazione cellulare. Molecole e cellule interagiscono producendo segnali diversi che inducono fenomeni di attivazione e/o soppressione. Il riconoscimento degli antigeni è effettuato nel sistema dei linfociti B dalle immunoglobuline di membrana, e il prodotto delle cellule B, trasformate in plasmacellule dopo l'incontro con l'antigene, è l'anticorpo appartenente a una delle cinque classi isotipiche (IgG, IgA, JgM, IgD e IgE). Fino a tre anni fa non si conosceva la struttura equivalente alle Ig nei linfociti T; oggi sappiamo che i linfociti T maturi possiedono un recettore per l'antigene (T1) o T idiotipo, costituito da una struttura binaria di catene alfa e beta unite da ponti disolfuro, come accade per le Ig, e fornite sia di una parte costante che di una frazione variabile. Il recettore dei linfociti T appare anche nelle cellule timiche mature e subisce fenomeni di riarrangiamento genetico. Il processo è analogo a quello che subiscono le cellule B immature, nelle quali il riarrangiamento genetico delle immunoglobuline precede le tappe finali che definiscono il fenotipo maturo. Il recettore dei linfociti T è unito alla molecola T3 (CD3) che è costituita a sua volta da tre catene polipeptidiche assemblate a formare il cosiddetto T3-complex o unità di riconoscimento dell'antigene.
2. Immunodeficienze primitive
a) Classificazione e diagnostica.
Il settore delle immunodeficienze primitive (IDP) comprende una patologia molto ampia la cui caratterizzazione nosografica subisce un costante aggiornamento e una continua revisione critica: la tab. I riporta la classificazione più aggiornata (1985) dell'OMS.

Le IDP sono state definite ‛esperimenti della natura' e, al di là dei problemi clinici e umani che pur rappresentano, hanno fornito un indubbio contributo alla conoscenza della struttura e delle funzioni delle varie componenti del sistema immunitario. Un modo relativamente semplice per inquadrare quanto è noto sulle modalità della risposta immunitaria consiste nel trattare separatamente i componenti e i fattori aspecifici da quelli con funzioni più selettivamente orientate dall'antigene. Le condizioni di immunodeficienza, parziale o totale, per causa sia congenita che acquisita, possono riguardare i due gruppi di funzioni del sistema immunitario: le aspecifiche e le specifiche. Quali che siano i difetti, ne deriva un'alterazione della risposta nei confronti dei numerosi agenti patogeni: virus, batteri, funghi e parassiti. Gli agenti patogeni, a seconda delle loro proprietà e caratteristiche, suscitano reazioni particolari. Il sistema muco-cutaneo rappresenta la prima difesa di ‛barriera': oltre che delle sue caratteristiche meccaniche, si giova di secrezioni contenenti enzimi o altre sostanze in grado di limitare l'aggressività dei microrganismi invasori (per esempio il lisozima, che può digerire alcuni mucopeptidi della parete batterica). Le difese contro batteri, virus e funghi utilizzano vari meccanismi. Gli anticorpi hanno una funzione opsonizzante che può esercitarsi contro i virus - determinando la loro agglutinazione e bloccando l'ingresso delle particelle dentro la cellula bersaglio - o contro alcuni batteri a crescita extracellulare. In questo caso le immunoglobuline prodotte verso gli antigeni di stafilococchi, streptococchi ed escherichie riconoscono la cellula e stimolano la fagocitosi da parte di alcuni globuli bianchi (neutrofili) o attivano la lisi mediante complemento. Se alcuni batteri crescono all'interno della cellula che li ha fagocitati, intervengono altri processi di controllo mediati dai macrofagi o dal sistema timo-dipendente. Una situazione analoga è rappresentata dalla difesa nei confronti delle infezioni fungine, che necessita dell'integrità del sistema cellulo-mediato. Negli ultimi anni hanno assunto un ruolo sempre più importante le informazioni acquisite sugli interferon (IFN), lipoproteine che svolgono azione antivirale ma anche immunoregolatrice. Le molecole di IFN (alfa, gamma, beta) sono in grado di influenzare i processi metabolici di varie cellule agendo a livello della sintesi di RNA e di catene amminoacidiche. Un organismo perfettamente integro può cadere vittima di patogeni ad alta virulenza (che temporaneamente sopprimono le difese immunitarie), ma, quando la risposta complessiva è efficace (grazie anche all'impiego di antibiotici), l'infezione viene mantenuta sotto controllo e si conclude con la guarigione. Viceversa, quando individui con lesioni congenite o acquisite del sistema immunitario (soggetti immunodeficienti) vengono a contatto con microrganismi altrimenti non patogeni (opportunisti), possono andare incontro a gravi malattie infettive.
Le IDP riguardano sia l'immunità umorale che quella cellulare; per uno studio dei difetti a carico dell'immunità cellulare e umorale con azione specificamente orientata si può fare riferimento allo schema della fig. 5, che illustra gli stadi di sviluppo e di differenziamento della linea linfocitaria.
Quando il difetto ha sede a livello (A) si hanno le conseguenze patologiche più gravi: viene bloccato ogni differenziamento e le tappe successive non sono possibili. La forma clinica prototipo è l'immunodeficienza grave combinata. Se l'alterazione riguarda il timo (B) si ha un difetto regolativo e anche il sistema umorale ne risente. La sindrome di DiGeorge ne è il prototipo completo. A livello (C) il sistema di regolazione è solo parzialmente noto e non si conoscono le molecole differenziatrici del microambiente bursale (fegato fetale e midollo osseo nella specie umana). Il prototipo delle forme cliniche che interessano questo livello è l'agammaglobulinemia legata al sesso (nota anche come sindrome di Bruton). In periferia (D), quando è superata la tappa maturativa degli organi linfatici centrali, si hanno alterazioni essenzialmente di tipo regolativo e i difetti che ne derivano dipendono da più cause concomitanti.
La più nota e drammatica delle ID acquisite è il difetto secondario conseguente a infezione da virus HIV (sindrome dell'immunodeficienza acquisita).
Descriveremo di seguito alcuni esempi delle forme cliniche più significative nell'ambito dei difetti dell'immunità cellulare e umorale. Fra le forme con prevalente difetto anticorpale verranno analizzate l'agammaglobulinemia legata al sesso e il difetto selettivo delle IgA; nel gruppo dell'immunodeficienza combinata saranno descritte l'immunodeficienza grave combinata e quella comune variabile. La sindrome di DiGeorge, la sindrome di Wiskott-Aldrich e l'atassia-teleangectasia verranno quindi analizzate per il loro importante significato clinico e biologico.
b) Agammaglobulinemia congenita legata al sesso
Definita anche ‛agammaglobulinemia di Bruton', dal nome del medico che la identificò all'inizio degli anni cinquanta (v. Bruton, 1952), l'agammaglobulinemia legata al sesso si può considerare la prima malattia da immunodeficienza descritta. Si caratterizza per l'assenza di immunoglobuline di tutti gli isotipi e per l'alta frequenza di infezioni batteriche ricorrenti. Sono noti due tipi genetici: la forma X-linked (tipo Bruton) recessiva e una forma autosomica anch'essa recessiva (v. Hoffman e altri, 1977).
Il difetto base è a carico della linea cellulare B (blocco maturativo pre-B). La maggior parte degli studi svolti sul sistema T ne hanno evidenziato la normalità. Sono assai rare sia le plasmacellule che i linfociti B circolanti. I linfociti pre-B appaiono in numero normale nel midollo osseo. Alcuni esperimenti condotti in vitro hanno dimostrato che linfociti B del sangue periferico provenienti da soggetti agammaglobulinemici sono in grado di produrre anticorpi se opportunamente stimolati.
Indagini condotte sulla catena μ del citoplasma derivato da linee linfoblastoidi confermano che essa è più piccola del normale e difetta di alcune zone della parte variabile. L'IgM di membrana, assieme all'immunoglobulina IgD, svolge un ruolo critico nella differenziazione della linea B, per cui, se difetti intrinseci o squilibri del microambiente modificano la regolarità dei vari passaggi, ne consegue il blocco funzionale (v. Coffman e Weissmann, 1983; v. Cooper e altri, 1984; v. Tedder e altri, 1985). Recenti dati propongono l'ipotesi che l'agammaglobulinemia X-linked dipenda da un difetto dei processi regolativi che controllano il riarrangiamento dei geni VH-D-JH. Gli enzimi che consentono la ricombinazione D-JH e VH-D possono essere malfunzionanti e quindi determinare un riarrangiamento non produttivo delle molecole Ig.
Esistono inoltre alcuni dati sulla presenza di un notevole numero di cellule con funzione suppressor (Ts) nel sangue periferico dei soggetti colpiti da questa malattia, ma non si posseggono prove definitive sul reale significato patogenetico dei Ts. Queste cellule possono essere un semplice fenomeno secondario (per esempio, coltivando in presenza di linfociti soppressivi cellule B, esse mantengono l'incapacità di sintesi se le cellule T ad azione soppressiva vengono eliminate dal medium).
Il quadro clinico è abbastanza tipico. I neonati di sesso maschile affetti da agammaglobulinemia congenita godono in genere di buona salute fino al quarto-sesto mese di vita. In pratica finché le gammaglobuline IgG materne sono in circolo il bambino è sufficientemente protetto. Verso il secondo anno cominciano i primi episodi infettivi sostenuti da pneumococchi, streptococchi, stafilococchi; è spesso possibile isolare Haemophilus influenzae. Si assiste alla comparsa di infezioni delle alte e basse vie respiratorie (sinusiti, faringiti, polmoniti) e dell'orecchio (otiti); non rare le sepsi; meno frequenti, sebbene non eccezionali, le forme di pioderma e di meningite. Questi pazienti, che prima dell'era antibiotica andavano inevitabilmente incontro all'exitus, raggiungono ai nostri giorni il terzo e quarto decennio di vita, ma spesso con gravi esiti quoad valetudinem (bronchiectasie e insufficienza respiratoria). Rari sono i fenomeni di malassorbimento, sebbene sia comune una giardiasi diffusa. I virus che provocano gli esantemi infantili non causano complicazioni particolari; lo stesso accade per miceti o protozoi. Le infezioni da Pneumocystis carinii possono, invece, essere frequentemente mortali. Le infezioni virali, alle quali gli agammaglobulinemici sembrano essere molto suscettibili, sono quelle provocate dal virus dell'epatite B, dai virus Echo (Echo 30) e dal gruppo dei polio (v. VIRUS). La vaccinazione con agenti polio attenuati può causare forme paralitiche ad andamento ingravescente. Si associano anche fenomeni allergici, reazioni da farmaci, eczema atopico. In oltre la metà dei soggetti agammaglobulinemici si sviluppa una sindrome assai simile all'artrite reumatoide: dati sull'isolamento di un micoplasma consentono di ipotizzare un'eziologia infettiva di questa complicazione (v. Stuckey e altri, 1978). È stata anche segnalata una sindrome connettivitica simildermatomiositica con rash cutanei, edema e indurimento delle masse muscolari. In apparente contrasto con la gravità e la frequenza degli episodi infettivi, l'esame bioptico degli organi linfatici rivela assenza di plasmacellule anche dopo stimolo e disorganizzazione strutturale con deplezione sia dei follicoli che dei centri germinativi. Caratteristica è l'ipoplasia tonsillare.
La diagnosi di laboratorio ha estremo valore. Il dato istologico ai fini di una corretta diagnosi di agammaglobulinemia congenita consiste nell'assenza di plasmacellule nei linfonodi e in altri distretti linfatici secondari. Le raccomandazioni dell'OMS consigliano tre tipi di indagine per lo studio dei difetti dell'immunità umorale: a) valutazione degli anticorpi naturali come le isoemoagglutinine A e B, le eterolisine o le eteroagglutinine, le streptolisine e gli anticorpi con capacità battericida nei confronti di E. coli; b) valutazione della presenza di anticorpi rivolti verso gli antigeni di comune impiego per le vaccinazioni (tetano, difterite, poliomielite); c) immunizzazione attiva.
Stimolando con antigeni un bambino con sospetto diagnostico e prelevando i linfonodi, la ricerca è positiva se si nota l'assenza di plasmacellule e di cellule attivate. Clinicamente indicativa è l'ipoplasia tonsillare. Non sono rilevabili alterazioni nel timo. Per il rischio di errore nei confronti dell'agammaglobulinemia transitoria dell'infanzia è utile l'esecuzione di controlli ravvicinati nel tempo del dosaggio di immunoglobuline.
La prognosi è oggi assai migliorata rispetto al passato, sia per l'uso di antibiotici che per il massiccio impiego endovenoso di immunoglobuline. Le lesioni acquisite nella prima infanzia riducono a volte le prestazioni fisiche nel corso degli anni e possono determinare seri problemi quoad vitam. Le complicazioni virali, soprattutto a carico del sistema nervoso, causano non di rado lesioni della corteccia ed exitus. L'instaurarsi della manifestazione simil-reumatoide in un certo numero di pazienti è un altro elemento negativo quoad valetudinem. È segnalata la comparsa di neoplasie a carico del sistema linfatico.
Nell'ultimo decennio si sono compiuti notevoli progressi nel settore della terapia sostitutiva con anticorpi. La somministrazione di immunoglobuline umane normali (di solito IgG ottenute con il metodo di preparazione etanolica) è tollerata per via intramuscolare ma non per via endovenosa. Si manifestano infatti gravi reazioni sistemiche che tendono a essere più frequenti proprio in soggetti agammaglobulinemici. L'origine delle reazioni è l'attività ‛anticomplementare' delle molecole iniettate (v. Barandun e altri, 1980), ma vari problemi sono correlabili con la quantità di prodotto somministrato e con l'attivazione indiretta di processi infiammatori (liberazione di sostanze vasoattive). I numerosi svantaggi della via intramuscolare (modeste quantità iniettabili, azione locale di proteolisi con ritardo di assorbimento, non buona disposizione psicologica del paziente, reazioni locali e sistemiche) hanno stimolato varie tecnologie farmaceutiche per produrre molecole di IgG adatte alla somministrazione endovenosa.
Per la preparazione di immunoglobuline somministrabili per via endovenosa sono stati proposti vari metodi: trattamento enzimatico (pepsina, plasmina), procedimenti fisicochimici (pH 4, betapropiolattone, riduzione-alchilazione, sulfonazione, miscelamento con polietilenglicole), trattamenti fisici (adsorbimenti). Tali metodi presentano vantaggi e svantaggi, ma in pratica nessuna preparazione è completamente priva di inconvenienti, ed effetti collaterali minori sono osservati sia durante che dopo l'infusione. I gravi fenomeni anafilattici sono eccezionali e in parte imputabili a iperreattività individuale; spesso si possono controllare con opportune modalità di somministrazione (per esempio minore velocità di gocciolamento per frazione tempo). Attualmente la dose media è di 200-400 mg/kg (peso) ogni 14-21 giorni, fino al raggiungimento di valori sierici di IgG non inferiori ai 500 mg/dl. Indubbi vantaggi clinici sono evidenti a breve-medio termine, e continuano nel corso degli anni garantendo al paziente una qualità di vita molto buona.
c) Difetto selettivo di IgA
Questa sindrome rappresenta il difetto più frequente nell'ambito della risposta umorale. Entrambi i sessi sono colpiti e le conseguenze cliniche sono in generale compatibili con una vita normale. In una quota variabile dal 20 al 40% dei casi si associano malattie infettive, fenomeni autoimmuni e allergici. Sembra accresciuto anche il rischio di contrarre alcuni tipi di neoplasia. L'incidenza dei fenomeni allergici e autoimmuni è significativamente più alta rispetto ai controlli normali.
Il difetto di IgA può essere ereditato come forma dominante autosomica (v. Ooldberg e altri, 1968) o come carattere recessivo (v. Stocker e altri, 1968). Talvolta sono associate anormalità cromosomiche (v. Burgio e altri, 1980).
Gli studi sulla distribuzione delle IgA di membrana hanno messo in evidenza nel sangue periferico dei portatori di questo difetto cellule B recanti l'isotipo specifico, per cui è stata prospettata l'ipotesi di un blocco terminale del differenziamento, conseguente a un danno proprio del linfocita B (v. Cooper e Lawton, 1972). Ulteriori dati sperimentali hanno però evidenziato un più complesso gruppo di alterazioni: poiché la fase finale di differenziamento cellulare dei linfociti B è sotto il controllo delle popolazioni T, si è ipotizzato che il difetto principale risieda in tali cellule (v. Elson, 1985). I pazienti si dividono in due gruppi: quelli con difetto prevalente delle funzioni helper e quelli con un eccesso dell'attività suppressor. L'osservazione (v. Adachi e altri, 1983) di un difetto nella popolazione di linfociti T che reca sulla membrana il recettore specifico per le IgA è estremamente interessante sotto il profilo patogenetico: il deficit di IgA può essere la conseguenza di un'anomala regolazione da parte di linfociti T. Più recenti contributi clinici e di laboratorio hanno permesso di acquisire un nuovo concetto: circa il 40% dei soggetti con difetto ‛selettivo' di IgA hanno bassi valori di IgG2 e/o IgG4; talvolta si può associare un difetto di IgG3. Tale associazione può spiegare almeno in parte la differenza tra individui con difetto di IgA senza sintomatologia (riscontro occasionale) e quelli con problemi infettivi (nei quali il difetto di IgA non è unico ma si accompagna ad alterazioni più estese della sintesi immunoglobulinica).
I pazienti con difetto di IgA hanno infezioni ricorrenti, con bronchiti croniche e frequenti fenomeni allergici. Esiste poi una serie di segni gastroenterici di varia gravità ed estensione: diarrea cronica, enterite da Giardia lamblia, enteropatia da glutine, colite ulcerosa, enterite regionale. Un ulteriore gruppo di malattie identificabili in portatori di deficit di IgA è quello dell'autoimmunità: lupus eritematoso sistemico, artrite reumatoide e sua variante giovanile, tiroiditi, anemia perniciosa, anemia emolitica e sindrome di Sjogren sono state descritte in numerosi pazienti. Va inoltre menzionata una particolare reattività alle trasfusioni di sangue o derivati del sangue, che in alcuni casi può dar luogo a fenomeni anafilattoidi anche molto gravi (talora mortali). Queste reazioni sono causate dalla presenza di anticorpi anti-Ig.
La diagnosi immunologica è stabilita quando i valori delle IgA sieriche sono inferiori a 5 mg/dl se il paziente ha raggiunto almeno l'età di due anni. L'assenza delle IgA sieriche si accompagna a quella delle IgA secretorie. Utile è quindi anche lo studio delle IgA nelle secrezioni, i cui valori standard sono comunque assai variabili da soggetto a soggetto. Per la valutazione dei casi con difetto parziale di IgA è molto importante l'età. Ulteriori indagini riguardano il dosaggio delle sottoclassi di IgG. Queste risultano occasionalmente diminuite, soprattutto per i due isotipi IgG2 e IgG4: si sottolinea il concetto che individui con difetto multiplo qualitativo possono essere trattati come ipogammaglobulinemici.
Per completare i dati sierologici sono utili lo studio dei linfociti B (Ig di membrana all'immunofluorescenza) e l'analisi del pannello dei linfociti T: numero totale e sottoclassi (helper e suppressor). È stato recentemente osservato che alcuni pazienti con difetto parziale di IgA si normalizzano con la crescita. La diagnosi differenziale presenta alcune difficoltà nella prima infanzia. La comparsa di fenomeni infettivi durante la crescita, con carattere recidivante, induce a sospettare anomalie della sintesi anticorpale; in questi casi è il laboratorio a dirimere ogni dubbio. Nell'età adulta il reperto può essere occasionale, ma alcuni farmaci anticonvulsivanti o immunosoppressivi possono agire selettivamente causando una transitoria riduzione delle IgA, con andamento reversibile.
La prognosi è in genere buona quoad vitam ma talvolta meno buona quoad valetudinem. Le complicazioni infettive sono frequenti in caso di associazione con i difetti di IgG, sebbene la letteratura sull'argomento non fornisca delucidazioni complete.
d) Immunodeficienza comune variabile
L'immunodeficienza comune variabile (ICV) è uno stato di depressione della sintesi di immunoglobuline che può manifestarsi in ogni periodo della vita (v. Zinnerman e altri, 1954; v. Grant e Wallace, 1954; v. Robbins e altri, 1966) e in entrambi i sessi. Esiste frequentemente una familiarità, sebbene i numerosi studi effettuati non abbiano ancora chiarito le modalità di trasmissione ereditaria (v. Geha e altri, 1974). Tre cause sono possibili: a) un difetto intrinseco dei linfociti B; b) uno squilibrio della regolazione; c) la presenza di autoanticorpi rivolti verso le cellule linfocitarie T o B. I linfociti B sono presenti in circolo ma non rispondono in modo efficace alla stimolazione antigenica in vitro (v. Ciccimarra e altri, 1976). Alcuni pazienti, pur essendo in grado di sintetizzare le immunoglobuline, non hanno capacità di secernerle per un difetto della maturazione (v. Schwaber e Rosen, 1979). Un eccesso di attivazione dei T suppressor è stato inoltre osservato in alcuni casi di ICV. Altri dati riportano presenza di autoanticorpi (v. Tursz e altri, 1977) verso cellule immunocompetenti. Nuovi studi sul ruolo di cellule circolanti di forma dendritica dimostrano anomalie nelle modalità di presentazione dell'antigene. Viene oggi formulata l'ipotesi che le cellule B non siano in condizione di rispondere per un difetto dello stimolo di differenziamento. L'arresto nel processo maturativo sarebbe quindi secondario a un'alterazione della presentazione antigenica (v. Farrant e altri, 1986).
Il quadro clinico non è molto diverso da quello descritto in caso di agammaglobulinemia legata al sesso. Si osserva aumento della suscettibilità a infezioni provocate da piogeni e i pazienti presentano storia di broncopolmoniti recidivanti con esiti bronchiectasici. Un quadro intestinale simil-spruetico, assai più raro nelle forme congenite, è provocato da Giardia lamblia: i soggetti con ICV accusano diarrea, steatorrea, sindrome proteinodisperdente. Sono descritti anche episodi di artrite, che però non sembrano molto frequenti. L'epatosplenomegalia è di frequente osservazione, così come una particolare iperplasia nodulare linfatica a carico dell'intestino (v. Johnson e altri, 1971). In circa un terzo dei soggetti si riscontra anche anemia perniciosa, mentre i granulomi caseosi hanno una distribuzione ubiquitaria e coinvolgono più organi. Rara è l'anemia emolitica autoimmune. Hanno alta incidenza i tumori a carico della mucosa dell'apparato gastroenterico.
I segni clinici dell'ICV sono sovrapponibili a quelli dell'agammaglobulinemia congenita. Un elemento importante di differenziazione è il periodo di comparsa (che in età infantile si manifesta anche dopo i 10 anni). Spesso carattere indicativo assume una sindrome di malassorbimento con enteropatia proteinodisperdente. I fenomeni autoimmuni possono precedere i segni tipici del difetto immunologico: in queste circostanze di solito la diagnosi viene posta tardivamente con danno per il paziente. Forme bronchiectasiche croniche progressive sono osservabili nell'adulto e devono insospettire il medico per un adeguato inquadramento di agnostico: non di rado si riscontra un'anamnesi positiva per lobectomia. Il livello delle IgO sieriche è inferiore a 300 mg/dl (talora con valori compresi tra 200 e 500). Difetti sono stati osservati nei rapporti delle sottoclassi di IgG. È utile approfondire lo studio dell'ICV analizzando la distribuzione delle sottopopolazioni linfocitarie (squilibrio del rapporto belper/suppressor) e le loro caratteristiche funzionali. I linfociti B circolanti nel sangue periferico sono in numero normale o poco al di sotto, mentre nell'agammaglobulinemia legata al sesso essi sono completamente assenti. La prognosi è estremamente variabile, sia quoad vitam che quoad valetudinem. La cronicizzazione della sintomatologia respiratoria è un grave impedimento e può progredire fino alla grave insufficienza. Le donne possono avere gravidanza normale, ma sono segnalati casi con prole affetta da deficit dell'immunità umorale.
L'approccio terapeutico è quello attuato con somministrazione endovenosa di immunoglobuline. La sintomatologia intestinale può risentire beneficio dall'adozione di particolari diete o, nel caso di infestazione con Giardia lamblia, dalla somministrazione di metronidazolo. L'impiego di terapia antibiotica ciclica può migliorare la prognosi nel corso degli anni, sebbene l'inoculazione endovenosa di IgG ad alte dosi abbia modificato sia la qualità che l'attesa di vita di questi pazienti.
e) Immunodeficienza grave combinata
L'immunodeficienza grave combinata (IGC, severe combined immunodeficiency, secondo la terminologia anglosassone) può essere ereditata come forma autosomica recessiva o legata al sesso. In circa la metà dei casi autosomico-recessivi si associa il difetto dell'enzima adenosindeaminasi. Il 70% dei soggetti colpiti dalla patologia sono maschi, ma indipendentemente dalle varianti genetiche l'andamento clinico è sovrapponibile, sebbene con diversa gravità.
La displasia timica è propria di tale quadro morboso, ma con ogni probabilità il difetto primitivo non è localizzato soltanto in questa sede. Il deficit immunologico sembra essere causato da un blocco maturativo o funzionale delle cellule provenienti dal midollo osseo. Una varietà clinicamente molto grave è la cosiddetta ‛disgenesia reticolare', cui è associata una grave neutropenia. Questo difetto è probabilmente a carico delle cellule staminali pluripotenti che non sono in grado di evolvere verso completa maturazione. Se le immunoglobuline sono presenti con una risposta ridotta dell'immunità cellulare e deplezione del tessuto linfoide, si usa di solito l'espressione ‛sindrome di Nezelof'. Quando il difetto è presente sulla linea linfoide staminale si parla di ‛tipo svizzero'. Comunque, sebbene il disordine a carico del timo possa giustificare gran parte dei casi di immunodeficienza grave combinata, il ripristino della branca T-dipendente (v. Hoyer e altri, 1968; v. Rosen, 1968) non si associa costantemente al recupero delle funzioni B. Si conoscono due forme di IGC, in ciascuna delle quali è riscontrabile una distinta anomalia enzimatica: si tratta della IGC associata al deficit dell'enzima adenosindeaminasi (ADA: v. Giblett e altri, 1972) e di quella associata al deficit dell'enzima nucleosidefosforilasi (PNP: v. Giblett e altri, 1975).
L'IGC è una grave malattia dell'età infantile; i soggetti colpiti muoiono nel corso dei primi due anni di vita. Le infezioni sono precoci, si manifestano attorno ai tre mesi di vita e hanno un andamento ricorrente e grave. Il ritardo della crescita, la presenza di una diarrea cronica e le polmoniti sono di riscontro comune. Tipica è la candidiasi dell'orofaringe, dell'esofago e della cute. Non rari sono gli ascessi polmonari da Pseudomonas aeruginosa e pneumoniti da Pn. carinii. La presenza di aftosi, indipendentemente dal trattamento antibiotico prolungato, è costante. L'uso di vaccino antipolio con virus attenuati può causare una poliomielite progressiva, mentre le comuni infezioni virali dell'infanzia assumono un decorso grave e spesso irreversibile (varicella generalizzata, complicazioni polmonitiche del morbillo, infezioni da Citomegalovirus). Una vaccinia progressiva è stata osservata in soggetti sottoposti a vaccinazione antivaiolosa. In sostanza i bambini colpiti da IGC hanno una sensibilità a un ampio spettro di agenti patogeni e diventano facilmente preda di ogni tipo di infezione. Talvolta la malattia è associata a una forma di nanismo ad arti corti. L'assenza o l'ipotrofia dei linfonodi e delle tonsille è un valido ausilio diagnostico. Le radiografie non evidenziano l'ombra timica. Il dato caratteristico dell'IGC è la considerevole eterogeneità delle anomalie immunologiche. La leucocitopenia si accompagna a una marcata linfocitopenia e i linfociti assumono l'aspetto di linfoblasti con i caratteri dell'immaturità. Se l'infezione è prolungata, pur in assenza di una leucocitosi, l'eosinofilia è un reperto comune. Le concentrazioni ematiche di immunoglobuline sono ridotte e spesso, dopo stimolazione in vitro, non si ottiene la produzione di alcun anticorpo. Lo studio dei linfociti B del sangue periferico può dimostrare una distribuzione normale delle cellule con immunoglobuline di membrana. I linfociti T sono ridotti di numero e le prove funzionali, per esempio i test di blastizzazione con fitoemoaggiutinine (PHA), risultano deficitarie (v. Geha, 1976; v. Buckley e altri, 1976; v. Businco e altri, 1981). Il midollo osseo è privo di plasmacellule, linfociti e linfoblasti. I centri germinativi sono assenti nei linfonodi e qualora sia possibile identificare degli isolotti linfocitari, si tratta di cellule che hanno perduto ogni organizzazione strutturale. Poiché il dato istopatologico specifico di una IGC si trova a livello timico, può essere risolutrice una biopsia dell'organo. Le prove in vitro di ipersensibilità ritardata dimostrano una ridotta o assente risposta ai diversi antigeni impiegati (per esempio la Candida) (v. Paulter e altri, 1982). L'IGC è la forma più estesa di difetto immunitario e quindi, sotto il profilo clinico, sia la gravità che la relativa precocità dei sintomi assumono un importante significato prognostico (v. O'Reilly e altri, 1978). La prognosi è inevitabilmente infausta se non vengono adottate alcune terapie immunologiche d'avanguardia.
Il primo tentativo terapeutico è stato fatto mediante trapianto di midollo osseo (v. Gatti e altri, 1968): il ripristino di un adeguato patrimonio cellulare di stem cells ha una funzione ricostitutiva. A tale scopo vengono utilizzati donatori HLA-compatibili. In alcuni individui sono stati ottenuti buoni risultati con trapianto di timo o fegato fetale o epitelio timico. Il miglior metodo per ottenere risultati rimane quello del trapianto di midollo osseo, ma le altre tecniche spesso non sono rinviabili poiché la disponibilità di donatori compatibili è bassa. Il trapianto di midollo osseo da gemelli HLA-identici sembra essere il trattamento di scelta. Infatti i 2/3 dei pazienti trattati in questo modo mostrano una ricostituzione dell'immunità cellulare e umorale e circa il 500/o dei trapiantati è vivo dopo lungo tempo (v. Kenny e Hitzig, 1979). È stato anche proposto il trapianto di cellule ematopoietiche fetali, basato sull'osservazione che l'impiego di tali cellule comporta un minore rischio di una reazione da GvHD (graft vs host disease). Allo stato attuale dell'esperienza, benché dopo trapianto di fegato fetale e timo la GvHD sia mite, non più del 30% dei soggetti con IGC presenta una netta evidenza dell'avvenuta ricostituzione. Nel corso degli anni sono stati ideati anche altri approcci: separazione di cellule staminali su gradienti di densità (v. Dicke e altri, 1973) o trattamento di linfociti con globulina antilinfocitaria (v. Salmon e altri, 1970). I successi sono stati parziali e in casi di IGC con deficit di ADA il miglioramento clinico è stato ottenuto trapiantando midollo come nelle forme senza deficit di ADA. Quando il donatore non è disponibile si possono utilizzare globuli rossi irradiati che, essendo ricchi di ADA, riescono comunque a svolgere una certa funzione ricostitutiva.
f) Sindrome di Wiskott-Aldrich
È una sindrome legata al sesso, di carattere recessivo, con una triade sintomatologica tipica: 1) infezioni ricorrenti; 2) trombocitopenia con gravi emorragie; 3) eczema. In circa un terzo degli individui si associa una neoplasia a carico del sistema linfatico. La patogenesi della sindrome di W-A è di difficile interpretazione. Probabilmente vi è un'inattivazione selettiva midollare a carico dei precursori delle piastrine (v. Akkerman e altri, 1982) e dei linfociti T, mentre questo non si verifica per le emazie, i macrofagi, i polimorfonucleati e i linfociti B. Il difetto riguarda l'immunità umorale, quella cellulare e parzialmente anche le resistenze aspecifiche. Le IgG sono normali mentre le IgM risultano diminuite. Si osserva inoltre un marcato ipercatabolismo per tutti gli isotipi di immunoglobuline, che viene spiegato come conseguenza di un'iperplasia reticoloendoteliale (v. Blaese e altri, 1971). Nei bambini i livelli sierici di isoemoagglutinine sono bassi (a volte non riscontrabili). È deficitaria la risposta nei confronti di antigeni polisaccaridici quali i tipi I e Il dello pneumococco e il tipo VI di E. coli. Le cellule B sono normali. I linfociti T tendono a un progressivo declino nel tempo, sia nel numero che nelle funzioni. La risposta a PHA, concanavalina A (ConA) e pokeweed mitogen (PWM) può essere normale o relativamente depressa; lo stesso si verifica per gli skin tests. I macrofagi sono normali, mentre tavolta è ridotta la capacità chemiotattica dei monociti (v. Altman e altri, 1974). Studi eseguiti sulle piastrine e sulla membrana dei linfociti T provenienti da soggetti con sindrome di Wiskott-Aldrich hanno dimostrato un difetto di glicosilazione a carico di particolari proteine.
Già alla nascita i neonati hanno petecchie e trombocitopenia. Nel primo anno compaiono l'eczema e le manifestazioni infettive (episodi ricorrenti di otite media, enteriti e diarrea). Sono stati descritti episodi di polmonite, sepsi e meningite. Il 20% dei pazienti muore per emorragie non controllabili (episodi intracranici). L'eczema, frequentemente complicato da superinfezioni (batteriche o fungine), è difficilmente distinguibile dall'eczema atopico, e ha una distribuzione irregolare sull'intera superficie cutanea. Le infezioni prevalenti sono batteriche (Streptococcus pneumoniae, Haemophilus influenzae, Staphylococcus aureus), tuttavia alcuni virus (virus dell'herpes simplex, Citomegalovirus, virus del morbillo) possono causare complicazioni fatali. È stata occasionalmente descritta anemia emolitica autoimmune (test di Coombs positivo) refrattaria a numerosi trattamenti terapeutici. La comparsa di linfomi non rappresenta un rischio minore.
L'assenza di IgM è un dato di laboratorio che può confermare la diagnosi; infatti le IgA (spesso aumentate) possono essere entro il range dei valori normali assieme alle IgG. È tipico inoltre il difetto in vivo della risposta al tentativo di immunizzazione con antigeni polisaccaridici. La prognosi non è molto buona e la media di sopravvivenza è inferiore ai sei anni. Molti giovani muoiono per le complicazioni emorragiche.
In alcuni pazienti si è prodotto un miglioramento transitorio usando estratto leucocitario noto come transfer factor (v. Giunchi e altri, 1981). In alcuni soggetti sono stati usati ormoni timici, con risultati incerti. Buono è il risultato del trapianto di midollo osseo: la sostituzione completa del midollo, dopo trattamento radiante o chemioterapico del soggetto per indurre deplezione cellulare, permette di correggere le anomalie presenti nella sindrome. La splenectomia è utile per determinare un aumento dei valori delle piastrine circolanti. L'uso di antibiotici e un accurato monitoraggio dei pazienti possono consentire un miglioramento della prognosi.
g) Sindrome della terza e quarta tasca branchiale (Di George's syndrome)
È una sindrome complessa, caratterizzata da difetti nelle strutture anatomiche che derivano dalla terza e dalla quarta tasca faringea. Sono presenti anomalie dell'arco aortico, delle paratiroidi e del timo. Il sistema immunitario T-dipendente è alterato a causa dell'ipoaplasia timica. La malattia sembra conseguente a danni subiti dal feto nel corso della vita intrauterina durante il periodo precedente la dodicesima settimana di gestazione.
Il danno fondamentale colpisce il timo (v. DiGeorge, 1968); tale organo (derivato dall'epitelio endodermico della terza e quarta tasca faringea) comincia a svolgere il suo ruolo immunologico primario attorno all'ottava settimana di gestazione, quando è popolato da cellule staminali. Conseguentemente il danno è variabile in relazione all'estensione del difetto: ci sono pazienti con gravi anomalie della risposta T e altri il cui assetto è sostanzialmente normale e nei quali sono prevalenti le altre malformazioni. Comunque la maggior parte dei soggetti con questa sindrome ha piccoli lobi timici ectopici. Il numero dei linfociti circolanti è normale o lievemente ridotto.
Estremamente importanti sono i test funzionali per l'immunità T-dipendente, che risulta nettamente depressa sia in vitro che in vivo. I linfociti proliferano poco se stimolati con PilA e gli skin tests forniscono risultati negativi. Inoltre è stato osservato un prolungamento della sopravvivenza nel caso di trapianto allogenico della cute. La biopsia dei linfonodi conferma una riduzione di cellularità nelle cosiddette aree timo-dipendenti (zona paracorticale).
La sindrome ha un esordio precoce a causa della comparsa di tetania neonatale accompagnata da ipocalcemia (iposviluppo paratiroideo). I difetti cardiaci, se gravi (tetralogia di Fallot), costituiscono causa di morte. Sono stati descritti anche atresia esofagea, uvula bifida e ipotiroidismo. Inoltre i soggetti colpiti hanno una facies tipica caratterizzata da micrognatia, ipertelorismo, anomalie del lobo auricolare e un particolare taglio degli occhi descritto in senso ‛antimongoloide'. Spesso i bambini decedono nell'infanzia. Candida albicans e alcuni protozoi sono gli agenti più frequentemente identificati nella genesi delle infezioni, che consistono essenzialmente in polmoniti ricorrenti, sinusite cronica ed enteriti con diarrea.
Sotto il profilo laboratoristico il dato immunologico è importante e può aiutare nella prognosi. Infatti l'esistenza di un sistema immunitario T-dipendente, anche parzialmente funzionante, è in grado di garantire una sufficiente difesa contro vari patogeni. È dunque necessario valutare il numero dei linfociti T circolanti, la risposta in vitro (test di blastizzazione con PilA) e in vivo (uso di multitest standardizzati) (v. Adami e altri, 1981). Il dosaggio delle immunoglobuline sieriche può confermare la normalità del sistema B-dipendente. Il difetto immunologico può essere corretto con impianti di timo fetale (v. Cleveland e altri, 1968) o con l'impiego di estratti timici umorali (v. Aiuti e altri, 1980; v. Aiuti e Businco, 1983). Naturalmente, assieme al trattamento del difetto immunitario, vanno garantiti altri presidi terapeutici, come l'impiego di gluconato di calcio, vitamina D e così via, per i gravi disturbi conseguenti all'ipoparatiroidismo e all'ipocalcemia.
h) A tassia-teleangectasia
L'A-T è una sindrome caratterizzata da un complesso quadro patogenetico nel quale l'anomalia di base sembra essere un difetto di riparazione del DNA. Forma autosomica recessiva (v. Spector e altri, 1982), si manifesta clinicamente con atassia cerebellare, teleangectasie oculocutanee, infezioni e alto rischio di neoplasie.
Rotture e translocazioni cromosomiche sono state riportate nella letteratura e riguardano prevalentemente i cromosomi 6, 7, 14 e X. È stato descritto un particolare effetto citolesivo delle radiazioni ionizzanti, mentre non sarebbero efficaci in tal senso i raggi ultravioletti. Un alterato controllo genetico viene oggi considerato il primum movens estrinsecantesi in diversi settori. Alcuni pazienti presentano alti livelli di alfa-fetoproteina sierica e di antigene carcinoembrionario. Sono state descritte alterazioni sia a carico dell'immunità cellulo-mediata che nel settore umorale (v. Strober e altri, 1968). Ipoplasia o aplasia timica sono spesso presenti, con linfocitopenia e ridotta risposta agli skin tests. Nell'80% dei pazienti si è osservato un difetto della sintesi anticorpale a carico delle IgA, IgE, IgG2, IgG4; è stata inoltre segnalata la presenza di un'IgM a basso peso molecolare. Un difetto selettivo nella risposta ad alcuni particolari antigeni può giustificare l'alta frequenza di infezioni. Studi sulla risposta immunitaria hanno messo in luce un ruolo delle cellule T-soppressive (v. Fiorilli e altri, 1983); altri studi sulla regolazione immunitaria hanno invece messo in evidenza un difetto dei T-helper, con normalità delle funzioni suppressor. Esiste l'ipotesi che le alterazioni immunologiche nell'A-T siano dovute ad alterazioni cromosomiche (v. Fiorilli e altri, 1986) e alcune osservazioni hanno trovato una spiegazione recentemente. Allo stato attuale delle conoscenze non è stato possibile fornire un'interpretazione patogenetica definitiva della malattia, soprattutto tenendo conto del coinvolgimento di molteplici sistemi; al rignardo appare comunque significativa l'esistenza di un'alterazione dei tessuti germime conseguente al difetto di riparazione del DNA. Un peptide con basso peso molecolare potrebbe avere la responsabilità delle lesioni strutturali del DNA (v. Shaham e altri, 1982); certamente l'identificazione del peptide nel liquido amniotico fornirebbe un'utile informazione per la diagnosi prenatale.
I segni clinici si manifestano quando il bambino con A-T inizia a camminare: compaiono atassia con movimenti coreo-atetosici, scosse miocloniche e aprassia oculomotoria. L'anatomia patologica rivela la natura degenerativa delle lesioni cerebrali, con scarsi caratteri infiammatori. A livello congiuntivale e quindi cutaneo appaiono dilatazioni venulari (teleangectasie). Nel corso del tempo si presentano i caratteri di atrofia generalizzata della cute, con diminuzione del contenuto adiposo, comparsa di vitiligine e di altre discromie. Il difetto immunitario si evidenzia con un accresciuto rischio infettivo che è presente in oltre il 90% dei soggetti colpiti. Il sistema respiratorio è spesso coinvolto e sono frequenti sia le bronchiti che le broncopolmoniti. In base a quanto segnalato nei Registri nazionali delle immunodeficienze primitive (v. Luzi e altri, Il Registro..., 1986) c'è un accresciuto rischio di neoplasie.
Le prove di laboratorio utili per uno screening generale riguardano il dosaggio delle immunoglobuline sieriche (IgG, IgA, IgM e IgE) e lo studio delle sottoclassi di IgG. I test di risposta in vitro con biastizzazione dopo stimolo con PHA, o utilizzando altre sostanze, sono deficitari. La maggior parte dei pazienti è priva di IgA secretorie e frequente è la presenza di anticorpi anti-IgA. I deficit dell'immunità cellulare hanno varia espressione: il timo è ipoplasico e la biopsia può rivelare una deplezione delle aree periferiche paracorticali. La prognosi è infausta, dal momento che una delle cause principali dei decessi è dovuta alla comparsa di neoplasie. Non esiste una terapia risolutiva per l'A-T. La somministrazione di gammaglobuline va attuata nei soggetti con particolari problemi infettivi e mantenuta comunque in funzione profilattica. Il transfer factor è stato usato in alcuni casi senza reale successo per il decorso generale della malattia.
3. La sindrome da immunodeficienza acquisita
a) Introduzione
Le immunodeficienze secondarie, che costituiscono un insieme eterogeneo di quadri patologici, sono associate a eziologie primitive o prevalenti e, almeno per gran parte dei casi, esiste una buona reversibilità se viene eliminata la causa del difetto. Le cause più note sono la malnutrizione (diminuito assorbimento di proteine, deficienza selettiva di alcuni oligoelementi quali lo zinco e il ferro), infezioni virali (morbillo) e batteriche (lebbra, lue, tubercolosi, infezioni ad andamento cronico), malattie neoplastiche, sindrome nefrosica e nefropatie croniche, impiego di farmaci immunosoppressori, alcune sindromi dismetaboliche. Il sistema immunitario, sia ad azione specifica che ad azione aspecifica, può essere coinvolto a diversi livelli dalle manifestazioni di immunodeficienza secondaria. Comunque la più drammatica espressione clinica di un difetto acquisito dell'immunità, che ha ricevuto nel corso degli ultimi anni una grande attenzione internazionale, è l'AIDS (acquired immunodeficiency syndrome). Ci soffermeremo quindi essenzialmente su tale sindrome, che ha aperto un nuovo e importante capitolo della medicina contemporanea.
Le prime indagini epidemiologiche sull'AIDS iniziarono negli Stati Uniti dopo l'osservazione di un elevato e inusuale numero di casi di polmonite da Pneumocystis carinii, segnalati già prima del 1981 in omosessuali attivi di sesso maschile residenti nelle grandi città americane o che avevano soggiornato ad Haiti. Dal sospetto che la sindrome fosse trasmissibile alla dimostrazione che l'agente responsabile è un virus trascorse poco tempo e sia negli Stati Uniti (gruppo di R. Gallo: v. Gallo e altri, 1984) che in Francia (gruppo di L. Montagnier: v. Montagnier e altri, 1984) vennero pubblicati i primi lavori sull'agente patogeno ritenuto responsabile: l'HIV (human immunodeficiency virus) (v. Broder e Gallo, 1984; v. Wong-Staal e Gallo, 1985).
b) L'HIV: un particolare retro virus
I Retrovirus costituiscono una famiglia di virus diffusa in numerose specie animali e presentano alcune caratteristiche genetiche e autoreplicative del tutto peculiari (v. VIRUS). Il loro genoma è ‛scritto' in RNA, ma, tramite l'azione di un enzima (transcriptasi inversa), può essere trascritto in DNA e così integrarsi nel genoma della cellula ospite. In linea di massima, per quel che ne sappiamo finora, il danno causato dai Retrovirus è piuttosto lento, di tipo cronico. Tuttavia l'HIV (già noto come HTLV-III) è un'eccezione: provoca un danno cellulare molto grave in tempi relativamente brevi e si associa a fenomeni clinici sia acuti che cronici (v. Schneider e altri, 1987). Il genoma di un retrovirus è costituito da tre geni strutturali che assumono il nome di GAG, POL ed ENV. Le proteine della parte più interna del virus (core) sono codificate dal GAG, il POL codifica per la transcriptasi inversa e l'ENV per il rivestimento esterno (envelope). Il processo chiave dell'infezione e delle premesse per il successivo danno cellulare è quello legato alla funzione della DNA-polimerasi RNAdipendente (transcriptasi inversa) che consente la lettura del genoma a RNA e la costituzione del DNA virale. Sono noti i due virus oncogeni HTLV-I e HTLV-II, molto simili tra loro e al virus della leucemia bovina. L'HTLV-I è correlato alla leucemia a cellule T dell'adulto e ha spiccate proprietà di attivazione della replicazione cellulare. L'HIV, al contrario, non produce trasformazioni nella cellula bersaglio contagiata, ma funge da virus litico: uccide la cellula. Sono comunque tutti virus linfotropi e la cellula preferenzialmente colpita dall'HIV è il linfocita T-helper (CD4+). Anche altre cellule del sistema immunitario vengono infettate e altri tessuti risultano colpiti (il sistema nervoso centrale costituisce un bersaglio altrettanto privilegiato e può subire danni diretti provocati dal virus o essere aggredito da vari agenti patogeni che non risultano più controllabili dalle regolari funzioni del sistema immunitario). I virus umani, rispetto a quelli animali, sono dotati di almeno un gene in più (TAT = gene transattivante, in grado di attivare varie porzioni di DNA) e l'organizzazione dell'HIV è ancora più complessa per almeno due fattori: l'esistenza di altri geni (non meno di tre) codificanti per proteine non strutturali e l'alta variabilità genetica (instabilità del genoma?). Le origini geografiche e biologiche dell'HIV non sono del tutto chiare, ma si ritiene che la diffusione del virus nella specie umana stia avvenendo ai nostri giorni per la prima volta nella storia dell'evoluzione. Probabilmente gli HTLV hanno origine animale e provengono dalla famiglia delle scimmie verdi africane. Il rapporto di vicinanza scimmia-uomo in alcune aree africane e l'affinità genetica tra i virus STLV-1 e STLV-III, propri della scimmia, e il virus HTLV-I giustificano questa ipotesi. D'altra parte, poiché sembra che i virus HTLV siano stati acquisiti anche dalle stesse scimmie, è assai arduo individuarne l'origine.
In Africa l'HIV è molto diffuso e le modalità di contagio sembrano essere dovute essenzialmente alla via eterosessuale. Due ipotesi ‛elementari' possono spiegare il coinvolgimento della specie umana in questa pandemia: l'STLV-III è riuscito a contagiare l'uomo e nel più prevedibile dei processi di adattamento si è evoluto in una forma altamente aggressiva (l'HIV), oppure il processo di adattamento è stato più lento e l'STLV-III si è trasformato in un primo tempo nel virus HIV-2 (recentemente identificato nell'Africa occidentale) per poi progredire ulteriormente e trasformarsi in HIV. Il virus HIV-2 è egualmente patogeno e dà una forma simile all'AIDS causata dall'HIV (oggi classificato anche HIV-1). Poiché il patrimonio del suo ENV varia del 57% rispetto al virus HIV è da considerare come un nuovo retrovirus umano.
c) Il danno immunologico e la patologia cellulare
La cellula bersaglio dell'HIV è il linfocita CD4+, che esprime sulla sua membrana un recettore selettivo per il virus. Quando è invaso dal virus, il linfocita CD4+ va incontro a una crescita alterata con un rallentamento progressivo e quindi la lisi. Il drammatico ridursi di questa popolazione linfocitaria ha conseguenze fondamentali: i linfociti B non ricevono le informazioni modulatrici per la sintesi di anticorpi specifici, i T-suppressor (CD8 +) sono privi dello stimolo regolatore e non controllano la sintesi di immunoglobuline (che, in forma policlonale, vengono sintetizzate e secrete in grande quantità), i mediatori chimici della risposta immunitaria sono quantitativamente e qualitativamente alterati (IL-2 è a livelli molto bassi, l'interferon gamma è depresso e sia i macrofagi che le cellule natural killer risultano ipostimolate). Il danno maggiore esercitato dal virus non è selettivamente ristrettò a livello di alcune cellule T: infatti è possibile che anche i linfociti B vengano colpiti, così come i macrofagi e probabilmente le piastrine. Per quanto riguarda i linfociti B, se analizzati in coltura essi dimostrano una spiccata tendenza alla secrezione spontanea di immunoglobuline e all'attivazione policlonale, sebbene risulti precocemente alterata la distribuzione degli isotipi (v. Clifford Lane e altri, 1983; v. Luzi e altri, Analysis..., 1986). È anche evidente una diminuzione (v. Pahwa e altri, 1985) della risposta linfoproliferativa a mitogeni B (T-indipendenti). È probabile che il danno nel settore linfocitario della risposta cellulare in corso di AIDS sia estremamente complesso e non si limiti all'unica sottopopolazione linfocitaria T (v. Clifford Lane e altri, 1983; v. Zolla-Pazner, 1984). Non solo, ma possono essere infettate cellule endoteliali, cellule epiteliali e glia. La glia, in modo particolare, può essere considerata una vera e propria sede privilegiata: essa funge da serbatoio di riserva per l'immissione in circolo di nuove particelle virali. Gli studi più recenti di immunologia molecolare hanno messo in luce l'esistenza di particolari fattori di soppressione, liberati dagli stessi linfociti infetti, che interferendo su altri linfociti-helper ne causerebbero una cecità di membrana bloccando la capacità di riconoscere altri antigeni. Studi sulla sensibilità individuale all'infezione da HIV hanno chiarito solo parzialmente il ruolo di alcuni fattori HLA-correlati, aprendo comunque uno spiraglio sul significato della predisposizione genetica. Dati ancora più interessanti derivano da alcune ricerche condotte su omosessuali, ricerche che hanno dimostrato l'esistenza di una proteina associata alla ridotta vulnerabilità all'AIDS. In particolare esisterebbero tre varianti fenotipiche, o sottotipi, di tale molecola, indicate con le sigle Gclf (fast), Gcls (slow), Gc2: i portatori del fenotipo Gc2 avrebbero una ridotta probabilità di contrarre il virus HIV. In corrispondenza di queste varianti, percentualmente diffuse nella popolazione normale secondo la distribuzione Gcls > Gc2 > Gclf, si è riscontrata una diversa concentrazione di acido sialico, che risulta massima in presenza di Gclf, nulla in presenza di Gc2, media in presenza di Gcls. Si è pensato che l'acido sialico svolgesse un ruolo non secondario nella fase di legame del virus HIV al recettore presente sulle cellule T-helper, ma ulteriori dati di più recente acquisizione non hanno confermato questa ipotesi.
d) L'anatomia patologica del linfonodo
La biopsia linfonodale dimostra alcuni reperti tipici che aiutano sia l'approccio clinico che l'eventuale prognosi (v. Racz e altri, 1986). Attualmente l'evoluzione della risposta tessutale del linfonodo all'infezione da HIV comprende almeno tre fasi: subito dopo il contagio iniziale si assiste a un'iperplasia reattiva cui fa seguito, con modalità e tempi diversi, una fase regressiva (clinicamente associata all'ARC, AIDS related complex, e a certe LAS, sindromi linfoadenopatiche), mentre il quadro finale associato all'ATDS è quello della deplezione linfocitaria. Tipico della fase iperplastica è il quadro istopatologico definito ‛a cielo stellato', con aumento numerico dei centri germinativi e loro ingrossamento volumetrico. L'iperattivazione dei centri germinativi giustifica il dato sierologico dell'aumentata produzione di immunoglobuline. In prima approssimazione l'anatomopatologo non attribuisce a questo aspetto morfologico i caratteri della specificità, sebbene risulti già possibile mettere in luce alcune cellule in fase regressiva precoce. La comparsa di emorragie nel centro germinativo unitamente a una diminuzione del numero delle cellule è un indice di regressione, che evolve gradualmente con fenomeni di necrosi centrale e perdita dell'intera struttura. Nel suo evolvere terminale il linfonodo appare depleto, con cellule similepiteliali e commistione dei vari elementi cellulari. Non sono più riconoscibili le aree funzionali e conseguentemente risulta impossibile un'adeguata risposta all'antigene. È estremamente importante l'azione svolta dal virus su particolari cellule accessorie (dendritiche o interdigitate), probabilmente già invase e danneggiate dall'HIV prima del coinvolgimento della popolazione linfocitaria. Studi compiuti con l'impiego di anticorpi monoclonali hanno evidenziato la presenza della proteina p24 (appartenente alla struttura più interna del virus) nelle cellule del centro germinativo (v. Baroni e altri, 1986).
e) Diagnosi di laboratorio
La diagnosi di infezione da HIV e delle relative forme cliniche (LAS-ARC) e di AIDS è oggi sufficientemente codificata, grazie anche alla possibilità di identificare gli anticorpi anti-HIV (v. Brun-Vézinet e altri, 1984). I progressi in campo laboratoristico sono stati molti nel corso degli ultimi tre anni: la caratterizzazione degli antigeni proteici del virus e la stessa individuazione del genoma delle proteine hanno consentito di perfezionare metodiche atte al rilevamento degli anticorpi specifici. Attualmente i metodi applicati per svelare gli anticorpi anti-HIV sono i seguenti: ELISA (enzyme-linked immunosorbent assay, test enzimatico) con miscela di vari antigeni, ELISA con le proteine note p41 e p24, RIA (radio immunological assay, test radioimmunologico) e immunofluorescenza, immunocompetizione e Western blot. I maggiori problemi sono emersi con l'impiego dell'ELISA e dell'immunofluorescenza. Per questo motivo sono stati successivamente impiegati dei test detti di conferma. Il test di conferma più applicato è il Western blot, che impiega una serie di procedimenti analitici in grado di rilevare la reattività anticorpale verso le singole componenti del complesso proteico degli antigeni virali. Senza analizzare ulteriori dettagli è però assai importante sottolineare il ruolo delle varie componenti proteiche virali per la definizione dei criteri di positività o negatività. In genere il criterio di positività di un siero è dato dalla presenza di due o più bande per una proteina virusspecifica (p15-18, p24, p31-33, gp41, gp120). In particolare se una banda correlata al gene GAG si associa a una banda riferita a quello ENV, il criterio di positività è ancora più chiaro. L'esperienza consiglia di ripetere il saggio se originariamente la positività riguarda una sola banda. In alcuni casi (in particolare quando si sospetta un'infezione recente o nel caso di bambini che possono essersi infettati durante la vita fetale) è utile senz'altro valutare la presenza di anticorpi di tipo IgM, sebbene difficoltà anamnestiche e tecniche possano non rendere di completo affidamento quest'ultimo approccio. È importante, inoltre, il monitoraggio dei pazienti, la possibilità di eseguire colture batteriche particolari e l'identificazione dei vari agenti patogeni (per esempio funghi). Il problema dei sieropositivi (asintomatici) è quello di stabilire l'appartenenza alle varie categorie di rischio ed eventualmente di identificare altri parametri di inquadramento diagnostico e/o prognostico (v. Aiuti e altri, 1985). Sono importanti: globuli bianchi e formula leucocitaria (con particolare riguardo alla leucocitopenia e/o linfocitopenia: significato del numero assoluto rispetto alla percentuale semplice), conteggio delle piastrine (anche se non ancora completamente codificato, un graduale ma sensibile e costante decremento del numero delle piastrine circolanti può essere un segno diagnostico di evoluzione, assieme alla splenomegalia), dosaggio delle immunoglobuline sieriche (aumento delle IgG e IgA e comunque incremento totale superiore al 2o-3o%). In caso di soggetti con sospetto di falsa positività (oggi rischio assai limitato) è bene eseguire il Western blot. La possibilità di isolare direttamente il virus è un altro aspetto del problema diagnostico-epidemiologico che non può essere ancora affrontato in sede routinaria. Non va dimenticato, comunque, sia in fase diagnostica che nel monitoraggio del paziente, l'insieme dei problemi neurologici connessi all'infezione da HIV. Si tratta di una patologia variabile (encefalite, neuropatie periferiche, forme demielinizzanti) che può essere completamente svincolata dal danno strettamente linfocitario e presentarsi con una relativa precocità.
f) Le classificazioni e l'inquadramento nosologico
Il gruppo dei soggetti sieropositivi va distinto in due categorie: gli asintomatici e i sintomatici. Gli asintomatici (impropriamente definiti portatori sani) presentano nella maggioranza dei casi alterazioni immunologiche rilevabili durante l'esecuzione di analisi di laboratorio. La sieroconversione dopo il contagio presenta periodi di tempo variabili e si può verificare anche a distanza di 6-8 mesi, mentre, secondo le informazioni più recenti, il periodo medio di incubazione intercorrente dalla diagnosi di positività alla manifestazione clinica di AIDS conclamata sarebbe più lungo. Attualmente vengono proposti tre tipi di classificazione, che consentono di inquadrare i pazienti secondo criteri clinico-immunologici.
Nella pratica clinica si fa spesso ricorso alla definizione di LAS (sindrome linfoadenopatica) o ARC (AIDS related complex), definizioni non generiche bensì derivanti da precise codificazioni. Nella tab. II è riportato lo spettro delle descrizioni note per la patologia correlata all'HIV.

Uno schema che tiene conto della situazione immunologica, con particolare riferimento al numero dei linfociti T-helper circolanti, è quello ottenuto dalla classificazione del Walter Reed Institute (v. Redfield e altri, 1986). Si tratta di una classificazione in stadi (da WRO a WR6) che sintetizza una serie di situazioni cliniche caratterizzate da assenza di infezioni opportunistiche. È abbastanza impiegata in clinica su grandi numeri, ma non sembra essere l'ideale per impostare un adeguato monitoraggio clinico, non consentendo una precisa descrizione del singolo caso (con tutte le specifiche caratteristiche). Molto utile è la più recente classificazione proposta nel maggio 1986 dal CDC (Center for Diseases Control) di Atlanta (v. Classification..., 1986). La diagnosi specifica di infezione si basa sull'identificazione diretta del virus nei tessuti del paziente o su risultati di test ELISA ripetutamente positivi, nei quali gli anticorpi sono anche identificati con l'impiego di metodi diversi (immunofluorescenza, Western blot). Il sistema di classificazione si applica ai pazienti con diagnosi di infezione da HIV e ha essenzialmente uno scopo diagnostico-descrittivo (v. tab. III).

g) Le conseguenze cliniche dell'infezione da HIV
Dopo il contagio iniziale appaiono gli anticorpi rivolti verso le proteine del virus. Questa fase di sieroconversione consente di porre la diagnosi di soggetto infetto ma non sempre si accompagna a sintomatologia clinica. Gli anticorpi prodotti inizialmente non sono dotati della capacità di neutralizzare il virus e quindi non svolgono un ruolo protettivo. In alcuni soggetti è stata descritta l'infezione acuta, caratterizzata da malessere generalizzato simile a un quadro influenzale, con febbre, profonda astenia, dolori muscolari, faringite, rash cutaneo e discreta linfoadenopatia diffusa alle principali stazioni linfonodali. Questo tipo di reazione è anche definita mononucleosis-like syndrome, per la sua somiglianza con la forma acuta di mononucleosi.
Dopo il periodo iniziale, sintomatico o asintomatico, si sviluppa la cosiddetta fase cronica. Questa può essere caratterizzata da un periodo silente sotto il profilo sintomatologico o progredire in breve verso la sindrome linfoadenopatica. Va comunque precisato che in quest'area di ‛transizione clinica', non ancora completamente inquadrata, sono presenti, in un gran numero di individui, ben definiti danni immunologici (v. Aiuti, 1983 e 1984; v. Aiuti e altri, Nuovi aspetti... e AIDS in Europe..., 1984). Questa forma può restare inalterata per lungo tempo, anche per anni, e addirittura regredire, tanto da far considerare il paziente impropriamente asintomatico, oppure può evolvere in peggioramento verso l'ARC, il cui quadro è stato già descritto in dettaglio. Giova comunque qui ripetere che questa sindrome si caratterizza sul piano clinico per la presenza di segni generali accompagnati da sintomi specifici: si ricordano le manifestazioni oculari, la candidiasi orale, i condilomi giganti, il mollusco contagioso, alcune infezioni cutanee da funghi, le encefaliti acute o croniche (quadro sommariamente definibile anche come pre-AIDS o lesser AIDS). Questa forma di ARC ha un'alta probabilità di progressione ulteriore in funzione della gravità e dell'estensione del danno immunitario. L'AIDS (forma conclamata) è la conseguenza più severa della storia naturale dell'infezione. La patologia che la caratterizza comprende tre elementi base: la presenza di infezioni opportunistiche, il sarcoma di Kaposi o il linfoma cerebrale, l'associazione tra infezioni opportunistiche e tumori. L'evoluzione dell'AIDS (invariabilmente fatale in circa il 90% dei soggetti entro 4 anni) dipende dalla progressione del danno cellulare (riduzione marcata dei linfociti CD4+) con riduzione delle risposte cellulo-mediate, attivazione policlonale del sistema B e iperproduzione di gammaglobuline (v. Zolla-Pazner, 1984; v. Luzi e altri, Comunicazione..., 1986). Il danno complessivo che subisce un paziente con AIDS deriva da due ordini di fattori: 1) l'immunodeficienza generalizzata; 2) le lesioni specifiche causate dall'HIV in tessuti diversi da quello linfatico. Il coinvolgimento neurologico rappresenta una tappa molto importante, che può condizionare il quadro clinico complessivo in circa 1/3 dei pazienti: si assiste a una grave forma di deterioramento, con cefalea, depressione, demenza progressiva e neuropatia periferica. In un primo momento si riteneva che le alterazioni a carico del sistema nervoso centrale derivassero da infezioni opportunistiche, ma oggi è chiaro che il virus stesso attacca direttamente le cellule cerebrali ed è responsabile delle sequele acute, subacute e croniche. La sindrome di riscontro più frequente è l'encefalite subacuta. È possibile, ed esistono alcune ipotesi al riguardo, che il danno neurologico sia del tutto indipendente dal linfotropismo del virus e che molti dei ‛portatori asintomatici' progrediscano lentamente verso una forma di interessamento del sistema nervoso clinicamente evidenziabile solo dopo molti anni (v. Snider e altri, 1984; v. Levy e altri, 1985).
h) I problemi epidemiologici
Il virus HIV si trasmette per via ematica e sessuale (v. Luzi e altri, 1985); la sua diffusione è mondiale e l'Organizzazione Mondiale della Sanità ha dichiarato lo stato di pandemia, poiché sono stati osservati 58.880 casi al 2 settembre 1987. I gruppi a rischio maggiore sono i tossicodipendenti e gli omosessuali, i figli di madre sieropositiva e i politrasfusi. Al 30 maggio 1987 i casi segnalati di AIDS conclamato in Europa erano 4.850. I dati sull'Africa sono estremamente difficili da valutare per le difficoltà di raccolta epidemiologica, sebbene alcune proiezioni statistiche indichino circa 10 milioni di sieropositivi. La distribuzione per milione di abitanti segnala un indice di 25,7 in Danimarca, 29,5 in Svizzera, 22,2 in Francia, 10,8 nel Regno Unito, 9,1 in Italia, 6,9 in Spagna. Negli Stati Uniti i casi segnalati sono circa 42.000 e i decessi superiori al 50%. Un recente convegno a Roma su AIDS e sindromi correlate (19-21/5/1987) ha consentito di stabilire, su un'imponente casistica di 52.900 soggetti, che circa la metà dei sieropositivi presenta sintomi diagnosticabili (6.834 LAS e 1.749 ARC) e che quindi in Italia gran parte dei portatori di anticorpi anti-HIV sono in fase sintomatologica al momento della diagnosi. I tossicodipendenti hanno una distribuzione media di sieropositività attorno al 47%. Negli omo-bisessuali la frequenza è del 25,6%. Si sono riscontrate alte quote di sieropositivi fra gli emofilici (32,6%), i politrasfusi adulti (5,6%) e bambini (6,1%). Il 50% dei figli di madre infetta nasce sieropositivo. Benché si siano accumulate molte informazioni sull'incidenza dell'AIDS, è assai difficile elaborare previsioni e stime attendibili sia sul numero dei sieropositivi sia sul rischio che i sieropositivi corrono di evolvere verso la forma clinica conclamata. È anche difficile, sotto il profilo statistico, ottenere parametri epidemiologici sufficientemente rigidi per il calcolo previsionale. L'incidenza dell'AIDS dipende infatti non soltanto dalla trasmissione (agente causale e cofattori), ma anche dalla durata del periodo di incubazione (ad altissima variabilità) e dalla frazione degli individui a maggior rischio (come sembrano indicare alcune recenti osservazioni sul significato genetico della risposta antivirale all'HIV). Un dato costante sembra l'irreversibilità della lesione immunologica, sebbene il tempo di evoluzione che definisce la soglia del danno non sia completamente noto. I dati più completi riguardano le osservazioni su numerosissimi omosessuali. È probabile che alcuni fattori ‛nascosti' non ancora noti condizionino l'estensione dell'infezione in particolari gruppi di rischio e in determinate aree geografiche. Il problema attualmente presenta due aspetti principali: 1) il rischio ‛cronologico' del singolo individuo portatore del virus nell'evoluzione verso l'AIDS; 2) il rischio di diffusione nella popolazione eterosessuale.
Si è scritto molto negli ultimi anni sull'AIDS e sulle vane modalità di prevenzione che possono essere adottate. È evidente che si tratta di un grande problema nazionale e internazionale che può essere risolto nel tempo con l'adeguato impegno di consistenti risorse finanziarie, agendo sia a livello scientifico-clinico che a livello profilattico-educativo. In particolare, una costante sorveglianza va attuata sia nei centri trasfusionali che a livello diagnostico-laboratoristico. È necessario affinare i metodi sotto il profilo sia della specificità che della sensibilità, per evitare di incorrere in errori sull'attribuzione della sieropositività.
i) La prevenzione
Le modalità di trasmissione del virus che causa l'AIDS, sovrapponibili a quelle del virus dell'epatite B, rendono essenziale un'adeguata conoscenza (da parte della popolazione in genere e di alcune categorie di operatori in particolare) dei rischi del contagio e dei mezzi per prevenirlo. L'HIV può essere trasmesso solo per via sessuale e parenterale: quindi attraverso il rapporto sessuale con persone infette, mediante trasfusione di sangue infetto (plasma o fattori della coagulazione), con l'uso di siringhe e di aghi contaminati con sangue infetto. Non è stata sinora messa in chiara evidenza alcuna trasmissione dell'infezione da virus HIV attraverso contatti occasionali in ambienti di lavoro, sui mezzi pubblici, attraverso la via oro-fecale. È in discussione il ruolo di alcuni insetti ematofagi, ma allo stato attuale delle conoscenze, almeno alle nostre latitudini, non sembra che questo rischio sussista. In attesa di farmaci efficaci o della messa a punto di un vaccino, vanno condotte, a scopo preventivo, campagne di informazione su larga scala, peraltro molto difficili da attuare, perché debbono indurre a modificare alcune abitudini sociali; ne deriva quindi l'esigenza di organizzare un sistema adeguato di rilevamento dati al fine di sapere se l'intervento educativo abbia o meno conseguito un reale successo.
l) La terapia
Il problema del controllo farmacologico delle infezioni causate da particelle virali è ancora aperto. La stessa ‛semplicità' del virus è il fattore che ha vanificato i vari approcci tentati sinora. Il nocciolo del problema consiste nella distinzione fra le funzioni virus-correlate e quelle proprie della cellula infettata. Per questo motivo si è ricorso a un controllo mediato delle infezioni da virus e sono stati introdotti, in varie epoche e con modalità differenti, i vaccini.
Soltanto i recenti progressi della genetica virale e l'adozione delle biotecnologie hanno modificato in parte i termini del problema, promuovendo il ricorso all'impiego di alcuni prodotti che svolgono un'azione antivirale diretta (v. Broder, 1987). Tra i prodotti più noti si ricordano l'azidotimidina, la didesossicitidina, l'acyclovir, la ribavirina e la vidarabina: si tratta dei cosiddetti analoghi dei nucleosidi. Come è noto i nucleosidi, mediante azione enzimatica, possono essere assemblati a formare catene di acidi nucleici. Se un analogo del nucleoside viene a interporsi durante la fase di sintesi, si ha un danno strutturale con il blocco della replicazione del virus. Nell'ambito dei Retrovirus e dell'HIV in particolare è stato proposto di recente l'impiego dell'AZT (azidotimidina). Questa molecola è nota sin dagli anni sessanta e fa parte dei prodotti elaborati per le terapie antitumorali. È un potente inibitore della transcriptasi inversa e svolge il suo ruolo bloccando la crescita della catena di DNA (chain terminator). L'affinità dell'AZT per la transcriptasi inversa è molto alta rispetto all'affinità dimostrata per la DNA-polimerasi: la molecola, poiché a livello del pentosio ha un N anziché un gruppo idrossilico, non trova punti di ancoraggio e la transcriptasi non può funzionare. Gli studi in vitro dimostrano una potente azione del farmaco sulla replicazione virale, senza che il danno si estenda alla popolazione dei linfociti T. Attualmente sono in corso negli Stati Uniti e in Europa diversi protocolli clinici di applicazione dell'AZT in pazienti con AIDS e sindromi correlate. Malgrado gli attuali dati sembrino incoraggiare l'impiego del prodotto a breve termine, non vanno dimenticati i rischi di una terapia protratta per lungo tempo, soprattutto a causa della potenziale tossicità della molecola nei confronti del midollo osseo. Altri approcci miranti a bloccare la replicazione dell'HIV si stanno attuando con l'interferon alfa umano, che può inibire la produzione di RNA virale e limitare o abolire la sintesi di proteine e il loro assemblaggio. Per il futuro sono previsti trials con farmaci sinergici (ad azione sia antivirale che immunopotenziante) che, producendo effetti tossici ridotti perché somministrati a dosi relativamente basse, avrebbero l'indubbio vantaggio di colpire il virus in diverse fasi della sua replicazione. Resta ovviamente aperta la problematica del vaccino: purtroppo l'alta variabilità genetica dell'HIV e il conseguente polimorfismo fenotipico non hanno consentito finora risolutivi progressi nel settore. Il tempo di attesa è comunque stimabile in almeno tre-cinque anni.
Negli ultimi due anni sono stati approntati tre tipi di vaccino: il primo ottenuto uccidendo il virus con formaldeide o al calore, il secondo utilizzando la metodica del DNA ricombinante e preparando pertanto antigeni dell'ENV (parti costanti) e/o del core, il terzo con un sistema di anticorpi anti-idiotipo. Il vaccino anti-idiotipo, pur essendo promettente, per ora non sembra in grado di fornire immunità altamente protettiva. Esso è stato ottenuto inoculando animali con anticorpo monoclonale anti-CD4+ (cioè contro il recettore per l'HIV) e quindi in grado di riconoscere anche il virus stesso per la complementarità della conformazione sterica.
Gli esperimenti compiuti con i tre vaccini su animali correlati all'uomo (scimpanzé) non hanno fornito una risposta immunitaria capace di protezione. I dodici casi umani vaccinati hanno manifestato una risposta immunitaria dopo circa due mesi, ma non si conosce ancora l'eventuale ruolo protettivo (Zaguri, comunicazione al III Congresso internazionale sull'AIDS, Washington, 1-6 giugno 1987).
4. Ruolo della risposta immunitaria nel controllo della crescita neoplastica
Durante gli anni settanta la teoria della sorveglianza immunologica nel controllo della crescita neoplastica ha rappresentato un entusiasmante modello di studio, fondato sul concetto della potenziale efficacia difensiva da parte del sistema immunitario nei confronti della cellula tumorale. In realtà l'esistenza di una risposta immunitaria non significa necessariamente risposta efficace; infatti gli antigeni neoplastici sono deboli immunogeni non in grado di stimolare adeguatamente una risposta valida nei propri confronti. Nello stesso tempo si può sostenere che appaiono ‛clinicamente' soltanto i tumori che sono riusciti a svincolarsi dalla sorveglianza immunitaria. Il problema è aperto e allo stato attuale delle conoscenze esistono varie situazioni cliniche o sperimentali che possono aiutarci a inquadrare questo aspetto del sistema tumore-immunosorveglianza, sebbene una risposta univoca e convincente sembri piuttosto lontana.
Ai fini di una discussione analitica riteniamo opportuno isolare alcuni concetti base che sono senz'altro di aiuto per la comprensione del problema. Dobbiamo a tal fine prendere in esame: a) la cellula tumorale; b) le modalità con le quali il sistema immunitario può contenere l'espansione neoplastica; c) i meccanismi attraverso i quali la cellula tumorale elude la sorveglianza immunologica.
a) La cellula tumorale
È ancora difficile fornire una definizione ‛assoluta' di cellula tumorale maligna; è certo, comunque, che il carattere peculiare è la proliferazione incontrollata, con la capacità di colonizzare a distanza dalla sede di origine. In linea di massima il ritmo di divisione di una cellula neoplastica non è molto diverso da quello di una cellula normale, tuttavia nel tessuto neoplastico si osserva un'incapacità di differenziamento insieme alla presenza di numerose unità in ciclo mitotico (cioè attivamente proliferanti). Grazie alla capacità di produrre sostanze chimiche (come i fattori di crescita angiogenetici), la massa neoplastica invece di andare incontro a necrosi si organizza (almeno temporaneamente) e dà luogo alla costituzione di una neovascolarizzazione di tutto il tessuto. In un sistema tissutale normale il contatto tra le cellule ne blocca la proliferazione (inibizione da contatto). Tale regolazione può essere vista come un preciso meccanismo di controllo, geneticamente determinato, ai fini di una specializzazione funzionale. Le cellule maligne perdono l'inibizione da contatto e alcune di loro sono in grado di migrare e proliferare in distretti corporei diversi da quello di origine (metastatizzazione). La trasformazione neoplastica e il processo di cancerogenesi non sono ancora completamente noti; ne sono state prese in esame molte cause: mutazioni somatiche, radiazioni, cancerogeni chimici, alterati meccanismi di riparazione del DNA. Di recente è stata dimostrata l'esistenza, sia nei virus sia nelle cellule, di geni in grado di conferire caratteristiche neoplastiche alla cellula nella quale si trovano localizzati. In particolare nelle cellule eucariotiche tali geni (oncogeni) sono presenti normalmente; una loro alterazione o trascrizione anomala si accompagna a quel complesso di trasformazioni fenotipiche in grado di dare origine alla crescita neoplastica. Molte conoscenze al riguardo sono derivate dallo studio di virus oncogeni a RNA (Oncornavirus), noti anche come Retrovirus. Comunque, se prescindiamo dal primum movens, i tumori hanno origine dalla trasformazione di una sola cellula che, proliferando, dà luogo all'espansione di una popolazione clonale. Accanto a questo fenomeno si deve considerare la serie di eventi trasformanti che precedono e che accompagnano l'espansione neoplastica: la cosiddetta crescita a tappe multiple. Con il tempo quindi tante popolazioni di cellule maligne finiscono con il costituire una massa neoplastica in situ o ‛colonizzante'. In pratica il clone cellulare più maligno finirà con l'assumere un ruolo dominante sugli altri e determinerà la cachessia dell'organismo ospite. Per arricchire il quadro delle informazioni acquisite vanno citate tutte le sostanze (o fattori) che, elaborate nel microambiente dal tumore stesso, ne condizionano in parte l'evoluzione rispetto ai sistemi di controllo. (V. anche neoplasie: Oncologia sperimentale; v. trasformazione delle cellule; v. virus; v. neoplasie).
b) Modalità con le quali il sistema immunitario può contenere l'espansione neoplastica
Indipendentemente dal modello di crescita neoplastica preso in considerazione, il sistema immunitario viene attivato da strutture antigeniche in grado di stimolare efficacemente le cellule immunocompetenti (immunogenicità). Di conseguenza un campo di ricerca essenziale è quello degli antigeni tumorali o tumore-associati (v. Coggin e Anderson, 1974; v. Bartorelli, 1980). Il progresso delle conoscenze in tale settore deriva dagli studi sui trapianti di cellule neoplastiche in animali geneticamente puri e dalla caratterizzazione delle proprietà immunochimiche delle molecole antigeniche. Il primo approccio riguarda quindi lo studio degli antigeni tumorali specifici e la risposta immunitaria da essi indotta. Va innanzi tutto precisato che è assai arduo trasferire sull'uomo quanto ricavabile dalla sperimentazione in vitro o su animali. A livello della membrana di una cellula neoplastica si trovano varie strutture antigeniche: antigeni di differenziazione, embrionali, ‛specifici' della stessa cellula tumorale e altri. È bene distinguere gli antigeni tumorali specifici (non presenti sulle cellule normali del tessuto d'origine della neoplasia) dagli antigeni tumoreassociati (non strettamente peculiari di un particolare tumore noto). Gli antigeni tumore-associati in condizioni sperimentali sono in grado di indurre risposte immunitarie che provocano il rigetto del tumore, ma la risposta espressa nei loro confronti non svolge sempre un effetto di controllo sulla crescita dello stesso tumore. Per alcuni tumori indotti chimicamente sono stati identificati antigeni specifici correlati al tipo di carcinogeno impiegato, così come gli studi svolti con virus oncogeni consentono l'identificazione di strutture molecolari correlate alle caratteristiche dei virus analizzati negli esperimenti di carcinogenesi. Comunque antigeni presenti sulla superficie cellulare di tessuti normali dell'adulto sono stati trovati anche su cellule neoplastiche. Esistono senz'altro problemi di redistribuzione quantitativa degli antigeni ‛normali' su cellule neoplastiche, ma mancano sino a oggi delle conoscenze significative su questo argomento. Lo sviluppo delle conoscenze sui ‛marcatori tumorali' ha consentito di mettere a punto adeguate tecniche di laboratorio utilizzabili in diagnostica clinica. Per esempio la ricerca dell'alfa-fetoproteina e quella dell'antigene carcinoembrionario (CEA) sono utili in alcune neoplasie specifiche; l'alfa-fetoproteina viene riscontrata in corso di epatocarcinoma e di alcuni teratomi maligni del testicolo, e la presenza di CEA consente di evidenziare neoplasie maligne a carico dell'apparato digerente. Qualche elemento utile è fornito da un altro antigene sulfoglicoproteico, che è stato evidenziato nel liquido di lavaggio gastrico in pazienti con tumori maligni dello stomaco. Gli antigeni oncofetali corrispondono sostanzialmente a una fase di derepressione genica che consente l'espressione fenotipica di molecole usualmente identificabili soltanto durante la vita embrionale. La teoria della sorveglianza immunologica parte dal presupposto che durante la divisione cellulare si originino doni mutanti e che una parte della risposta immunitaria si sia evoluta per eliminare le cellule potenzialmente neoplastiche. Esistono indirizzi sperimentali e osservazioni cliniche favorevoli alla teoria della sorveglianza immunologica, ma esistono anche dati non coerenti con l'impostazione generale del problema. Per esempio è noto che quando l'immunocompetenza è relativamente diminuita (età, trattamenti immunodepressivi) si ha una maggiore suscettibilità alla carcinogenesi negli animali da esperimento, ed è noto che la terapia adottata per contenere il rigetto di trapianti induce un maggior rischio di crescita neoplastica in alcuni distretti (labbra, pelle). Non solo, ma è esperienza clinica l'osservazione che il controllo delle metastasi (e quindi la loro regressione) è talvolta secondario all'asportazione chirurgica del tumore principale. Approfondiremo questi concetti nel paragrafo prossimo: in questo limitiamo l'analisi al complesso dei fattori immunologici in grado di inibire la crescita tumorale. Mentre il ruolo degli anticorpi può essere di una qualche rilevanza biologica in certe fasi della crescita neoplastica (v. Klein e Klein, 1985), gli studi in vitro hanno consentito di focalizzare tre tipi di citotossicità mediata da cellule: citotossicità diretta mediata dai linfociti, citotossicità anticorpo-mediata e citotossicità legata alle funzioni macrofagiche. Recentemente è stato descritto un tipo cellulare che presenta alcune funzioni caratteristiche: la cellula NK (natural killer: v. Trinchieri e Perussia, 1984). Queste cellule, identificate in ceppi di topi privi di pelo e congenitamente atimici (nude mice), sono morfologicamente descritte come LGL (large granular lymphocytes) e svolgono un ruolo primario nei confronti delle cellule tumorali, senza una precedente immunizzazione specifica nei loro confronti.
c) Meccanismi attraverso i quali la cellula tumorale elude la sorveglianza immunologica
Dal momento che gran parte dei pazienti colpiti da una neoplasia non presenta gravi danni a carico del sistema immunitario e dal momento che una risposta immunologica verso gli antigeni neoplastici non sempre significa risposta efficace contro la cellula neoplastica, l'intera teoria della sorveglianza immunologica è stata sottoposta a revisione critica. Non solo, ma il modello offerto dagli ‛esperimenti della natura', le immunodeficienze primitive, è esso stesso non privo di contraddizioni. Infatti il tumore associato ai difetti immunologici primitivi è di tipo sistemico e prevalentemente a carico della serie linfocitaria, mentre soltanto in alcuni deficit vengono segnalati tumori di altri tessuti. Quindi è necessario usare molta prudenza nell'interpretare i dati, per attribuire loro il giusto significato. È indubbio infatti che la cellula neoplastica presenti dei vantaggi ‛selettivi' nei confronti del sistema di sorveglianza: essa è dotata di antigeni deboli e quindi suscita una risposta lenta e debole. Un procedimento di immunoselezione potrebbe consentire la proliferazione di particolari doni che, cresciuti ‛in sordina', finirebbero con attuare uno sneak through e deviare la risposta anticorpo-mediata dell'organismo ospite. Non solo, ma spesso la membrana della cellula tumorale libera frazioni di antigene o antigeni tumorali dando luogo al cosiddetto shedding (v. Alexander, 1974). La risposta anticorpale indurrebbe di conseguenza la formazione di complessi immuni e le macromolecole in tal modo costituite finirebbero con l'esercitare una funzione di blocco sul sistema efferente-citotossico cellulo-mediato. Altre situazioni ‛di fuga' potrebbero associarsi alla sede topografica e alla vascolarizzazione: infatti se il processo di espansione neoplastica non consente un rapido utilizzo di vasi neoformati, la risposta cellulare attraverso il circolo risulta altamente compromessa e le modalità di sensibilizzazione indurrebbero (grazie anche alla frequente bassa immunogenicità) un fenomeno di tolleranza. È anche osservabile un fenomeno di amplificazione della crescita tumorale causata da particolari circostanze di ‛facilitazione' (enhancement), riconducibili alla presenza di immunocomplessi circolanti (con funzione di schermo rispetto alle cellule effettrici) o a un'azione ‛diretta' di anticorpi che modulano sulla stessa membrana della cellula neoplastica l'espressione di particolari antigeni (v. Prehn, 1972). L'interazione tra anticorpo e antigene tumorale comporterebbe l'insorgere di un segnale che stimolerebbe l'espansione proprio della cellula tumorale.
5. Il concetto di monitoraggio immunologico
La disponibilità di varie tecniche di laboratorio e il rapido evolvere delle conoscenze immunologiche nelle loro diverse ramificazioni rendono assai complesso il significato dei numerosi contributi. Del resto è arduo definire in termini netti l'area di competenza di ciascuna sottodisciplina immunologica, dovendosi ogni volta premettere quale tipo di ‛conoscenza' si vuole raggiungere o utilizzare. È comunque indubbio che l'immunologia (come immunologia generale, immunochimica, immunogenetica, immunologia clinica) ha modificato gran parte dell'interpretazione della patogenesi di numerose malattie, consentendo nello stesso tempo di chiarire le modalità di risposta dell'organismo sano nei confronti dei vari patogeni noti. Ma risposta immunitaria significa anche equilibrio omeostatico, significa regolazione e modulazione. Se teniamo conto di questa breve premessa si comprendono meglio le difficoltà che l'immunologo clinico trova sia nella diagnostica delle malattie che hanno una precisa genesi immunologica sia nell'ambito della medicina interna, che si avvale delle tecniche diagnostiche dell'immunologia e delle sue conoscenze specifiche (basti pensare alle epatopatie croniche, alle malattie delle ghiandole endocrine, ad alcune forme di infertilità, alle malattie allergiche). Si intuisce inoltre che i dati di laboratorio e quelli derivati dalla sperimentazione in vitro non possono avere un immediato riscontro applicativo e quindi il concetto di ‛monitoraggio immunologico' va limitato a quell'insieme di tecniche e metodologie che, allo stato attuale delle conoscenze, contribuiscono alla formulazione di una diagnosi e alla valutazione del coinvolgimento del sistema immunitario durante l'evolvere di ‛altra' patologia a eziologia non immunitaria.
Le attuali metodiche del monitoraggio immunologico comprendono le tecniche di laboratorio relative allo studio dell'immunità cellulare, le indagini sulla risposta antigene-anticorpo e l'insieme delle nuove tecnologie relative all'‛immunità umorale'.
Le tecniche di laboratorio di uso più comune nel settore degli studi sull'immunità umorale sono le seguenti: elettroforesi; immunoelettroforesi; immunodiffusione; vari metodi immunochimici e fisico-chimici; radioimmunologia; alcune applicazioni dell'immunofluorescenza; cromatografia; varie metodiche di agglutinazione; analisi delle funzioni complementari; applicazione di anticorpi monoclonali.
Nel settore degli studi sulla risposta cellulare si possono citare alcune metodiche: numero e funzioni dei linfociti (stimolazione in vitro); coltura mista linfocitaria; tecniche varie di rosetting (rosettazione); alcune applicazioni dell'immunofluorescenza; studio dell'immunità specifica (funzioni analizzate: test di degranulazione, killing intracellulare, chemiluminescenza, chemiotassi e fagocitosi).
L'analisi dettagliata di tutte le metodiche elencate esula dall'ambito di questo articolo, tuttavia sembra opportuno soffermarsi sui principi di alcune delle tecniche sopracitate che hanno contribuito grandemente alla diagnostica di laboratorio. Per esempio, combinando la separazione elettroforetica con l'immunoprecipitazione, si possono individuare le componenti sieriche e scoprire eventuali alterazioni in senso monoclonale (per esempio nei mielomi), così come il dosaggio per immunodiffusione radiale secondo Mancini è oggi applicato routinariamente per ‛pesare' diverse componenti plasmatiche. In base alla relazione esistente tra la concentrazione dell'antigene posto in un pozzetto scavato in agar-gel e il quantitativo di anticorpo diffuso nel supporto gelificato, e in base al diametro dell'anello di migrazione, si ricava il valore della concentrazione dell'antigene (questa tecnica è usata in molti laboratori per il dosaggio delle immunoglobuline sieriche). In alcuni casi è possibile applicare l'immunodiffusione doppia di Ouchterlony, per valutare reazioni di identità e non identità tra anticorpo e antigene. Per il dosaggio di quantità molto piccole si usa oggi la radioimmunologia (per esempio per il monitoraggio degli ormoni tiroidei, o per riconoscere gli anticorpi IgE responsabili di alcune sindromi allergiche, o per identificare l'allergene responsabile di una particolare sensibilizzazione). Di solito in radioimmunologia si usa un anticorpo marcato con tracciante radioattivo e, con opportuni sistemi chimico-fisici (per esempio di inibizione del legame tra antigene e anticorpo), si può dosare la molecola ricercata. Talvolta, però, le metodiche di base vengono leggermente modificate o sensibilizzate per una ottimizzazione del rendimento finale (questi procedimenti sono prerogativa frequente dell'industria dei diagnostici che ha avuto una grandissima espansione, soprattutto nel settore dell'indagine allergologica). A introdurre i RIA (radio immunological assays) furono, all'inizio degli anni sessanta, Berson e Yalow che si occuparono essenzialmente del dosaggio dell'insulina plasmatica. Da allora i progressi sono stati molteplici e oggi tale metodica si distingue per la sua peculiare alta sensibilità e l'ottima specificità (quest'ultima dipende ovviamente anche da altri fattori). L'immunofluorescenza è essenzialmente una tecnica citochimica o istochimica, che si basa su un anticorpo rivelatore marcato con fluorocromo (isotiocianato di fluorescina o rodamina). Dopo la reazione tra substrato antigenico e anticorpo fluorescente si forma un immunocomplesso stabile che può essere facilmente identificato in campo oscuro ricorrendo a una sorgente di luce ultravioletta. Il suo significato in sierologia è importante poiché questa tecnica permette l'identificazione indiretta di anticorpi. In pratica si può considerare una derivazione della reazione di Coombs. Esiste anche un sistema di immunofluorescenza ‛quantitativa': usando un microfluorimetro opportunamente tarato è possibile convertire la quantità di luce emessa in funzione del peso e calcolare la concentrazione dell'anticorpo. Tale metodica ovviamente può essere adattata a diversi sistemi, ma viene in pratica utilizzata per misurare le immunoglobuline sieriche, frazioni del complemento e anticorpi antinucleo. Un ulteriore progresso è stato reso possibile dall'impiego dei metodi enzimatici, che utilizzano la coniugazione di una molecola sull'anticorpo rivolto verso un particolare antigene. Poiché l'insieme anticorpo-enzima è funzionante sia sotto il profilo immunologico che per quanto riguarda la reattività enzimatica, in pratica si applica un principio del tutto sovrapponibile a quello dell'immunofluorescenza. Dopo l'avvenuta reazione antigene-anticorpo si aggiunge al sistema il substrato per l'enzima (che, per esempio, può essere una perossidasi) e quindi con una lettura adeguata si evidenzia il tipo di risposta cercata: qualitativa e/o quantitativa. Le due varianti di questa metodica che hanno maggiore applicazione sono la competitiva e la più nota ELISA (enzyme-linked immunosorbent assay). I test enzimatici trovano largo impiego nella diagnostica clinica e sono attualmente applicati per la determinazione quantitativa di numerose sostanze (ormoni, anticorpi, molecole farmacologiche). Una recente estensione si è avuta nella ricerca di anticorpi rivolti contro antigeni associati al virus che provoca l'AIDS (HIV).
Una metodica di purificazione, estremamente importante per il frazionamento delle proteine e l'isolamento di diversi tipi di immunoglobuline anticorpali, è la cromatografia. Esistono due principi generali di applicazione di tale metodica: lo scambio ionico e la cosiddetta ‛gel-filtrazione'. In apposite colonne si dispongono molecole cariche elettricamente che formano una matrice di separazione (scambio ionico). Le sostanze che debbono essere separate (di carica opposta o elettricamente neutre) vengono eluite con soluzioni a pH opportuni. Nel sistema con gel-filtrazione le molecole vengono separate tenendo conto della loro grandezza e forma, e scorrono ‛attraverso' biglie che formano la matrice della colonna. Ottimizzando questi procedimenti si possono conseguire notevoli risultati analitici utilizzabili in particolari ricerche di diagnostica clinica.
La moderna indagine nel settore sierologico utilizza in modo sempre più vasto gli anticorpi monoclonali. Nel 1975 Kohler e Milstein hanno introdotto la tecnica di preparazione degli anticorpi monoclonali, che consiste essenzialmente nell'ibridazione somatica di cellule in grado di formare anticorpi con cellule in attiva replicazione (v. biotecnologie, suppl.). Con tale procedimento e opportune metodiche di purificazione gli immunologi sono in grado di produrre oggi una quantità illimitata di molecole con caratteristiche fisico-chimiche di assoluta omogeneità. Gli ibridomi o ibridi di cellule somatiche si possono ottenere fondendo una sospensione di cellule spleniche o linfociti ottenuti da topi o ratti immunizzati con cellule neoplastiche in continua replicazione (per esempio di linfomi o mielomi). La scelta delle linee cellulari in replicazione si basa su: 1) assenza di produzione o secrezione di anticorpi; 2) assenza di un enzima molto importante noto come HPRT: ipoxantina-adenina-fosforibosil-transferasi. Una sostanza conosciuta come polietilengucole permette la fusione. In pratica i passaggi hanno inizio da spienociti di topo opportunamente immunizzato, che vengono fusi con una sospensione di cellule di mieloma di topo: un terreno particolare noto come HAT (ipoxantina, aminopterina e timidina) consente la selezione delle cellule che interessano. I doni HPRTche non possono usufruire di ipoxantina esogena muoiono, e la stessa sorte hanno le cellule di milza che non sono andate incontro a fusione. Sopravvivono soltanto gli ibridi che sono dotati delle due proprietà conferite dalla fusione: espansione e capacità di sopravvivenza nel terreno HAT. Ciascun ibrido sintetizza un anticorpo con proprie specificità. Il tempo medio di raddoppio è di 48 h; le colonie si formano in opportuni micropozzetti dove è stata originariamente deposta una sola cellula (metodo della diluizione limite).
La tecnologia degli anticorpi monoclonali ha avuto un immediata applicazione clinica e nella diagnostica medica. Sono in corso i vari approcci sperimentali per un'opportuna applicazione nel settore della terapia oncologica. Un impiego oggi routinario deriva dalla possibilità di identificare sottopopolazioni linfocitarie con diverse caratteristiche funzionali. Per esempio, un anticorpo monoclonale è selettivamente specifico per una struttura nota come CD4, che caratterizza i linfociti a prevalente funzione helper. In corso di AIDS, come è noto, sono proprio i linfociti CD4 che risultano maggiormente colpiti dal virus e quindi, con una metodica relativamente semplice (IF, immunofluorescenza, o FACS, fluorescence activated cell sorting), è possibile monitorizzare il danno provocato dall'HIV. Notevoli progressi sono stati compiuti nello studio dei vari momenti evolutivi delle popolazioni cellulari timo-dipendenti, grazie alla possibilità di disporre di molecole di anticorpo specifiche per particolari antigeni di maturazione.
I metodi di laboratorio volti a studiare le funzioni delle cellule immunocompetenti o le loro caratteristiche morfologiche, pur con notevoli difficoltà per i problemi di standardizzazione e riproducibilità, hanno contribuito grandemente alla correlazione tra fenomeni clinici e risposte in vitro. La tecnica della ‛rosettazione' ha un posto privilegiato nella storia della moderna immunologia. Infatti i linfociti T umani (a differenza dei B) possono legarsi a emazie di montone e dar luogo alla suggestiva immagine di una cellula circondata da globuli rossi, la ‛rosetta'. L'applicazione dei vari pannelli di anticorpi monoclonali ha reso oggi più sofisticato il riconoscimento dei vari fenotipi cellulari, ma il ricorso al legame tra emazie di montone e linfociti T è ancora molto usato per alcuni passaggi durante vari metodi di separazione cellulare. I linfociti B non si legano a emazie di montone ma presentano sulla superficie di membrana le immunoglobuline. Utilizzando opportuni antisieri fluorescinati è possibile identificare le immunoglobuline di membrana o altri recettori. La possibilità di utilizzare anticorpi fluorescenti monoclonali tramite l'impiego della citometria a flusso e del cell sorting ha consentito di superare certi limiti delle precedenti tecniche, sebbene il costo di apparecchi come il FACS (fluorescence activated cell sorter) renda ancora validissime alcune delle metodiche ‛classiche'. Il legame tra antigene e cellula sensibilizzata induce un processo di attivazione citoplasmatica con diverse caratteristiche. Per esempio i linfociti ‛attivati' si trasformano in linfoblasti se vengono messi a contatto con alcune particolari sostanze. Molto utile è la fitoemoagglutinina (PHA), comunemente impiegata per studiare determinate funzioni linfocitarie. Le sostanze impiegate in queste metodiche si chiamano genericamente lectine e hanno uno spettro variabile di specificità: per esempio la PHA e la ConA (concanavalina A) sono mitogeni (cioè inducono mitosi) nell'ambito dei linfociti T, mentre il PWM (pokeweed mitogen) stimola i B. Per scopi di diagnosi clinica vengono usati anche i lipopolisaccaridi o la proteina A dello stafilococco, che è uno specifico stimolante dei linfociti B. Per valutare il livello di risposta si usa del materiale radioattivo (timidina marcata con 3H): calcolando l'entità di incorporazione della molecola radioemittente si può ottenere l'indice di attivazione. Naturalmente è possibile usare vari antigeni e studiare la risposta specifica verso di essi.
La coltura mista linfocitaria rappresenta un altro caso di stimolazione, in cui l'attivatore è una cellula opportunamente trattata che viene messa a contatto con il potenziale responder. Usando anche in questo sistema dei traccianti radioattivi, si può studiare la sintesi del DNA nelle cellule in grado di rispondere. L'applicazione clinica di tale metodica è mirata a identificare gli antigeni di istocompatibilità che possono condizionare l'esito dei trapianti d'organo.
La scoperta delle cellule NK (natural killers), morfologicamente caratterizzate dall'appartenere ai LGL (large granular lymphocytes), ha rappresentato un ulteriore successo dell'indagine di laboratorio. La metodica classicamente impiegata per valutare la funzione killer consiste nel mettere a contatto una cellula bersaglio (clone K-562) con le cellule da sottoporre al test. Dalla quantità di 51Cr liberato per lisi dalle K-562 (monitorizzabile perché radioattivo) si può ricavare un'analisi del funzionamento delle cellule NK.
Per quanto riguarda l'immunità aspecifica (funzioni dei neutrofili polimorfonucleati), si debbono distinguere le cinque tappe che completano l'attività microbicida: 1) movimento rivolto verso il bersaglio (chemiotassi); 2) legame del recettore sulla molecola opsonizzante l'antigene (binding o riconoscimento); 3) formazione del fagosoma dopo ingestione; 4) azione dei lisosomi che ‛versano' il loro contenuto enzimatico (perossidasi-idrolasi) all'interno del vacuolo digestivo (degranulazione); 5) attivazione del killing intracellulare. Esistono comunque varie altre funzioni che producono un killing intracellulare e tra queste si ricorda il cosiddetto burst respiratorio. La possibilità di studiare l'emissione elettromagnetica da parte dei neutrofili dopo ingestione dei microrganismi è stata sfruttata in laboratorio per valutare l'efficacia del killing intracellulare. Sfruttando la liberazione di un particolare tipo di ossigeno altamente reattivo con i substrati lisosomiali, è possibile una valutazione diretta del processo ossidativo, che è proporzionale all'emissione di luce osservabile con un sistema di amplificazione delle radiazioni emesse. Questo processo, noto come chemiluminescenza, risulta deficitario, per esempio, in una grave condizione patologica nota come malattia cronica granulomatosa (v. Stossel e altri, 1972; v. immunologia e immunopatologia).
6. Prospettive della terapia immunologica
Sin dall'inizio del secolo XX, quando gli studi sull'immunità cellulare e sulle vaccinazioni facevano sperare in un rapido ‛controllo' della risposta immunitaria, i ricercatori hanno coltivato il sogno di poter intervenire per finalità terapeutiche nelle diverse fasi dell'interazione antigene-sistema immunitario. Tuttavia, al contrario di quanto auspicato, i successi dei tentativi di attuare una terapia immunologica non sono stati numerosi, escludendo ovviamente l'applicazione su vasta scala delle vaccinazioni, che hanno portato al controllo di gravi malattie infettive e all'eradicazione del vaiolo. La storia della medicina e la storia dell'evoluzione biologica dell'uomo ci insegnano che non esiste un vero punto di arrivo e il recente episodio della pandemia di AIDS, tutt'altro che sotto controllo, dimostra come sia necessario adattare volta per volta le conoscenze acquisite alle esigenze del momento. Infatti per il virus (i virus) dell'AIDS non esiste una prospettiva di vaccino in senso ‛classico' e quindi ancora molto lavoro sarà necessario in questo settore. Comunque in questa sede si vuole prendere in esame quanto è stato fatto nell'ambito delle terapie immunologiche in grado di agire nelle diverse fasi della risposta immunitaria. Si deve innanzitutto partire dai due concetti di immunosoppressione e immunopotenziamento o immunoattivazione (v. Giunchi e altri, 1981), ma va precisato che certe espressioni del linguaggio enfatizzano a volte concetti non chiari e non ben definiti. Per esempio, nel corso degli anni settanta si è molto parlato (e si continua oggi a parlare) di immunomodulazione e di farmaci immunomodulanti. Ci sembra doveroso sottolineare come le nostre conoscenze non siano ancora così approfondite da permetterci di utilizzare molecole biologiche, o costruite in laboratorio, in modo talmente sofisticato da privilegiare selettivamente un certo tipo di risultato rispetto a un altro. Qualcuno ha parlato, forse con spirito critico, di immunointerferenza e prudentemente gli anglosassoni hanno coniato la sigla BRM (biological response modifiers) per raggruppare una serie di molecole con la proprietà di modificare la risposta biologica.
Per ottenere l'immunosoppressione si fa ricorso a: sieri antilinfocitari; farmaci ad azione citotossica; cortisonici; azatioprina; ciclofosfammide; ciclosporina; radiazioni; chirurgia.
Per l'immunoattivazione o immunopotenziamento si utilizza invece una serie di altri prodotti, in parte di estrazione biologica e in parte di origine sintetica, fra cui: alcuni prodotti batterici (BCG, bacillo Calmette-Guerin); nucleotidi ciclici; alcuni peptidi sintetici o derivati imidazolici (levamisole); il transferfactor (estratto leucocitario); estratti timici (ormoni timici); il sistema interferon (IFN); il sistema delle interleuchine (IL-2).
a) Immunosoppressione
Gli effetti immunosoppressivi dei farmaci possono variare in funzione di diversi parametri: la frequenza di somministrazione, il dosaggio, il rapporto tra tempo di somministrazione ed eventuale carico antigenico. Inoltre l'effetto di una molecola farmacologica può variare a seconda delle modalità con le quali viene somministrata. Per esempio, i corticosteroidi e alcune immunoglobuline antilinfocitarie (sieri antilinfocitari) esercitano un'azione marcatamente soppressiva se vengono somministrati assieme all'antigene, ma se questo è introdotto molto tempo prima gli effetti sono scarsi. Ovviamente il danno prodotto da radiazioni, agenti alchilanti e così via, pur essendo selettivo nei confronti di particolari cellule bersaglio, è esteso e si manifesta sia su cellule immunocompetenti che su altri distretti. Le informazioni raccolte sull'effetto dei farmaci immunosoppressori a livello delle diverse popolazioni linfocitarie sono incomplete, ma l'impiego degli anticorpi monoclonali ha consentito di selezionare meglio alcuni dati sperimentali. Si può dire in generale che alcune molecole privilegiano specifiche sottopopolazioni linfocitarie: la ciclofosfammide agisce primariamente sulla serie B e in un secondo tempo sui linfociti T; questi risentono selettivamente dell'azione dei corticosteroidi, che hanno un effetto linfocitolitico anche sui linfociti B. Allo scopo di distruggere una particolare popolazione cellulare sono stati preparati dei sieri antilinfocitari (ALS) iniettando i linfociti in riceventi xenogenici. Qualche risultato è stato ottenuto in particolari condizioni (per esempio in alcune sindromi autoimmuni), ma la preparazione iniziale, la variabilità dei quantitativi usati e gli svantaggi dovuti a frequenti effetti collaterali di intolleranza ne hanno fortemente sconsigliato l'uso. Non va inoltre dimenticato quanto esposto nel capitolo sull'immunoregolazione: gli effetti farmacologici e biologici sono tra loro connessi e il risultato finale passa attraverso un reticolo di interazioni. Per esempio, in corso di una sindrome autoimmune nella quale sia dimostrabile un incremento numerico e/o funzionale dei linfociti CD4+ (T helper), potrebbe essere utile un farmaco ‛immunostimolante' nei confronti di cellule a prevalente azione ‛soppressiva'. Per poter ottenere una situazione di questo tipo occorre prevedere l'esistenza di un ‛recettore' selettivo a livello della membrana dei linfociti suppressor che sia assente però sulle altre popolazioni cellulari.
Il tipo di risposta finale è quindi il risultato ottenuto bilanciando diverse azioni di interferenza.
Un farmaco immunosoppressore che ha contribuito grandemente alla riuscita dei trapianti ed è stato impiegato su larga scala è la ciclosporina. Si tratta di un polipeptide ciclico composto da 11 amminoacidi, che si è dimostrato efficace nell'inibizione dello sviluppo delle reazioni immunitarie mediate da cellule, comprendenti i fenomeni da ipersensibilità cutanea ritardata, reazione da trapianto verso l'ospite e produzione di anticorpi dipendente dai linfociti T. L'azione della molecola è specificamente diretta sui linfociti T, in modo reversibile. E l'immunosoppressore di scelta nella prevenzione del rigetto del trapianto allogenico di organi e nel trapianto di midollo osseo ha indicazione nella profilassi della graft vs host disease.
b) Immunostimolazione
Nell'ambito dell'immunostimolazione prenderemo in esame le molecole che fanno ritenere possibile per il futuro una loro applicazione in diverse situazioni cliniche.
La famiglia degli interferoni. - Si tratta di proteine osservate in diversi animali e che somigliano, per certe loro caratteristiche, agli ormoni. Gli IFN hanno funzioni diverse e originariamente sono stati caratterizzati grazie alle loro specifiche proprietà antivirali. Vengono prodotti dalle cellule infettate da virus e le molecole sono liberate nell'ambiente extracellulare subito dopo la loro sintesi (v. Isaacs e Lindenmann, 1957). Possono quindi legarsi a recettori specifici su altre cellule o seguire il flusso dei liquidi organici. Quando gli IFN raggiungono il sito specifico recettoriale sulla membrana della cellula danno luogo a un segnale citoplasmatico che innesca una serie di reazioni interne, il cui risultato finale consiste nel rendere la cellula temporaneamente non recettiva nei confronti di altri virus (v. Borden e Murphy, 1971). Le funzioni protettrici mediate dagli IFN si esplicano con la sintesi di altre proteine antivirali che svolgono essenzialmente una funzione enzimatica. I principali tipi di IFN sono: IFN alfa, di origine leucocitaria; IFN beta, proveniente da fibroblasti; IFN gamma, ottenuto da linfociti sensibilizzati.
L'approfondimento delle conoscenze sul significato dell'IFN in fisiologia e patologia ha consentito una visione più completa del ruolo biologico svolto da tali famiglie molecolari. Per esempio, oltre all'azione antivirale, è stata messa in evidenza una qualche azione antitumorale e sono stati proposti i primi protocolli di terapia antineoplastica. Successivamente si è dimostrata la presenza di IFN nel siero di soggetti con diverse malattie immunologiche (sindromi autoimmuni, deficit immunitari) e si è parlato degli IFN come di un complesso sistema ‛immunomodulante' in grado di interagire con diversi substrati cellulari durante l'esplicarsi di una risposta immunitaria. Per esempio è di grande interesse la capacità degli IFN di rallentare o bloccare la moltiplicazione cellulare (sia di cellule normali che in corso di crescita neoplastica), di solito con un semplice effetto citostatico reversibile nel corso del tempo. L'azione immunomodulante degli IFN si esplica sia a carico dell'immunità umorale che di quella cellulare: riduzione della sintesi di immunoglobuline (quando impiegati a bassa concentrazione), stimolo alla secrezione di anticorpi con aumento della concentrazione di Ig (se vengono impiegate alte dosi), aumento dell'attività natural killer e incremento delle funzioni citotossiche mediate dal sistema ADCC (citotossicità anticorpo-mediata). L'impiego degli IFN in terapia è stato limitato per molto tempo a causa della ridotta disponibilità del prodotto. Oggi, grazie alla tecnologia del DNA ricombinante, se ne possono avere quantità inesauribili. I maggiori successi si sono avuti in corso di alcune infezioni virali (cheratiti erpetiche, forme virali ginecologiche e dermatologiche), mentre la sperimentazione nel campo delle neoplasie è tuttora in fase preliminare (v. Arvin e altri, 1978; v. Marcovici e altri, 1983). Buoni risultati comunque vengono segnalati ai nostri giorni nel trattamento delle forme cutanee di sarcoma di Kaposi durante AIDS, nella leucemia a cellule ‛capellute' (hairy cell leukemia degli anglosassoni; v. sangue: Leucemie), nella leucemia mieloide cronica e nel mieloma.
Forse è giunto il momento, per questa particolare forma di terapia, di uniformare i protocolli terapeutici con l'attivazione di studi multicentrici: infatti spesso i risultati acquisiti con l'IFN non sono facilmente confrontabili tra di loro per la notevole eterogeneità dei trattamenti attuati e per le consistenti variazioni sia nelle dosi applicate che nella durata dei trattamenti stessi.
Ormoni timici. - L'estendersi delle conoscenze nell'ambito dell'immunità cellulare ha consentito di approfondire il ruolo cruciale del timo nell'ontogenesi e nell'assetto omeostatico del sistema immunitario. Tra le sue numerose funzioni, il timo rende possibile la risposta agli alloantigeni e permette di riconoscere il self distinguendolo da ciò che è estraneo all'organismo. Organo di forma grossolanamente piramidale, viene diviso in una zona corticale e una midollare. Cellule linfoidi e similmacrofagiche unitamente alla componente epiteliale ne costituiscono le principali popolazioni. La componente epiteliale elabora e secerne dei polipeptidi che vengono, almeno in parte, liberati nel flusso ematico (v. Bach e Goldstein, 1980). Il complesso di tali peptidi prende il nome comprensivo di ‛ormoni timici'; infatti possiedono le caratteristiche peculiari delle molecole ormonali, in quanto sono liberati in circolo e vanno ad agire in periferia sul bersaglio dei linfociti T. Il loro effetto si manifesta non solo sui linfociti pre-timici, ma anche su quelli T maturi. Vari ricercatori si sono occupati dello studio e della purificazione degli ormoni timici. Sono stati loro attribuiti diversi nomi e sigle e comunque viene generalmente ammesso che essi non svolgano azioni sovrapponibili. Nell'ambito dei preparati studiati in laboratorio o già in commercio è assai interessante un complesso polipeptidico estratto e purificato dal timo bovino: si tratta della timostimolina (v. Falchetti e altri, 1977), che è in grado di indurre la maturazione dei linfociti T e modula la risposta cellulo-mediata interferendo anche con la sintesi anticorpale. Utile soprattutto nelle immunodeficienze primitive (sindrome di DiGeorge, candidiasi cronica mucocutanea, sindrome di Wiskott-Aldrich, deficit T periferico, atassia-teleangectasia), trova un'utile possibilità di impiego nelle gravi infezioni virali (virus dell'herpes simplex, virus vancella-zoster, Citomegalovirus, Adenovirus) di pazienti a rischio e nelle complicanze infettive concomitanti a malattie neoplastiche (v. Aiuti e altri, 1979; v. Aiuti e altri, 1980; v. Aiuti e altri, Placebo..., 1984). Qualche dato incoraggiante nella letteratura su un'azione diretta al controllo delle metastasi in particolari tumori non sembra aver avuto un'ulteriore conferma. Nelle malattie autoimmuni l'uso è quanto mai discutibile, se non del tutto controindicato. Di interesse analogo alla timostimolina è anche un prodotto di sintesi: la timopentina (v. Goldstein e altri, 1979). Si tratta di un polipeptide che rappresenta una frazione attiva della timopoietina (ormone timico naturale), composto da 5 amminoacidi (arginina, lisina, asparagina, valina, tirosina) distribuiti nelle sequenze 32-36 della molecola naturale (v. Heavner, 1985). Le azioni sui linfociti T vengono mediate dal GMPc intracellulare e si svolgono in vario modo, con il risultato finale di riequilibrare alcune funzioni immunitarie. Il polipeptide ha una capacità differenziatrice sia in senso fenotipico che funzionale sui linfociti T e risulta di utile applicazione nelle infezioni correlate a difetto dei T helper. Alcuni studi sembrano dimostrarne una qualche efficacia nell'attivazione di T suppressor, facendo quindi ipotizzare il suo impiego nelle malattie autoimmuni (sebbene valgano anche in questo caso le riserve già avanzate per la timostimolina).
Linfochine e prospettive di terapia antineoplastica. - Si è visto come la risposta immunitaria e la sua regolazione dipendano sia da fattori solubili che dall'interazione fra cellule modulatrici. Volendo schematizzare le varie tappe che portano a una risposta immunitaria completa, dobbiamo partire dal contatto tra antigene e cellule ‛presentanti' l'antigene stesso. Il linfocita T, dopo l'interazione tra APC (antigen presenting cell) e molecole di MHC (complesso maggiore di istocompatibilità) esposte sulla membrana, esprime il recettore per una particolare sostanza nota come IL-2 ed esso stesso la produce. L'IL-2 (interleuchina 2) stimola il linfocita che l'ha prodotta e conduce a un'amplificazione del meccanismo di proliferazione. L'IL-2 aumenta la secrezione di interferon e amplifica l'attività sia delle cellule natural killers che del gruppo delle cellule note come LAK (cellule con funzione killer, attivate dall'interleuchina). L'IL-2 umana è un polipeptide dotato di un solo ponte disolfuro e costituito da 133 amminoacidi. Essa provoca, dopo il legame con il suo recettore specifico, il passaggio dalla fase G1 del ciclo cellulare alla fase S (v. cellula: Fisiologia della). Questa attivazione implica l'innesco di successive tappe regolatorie sul sistema T e B. Infatti il linfocita T elabora il BCGF (B cell growth factor) e il BCDF (13 cell differentiation factor), i quali agiscono sulla linea filogenetica di maturazione B, amplificando sia il meccanismo di proliferazione che quello di differenziamento. Ma il linfocita T-helper può iniziare il complesso meccanismo descritto soltanto se, oltre al contatto con l'antigene (Ag), riceve anche un segnale che parte dalle cellule macrofagiche presenti nel contesto di ogni processo infiammatorio. Tale segnale è rappresentato da un'altra linfochina (glicoproteina di circa 15 kdalton di peso molecolare) che prende il nome di IL-1. L'IL-1 è dotata di numerose proprietà (induce la proliferazione dei timociti, aumenta l'attività NK, è un potente pirogeno) e funziona sinergeticamente con l'IL-2. Lo schema seguente riassume i punti fondamentali del processo
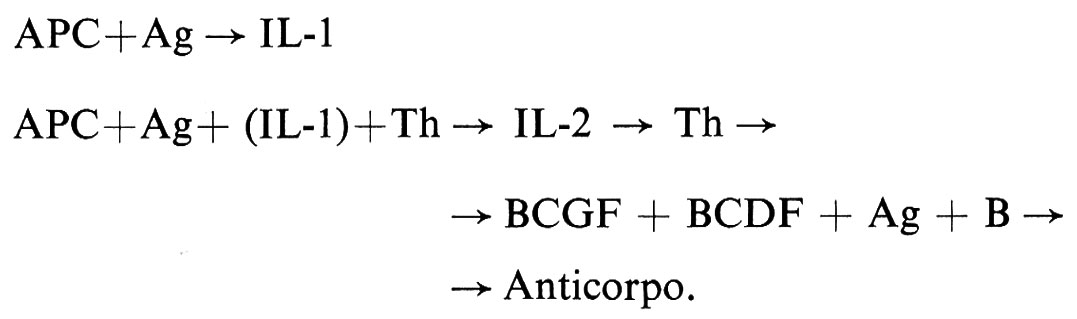
Nel corso degli studi sull'immunità cellulare è stato individuato il gene codificante l'IL-2 e oggi sono disponibili anticorpi monoclonali diretti contro l'IL-2 e il suo recettore. Altre interleuchine identificate sono l'IL-3, l'IL-4 e l'IL-5, ma il loro ruolo è tuttora da definire.
Durante un processo infiammatorio si affollano nella sede di reazione numerose cellule con funzioni diverse, assieme ai loro prodotti. Si possono ricordare alcuni dei più noti: MIF (fattore di inibizione macrofagica), MAF (fattore di attivazione macrofagica), PAF (fattore di attivazione piastrinica) e numerosi altri.
L'IL-2, come si è visto, svolge un ruolo centrale nell'attivazione di alcune cellule linfocitarie. Poiché queste, opportunamente stimolate, possono acquisire proprietà citotossiche nei confronti di altre cellule bersaglio (per esempio neoplastiche) si è pensato di usare l'IL-2 in alcuni modelli di terapia oncologica. L'approccio è reso ancora più interessante dalla dimostrazione dell'esistenza di TIL (tumor infiltrating lymphocytes) che farebbero parte di una sottopopolazione di cellule LAK (lymphokine activated killer cells). Nell'uomo sarebbero incluse in un gruppo fenotipicamente CD8 + (che comprende le cellule a funzione suppressor o citotossiche). Il razionale dello schema terapeutico prevede il pretrattamento in vitro con IL-2 di cellule LAK provenienti da donatori con tumore solido metastatizzato e la successiva reinoculazione associando IL-2. I protocolli iniziali hanno avuto un discreto successo, sebbene gli effetti collaterali di tipo congestizio ne abbiano limitato l'applicazione. Attualmente il tentativo più avanzato consiste nell'isolare TIL (che sono dotati di maggiore selettività rispetto alle LAK) e nello stimolare le loro funzioni associando IL-2 dopo trattamento con antiblastici (ciclofosfammide). Alcuni protocolli sono in sperimentazione.
bibliografia
Adachi, M., Yodoi, J. e altri, Altered expression of lymphocyte FC alfa receptor in selective IgA deficiency and IgA nephropathy, in ‟Journal of immunology", 1983, CXXXI, pp. 1246-1252.
Adami, G. F., Vitale, B., Bachi, V., Delayed hypersensitivity serial evaluatin by means of a multitest system IRCS, in ‟Medical science", 1981, IX, pp. 1132-1140.
Aiuti, F., La sindrome da immunodeficienza acquisita (AIDS), in ‟Immunologia clinica e sperimentale", 1983, II, pp. 67-74.
Aiuti, F., AIDS e sindromi correlate, in Il Convegno su AIDS e sindromi correlate. Status quo 1984. Roma, dicembre 1984, Roma 1984.
Aiuti, F., Allen, J. R., Bijerk, H., AIDS in Europe. Status quo 1983, in ‟European journal of cancer and clinical oncology", 1984, XX, pp. 155-173.
Aiuti, F., Ammirati, P., Fiorilli, M., D'Amelio, R., Franchi, F., Calvani, M., Businco, L., Immunological and clinical investigation on a bovine thymic extract therapeutic applications in primary immunodeficiencies, in ‟Pediatric rsearch", 1979, XIII, pp. 797-802.
Aiuti, F., Businco, L., Effects of thymic hormones in immunodeficiency, in ‟Clinics in immunology and allergology", 1983, III, pp. 187-191.
Aiuti, F., Businco, L., Rossi, P., Quinti, I., Response to thymopoietin pentapeptide in a patient with DiGeorge syndrome, in ‟Lancet", 1980, I, pp. 91-93.
Aiuti, F., Ensoli, B., Scarpati, B., Carbonari, M., Scano, G., Papetti, C., Sirianni, M. C., Rossi, P., Nuovi aspetti etiologici, epidemiologici e immunologici della sindrome da immunodeficienza acquisita e delle sindromi ad essa correlate in Italia, in ‟Medicina. Rivista dell'Enciclopedia medica italiana", 1984, IV, pp. 383 ss.
Aiuti, F., Rossi, P., Sirianni, M. C., Carbonari, M., Popovic, M., Sarngadharan, M. G., Contu, L., Moroni, M., Romagnani, S., Gallo, R. C., IgM and IgG antibodies to human T cell lymphotropic retrovirus (HTLV-III) in LAS syndrome and subjects at risk for AIDS in Italy, in ‟British medical journal", 1985, CCXCI, pp. 165-166.
Aiuti, F., Sirianni, M. C., Fiorilli, M., Paganelli, R., Stella, A., Turbessi, G.A., Placebo controlled trial of thymic hormone treatment of recurrent herpes simplex labialis infection in immunodeficient host: results after a one year follow-up, in ‟Clinical immunology and immunopathology", 1984, XXX, pp. 11-18.
Akkerman, J. W. N., Van Breerode, W., Gorter, G., Zegers, B. J. M., Kmis, W., The Wiskott-Aldrich syndrome: studies on a possible defect in mithocondrial ATP resynthesis in platelets, in ‟British journal of haematology", 1982, LI, pp. 561-568.
Alexander, P., Escape from immune destruction by the host through shedding of surface antigens: is this a characteristic shared by malignant and embryonic cell?, in ‟Cancer research", 1974, XXXIV, pp. 2077-2084.
Altman, L., Snyderman, R. M., Blaese, R. M., Abnormalities of chemotactic lymphokine synthesis and monocyte leukocyte chemotaxis in Wiskott-Aldrich syndrome, in ‟Journal of clinical investigation", 1984, LIV, pp. 486-493.
Arvin, A. M., Feldman, S., Merignan, T. C., Human leukocyte interferon in the treatment of varicella in children with cancer: a preliminary controlled trial, in ‟Antimicrobial agents and chemotherapy", 1978, XIII, pp. 605-612.
Bach, J. F., Goldstein, G., New concept of thymic hormones, in ‟Thymus", 1980, II, pp. 1-8.
Barandun, S., Morell, A., Skvaril, F., Clinical experiences with immunoglobulin for intravenous use, in Immunoglobulins: characteristics and uses of intravenouis preparations. Proceedings", Bethesda 1980.
Baroni, C.D., Pezzella, F., Mirolo, M., Ruco, L. P., Rossi, G. B., Immunohistochemical demonstration of p24 HTLV-III major core protein in different cell types within lymph nodes from patients with lymphadenopathy syndrome (LAS), in ‟Histopathology", 1986, X, pp. 5-13.
Bartorelli, A., L'antigene CEA e gli antigeni associati a carcinoma della mammella, in Immunologia dei tumori (a cura di E. Sega), Roma 1980, pp. 293-301.
Blaese, R. M., Strober, W., Levy, A. L., Waldmann, T. A., Hypercatabolism of IgG and IgM and albumin in the Wiskott-Aldrich syndrome, in ‟Journal of clinical investigation", 1971, L, pp. 2331-2338.
Borden, E. C., Murphy, F. A., The interferon refractory state: in vivo and in vitro studies of its mechanisms, in ‟Journal of immunology", 1971, CVI, pp. 134-140.
Broder, S., Strategies for experimental therapy of the retrovirus which causes AIDS, in Acquired ID Syndrome (a cura di J.C. Gluckman e J. Vilmer), Amsterdam 1987.
Broder, S., Gallo, R. C., A pathogenic retrovirus (HTLV-III) linked to AIDS, in ‟New England journal of medicine", 1984, CCCXI, pp. 292-296.
Brun-Vézinet, F., Barré-Sinoussi, F., Saimot, A. G., Christol, D., Montagnier, L., Rouzioux, C. e altri, Detection of IgG antibodies to lymphoadenopathy associated virus in patients with AIDS and LAS, in ‟Lancet", 1984, I, pp. 1265-1268.
Bruton, O.C., Agammaglobulinemia, in ‟Pediatrics", 1952, I, pp. 1265-1268.
Buckley, R. H., Gilertesen, R. B., Schiff, R. I., Ferreira, E., Sanai, S. O., Waldmann, T. A., Heterogeneity of lymphocyte subpopulations in SCID: evidence against a stem cell defect, in ‟Journal of clinical investigation", 1976, LVIII, pp. 130-139.
Burgio, G. R., Duse, M., Monafo, V. e altri, Selective IgA deficiency: clinical and immunological evaluation of 50 pediatric patients, in ‟European journal of pediatrics", 1980, CXXXIII, pp. 101-106.
Burnet, F. M., The clonal selection theory of immunity, Nashville 1959.
Businco, L., Pandolfi, F., Rossi, P. e altri, Selective defect of a T-helper subpopulation in SCID, in ‟Journal of clinical immunology", 1981, I, pp. 125-130.
Ciccimarra, F., Rosen, F. S. e altri, Failure of heavy chain glycosylation of IgG in some patients with commonv ariable agammaglobulinemia, in ‟Journal of clinical investigation", 1976, LVII, pp. 1386-1391.
??? Classification system for HTLV-III/LAS associated infection, in ‟Mortality morbidity weekly report (MMWR)", 1986, XXXV, pp. 336-341.
Cleveland, W. W., Fogel, B. J., Brown, W. T., Kay, H. E. M., Foetal thymic transplant in a case of DiGeorge's sydnrome, in ‟Lancet", 1968, II, pp. 1211-1214.
Clifford Lane, H., Masur, H., Edgar, L. C. e altri, Abnormalities of B cell activation and immunoregulation in patients with the acquired immunodeficiency syndrome, in ‟New England journal of medicine", 1983, CCCIX, pp. 153-157.
Coffman, R. L., Weissmann, I. I., Immunoglobuline gene rearrangement during pre-B cell differentiation, in ‟Journal of molecular and cellular immunology", 1983, I, pp. 31-37.
Coggin, J. H., Anderson, N. G., Cancer differentiation and embryonic antigens: some central problems, in ‟Advances in cancer research", 1984, XIX, pp. 105-165.
Cooper, M. D., Kearney, J., Scher, I., B lymphocytes, in Fundamental immunology (a cura di W. E. Paul), New York 1984, pp. 43-45.
Cooper, M. D., Lawton, A. R., Circulating B cells in patients with immunodeficiency, in ‟American journal of pathology", 1972, LXIX, pp. 513-517.
Dicke, K. A., Schafer, U. W., Van Bekkum, D. W., The use of stem clel concentrates as bone marrow grafts in combined immunodeficiency, in ‟Transplantation", 1983, V, pp. 909-912.
DiGeorge, A.M., Congenital absence of the thymus and its immunologic consequences. Concurrence with congenital hypoparathyroidism, in Immunologic deficiency diseases (a cura di D. Betgsma e F. A. McKusik), Baltimore 1968, pp. 116-122.
Elson, C. O., Is IgA production T-cell dependent!, in ‟Immunology today", 1985, V, pp. 5-6.
Falchetti, R., Bergesi, G., Eshkol, A. e altri, Pharmacological and biological properties of a calf thymus extract, in ‟Drugs under experimental and clinical research", 1977, III, pp. 39-43.
Farrant, J., Bryiant, A. E., Knight, S. C., Webster, A. D. B., Defective mitogen presentation by dendritic-like cells in patients with common variable hypogammaglobulinemia, in Primary immunodeficiency diseases (a cura di M. M. Eibl e F. S. Rosen), New York 1986, Pp. 277-280.
Fiorilli, M., Businco, L., Pandolfi, G. F., Paganelli, R., Russo, G., Aiuti, F., Heterogeneity of immunological abnormalities in ataxiateleangiectasia, in ‟Journal of clinical immunology", 1983, III, pp. 135-141.
Fiorilli, M., Russo, G., Paganelli, R., Papetti, C., Carbonari, M., Crescenzi, M., Calvani, M., Quinti, I., Aiuti, F., Hypogammaglobulinemia with hyper IgM severe T-cell defect and abnormal recirculation of OKT4+ lymphocytes in a girl with chronic lymphadenopathy, in ‟Clinical immunology and immunopatology", 1986, XXXVIII, pp. 256-264.
Gallo, R., Salahuddin, S. S., Popovic, M., Shearer, G. M., Kaplan, M., Haynes, B. F., Palker, T. J., Redfield, R., Oleske, J., Safai, B., White, G., Foster, P., Markam, V., Frequent detection and isolation of cytopathic retrovirus (HTLV-III) from patients with AIDS and at risk for AIDS, in ‟Science", 1984, CCCXI, pp. 500-503.
Gatti, R. A., Menwissen, H. J., Allen, H. D., Hong, R., Good, R. A., Immunologic reconstitution of sex-linked lymphopenic immunologic deficiency, in ‟Lancet", 1968, II, pp. 1366-1369.
Geha, R. S., Is the B-cell abnormality secondary to T-cell abnormality in SCID?, in ‟Clinical immunology and immunopathology", 1976, VI, pp. 102-112.
Geha, R. S., Scheneeberger, E., Merler, E., Rosen, F.S., Heterogeneity of acquired' or common variable agammaglobulinemia, in ‟New England journal of medicine", 1964, CCXCI, pp. 1-6.
Giblett, E. R., Ammann, A. J., Wara D. W., Sandman, R., Diamond, L. K., Nucleoside-phosphorilase deficiency in a child with severely defective T-cell immunity and normal B-cell immunity, in ‟Lancet", 1985, I, pp. 1010-1013.
Giblett, E. R., Anderson, J. E., Cohen, F., Pollara, B., Menwissen, H. J., ADA-deficiency in two patients with severely impaired cellular immunity, in ‟Lancet", 1972, II, pp. 1067-1069.
Giunchi, G., Aiuti, F., D'Amelio, R., Concetti attuali di immunoterapia, Padova 1981.
Goldberg, L.S., Barnett, E. V.,Fudenberg, H. H., Selective absence of IgA: a family study, in ‟Journal of laboratory and clinical medicine", 1968, LXXII, pp. 204-212.
Goldstein, G., Scheid, M. F., Boyse, E.A., Schlesinger, D. H., Van Wauve, J. A., A synthetic pentapeptide with biological activity characteristic of the thymic hormone thymopoietin, in ‟Science", 1979, CCIV, pp. 1309-1310.
Grant, G. H., Wallace, W. D., Agammaglobulinemia, in ‟Lancet", 1954, II, pp. 671-674.
Heavner, G. A., Thymopentin structure and relation to thymic factors, in ‟Survey of immunological research", 1985, IV, pp. 11-14.
Hoffman, T., Winchester, R., Schulkind, M. e altri, Hypogammaglobulinemia with normal T cell function in female siblings, in ‟Clinical immunology and immunopathology", 1977, VII, pp. 364-371.
Hoyer, J.R., Cooper, M. D., Gabrielsen, A. E., Lymphopenic forms of congenital immunologic deficiency diseases, in ‟Medicine", 1968, XLVII, pp. 201-210.
Isaacs, A., Lindemann, J., Virus interference. I. The interferon, in ‟Proceedings of the Royal Society. Biological section", 1957, CXLVII, pp. 258-267.
Jerne, N. K., Towards a network theory of the immune system, in ‟Annales d'immunologie", 1974, CXXV c, pp. 373-381.
Johnson, B. L. Jr., Goldberg, L. S. e altri, Clinical and immunological studies in a case of nodular lymphoid hyperplasia of the small bowel, in ‟Gastroenterology", 1971, LXI, pp. 369-374.
Kenny, A. B., Hitzig, W. H., Bone marrow transplantation for severe combined immunodeficiency, in ‟European journal of pediatrics", 1979, CXXXI, pp. 155-159.
Klein, G., Klein, E., Evolution of tumors and the impact of molecular oncology, in ‟Nature", 1985, CCCXV, pp. 190-195.
Levy, R. M., Bredesen, D. E., Rosenblum, M. L., Neurological manifestations of the acquired immunodeficiency syndrome (AIDS): experience at UCSF and review of the literature, in ‟Journal of neurosurgery", 1985, LXII, pp. 475-495.
Luzi, G. e altri, Comunicazione al X Convegno del Gruppo Italiano di Cooperazione in Immunologia, Cortona 1986.
Luzi, G. e altri, Il Registro italiano delle immunodeficienze primitive, in ‟Federazione medica", 1986, IX, pp. 1179-1197.
Luzi, G., Ensoli, B., Turbessi, G., Scarpti, E., Aiuti, F., Transmission of HTLV-III infection by heterosexual contact, in ‟Lancet", 1985, II, p. 1018.
Luzi, G., Kubagawa, H. e altri, Analysis of IgG subclass production in cell cultures from IgA deficient patients and in normal controls as a function of age, in ‟Clinical and experimental immunology", 1986, LXV, pp. 434-442.
Marcovici, R., Peretz, B. A., Paldi, E., Human fibroblast IFN in patients with condiloma acuminata, in ‟Israel journal of medical science", 1983, XIX, pp. 104-112.
Milstein, C., Gli anticorpi monoclonali, in Le difese immunitarie (a cura di F. Celada), Milano 1986, pp. 209-217.
Montagnier, L., Chermann, J. C., Barré-Sinoussi, F. e altri, A new human T-lymphotropic retrovirus: characterization and possible role in LAS and AIDS, in Human T-cell leukemia/lymphoma virus (a cura di R. C. Gallo, M. Essex e L. Gross), New York 1984, pp. 363-370.
Nossal, G. J. V., Lederberg, J., Antibody production by single cells, in ‟Nature", 1958, CLXXXI, pp. 1419-1434.
Nossal, G. S. V., The basic components of the immune system, in ‟New England journal of medicine", 1987, CCCXVI, pp. 1320-1325.
O'Reilly, R., Aiuti, F., Kirkpatrick, C., Stiehm, R., Griscelli, C., Criteri per la diagnosi delle SCID, in ‟Recenti progressi in medicina", 1978, LXIV, pp. 265-273.
Pahwa, S., Pahwa, R., Saxinger, C., Gallo, R. C., Good, R. A., Influence of the human T-lymphotropic virus on functions of human lymphocytes: evidence for immuno suppressive effects and polyclonal B-cell activation by banded viral preparation, in ‟Proceedings of the National Academy of Sciences", 1985, LXXXII, pp. 8198-8202.
Paulter, L. W., Seymour, G. J., Duke, O., Janossy, G. e altri, Immunohistological analysis of delayed-type hypersensitivity in man, in ‟Cellular immunology", 1982, LXXIV, pp. 358-369.
Prehn, R. T., The immune reaction as a stimulator of tumor growth, in ‟Science", 1972, CLXXVI, pp. 170-186.
Racz, P., Tenner-Racz, K., Kahl, C., Feller, A. C., Kern, P., Dietrich, M., Spectrum of morphologic changes of lymph nodes from patients with AIDS or AIDS-related complex, in ‟Progress in allergy", 1986, XXXVII, pp. 81-181.
Redfield, R.R., Craig, D.W., Tramont, E. C., The Walter Reed staging classification for HTLV-III/LAV infection, in ‟New England journal of medicine", 1986, CCCXIII, pp. 131-132.
Resnick, L., Di Marzo-Veronese, F., Schupbach, J., Tourtellotte, W. W. e altri, Intra-blood-brain-barrier synthesis of HTLV-III specific IgG in patients with neurologic symptoms associated with AIDS or AIDS-related complex, in ‟New England journal of medicine", 1985, CCCXIII, pp. 1498-1504.
Rich, K. C., Richman, C. M., Nejias, E., Daddona, P., Immunoreconstitution by peripherla blood leukocytes in adenosine deaminasedeficient severe combined immunodeficiency, in ‟Journal of clinical investigatoin", 1980, LXVI, pp. 389-395.
Robbins, J. B., Eitzman, D. V., Ellis, E. F., Immunochemical evidence for development of acquired' hypogammaglobulinemic state, in ‟New England journal of medicine", 1966, CCLXXIV, pp. 607-614.
Rosen, F. S., The lymphocytes and the thymus gland congenital and hereditary abnormalities, in ‟New England journal of medicine", 1968, CCLXXIX, pp. 643-651.
Salmon, S. E., Smith, B. A., Lehner, R. I. e altri, Modification of donor lymphocytes for transplantation in lymphopenic immunological deficiency, in ‟Lancet", 1970, II, pp. 149-152.
Schneider, J., Junkiewicz, E., Hayami, M., Desroisiers, R., Marx, P., Hunsmann, G., Serlogical and structural comparison of HIV, SIVmac, SIVagm, and SIVsm, four primate lentiviruses, in Acquired ID syndrome (a cura di J. C. Gluckman e J. Vilmer), Amsterdam 1987, pp. 63-69.
Shcwaber, J., Rosen, F. S., Somatic cell hybrids and mouse myeloma cells and B lymphocytes from a patient with agammaglobulinemia: failure to secrete human immunoglobulin, in ‟Journal of immunology", 1979, CXXII, pp. 1849 ss.
Shaham, M., Voss, R., Becker, Y., Yarkoni, S. e altri, Prenatal diagnosis of ataxia-telangiectasia, in ‟Journal of pediatrics", 1982, C, pp. 134-137.
Snider, W. D., Simpson, D. M., Nielsen, S., Gold, J. W. M., Metroka, C. E., Posner, J. B., Neurological compliations of acquired immune deficiency syndrome: analysis of 50 patients, in ‟Annals of neurology", 1984, XIV, pp. 403-418.
Spector, E. D., Filipovich, A. H., Perry, G. S. III, Kersey, J. H., Epidemiology of cancer in ataxia-telangiectasia, in Ataxia-telangiectasia: a cellular and molecular link between cancer neuropathology and immune deficiency (a cura di B. A. Bridges e D. G. Harnden), New York 1982, pp. 103-138.
Stocker, F., Amman, P., Rossi, E., Selective A-globulin deficiency with autosomal inheritance in a Swiss family, in ‟Archives of disease in childhood", 1968, XLIII, pp. 585-591.
Stossel, T. P., Root, R. K., Vaughan, M., Phagocytosis in chronic granulomatous disease and the Chediak-Higashi syndrome, in ‟New England journal of medicine", 1972, CCLXXXVI, pp. 120-128.
Strober, W., Wochner, R. D., Barlow, M. H., McFarlin, D. E., Waldmann, T. A., Immunoglobulin metabolism in ataxia-telangiectasia, in ‟Journal of clinical investigation", 1969, XLVII, pp. 1905-1915.
Stuckey, M., Quinn, P. A., Gelfand, E. W., Identification of ureaplasma urealyticum (T-strain mycoplasma) in patient with polyarthritis, in ‟Lancet", 1978, II, pp. 917-920.
Tedder, F. F., Cooper, M. D., Clement, L.T., Human lymphocyte differentiation antigens HB-10 and HB-11. II. Differential production of B cell growth and differentiation. Factors by distinct helper T cell subpopulations, in ‟Journal of immunology", 1985, CXXXIV, pp. 2989-2994.
Tonegawa, S., Somatic generation of antibody diversity, in ‟Nature", 1983, CCCII, pp. 575-581.
Trinchieri, G., Perussia, B., Human natural killer cells: biologic and pathologic aspects, in ‟Laboratory investigation", 1984, L, pp. 489-496.
Tursz, R., Preud'homme, J. L., Labaume, S. e altri, Autoantibodies to B lymphocytes in a patient with hypogammaglobulinemia: characterization and pathogenic role, in ‟Journal of clinical investigations", 1977, LX, pp. 405-410.
Wong-Staal, F., Gallo, R. C., Human T-lymphotropic retroviruses, in ‟Nature", 1985, CCCXVII, pp. 395-403.
Zinkernagel, R. M., Doherty, P. C., MHC-restricted cytotoxic T cells: studies on the biological role of polymorphic major transplantation antigens determining T-cell restriction specificity, function and responsiveness, in ‟Advances in immunology", 1979, XXVII, pp. 51-64.
Zinnermann, H. H., Hall, W. H., Heller, B. I., Acquired agammaglobulinemia: report of 3 cases, in ‟Journal of American Medical Association", 1954, CLVI, pp. 1390-1394.
Zolla-Pazner, S., B cells in the pathogenesis of AIDS, in ‟Immunology today", 1984, V, pp. 289-290.