
Genetica. Analisi genetica della durata della vita e dell'invecchiamento
Genetica. Analisi genetica della durata della vita e dell'invecchiamento
Invecchiamento, senescenza e longevità sono concetti correlati, relativi a processi per lungo tempo ritenuti ineluttabili e quindi impossibili da analizzare mediante le comuni tecniche della biologia molecolare e cellulare e della genetica. In realtà non è così: tali processi sono sottoposti a controllo genetico; inoltre l'analisi genetica dell'invecchiamento e della senescenza è oggetto di studio ed è anzi probabile che questi, in futuro, possano essere controllati e perfino manipolati a piacimento. Lo sviluppo delle metodologie genetiche nell'ultimo quarto del XX sec. ha offerto la possibilità di identificare e alterare geni, quasi a piacimento, in molte specie di invertebrati e Vertebrati. Per quanto riguarda la specie umana, sebbene la terapia genica stia procedendo a un ritmo che pochi sarebbero stati in grado di predire fino a non molti anni fa, l'analisi genetica non può essere applicata nella sua piena potenzialità e così l'analisi fine dei processi d'invecchiamento è stata condotta su varie specie di invertebrati e, solo in modo limitato, sul topo.
Tali modelli sperimentali fungono da banco di prova per la determinazione dei processi coinvolti nella senescenza e possono essere poi sottoposti a verifica nella specie umana. Sono stati proposti diversi criteri che debbono essere presi in considerazione nella scelta di un sistema modello per l'invecchiamento, che includono una vita breve, l'accessibilità alla sperimentazione genetica e la disponibilità di ulteriori informazioni di base. I sistemi modello più usati per comprendere le basi genetiche dei processi d'invecchiamento includono quelle specie viventi, per le quali gli strumenti per l'analisi genetica sono stati maggiormente affinati nel corso degli anni. Non si può inoltre prescindere dai costi della sperimentazione, che variano considerevolmente da specie a specie, così come il tempo di realizzazione degli esperimenti, da pochi giorni nei funghi a più di tre anni nei topi.
L'analisi genetica dell'invecchiamento è stata oggetto di ricerca per almeno settant'anni, ma è solo negli ultimi decenni che gli approcci genetici relativi alla comprensione della durata della vita sono stati coronati da successo. Ciò è stato possibile dimostrando che i geni che prolungano la durata della vita possono essere identificati e manipolati usando tecniche classiche e di tipo quantitativo. Dall'inizio degli anni Ottanta del Novecento, in diverse specie, soprattutto di invertebrati, sono stati isolati molti mutanti 'longevi' e sono state create linee selezionate. Sono stati identificati e clonati geni coinvolti strettamente nel controllo dei processi d'invecchiamento. Nella specie umana sono stati isolati i geni responsabili di numerose malattie ereditarie, la più importante delle quali è il morbo di Alzheimer, ritenute in precedenza collegate al processo di invecchiamento. L'identificazione di geni che stanno alla base della longevità e delle patologie legate all'età contribuisce a chiarire la natura del processo che viene definito 'invecchiamento'. Sebbene le basi molecolari dell'invecchiamento siano ancora poco conosciute, si stanno compiendo progressi significativi e stanno emergendo numerose e importanti novità.
La senescenza nei funghi
Delezioni mitocondriali
I funghi sono stati utilizzati a lungo come modelli per lo studio dell'invecchiamento e della senescenza, per i vantaggi legati alla loro genetica e alla loro relativa semplicità di analisi. In entrambe le specie Podospora anserina e Neurospora crassa, la perdita della capacità di crescita (senescenza) è stata associata a riarrangiamenti del DNA mitocondriale. In P. anserina l'accumulo di genomi malfunzionanti e la conseguente proliferazione dei plasmidi determinano un fenotipo senescente. La senescenza può essere prevenuta da mutanti doppi in una coppia di geni nucleari, i viv o gr viv. Fino a oggi, l'unico gene clonato è gr (grisea), omologo alla famiglia ACE1 dei fattori di trascrizione eucariotici che svolgono un ruolo chiave nella regolazione dello scambio intracellulare del rame. Sebbene il suo ruolo nel mantenimento dell'integrità mitocondriale sia ancora poco chiaro, gr potrebbe regolare il livello della superossidodismutasi (SOD) a Cu/Zn. Anche in diversi ceppi di Neurospora la proliferazione di plasmidi mitocondriali è associata alla perdita della capacità di crescita.
Senescenza vegetativa
Kenneth Munkres e C.A. Furtek (1984) hanno identificato alcuni mutanti di N. crassa che presentano una prolungata sopravvivenza dei conidi (spore vegetative). Tali mutazioni risiedono in un gene denominato age-1, e sono correlate a un incremento delle attività di SOD e catalasi. Gli autori hanno ipotizzato che le cinque SOD, evidenziabili in Neurospora, siano tutte sotto il controllo del regulone age-1. Ceppi di Neurospora meno resistenti alle alte temperature erano invece correlati a mutazioni nei geni strutturali codificanti le SOD. Anche in lievito sono importanti per la sopravvivenza entrambi gli isoenzimi SOD (a Cu/Zn e a Mn); entrambi sono inoltre necessari in normali condizioni di aerazione, benché la delezione della SOD a Cu/Zn sia più deleteria; in condizioni di scarsa aerazione, la mancanza della forma a Mn causa una perdita più marcata di vitalità.
Durata limitata della vita proliferativa nel lievito
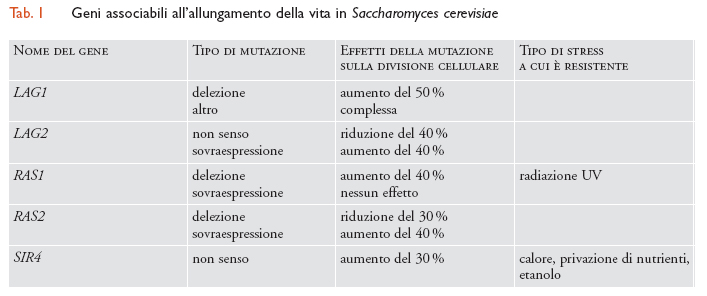
Alcuni trascritti variano quantitativamente durante la vita replicativa del lievito. Uno di questi, LAG1, quando è assente, determina l'allungamento della vita del lievito (tab. 1). LAG1 è una proteina, probabilmente integrale, di membrana e raggiunge il livello massimo di espressione a metà del corso della vita. Anche l'espressione dei geni RAS1 e RAS2 influenza la durata della vita: delezioni di RAS1 la prolungano di circa il 40% (e mostrano anche un'aumentata resistenza alle radiazioni UV), mentre delezioni del gene RAS2 la accorciano; una sovraespressione di RAS2 aumenta, parimenti, la longevità. Leonard Guarante e i suoi collaboratori sono invece andati a esaminare mutanti di lievito che presentavano una maggiore vitalità in condizioni di scarsi nutrienti e bassa temperatura; tali mutanti non solo mostravano un aumento della durata della vita, ma erano anche più resistenti agli stress termici da elevata temperatura.
Clonando, per complementazione, un gene con associato un fenotipo sterile, si è trovato che si tratta di una mutazione nel locus SIR4: questo gene svolge un ruolo nel silenziamento della cromatina sia nei loci del tipo sessuale (mating type) sia nei telomeri. La sir4-42 è una mutazione non senso (non sense) nel codone 1237 che elimina gli ultimi 121 amminoacidi della proteina ed è semidominante per l'aumento della durata della vita sia nelle cellule aploidi che in quelle diploidi. Sorprendentemente, le delezioni di SIR4 non generano un fenotipo 'vita più lunga' suggerendo che quest'ultimo sia il risultato, in sir4-42, dell'espressione di una qualche nuova funzione. Poiché la mutazione sir4-42 dà luogo all'accorciamento dell'estremità carbossiterminale, Brian Kennedy e i suoi collaboratori hanno creato nuovi mutanti in grado di sovraesprimere solo gli ultimi 154 amminoacidi carbossiterminali scoprendo che tali mutanti aumentavano la durata della vita in modo SIR4-dipendente. L'estensione della vita dipendeva da SIR2 e SIR3 ma non da SIR1, escludendo così i loci silenti del tipo sessuale e suggerendo un sito di azione telomerico. Una possibile spiegazione è che un gene Age (età) e un secondo gene nella regione telomerica, che conferisce resistenza allo stress, vengano inversamente modulati da un meccanismo dipendente da SIR. Molti ricercatori dubitano che il lievito costituisca un modello ottimale per questo tipo di studi sui Metazoi, in ragione della sua unicellularità e, quindi, dell'assenza di separazione tra linea germinale (destinata all'immortalità) e soma. Quanto tutto ciò sia vero, sarà chiarito certamente in un prossimo futuro.
Genetica dell'invecchiamento in Caenorhabditis elegans
Identificazione di mutanti e resistenza allo stress
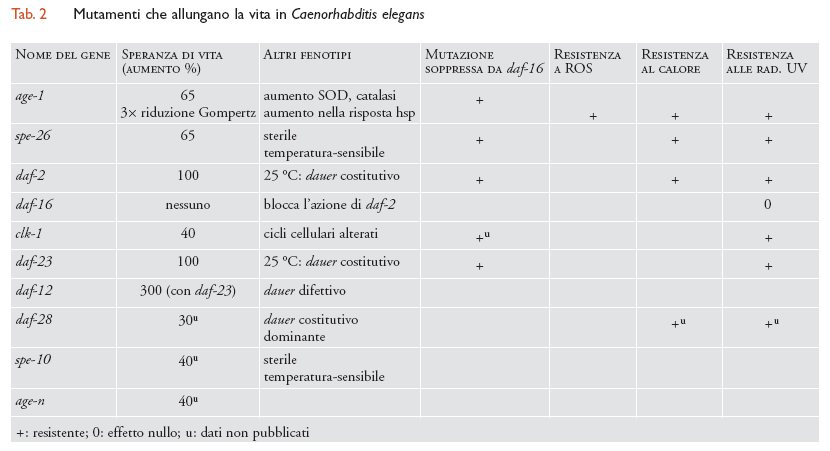
Caenorhabditis elegans è un nematode ampiamente usato negli studi di biologia dello sviluppo e sta diventando un modello sempre più importante anche per lo studio dell'invecchiamento, poiché il ciclo vitale di ermafrodito autofecondante consente un rapido inincrocio minimizzandone la conseguente depressione. Il primo mutante age-1 identificato ha una speranza di vita (life expectancy) aumentata del 65%, una durata massima della vita aumentata del 105% e un tasso tre volte più basso di mortalità. Tale mutazione ha uno scarso effetto sulla fecondità, sulla durata della riproduzione e sulla velocità dello sviluppo. I mutanti del gene age-1 sono inoltre più resistenti a danni da parte di ossidanti quali paraquat e H2O2, l'alta temperatura e le radiazioni UV. Tali mutanti accumulano delezioni del DNA mitocondriale (mtDNA) a un livello più basso e, oltre all'aumento della durata della vita, sono più resistenti agli stress ambientali e generalmente più sani (tab. 2).
Mutazioni in altri quattro geni in C. elegans conducono al fenotipo 'vita più lunga': (a) i mutanti spe-26 allungano del 65% la vita degli ermafroditi e dei maschi accoppiati; (b) a 16 °C, e in condizioni di normale tensione d'ossigeno, i mutanti rad-8 mostrano una durata media della vita più lunga del 50%, per lo più giustifica ta da uno sviluppo più lento; (c) i mutanti daf-2 presentano un raddoppiamento della durata media della vita; (d) le probabilità di raddoppiare la durata della vita, proprie dei mutanti daf-23, si quadruplicano in associazione con daf-12. Le mutazioni daf impediscono lo sviluppo di larve dauer, cioè un tipo di sviluppo alternativo intrapreso da C. elegans in condizioni definite 'tempi duri'; (e) alcuni alleli, ma non tutti, del nuovo gene clk-1, responsabile di uno sviluppo e di cicli cellulari più variabili, presentano un allungamento della vita. Esistono, inoltre, diversi altri casi, non ancora pubblicati, di loci per l'allungamento della durata della vita, che suggeriscono così che i gerontogeni identificati in C. elegans siano, in totale, circa dieci.
L'allungamento della vita indotto da daf-2 e daf-23 è soppresso dai mutanti dauer-difettivi del gene daf-16 e forse del gene daf-18; anche l'allungamento della vita indotto da age-1, spe-26 e clk-1 viene soppresso dai mutanti daf-16. Tutti i mutanti gerontotecnici finora studiati sono anche resistenti a stress ambientali, come quelli provocati dalle specie reattive dell'ossigeno (ROS, Reactive oxygen species), l'elevata temperatura e le radiazioni UV. Tra queste capacità di resistenza agli stress, quella alle radiazioni UV costituisce un segno più evidente di alungamento della vita; studi più completi con ROS portano per lo più a risultati identici. L'aumentata resistenza, conferita dai gerontogeni, a una varietà di agenti ambientali stressanti come quelli provocati dalle specie reattive dell'ossigeno, l'elevata temperatura e soprattutto le radiazioni UV, e i presunti ruoli di age-1, daf-2, daf-23 e daf-16 in una via di trasduzione del segnale mediato dalla PI3 chinasi sono compatibili con un modello in cui la durata della vita sia determinata dalla sensibilità dell'organismo all'ambiente e dalla capacità di resistere o porre riparo ai danni ambientali. È importante sottolineare come questi risultati suggeriscano che la capacità di riparare i danni non sia massima nei Metazoi, che vi sia ulteriore capacità latente all'interno del genoma e che un aumento di tale capacità conduca a un allungamento della vita.
Analisi di ceppi ricombinanti inincrociati
Si possono generare varianti genetiche dotate di vita più lunga anche mediante la produzione di ceppi ricombinanti inincrociati (RI). I ceppi RI, ottenuti da incroci fra i due ceppi selvatici N2 e Bergerac, sono stati utilizzati per verificare le teorie evolutive basate sull'ipotesi che la durata della vita possa essere definita da geni con pleiotropia antagonistica, ossia geni che hanno effetti opposti su caratteri adattativi nel primo e nell'ultimo periodo della vita. Quattro QTL (Quantitative trait loci, ovverosia geni responsabili di parte della differenza tra le linee di discendenza) che determinano un allungamento della vita sono stati mappati sui cromosomi 2, 4, 5 e X. Ricerche indipendenti hanno invece trovato QTL responsabili per il 90% della variabilità della durata della vita sui cromosomi 2, 4 e X, e QTL per l'autofecondità negli ermafroditi nei gruppi di associazione 2, 3 e 4, a loro volta responsabili per l'80% della variabilità genetica relativa alla fecondità. Tuttavia, gli autori non hanno trovato evidenze che suggeriscano un'associazione dei QTL per la fecondità con quelli per la durata della vita, come proposto dalla teoria evolutiva; esistevano, invece, alcune variazioni epistatiche fra loci, importanti per spiegare come la variabilità genetica sia mantenuta nelle popolazioni naturali.
Analisi genetica dell'invecchiamento in Drosophila
Assenza di mutanti a vita lunga
Mutanti a vita lunga sono stati identificati in C. elegans, nel lievito e nei funghi, ma è noto un solo mutante di questo genere (grandehildless) in Drosophila subobscura. Nonostante esistano in Drosophila melanogaster molti mutanti con fenotipo femmina sterile, non sono state ripetute in questa specie meglio studiata le osservazioni precedentemente fatte in D. subobscura. Non è stato perciò ancora stabilito se mutazioni in geni singoli possano portare o meno a un aumento della durata della vita in altre specie oltre a C. elegans e alcuni funghi. È possibile che specie prive di una lunga aplofase (presente invece nei funghi) o che non siano inincrociate naturalmente (come accade in C. elegans) accumulino mutazioni recessive deleterie, che determinano depressione da inincrocio. In questi casi, quindi, sarebbe più opportuna un'analisi in popolazioni in cui vi sia libero incrocio (outcross), benché si presentino, in tal caso, numerosi problemi per l'identificazione e il successivo clonaggio dei geni.
Incrocio selettivo per l'allungamento della vita
Sono stati conseguiti successi sperimentali sia incrociando individui con più alta età riproduttiva sia tramite selezione diretta in funzione della lunghezza della vita. È stato così possibile aumentare la durata della vita in individui di Drosophila di circa 30 giorni, usando una popolazione di laboratorio recentemente stabilita partendo da moscerini selvatici da poco catturati, e in realtà è stato sufficiente adattare alle condizioni di laboratorio per 130 generazioni una popolazione selvatica, per ottenere un incremento della durata della vita di 10 giorni per le femmine e di 6 giorni per i maschi. B. Zwaan e collaboratori hanno suddiviso una popolazione in subpopolazioni replicate, mantenute in condizioni di bassa temperatura, mentre i singoli individui erano analizzati e selezionati per la durata della vita; moscerini selezionati, discendenti dalle subpopolazioni, venivano quindi utilizzati come riproduttori per le generazioni successive. Le linee longeve mostravano una lieve diminuzione della fecondità negli stadi precoci della vita, suggerendo l'esistenza di un'alternativa tra fecondità nelle prime fasi vitali e fecondità in fasi più avanzate.
Resistenza allo stress
Oltre alla riduzione della fecondità nei giovani, è stato identificato un certo numero di alterazioni fisiologiche che potrebbero essere alla base dell'allungamento della vita. Il peso dell'ovario in individui giovani di ceppi a vita lunga è ridotto; inoltre, questi ceppi erano resistenti alla privazione di nutrienti, alla disidratazione e ad altri stress ambientali. Parimenti, incroci selettivi per resistenza alla privazione di nutrienti o alla disidratazione danno un aumento di longevità pari al 40%. Ci sono inoltre modificazioni metaboliche: la glucosio-6-fosfatodeidrogenasi è espressa maggiormente nelle linee a vita lunga ed è anche riscontrabile una certa resistenza alle ROS (radicali liberi) e, infatti, alcuni studi suggerirebbero che sia implicata una SOD iperattiva, anche se questi dati necessitano ancora di una definitiva conferma.
Identificazione dei geni alla base della longevità
L'analisi a livello dei singoli geni coinvolti nell'aumento della durata della vita ha presentato difficoltà quasi insuperabili. L'aumento della longevità risulta dall'azione combinata di centinaia di loci. La natura non inincrociata (outbred) delle linee rende estremamente difficile l'identificazione di geni individuali. Diversi gruppi di ricerca hanno comunque dimostrato che linee distinte, selezionate per il fenotipo longevo, possiedono geni, implicati nell'allungamento della vita, situati sul cromosoma 3. Un altro problema di cui si deve tenere conto in specie a riproduzione sessuale, come Drosophila e il topo, è la depressione da inincrocio. Le linee selezionate sono, relativamente a molti loci, meno inincrociate dei ceppi originari di partenza o dei controlli; ciò suggerisce che la sovradominanza possa svolgere un ruolo significativo nell'allungamento della vita di queste linee. Se la risposta alla selezione deriva da un aumento della eterozigosi, il fenotipo vita lunga non può essere studiato in ceppi inincrociati e l'analisi fine di caratteri genetici risulta particolarmente problematica, dal momento che la genetica molecolare può essere applicata solo a uno o a pochi geni alla volta.
Analisi genetica della durata della vita nei Mammiferi
L'analisi genetica della durata della vita nei Mammiferi, prevalentemente condotta sul topo, ha ottenuto pochi risultati in confronto ai dati ottenuti sugli invertebrati. La maggiore lunghezza della vita dei Mammiferi e il costo più elevato degli animali da sperimentazione non hanno consentito di approfondire sufficientemente gli studi sulla longevità.
Studi sulle funzioni immunitarie
È stato ipotizzato che la maggior parte dei geni specifici per la durata della vita debbano essere ricercati nella regione del complesso maggiore di istocompatibilità (MHC, Major histocompatibility complex) del topo. Sono state identificate diverse regioni, situate su otto cromosomi, come possibili QTL per la longevità, ma sfortunatamente studi indipendenti non hanno identificato i medesimi siti e non hanno confermato il ruolo svolto dalla regione MHC nella determinazione della speranza di vita. Tuttavia, la regione H-2 mostrava l'effetto maggiore in individui F2, anche se sembra che i risultati siano più significativi per la determinazione dei loci per vita breve e non per vita lunga. In contrasto con quanto detto, la selezione per un aumento o una diminuzione della funzione immunitaria altera drasticamente la durata della vita. Purtroppo la risposta più evidente alla selezione è stata un accorciamento della vita (speranza di vita di solo 346 giorni) nella linea caratterizzata da funzione immunitaria ridotta. Analisi successive su incroci fra linee ad alta e bassa funzione immunitaria hanno consentito l'identificazione di diversi QTL presunti, fra cui i loci MHC e lgh, così come le regioni già note sui cromosomi 2, 4 e 6, anche se questi risultati necessitano di conferme indipendenti.
Lunghezza della riproduzione
Diversi dati suggeriscono l'esistenza di una correlazione positiva tra la lunghezza del processo riproduttivo e la durata della vita. S. Lerner e collaboratori hanno studiato l'effetto dell'MHC sulla capacità riproduttiva delle femmine. J.F. Nelson e collaboratori hanno dimostrato l'esistenza di un controllo genetico sugli eventi puberali (apertura vaginale, prima corneificazione vaginale) e sulla ciclicità estrale, sia in termini di lunghezza sia di frequenza. J. Nagai e collaboratori (1990) sono riusciti a selezionare due linee di topo caratterizzate da aumenti significativi nella capacità riproduttiva e nella durata della vita rispetto alle linee di controllo non selezionate. Esistono, tuttavia, diversi problemi connessi a tali studi: (a) le linee selezionate mostravano livelli riproduttivi inferiori a quelli osservati in molti ceppi inincrociati; (b) gli individui di queste linee hanno sofferto della presenza di numerosi parassiti; (c) la longevità delle linee era molto ridotta (377 e 436 giorni), assai più che non in molti ceppi inincrociati; (d) inoltre, la capacità riproduttiva non si è mostrata così duratura ed elevata in linee selezionate per un aumento della riproduzione, mediamente pari a solo 241 giorni, un valore inferiore a quanto riscontrato in molti ceppi inincrociati.
Topi transgenici
Sono stati intrapresi diversi studi su animali transgenici, ma solo due hanno dimostrato un effetto significativo sull'allungamento della vita. Tongying Zhou e collaboratori (1995), usando topi transgenici per Fas, una molecola bifunzionale coinvolta nella apoptosi e nella stimolazione dei linfociti T, hanno mostrato che individui vecchi (26 mesi) hanno livelli di apoptosi indotta da Fas e proliferazione di cellule T confrontabili con quelli caratteristici degli individui giovani; in questo esperimento si verificava però solo un aumento limitato (ca. un mese) della speranza di vita. Topi transgenici per l'attivatore del plasminogeno di tipo urochinasico (αMUPA, Urokinase-type plasminogen activator) posto sotto il controllo del promotore dell'αA-cristallina (espresso per lo più nel cristallino dell'occhio), sono stati prodotti da Ruth Miskin e collaboratori (1990). Poiché tali topi consumavano meno cibo ed erano di minori dimensioni, se ne è controllata la durata della vita, scoprendo che presentavano un incremento del 30% del tasso di sopravvivenza per ogni classe di età. L'assenza di pubblicazioni su animali transgenici per la SOD e la catalasi è emblematica: poiché questi topi sono ormai disponibili da molti anni, se ne può dedurre che la sovrapproduzione di SOD verificatasi in queste linee non abbia portato a un allungamento della vita.
Analisi genetica dei fenomeni demografici negli invertebrati
Il tasso di mortalità dipendente dall'età aumenta durante la vita con una cinetica pressoché esponenziale per la maggior parte delle specie; questa osservazione è così generale che è quasi divenuta garanzia di 'vero' invecchiamento dell'organismo. Questo aumento esponenziale del tasso di mortalità è stato modificato geneticamente sia in C. elegans sia in Drosophila. I mutanti age-1 di C. elegans sono caratterizzati da una riduzione di tre volte del tasso di accelerazione della mortalità rispetto al tipo selvatico, ma non presentano una modificazione statisticamente significativa del tasso di mortalità in età giovanile. Gli individui maschi presentano tassi di accelerazione della mortalità anche notevolmente più alti e una durata della vita più ridotta rispetto agli ermafroditi; questo aumento è più lento nel mutante age-1, ma in misura inferiore a quanto avviene negli ermafroditi. Questa riduzione del tasso di accelerazione della mortalità è una caratteristica recessiva dell'allele mutante age-1. Il tasso di mortalità può anche essere modificato poligenicamente e i ceppi RI mostrano una variazione di tre volte nel tasso stesso, alcuni aumentando più rapidamente rispetto al ceppo selvatico, altri più lentamente. Il tasso di mortalità legato all'età varia in diversi ceppi inincrociati di Drosophila e sia il tasso di mortalità iniziale sia quello di accelerazione della mortalità risultano alterati nelle linee di Drosophila selezionate per la longevità.
Osservazioni sulla mortalità in età avanzata possono essere eseguite solo in popolazioni di grandi dimensioni. Un'interruzione dell'incremento esponenziale della mortalità si osserva, nei Nematodi, all'incirca al termine del periodo fecondo. Anche in Drosophila e nell'insetto Ceratitis capitata, il tasso di mortalità aumenta esponenzialmente per poche settimane per poi bloccarsi o perfino diminuire in tarda età. Questi risultati sono in disaccordo con il modello di Gompertz, secondo cui l'unico parametro da tenere in considerazione è una componente esponenziale della mortalità legata all'età. I tassi di mortalità sono costanti in età avanzata, usando una popolazione geneticamente eterogenea costituita da settantanove genotipi RI. La mortalità legata all'età aumenta esponenzialmente sino a 17 giorni, per mantenersi poi essenzialmente costante sino all'ultimo decesso avvenuto a circa 60 giorni. Questo periodo di costante tasso di mortalità legata all'età è un effetto dell'eterogeneità genetica e non è intrinseco all'invecchiamento: infatti ogni genotipo RI ha una durata della vita caratteristica e tassi di mortalità quasi esponenziali seppur differenti, finché quasi tutti gli individui sono morti.
Vitalità degli animali longevi
Individui di Drosophila e C. elegans geneticamente longevi sono stati studiati per verificare se la motilità (usata come indice generico di vitalità o di salute) risultasse alterata in ceppi a vita lunga. I mutanti age-1 sembrano essere caratterizzati da maggiore motilità in tutte le classi di età e sono in grado di mantenere questa caratteristica più a lungo, suggerendo così che le mutazioni che incrementano la longevità aumentano anche, in generale, vitalità e salute. I ceppi longevi RI mostravano un tasso di diminuzione della motilità che forniva un'accurata indicazione della durata media o massima della vita. Nelle linee a vita lunga di Drosophila era presente una chiara associazione tra una senescenza tardiva e un aumento della durata del volo. È quindi chiaro che la maggior parte dei meccanismi che sono alla base dell'allungamento della vita non coinvolgono un'estensione del periodo di senescenza alla fine della vita.
Genetica dell'invecchiamento nell'uomo
La teoria evolutiva suggerisce che, poiché l'invecchiamento non costituisce un carattere adattativo, la senescenza potrebbe essere dovuta alla mancata selezione contro geni deleteri nel periodo terminale della vita (ovvero, nella vita postriproduttiva). Esistono esempi specifici di tali geni? Essi svolgono un ruolo nell'invecchiamento umano? Noi riteniamo che i geni coinvolti nelle amiloidosi familiari a insorgenza tardiva siano un esempio particolarmente chiaro di tali geni.
Amiloidosi
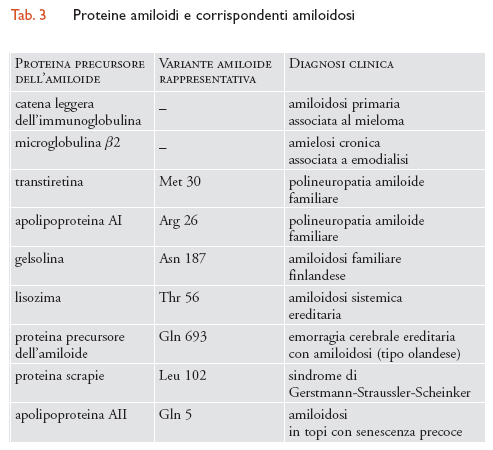
Il termine 'amiloide' si riferisce, genericamente, a depositi insolubili protidici in cui la sostanza amiloide assume un'evidente struttura fibrillare, e fu coniato, per errore, quando si pensava che i depositi fossero composti di materiale simile all'amido. La presenza dell'amiloide è rilevata, di solito, mediante l'uso di coloranti specifici, come il rosso congo, che si legano, nelle fibrille amiloidi, alla conformazione a foglietto β pieghettato delle proteine. Depositi di sostanza amiloide possono essere rilevati in diversi tessuti del corpo umano, spesso in condizioni chiaramente patologiche. Molte proteine differenti, apparentemente con nessuna relazione, possono formare amiloidi; tipi diversi di amiloidi sono associati a differenti malattie (tab. 3). Le amiloidosi possono essere suddivise in forme ereditarie e non ereditarie. Queste ultime sono spesso associate a disfunzioni fisiologiche specifiche e non hanno una relazione intrinseca con l'età; per esempio, l'amiloidosi della catena leggera delle immunoglobuline è associata al mieloma multiplo o alla macroglobulinemia primaria, mentre l'amiloidosi della microglobulina β2 è associata alla emodialisi. Le amiloidosi ereditarie presentano invece un'insorgenza tipicamente tardiva.
Transtiretina
La transtiretina, precedentemente denominata 'prealbumina', è una proteina di 127 amminoacidi secreta nel fegato e nel plesso corioideo del cervello. Essa lega sia la tirosina sia la proteina legante il retinolo e si rinviene in forma tetramerica nel siero, dove svolge un ruolo nel trasporto di tirosina e retinolo. La transtiretina inizialmente venne associata a una amiloidosi ereditaria, poiché fu dimostrato che formava i depositi amiloidi trovati in pazienti affetti da polineuropatia amiloide familiare (FAP, Familial amyloid polyneuropathy), una malattia autosomica dominante a insorgenza tardiva (quarta o quinta decade di vita), caratterizzata da una grave patologia dei nervi periferici e, in ultimo, da morte. Successivamente, studi sulla FAP hanno mostrato la presenza di una sostituzione valina-metionina nel gene per la transtiretina. Si ritiene che tale mutazione aumenti la capacità della transtiretina di assumere una conformazione amiloide. In seguito, sono state identificate molte altre amiloidosi sistemiche familiari determinate dalla transtiretina e tutte (più di quaranta) sono associate a sostituzioni amminoacidiche nella sequenza codificante la transtiretina. Si noti che, in generale, queste varianti della transtiretina, non possiedono alterate capacità di legame alla tirosina o al retinolo e la patologia amiloide non è associata alla compromissione delle funzioni della transtiretina stessa. Tali varianti sono perciò tipicamente dannose solo in età avanzata.
La polineuropatia amiloide familiare è una malattia rara, con un'incidenza di circa un caso su 100.000, e ha, quindi, scarsa rilevanza nei processi di invecchiamento della maggior parte delle persone. Tuttavia, anche i depositi amiloidi rinvenuti nell'amiloidosi sistemica senile, osservata in più del 25% di persone con oltre 80 anni, sono composti di transtiretina. In questo caso, l'amiloide è composta di transtiretina di 'tipo selvatico', cioè non mutata, e tutte le patologie associate con tali depositi amiloidi sono generalmente non gravi. Un'interpretazione ragionevole di queste osservazioni è che la transtiretina sia una proteina intrinsecamente amiloidica e, quindi, potenzialmente tossica. Tuttavia, poiché cospicui depositi amiloidi di transtiretina si formano tipicamente solo in età avanzata, non si è avuta alcuna selezione contro di essa. Le varianti rinvenute nella FAP esaltano questa tossicità intrinseca, determinando situazioni che sono senz'altro riconosciute come stati patologici. Vari altri geni possono risultare lievemente alterati producendo proteine predisposte alla formazione di amiloidi tossici (tab. 3), quali per esempio, la gelsolina, la cistatina, l'apolipoproteina AI e il lisozima. Inoltre, depositi amiloidi in un modello murino a senescenza accelerata sono associabili a una variante della apolipoproteina AII. Tali esempi non permettono di concludere che l'invecchiamento umano sia guidato dalla formazione di amiloidi tossici: geni con potenziale amiloidico a insorgenza tardiva sono gli unici ad avere effetti dominanti e a lasciare un'impronta istologica prontamente identificabile; chiaramente, geni che aumentassero la senescenza ma che fossero recessivi o debolmente semidominanti, sarebbero molto più difficili da individuare, specie se il loro meccanismo d'azione fosse ignoto o difficile da rilevare. Così i geni coinvolti nell'amiloidosi potrebbero essere solo i rappresentanti più evidenti di un'ampia classe di geni ignoti che influenzano la senescenza umana.
Il morbo di Alzheimer
Sebbene l'amiloidosi non consenta di spiegare molti aspetti dell'invecchiamento, il morbo di Alzheimer (AD, Alzheimer's disease), malattia su base amiloide, riveste un ruolo importante nell'invecchiamento dell'uomo. Questa demenza senile progressiva è caratterizzata da un aumento nella perdita della memoria, della capacità di ragionare e di comunicare e può essere direttamente o indirettamente fatale. Dati in letteratura riportano, complessivamente, l'incidenza della demenza senile tra 4,7%, all'età di 70 anni, e 49,6%, all'età di 90 anni. L'AD segrega come un tratto autosomico dominante ed è geneticamente eterogeneo. La principale caratteristica patologica è la formazione di placche senili nel cervello degli individui che ne sono affetti. Tali placche contengono tipicamente un nucleo di materiale amiloide extracellulare insolubile, chiamato β-amiloide, circondato da un alone di trame neurofibrillari e neuroni distrofici. La ben nota osservazione che la sindrome di Down (trisomia del cromosoma 21) sia quasi sempre associata all'insorgenza precoce dell'AD e la mappatura sul cromosoma 21 di alcuni casi di AD familiare, hanno portato i ricercatori a cercare mutazioni del gene APP associate alla malattia.
Geni per la longevità umana?
La correlazione fra l'età raggiunta alla morte in gemelli monozigoti e dizigoti suggerisce l'esistenza di un 20÷40% di ereditarietà per la lunghezza della vita nella specie umana. Anche i geni coinvolti in altre malattie legate all'età, come quelle cardiovascolari e il cancro, possono essere considerati validi candidati ad avere alleli associati alla longevità. In studi di associazione di marcatori genetici si è visto che la longevità è associata a regioni definite del genoma umano; entrambi i loci ACE e APOE sono stati analizzati in qualità di candidati gerontogeni e si è trovato che alleli di entrambi i geni sono associati all'aumento della longevità, sebbene non necessariamente nella direzione prevista in origine. Tali osservazioni non sono affatto sorprendenti, in ragione delle forti associazioni esistenti tra gli alleli studiati e i loro effetti sulla salute. Questi studi con marcatori genetici devono essere comunque convalidati da ulteriori ricerche. Molte malattie sono state denominate 'progeroidi segmentali', poiché simulano alcuni aspetti del normale invecchiamento. In particolare, due di esse simulano bene il normale invecchiamento umano: la progeria di Wemer, sul cromosoma 8, e quella di Hutchison-Gilford, mutazione spontanea dominante. La capacità limitata delle cellule somatiche umane di proliferare in colture tissutali ha portato all'adozione di tali linee cellulari come modelli dell'invecchiamento dei Mammiferi, anche se più che l'invecchiamento nell'uomo sembra rappresentino modelli utili per studiare il differenziamento terminale in vitro, il controllo del ciclo cellulare e l'oncogenesi.
Modificazioni genetico-molecolari alla base dell'invecchiamento
La riparazione del DNA e la mutazione somatica
È ben noto che nell'uomo la frequenza di mutazione somatica, nonché le frequenze di riarrangiamenti somatici soprattutto nei linfociti, aumentano con l'età cronologica. Anche diverse progenie umane mostrano elevate frequenze di mutazioni somatiche, di riarrangiamenti cromosomici o di entrambi. Tuttavia, non è stato mai possibile dimostrare in nessuna specie il ruolo causale delle mutazioni somatiche nell'invecchiamento. La teoria è stata sottoposta a verifica nei Nematodi e nella vespa Mormoniella vitripennis, usando ceppi difettivi per il sistema di riparazione dei danni da radiazione. Il risultato è che, in queste specie, la mutazione somatica non è il processo che limita la lunghezza della vita. Si è proposto che l'aumentata probabilità di contrarre il cancro con l'invecchiamento, la senescenza cellulare e, forse, anche l'invecchiamento stesso siano, nell'uomo, il risultato della progressiva perdita nel corso della vita di sequenze telomeriche nelle cellule somatiche. Questa 'ipotesi telomerica' per la senescenza cellulare non è stata mai dimostrata, sebbene un possibile ruolo della perdita di materiale telomerico nella senescenza replicativa di cellule umane e nell'invecchiamento in vivo sia corroborato da una grande quantità di dati derivanti dallo studio di diversi tipi di cellule somatiche. Non esiste, tuttavia, prova diretta del fatto che la perdita del telomero causi senescenza o invecchiamento nell'uomo o in altri Mammiferi. Ancor più dibattuta è la proposta che delezioni o altre alterazioni nel DNA mitocondriale possano causare direttamente l'invecchiamento. Esiste una chiara relazione tra età cronologica e alterazioni mitocondriali in numerose specie, e il tasso di tali delezioni è probabilmente controllato geneticamente. Tuttavia, il nesso causale tra invecchiamento e alterazioni del DNA mitocondriale non è stato dimostrato e non è affatto chiaro se l'aumento della frequenza delle alterazioni sia sufficiente a causare modificazioni funzionali rilevabili.
Effetto dell'età sulla sintesi dell'RNA
Sembra esserci una diminuzione generale, legata all'età, del tasso di sintesi e di degradazione dell'RNA, che mantiene per lo più costante il livello dell'RNA totale. Al livello di mRNA, si osserva una considerevole variabilità fra geni, alcuni dei quali sono attivati e altri repressi in funzione dell'età cronologica; per la maggior parte di essi, invece, il livello di espressione non risulta modificato. L'unico studio sull'espressione dell'mRNA in relazione a differenti genotipi è quello relativo al ceppo a vita lunga age-1 di C. elegans, dove non è stata osservata, rispetto al ceppo di controllo, alcuna differenza nell'espressione temporale di quei pochi trascritti che presentavano diversi livelli d'espressione in funzione dell'età cronologica. L'espressione genica è stata studiata anche in roditori sottoposti a limitazioni nella dieta (DR, Dietary restriction) e in Drosophila allevata a diverse temperature. La DR altera differenzialmente, per geni diversi, l'abbondanza dei trascritti corrispondenti. Essa aumenta significativamente l'espressione della α2u-globulina, probabilmente a livello del trascritto, alterando l'espressione dei fattori di trascrizione che regolano questo gene. Il gene hsp70 è stato studiato a lungo in epatociti di ratto dove viene indotto, a livello della trascrizione, dalla DR. Si è poi visto che questa attivazione è il risultato di un'alterazione del fattore di trascrizione da shock termico (HSTF, Heat shock transcription factor) specifico per hsp70, tale che un eccesso di molecole di questo fattore si trova in forma attiva.
Effetto dell'età sulla sintesi proteica
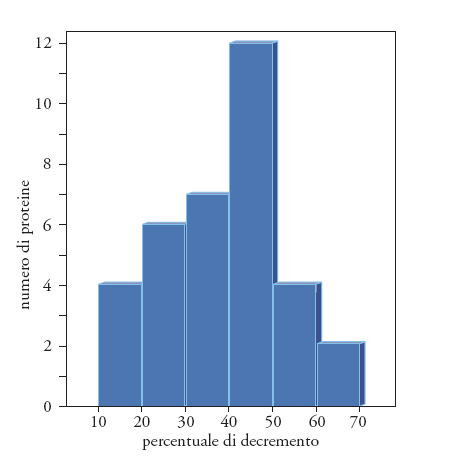
Si è cercato di connettere, a diversi livelli, la sintesi proteica con le modificazioni legate all'invecchiamento, ma sono state trovate poche o nessuna modificazione qualitativa nell'espressione delle proteine durante l'invecchiamento. C'è però un generale abbassamento dei tassi di sintesi e di degradazione proteica con l'aumentare dell'età cronologica. Per esempio, in Drosophila, J.E. Fleming e collaboratori hanno rilevato che trentasei proteine su quarantatré mostravano tassi di sintesi ridotti con l'aumentare dell'età, mentre solo una proteina aumentava enormemente il suo livello di sintesi. J. Butler e collaboratori (1989) hanno esaminato la sintesi proteica in epatociti isolati di ratto, trovando che trentacinque proteine su trentasei (di cui diciannove in modo statisticamente significativo) diminuivano in quantità con l'aumentare dell'età (fig. 3).
Invecchiamento e attività genica differenziale
Il ruolo di un'espressione genica alterata, come causa della senescenza, è oggetto di notevole interesse. La teoria evolutiva dell'invecchiamento propone che i geni alla base dell'invecchiamento sarebbero espressi o maggiormente espressi solo in età avanzata. In C. elegans, per esempio, l'analisi degli mRNA isolati da Nematodi di 14 giorni di età (corrispondenti alla durata media della vita del nematode) ha mostrato come meno del 2% dei geni fosse soggetto a livelli d'espressione quantitativamente diversi durante l'invecchiamento: nove trascritti diminuivano, due aumentavano lievemente e uno raggiungeva il suo massimo nel periodo medio della vita adulta. Mutazioni nel gerontogene age-1 non alteravano l'espressione genica. Tre degli mRNA che diminuivano in abbondanza erano già stati identificati come geni per la vitellogenina; l'unico trascritto che aumentava in abbondanza con l'età era omologo al fattore di traduzione EF1-α; gli altri cinque cloni rappresentano nuovi geni di Nematodi. È chiaro, quindi, che l'invecchiamento differisce dallo sviluppo: esso, in questi sistemi, non è guidato da un'espressione genica differenziale.
Per Drosophila si sta delineando un quadro in qualche modo diverso. Inizialmente fu proposto che EF1-α fosse un possibile agente della senescenza, come mostrato da studi che ne rivelavano una diminuzione durante la vita e, in seguito, da J.C.W. Shepherd e collaboratori, che dimostrarono come i moscerini con transgeni supplementari per EF1-α fossero più longevi. Studi successivi hanno dimostrato che, sebbene la linea transgenica sia realmente longeva, essa non produce maggiori quantità di EF1-α e non è, in realtà, più longeva del tipo selvatico. Inoltre, studi dettagliati su linee transgeniche diverse, in molteplici contesti genetici, hanno dimostrato che non è il transgene in quanto tale ma la posizione della sua inserzione e il contesto genetico a causare l'allungamento della vita. È stata osservata anche una chiara relazione inversa tra longevità e fecondità. Studi effettuati da Stephen L. Helfand e collaboratori (1995) hanno messo in evidenza come l'espressione genica non sia funzione dell'età assoluta ma piuttosto dell'età relativa (normalizzando i dati rispetto alle diverse durate della vita), in linea con un modello di invecchiamento programmato. Ciò è in netto contrasto con gli studi di Thomas J. Fabian e Thomas E. Johnson, i quali, lavorando con un ceppo age-1 a vita lunga, dimostrarono che non vi è alcun cambiamento, rispetto al tipo selvatico, nell'età assoluta dell'espressione genica. La genetica è un importante strumento che viene applicato all'analisi dei processi d'invecchiamento in una varietà di specie differenti e a diversi livelli, dal molecolare all'organismico, sui quali si stanno compiendo rapidi progressi e si spera che una comprensione dei processi d'invecchiamento sia raggiungibile in un futuro non lontano.
Bibliografia
Brooks 1994: Brooks, Anne - Lithgow, Gordon, J. - Johnson, Thomas E., Mortality rates in a genetically heterogenous population of Caenorhabditis elegans, "Science", 263, 1994, pp. 668-671.
Carey 1992: Carey, James R. e altri, Slowing of mortality rates at older ages in large medfly cohorts, "Science", 258, 1992, pp. 457-461.
Curtsinger 1992: Curtsinger, James W. e altri, Demography of genotypes: failure of the limited life-span paradigm in Drosophila melanogaster, "Science", 258, 1992, pp. 461-463.
Goto 1992: Goto, Makoto e altri, Genetic linkage of Werner's syndrome to five markers on chromosome 8, "Nature", 355, 1992, pp. 735-738.
Helfand 1995: Helfand, Stephen L. e altri, Temporal patterns of gene expression in the antenna of the adult Drosophila melanogaster, "Genetics", 140, 1995, pp. 549-555.
Johnson 1990: Johnson, Thomas E., The increased life-span of age-1 mutants in Caenorhabditis elegans results from lowering the Gompertz rate of aging, "Science", 249, 1990, pp. 908-912.
Kennedy 1995: Kennedy, Brian K. e altri, Mutation in the silencing gene SIR4 can delay aging in S. cerevisiae, "Cell", 80, 1995, pp. 485-496.
Miskin, Masos 1997: Miskin, Ruth - Masos, Tamar, Transgenic mice overexpressing urokinase-type plasminogen activator in the brain exhibit reduced food consumption, body weight and size, and increased longevity, "Journals of gerontology. Series A, Biological sciences and medical sciences", 52, 1997, pp. 118-124.
Morris 1996: Morris, Jason Z. - Tissenbaum, Heidi A. - Ruvung, Gary, A phosphatidylinositol-3-OH kinase family member regulating longevity and diapause in Caenorhabditis elegans, "Nature", 382, 1996, pp. 536-539.
Munkres, Furtek 1984: Munkres, Kenneth D. - Furtek, C.A., Selection of conidial longevity mutants of Neurospora crassa, "Mechanisms of ageing and development", 25, 1984, pp. 47-62.
Nagai 1995: Nagai, Junya - Lin, Ching-Yao - Sabour, Parviz M., Lines of mice selected for reproductive longevity, "Growth, development, and aging", 59, 1995, pp. 79-91.
Zhou 1995: Zhou, Tongying - Edwards, Carl K. III - Mountz, John D., Prevention of T age-related cell apoptosis defect in CD2-fas-transgenic mice, "Journal of experimental of medicine", 182, 1995, pp. 129-137.