
Citoscheletro
Citoscheletro
La funzione principale svolta dagli organismi come i batteri è limitata alla trasformazione in materia vivente dell'energia derivata da sorgenti semplici. In relazione a ciò, i batteri presentano forme piuttosto semplici e uniformi. Gli eucarioti unicellulari, le cui dimensioni sono decisamente maggiori rispetto a quelle dei procarioti, possono invece mostrare comportamenti estremamente complicati come la predazione; di conseguenza, le loro forme possono essere molto complesse. Le cellule eucariotiche sono dotate di organi, quali ciglia sensoriali, fotorecettori, apparati boccali, e così via, che, a livello della singola cellula, sono foggiati da componenti del citoscheletro. Negli organismi pluricellulari le diverse funzioni essenziali per la sopravvivenza sono svolte da organi specifici, le cui cellule si sono differenziate in forme sofisticate. Anche in questo caso, tali forme sono determinate per la maggior parte da variazioni nell'organizzazione tridimensionale del citoscheletro.
I due principali componenti del citoscheletro implicati nei processi morfogenetici sono i microtubuli e i microfilamenti. I microtubuli, costituiti da tubulina, sono coinvolti in maniera prevalente nell'organizzazione della forma complessiva della cellula, nella direzione dei movimenti e nella localizzazione degli organelli nel citoplasma. Inoltre essi sono assemblati in strutture specifiche, quali ciglia vibratili e sensoriali e fuso mitotico. I microfilamenti, costituiti da actina, sono implicati, maggiormente, in tutti gli aspetti della motilità cellulare e nel trasferimento dell'informazione dall'esterno all'interno della cellula. Tanto i microfilamenti quanto i microtubuli sono polimeri che presentano una struttura dinamica, orientata nello spazio, responsabile delle loro proprietà morfogenetiche. La loro asimmetria (o polarità) è letta da motori molecolari, i quali sono responsabili del movimento degli organelli verso domini cellulari specifici. La comprensione dei principî che sono alla base dell'organizzazione tridimensionale delle fibre del citoscheletro può portare, almeno in parte, a valutare la diversità delle forme e delle funzioni cellulari degli organismi eucariotici. Il sistema del citoscheletro possiede proprietà di autoorganizzazione che contribuiscono a spiegare i meccanismi responsabili delle diverse forme e funzioni cellulari.
Forma e funzione
Per quanto in biologia sia comune correlare struttura e funzione, non va dimenticato tuttavia che questo concetto è prevalentemente limitato alla materia vivente. I componenti della materia vivente, infatti, hanno strutture sofisticate che consentono loro di svolgere attività specifiche. Il problema della forma cellulare e della sua relazione con la funzione pone immediatamente l'interrogativo su quale sia la funzione di una cellula. Fondamentalmente, ogni sistema vivente deve sopravvivere e riprodursi, cioè trasformare l'energia in un ordine 'stabile' ma dinamico. Negli organismi pluricellulari si sono differenziate, nel corso dell'evoluzione, molte cellule di tipo diverso che hanno specifiche funzioni essenziali alla sopravvivenza dell'intero organismo; tutte devono, però, in qualche modo assorbire energia e trasformarla in una forma specifica, indispensabile per compiere una particolare funzione. Tre diverse questioni riguardano la relazione tra forma e funzione. La prima ha a che vedere con l'evoluzione, in particolare con il modo in cui evolvono le forme e le modalità con cui la pressione selettiva agisce sulla loro evoluzione. La seconda questione riguarda il meccanismo responsabile di trasformare l'informazione lineare, immagazzinata nel genoma, in strutture a tre o persino a quattro dimensioni. Infine, rimane da chiarire il modo in cui forma e funzione siano legate a livello cellulare.
L'evoluzione delle forme cellulari deve in qualche modo essere determinata dalle variazioni nella natura e nella combinazione dei prodotti dei geni coinvolti nella determinazione delle forme. Anche la sequenza temporale dell'espressione di questi geni multipli è certamente importante. Alcune proteine hanno un ruolo morfogenetico diretto in quanto sono gli elementi costitutivi del citoscheletro; altre proteine hanno, anch'esse, una funzione morfogenetica, ma il loro ruolo non è così ovvio, poiché si tratta di enzimi che controllano l'organizzazione tridimensionale del citoscheletro. L'evoluzione delle forme cellulari è, quindi, correlata in maniera diretta alla coevoluzione di un elevato numero di geni che codificano sia proteine strutturali sia enzimi. Si tratta, pertanto, dell'evoluzione di una rete di varie entità molecolari correlate tra loro. Iniziamo a comprendere come le proprietà fisiche rendano il citoscheletro capace di generare la molteplicità delle forme cellulari conosciute. Al contrario, il ruolo svolto dall'organizzazione della membrana cellulare nella morfogenesi, per quanto molto importante, non è stato ancora completamente chiarito, sebbene, anche in questo campo, si stiano compiendo rapidi e significativi progressi.
Cercheremo di illustrare, a questo punto, come le molecole responsabili della forma cellulare possano essere coinvolte contemporaneamente in più di una funzione cellulare specifica. Sarà allora chiaro che il dogma 'un gene, una proteina, una funzione' non è valido. Una singola proteina può partecipare a molte funzioni cellulari diverse; viceversa, il comportamento complessivo dei prodotti di molti geni è all'origine di una funzione cellulare specifica. L'interazione di più proteine del citoscheletro, ciascuna con le proprie specifiche proprietà fisiche, dà luogo a forme dotate di una funzione particolare.
La diversità delle forme e delle funzioni cellulari
I procarioti presentano forme molto semplici, da sferiche ad allungate, comunemente di dimensioni lineari dell'ordine di qualche micron. Di regola hanno un rivestimento protettivo (una parete cellulare) all'interno del quale una membrana plasmatica circonda un unico compartimento citoplasmatico contenente DNA, RNA, proteine e piccole molecole. Le cellule procariotiche possono derivare la loro energia da sorgenti molto semplici come il glucosio, trasformarla in ATP e quindi utilizzare gli atomi di carbonio per sintetizzare tutti gli enzimi e le altre molecole organiche di cui hanno bisogno. Gli eucarioti unicellulari hanno forme molto più complesse. Una delle maggiori differenze tra procarioti ed eucarioti è la fonte di energia da cui attingono: mentre i procarioti utilizzano fonti molto semplici, gli eucarioti si riforniscono da altri organismi viventi o direttamente dalla luce e dalle molecole presenti nel suolo. Di conseguenza, dimensioni e forme variano notevolmente; le dimensioni degli eucarioti sono, infatti, decisamente maggiori rispetto a quelle dei procarioti (generalmente più di 10 μm). Gli eucarioti hanno diversi compartimenti subcellulari; il DNA è racchiuso in un nucleo e il citoplasma contiene un'elaborata rete di membrane coinvolte negli scambi di sostanze con l'ambiente circostante tramite i processi di secrezione e di assorbimento. L'energia è prodotta dai mitocondri e, negli organismi fotosintetici, dai cloroplasti. Tali organelli contengono DNA e si pensa si siano originati come batteri simbiotici.
Molto interessante è il fatto che il citoplasma delle cellule eucariotiche è organizzato dal citoscheletro, composto di filamenti di actina, filamenti intermedi e microtubuli. I filamenti intermedi agiscono come un supporto strutturale per il citoplasma, mentre i microtubuli organizzano il citoplasma e servono da cavi lungo i quali si verifica il trasporto di sostanze diverse all'interno della cellula; i filamenti di actina sono generalmente interessati alla motilità e alla forma cellulare, insieme ai microtubuli. I protisti sono eucarioti che presentano un'anatomia complessa che include strutture quali ciglia sensoriali, flagelli, fotorecettori, apparati boccali e fasci contrattili di tipo muscolare. Essi mostrano, inoltre, comportamenti diversi: alcuni sono carnivori, altri si basano sulla fotosintesi; possono essere mobili o sedentari e la loro forma è adattata di conseguenza. Gli organismi mobili hanno ciglia o flagelli, quelli sedentari hanno un piede per ancorarsi alla superficie e una bocca circondata da ciglia per catturare la preda di passaggio. In questo modo, ciascuna forma cellulare determina un comportamento specifico che comprende numerose funzioni. Esistono differenti modalità per muoversi o per essere sedentari e le forme cambiano in conformità con esse; ciascuna forma, però, potrebbe essere leggermente diversa senza necessariamente compromettere la vita dell'organismo. Tutto ciò suggerisce che nel corso dell'evoluzione compaiono quelle forme che indirizzano le cellule verso comportamenti particolari.
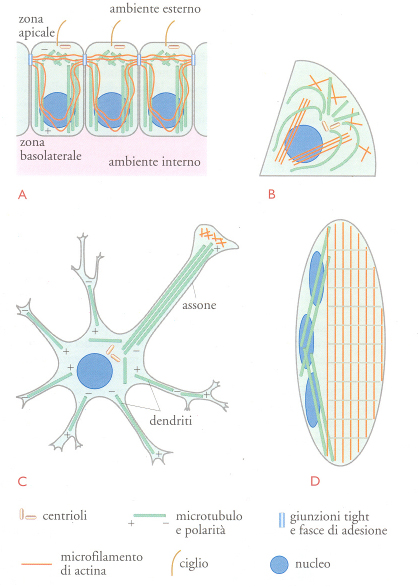
La multicellularità permette l'assemblaggio di organismi di grandi dimensioni che, almeno nel regno animale, devono nutrirsi di altre forme di vita. Di conseguenza, l'alimentazione, l'elaborazione del cibo, la riproduzione, il movimento e i comportamenti complessi dipendono dalla formazione di particolari organi costituiti da tipi cellulari diversi (fig. 2), con forme e funzioni specifiche. Le cellule epiteliali tappezzano le cavità corporee e costituiscono la pelle, interfaccia fra un organismo pluricellulare e il mondo esterno. Sono cellule di tipo epiteliale quelle del tubo digerente, dei reni, delle ghiandole di secrezione e quelle degli organi di senso; sono tutte polarizzate, con una zona apicale rivolta verso l'ambiente esterno e una zona basolaterale rivolta verso quello interno (fig. 2A). Questa asimmetria è essenziale per la loro funzione di scambio di materia o di informazione con l'ambiente circostante. Tali cellule sono strettamente legate l'una all'altra, condizione, anche questa, indispensabile per la loro funzione di barriera. Le cellule sensoriali hanno sviluppato strutture speciali, come le ciglia o le complesse reti di membrana, adatte al tipo di segnale fisico o chimico che devono individuare. Anche le cellule nervose, i neuroni, sono di origine epiteliale ma presentano numerosi prolungamenti distinti in due categorie: gli assoni e i dendriti. Recentemente è stato dimostrato che l'assone corrisponde alla zona apicale delle cellule epiteliali, mentre i dendriti sono più simili a quella basolaterale. È chiaro che l'organizzazione di queste cellule è adeguata alla loro funzione, che è quella di generare reti interconnesse che immagazzinano ed elaborano le informazioni.
I linfociti, i macrofagi e i fibroblasti sono cellule in grado di circolare liberamente e di spostarsi con movimenti attivi strisciando su altri tessuti. La forma sferica dei linfociti si adatta bene alla loro funzione di secernere anticorpi o di riconoscere agenti estranei da neutralizzare tramite la secrezione di sostanze tossiche. I macrofagi e i fibroblasti hanno forme allungate e sono equipaggiati per migrare nei vari substrati, i primi per cercare corpi estranei da distruggere attraverso la fagocitosi, i secondi per rimarginare lesioni nei tessuti.
Le cellule muscolari costituiscono uno straordinario esempio di differenziazione cellulare e di morfogenesi. Cellule chiamate mioblasti si fondono per formare i miotubi multinucleati; successivamente, si formano le miofibrille all'interno delle quali actina e miosina si organizzano nei sarcomeri, strutture capaci di contrarsi in risposta a un impulso nervoso. In questo caso, l'interrelazione tra forma e funzione è particolarmente evidente e illustra in modo chiaro come le proprietà fisiche di un particolare gruppo di proteine possano determinare una data forma e come questa sia correlata alla funzione cellulare.
Infine, consideriamo la relazione tra forma e funzione nei gameti: sia quello maschile sia quello femminile sembrano avere la stessa funzione, ossia fondersi per dare origine a un nuovo organismo; tuttavia, sono caratterizzati da forme del tutto diverse. Ancora una volta, il concetto di funzione è ambiguo. Nelle rane, per esempio, l'uovo consiste in una cellula sferica molto grande che contiene tutte le sostanze necessarie alla formazione di 20.000 cellule. La cellula uovo, pertanto, racchiude in sé il potenziale per sviluppare un nuovo organismo, svolgendo, al tempo stesso, la funzione di magazzino. Lo spermatozoo, invece, con il suo piccolo compartimento citoplasmatico e la grande coda mobile, è adatto a nuotare e a portare il genoma maschile all'interno dell'uovo. Per quanto la funzione globale dei gameti sia la stessa, il loro obiettivo specifico, nel breve termine, è diverso e la loro forma è in evidente relazione con la loro funzione immediata.
È interessante notare come la maggior parte delle funzioni svolte da cellule specifiche negli organismi pluricellulari, come l'endocitosi, le funzioni sensoriali, la motilità actino-dipendente e la capacità di prendere decisioni, sia già presente negli eucarioti unicellulari; infatti, ognuna di queste funzioni coinvolge proteine e strutture subcellulari specifiche. Per esempio, le stesse proteine, actina e miosina, sono implicate tanto nella motilità degli organismi unicellulari, quanto nella formazione delle cellule muscolari degli eucarioti pluricellulari. Il medesimo assemblaggio supramolecolare, basato sulla tubulina, interviene sia nella formazione delle ciglia dei protozoi, sia in quella del flagello dello spermatozoo. Sembra, dunque, che nel corso dell'evoluzione un gruppo ristretto di proteine sia stato utilizzato per assolvere a funzioni cellulari specifiche e che, insieme ad altre proteine regolatrici, determini sia la forma sia la funzione dei diversi tipi cellulari che si possono osservare attualmente in natura. Nonostante sia composto di proteine estremamente simili tra loro e dotate di una struttura che si è conservata nel corso dell'evoluzione, il citoscheletro può assumere una molteplicità di forme, ciascuna delle quali dà origine a funzioni specifiche.
Diversificazione delle forme e delle funzioni cellulari durante lo sviluppo
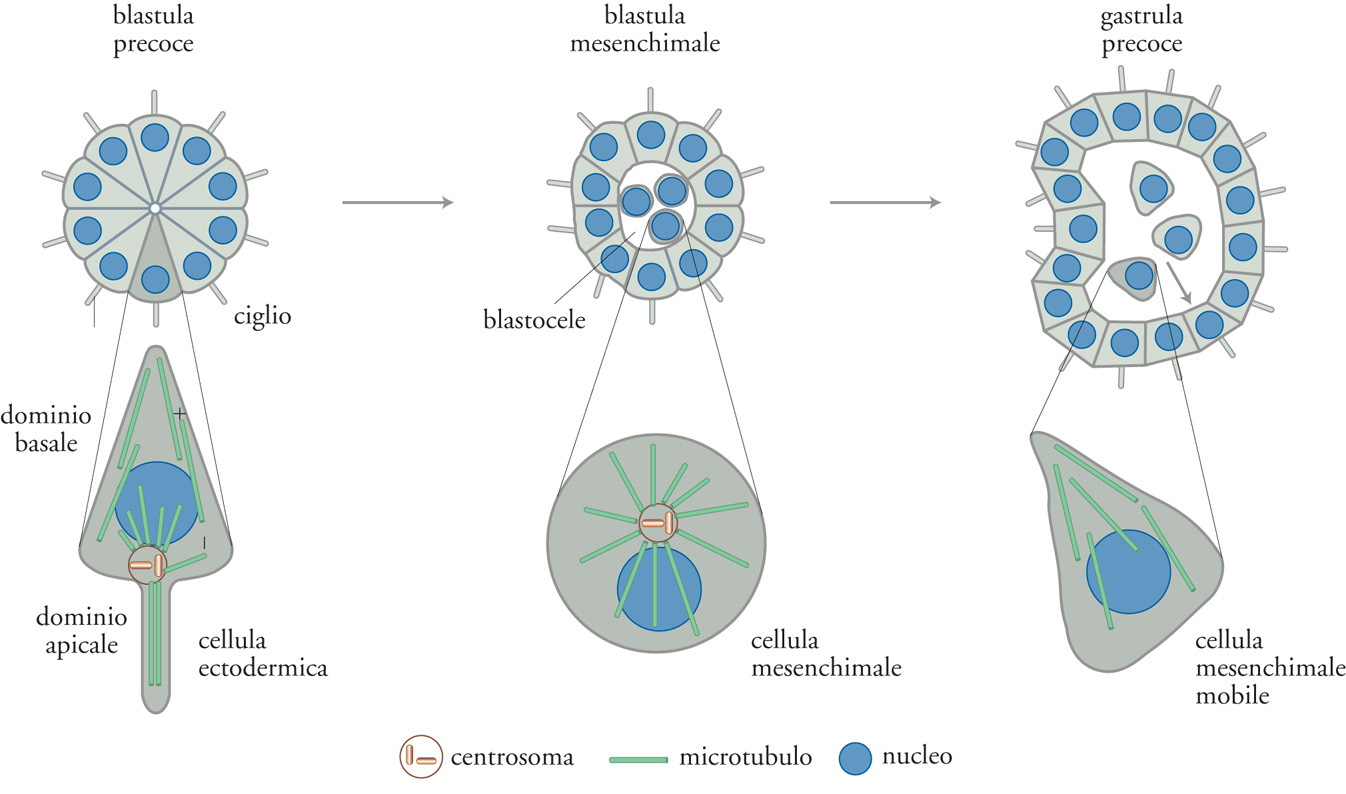
Nel corso dell'embriogenesi, le prime cellule che si differenziano sono quelle presenti sulla superficie dell'embrione allo stadio di blastula; in tutti gli embrioni, dalla drosofila all'uomo, queste cellule diventano cellule epiteliali polarizzate (fig. 3). Nella fase di gastrula, l'epitelio si introflette per formare il futuro endoderma. Alcune cellule endodermiche migrano nella cavità che si è formata tra i due strati cellulari, dove costituiranno il futuro mesoderma. L'ectoderma dà origine all'epidermide, al sistema nervoso centrale e agli organi di senso, mentre l'endoderma all'abbozzo del tubo digerente. I polmoni e il fegato si formano dalla parete dell'abbozzo del canale digerente che, nei primi stadi dello sviluppo, consiste in un tubo molto semplice che unisce la futura bocca con il futuro ano. Tutti gli elementi cellulari descritti, neuroni compresi, mostrano la stessa organizzazione ad asimmetria apico-basale e tutti traggono origine dalla superficie dell'embrione, cioè nell'interfaccia fra l'ambiente esterno e quello interno. È quindi verosimile che l'asimmetria apico-basale di queste cellule abbia origine nell'asimmetria fisica presente sulla superficie della blastula. Tale asimmetria è, ovviamente, mantenuta nelle cellule epidermiche e in quelle endodermiche che si affacciano sempre su due ambienti diversi e che costituiscono le superfici esterna e interna dell'embrione.
A differenza delle cellule dell'ectoderma e dell'endoderma, le cellule che dall'endoderma migrano verso il mezzo interno perdono l'asimmetria apico-basale, probabilmente perché sono immerse in un ambiente omogeneo. Tali cellule costituiscono il mesoderma, a partire dal quale si formano il mesenchima, le ossa, i muscoli, il cuore e le cellule del sangue, come pure le cellule del tratto urogenitale. Sebbene la maggior parte di queste cellule perda la tipica organizzazione apico-basale delle cellule epiteliali, alcune la riacquistano, come nella morfogenesi del rene, attraverso interazioni con le cellule epiteliali stesse.
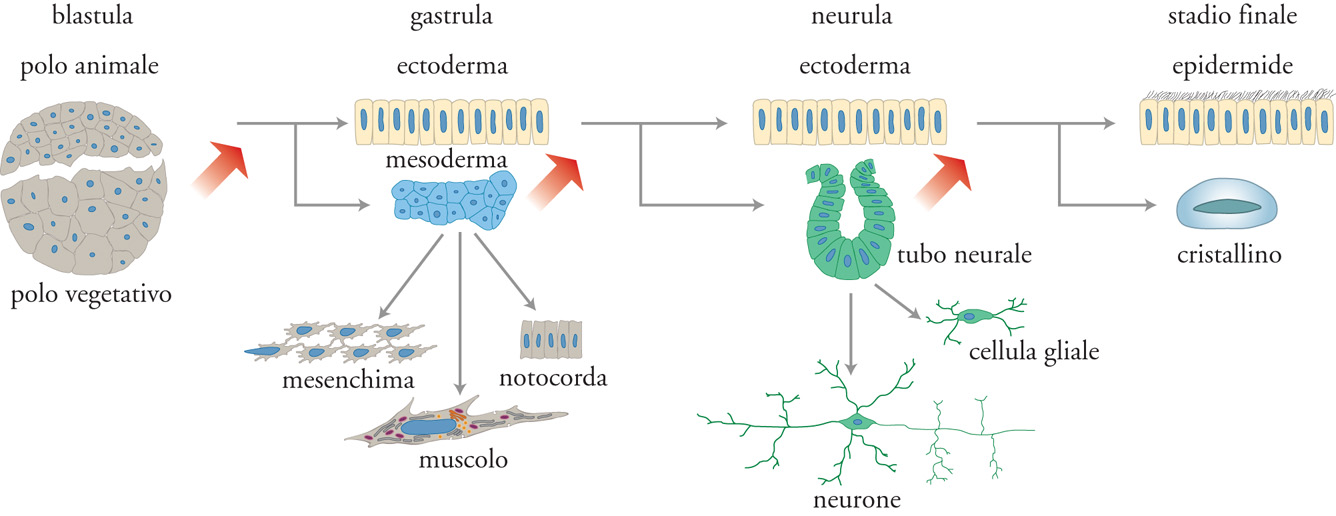
Tutti gli organi adulti, probabilmente in tutti gli organismi, si formano attraverso interazioni tra le cellule di origine ecto-endodermica e quelle di origine mesodermica, come mostra la fig. 4. Ciò indica che nell'embriogenesi la morfogenesi si realizza attraverso una progressiva amplificazione della varietà di forme cellulari. In altre parole, l'originale asimmetria dell'embrione sferico è essenziale per la formazione di tutti i tipi cellulari presenti nell'organismo, attraverso la produzione del primo tipo di cellule differenziate, quelle epiteliali. Queste cellule, pompando ioni nel blastocele, creano un ambiente interno omogeneo in cui nuove cellule possono differenziarsi, attraverso interazioni reciproche, restando isolate dall'ambiente esterno. Quando queste cellule acquisiscono un'asimmetria, essa è dovuta a segnali quali la presenza di altre cellule, di fattori di crescita o di gradienti di morfogenesi, più che alla differenza tra ambiente esterno e interno.
La differenziazione cellulare, il meccanismo attraverso il quale le cellule assumono forme e funzioni diverse nel corso dell'organogenesi, è ancora lontana dall'essere compresa. È chiaro tuttavia che le cellule si differenziano non perché cambiano i loro genomi, ma perché varia la combinazione dei geni espressi. Durante lo sviluppo, la differenziazione è il risultato di un controllo regionale dell'espressione genica, raggiunto, in parte, tramite la localizzazione eterogenea dei determinanti citoplasmatici nell'uovo, prima della gastrulazione, e attraverso processi di induzione che coinvolgono le interazioni fra cellule, come descritto sopra. Il contributo relativo di ciascun meccanismo varia da specie a specie. Nei Mammiferi, il citoplasma di tutti i blastomeri è identico all'inizio dello sviluppo e la differenziazione cellulare dipende prevalentemente da processi di induzione. Nelle ascidie, invece, il citoplasma di ciascun blastomero è diverso e determina il tipo di cellula cui darà origine; si parla in questo caso di embrione a mosaico. Nelle rane ci si trova di fronte a una situazione intermedia, in cui i determinanti vengono distribuiti asimmetricamente tramite movimenti citoplasmatici, durante il primo ciclo cellulare che segue la fecondazione, allo stadio di singola cellula, con il risultato che si formano due tipi di blastomeri: da una parte, quelli che, nel polo animale dell'embrione, danno origine alle cellule ectodermiche e, dall'altra, quelli che, nel polo vegetativo, danno origine alle cellule endodermiche. Le cellule mesodermiche vengono prodotte successivamente, tramite l'induzione delle cellule ectodermiche che, a loro volta, interagiscono con quelle endodermiche. Dopo la gastrulazione, nuove interazioni provocano un'ulteriore induzione che porta alla formazione di tutti i tipi cellulari, secondo quanto mostrato nella fig. 4.
Al di là di questi percorsi apparentemente differenti, sarebbe interessante capire se esistono principî fondamentali comuni sui quali si basa la generazione della diversità cellulare in tutti i phyla. In precedenza si è visto come l'asimmetria iniziale sia essenziale per generare diversità. Il problema è, dunque, stabilire in che momento e in che modo essa si generi. Negli embrioni a mosaico, l'asimmetria si forma a causa della localizzazione di determinanti nell'ovocita e si pensa sia mediata dal citoscheletro. Alcuni esperimenti hanno recentemente dimostrato che nell'ovocita della drosofila determinanti sotto forma di RNAm che codificano proteine specifiche, e forse le stesse proteine specifiche, sono localizzati tramite il citoscheletro. L'eterogeneità citoplasmatica, così generata, determina la formazione di cellule differenti che, nel corso della segmentazione, presentano un contenuto citoplasmatico diverso. Resta da capire come il citoscheletro dell'ovocita sia organizzato per posizionare i determinanti dei successivi processi di sviluppo. Esperimenti recenti hanno dimostrato in maniera elegante che l'obiettivo è raggiunto tramite la localizzazione di molecole segnale che sono posizionate in seguito all'interazione tra il citoplasma dell'ovocita e le cellule follicolari che lo circondano. Persino negli embrioni a mosaico, in ultima analisi, sono le interazioni cellulari a stabilire la localizzazione dei determinanti che governano il destino delle cellule embrionali nel futuro adulto.

In Xenopus si verifica una situazione simile, in quanto alcuni determinanti sono localizzati nel polo animale e altri nel polo vegetativo dell'ovocita. Tale disposizione è frutto del posizionamento dei microtubuli nel corso dell'ovogenesi. In questo caso, si determina una seconda asimmetria in funzione del punto di ingresso del gamete maschile e, probabilmente, dell'orientamento dei microtubuli in formazione a partire dal centrosoma associato al gamete maschile. Questa seconda asimmetria è all'origine dell'asse dorso-ventrale dell'embrione. Nei Mammiferi tutte le cellule dell'embrione precoce sono equivalenti fino allo stadio di otto cellule. A partire da questo stadio, ciascuna cellula si polarizza in seguito all'interazione con le cellule circostanti. Come conseguenza si verifica una divisione asimmetrica nel ciclo cellulare successivo; ogni cellula, all'interno dell'embrione, produce una cellula figlia completamente isolata dall'ambiente esterno (fig. 5). Queste danno origine alla massa cellulare interna e, in seguito, all'individuo adulto, mentre le cellule superficiali vanno a costituire il trofoectoderma e, successivamente, la placenta. Dalla massa cellulare interna, le cellule si differenziano completamente, attraverso continui processi di induzione che seguono la gastrulazione.
Da questa panoramica sulla possibile origine della diversità cellulare, nel corso dell'embriogenesi, si ricava che in tutti gli organismi la differenziazione cellulare viene indotta da un'asimmetria iniziale. Dal momento che tutti gli embrioni assumono, in un primo tempo, la forma di una sfera o di un ovoide, l'origine più ovvia dell'asimmetria iniziale è la superficie stessa dell'embrione. Negli embrioni a mosaico, come in drosofila, è l'interazione tra la superficie dell'ovocita e le cellule follicolari a provocare l'asimmetria iniziale essenziale per la successiva differenziazione cellulare, mentre nei Mammiferi l'asimmetria emerge più tardi; i principî di base non sono, comunque, tanto diversi. In tutti i casi vengono prodotti tre strati di cellule: l'ectoderma, l'endoderma e il mesoderma; dalle interazioni fra questi tre strati e per successiva induzione si differenziano i tipi cellulari.
Per comprendere come si generi la diversità cellulare nel corso dello sviluppo, bisogna risolvere tre importanti problemi. Il primo riguarda la relazione tra l'asimmetria cellulare e la differenziazione; il secondo, i meccanismi molecolari che legano le interazioni cellulari all'induzione e come, durante questo processo, l'espressione genica venga modificata in modo stabile; il terzo riguarda il meccanismo attraverso il quale l'espressione di una data combinazione di geni, come risultato della differenziazione, può condurre alla generazione di forme e funzioni cellulari specifiche. Siamo ancora lontani dalla comprensione dettagliata di questi processi; tuttavia, sembra che l'asimmetria cellulare venga provocata da eventi che si verificano sulla superficie cellulare e che comportano riarrangiamenti del citoscheletro, determinando, in tal modo, la localizzazione asimmetrica delle molecole coinvolte nella regolazione dell'espressione genica.
Resta da capire come un cambiamento dell'espressione genica sia reso stabile, così che le generazioni successive di cellule rimangano differenziate; tuttavia i modelli teorici stanno fornendo indicazioni al riguardo. Gli eventi ascrivibili alla superficie cellulare e responsabili della differenziazione potrebbero essere di natura fisica, come nella fase di compattazione degli embrioni precoci di mammifero o durante la formazione delle prime cellule epiteliali degli anfibi, oppure mediati da fattori, come quelli di crescita, che si legano a recettori. Quest'ultimo meccanismo si verifica, in genere, durante i processi di induzione tra le cellule generate da uno dei tre strati germinativi. La differenziazione cellulare è il risultato dell'espressione di una combinazione specifica di geni. Particolari isoforme proteiche del citoscheletro e proteine di regolazione porterebbero, insieme alle interazioni cellulari, alla formazione degli organi. Finora le conoscenze a riguardo sono piuttosto limitate, anche se si iniziano a comprendere le modalità con cui si differenziano forme cellulari specifiche e con cui la forma si lega alla relativa funzione. Una cosa è chiara: lo sviluppo di un organismo è un processo di autoorganizzazione in cui eventi locali provocano modificazioni locali nel comportamento delle cellule e nell'espressione genica che, in qualche modo, si stabilizzano e si propagano fino al raggiungimento di una situazione di equilibrio. Le proteine del citoscheletro mostrano proprietà fisiche sorprendenti che conferiscono loro potenzialità morfogenetiche e, in particolare, la capacità di attuare processi di autoorganizzazione. Il citoscheletro è infatti il più grande architetto nel generare forme cellulari specifiche. Uno dei principali problemi da affrontare riguarda il modo in cui si determinano le nuove forme cellulari attraverso modificazioni nella combinazione dei geni che vengono espressi in modo stabile.
Proprietà morfogenetiche del citoscheletro e ruolo dei segnali esterni nella morfogenesi cellulare
Si tratteranno qui, in primo luogo, le proprietà fondamentali dei microfilamenti e dei microtubuli; successivamente, verrà descritto il ruolo di questi due polimeri nella morfogenesi cellulare. Si inizierà dalla descrizione delle modalità con cui i microtubuli formano il fuso mitotico, processo che segue un percorso basato su principî morfogenetici generali, per poi passare a descrivere la morfogenesi delle cellule epiteliali e illustrare le generalità di alcuni principî morfogenetici dell'assemblaggio del fuso mitotico. Anche le cellule nervose e quelle muscolari sono molto interessanti perché rappresentano due casi estremi. Nei neuroni, infatti, il sistema dei microtubuli è molto sviluppato e gioca un ruolo fondamentale nell'organizzazione e nella funzione neuronale. Inoltre, si riscontrano alcune analogie tra lo sviluppo del cono di crescita dei neuroni e il modo in cui si stabilizza il dominio apicale delle cellule epiteliali. Nelle cellule muscolari, invece, prevale il sistema actomiosinico, nonostante i microtubuli sembrino rivestire un ruolo essenziale nella morfogenesi delle miofibrille.
Nelle cellule eucariotiche sono ben delineati tre sistemi del citoscheletro: i microfilamenti, i filamenti intermedi e i microtubuli. La funzione dei filamenti intermedi nel generare e mantenere la forma cellulare non è ancora stata del tutto chiarita. Al contrario, i microfilamenti e i microtubuli hanno proprietà fisiche che conferiscono loro una notevole potenzialità morfogenetica: entrambi sono sistemi fibrosi altamente dinamici, sono polimeri orientati e interagiscono con motori di locomozione che possono sia disporre i filamenti nello spazio, sia spostare carichi lungo di essi. Vengono qui presi in considerazione solo questi due sistemi filamentosi.
I microfilamenti


I microfilamenti sono costituiti da subunità monomeriche di actina, molecola caratterizzata da una struttura asimmetrica che conferisce polarità al filamento stesso (fig. 6). Ogni microfilamento è costituito da due protofilamenti di actina avvolti a elica. L'actina contiene un sito di legame per l'ATP, in corrispondenza del quale tale molecola è idrolizzata ad ADP durante l'assemblaggio; l'idrolisi serve unicamente per generare un certo grado di instabilità nel polimero. La polarità del polimero comporta due conseguenze: l'assemblaggio avviene preferenzialmente a un'estremità dei filamenti e i microfilamenti possono essere orientati all'interno della cellula. Esiste un unico tipo di motore in grado di spostarsi lungo di essi: la molecola di miosina, della quale esistono diversi tipi. Questa proteina si muove lungo i microfilamenti verso le loro estremità positive. In tal modo si verifica anche la contrazione muscolare, perché l'estremità positiva dei filamenti sottili di actina è localizzata a livello delle linee Z e i filamenti spessi di miosina sono bipolari (fig. 6). Oltre alla miosina della muscolatura striata, ne esistono molti altri tipi (fig. 7), le cui diverse localizzazioni intracellulari sono determinate da porzioni della proteina distinte dal dominio motore vero e proprio.
Organizzazione dei microfilamenti da parte di proteine associate
Molte proteine possono interagire con l'actina e regolare, così, la lunghezza dei singoli filamenti e la loro organizzazione nello spazio. Alcune, come la tropomiosina, si legano per tutta la lunghezza dei filamenti di actina, mentre altre, come la α-actinina, si legano solo alle estremità positive. Queste molecole sono coinvolte nell'organizzazione dei filamenti di actina in strutture contrattili. La filamina, invece, fa da ponte tra i microfilamenti in un sistema a rete lassa, simile a quello presente nella corteccia cellulare, lo strato di citoplasma che si trova immediatamente sotto la membrana plasmatica, dove i filamenti di actina sono legati alla membrana plasmatica dalle molecole di spectrina. Da queste osservazioni risulta che l'organizzazione tridimensionale e la formazione delle reti dei microfilamenti sono determinate dall'espressione di una specifica combinazione di proteine a essi associate, diversa per le cellule muscolari, per i fibroblasti, per le cellule epiteliali e per quelle nervose.
L'actina può essere presente in differenti organizzazioni tridimensionali anche in una singola cellula. L'ambiente gioca, dunque, un ruolo decisamente importante nel determinare quale tipo di molecole interagisca con il sistema dei microfilamenti e ne influenzi l'assemblaggio. Occorre, inoltre, mettere in evidenza come l'interazione tra le molecole di miosina e i filamenti di actina nelle cellule non muscolari possa contribuire, oltre che a produrre forze, a ordinare i microfilamenti di actina in orientamenti specifici. Ciò è dovuto al fatto che la miosina si lega ai filamenti secondo un orientamento specifico e scivola lungo di essi in una determinata direzione; un meccanismo dello stesso tipo interviene nell'organizzazione tridimensionale dei microtubuli.
Nelle cellule, quindi, l'esistenza di polimeri lineari dinamici e orientati e di una serie di molecole che regolano la lunghezza dei filamenti nello stato di equilibrio, la loro rigidità, la loro capacità di formare legami trasversali e la loro organizzazione dà luogo a un intero repertorio di forme in grado di svolgere funzioni specifiche. Tali forme si originano dall'autoorganizzazione dei componenti molecolari, ossia esse emergono dall'interazione dinamica esistente tra le molecole, e raggiungono specifici stati di equilibrio mantenuti grazie a un costante consumo di energia proveniente dall'idrolisi di ATP. Inoltre, molte forme possono essere create a partire da un determinato insieme di componenti, in funzione della loro concentrazione e delle piccole variazioni nelle loro proprietà.
I microtubuli
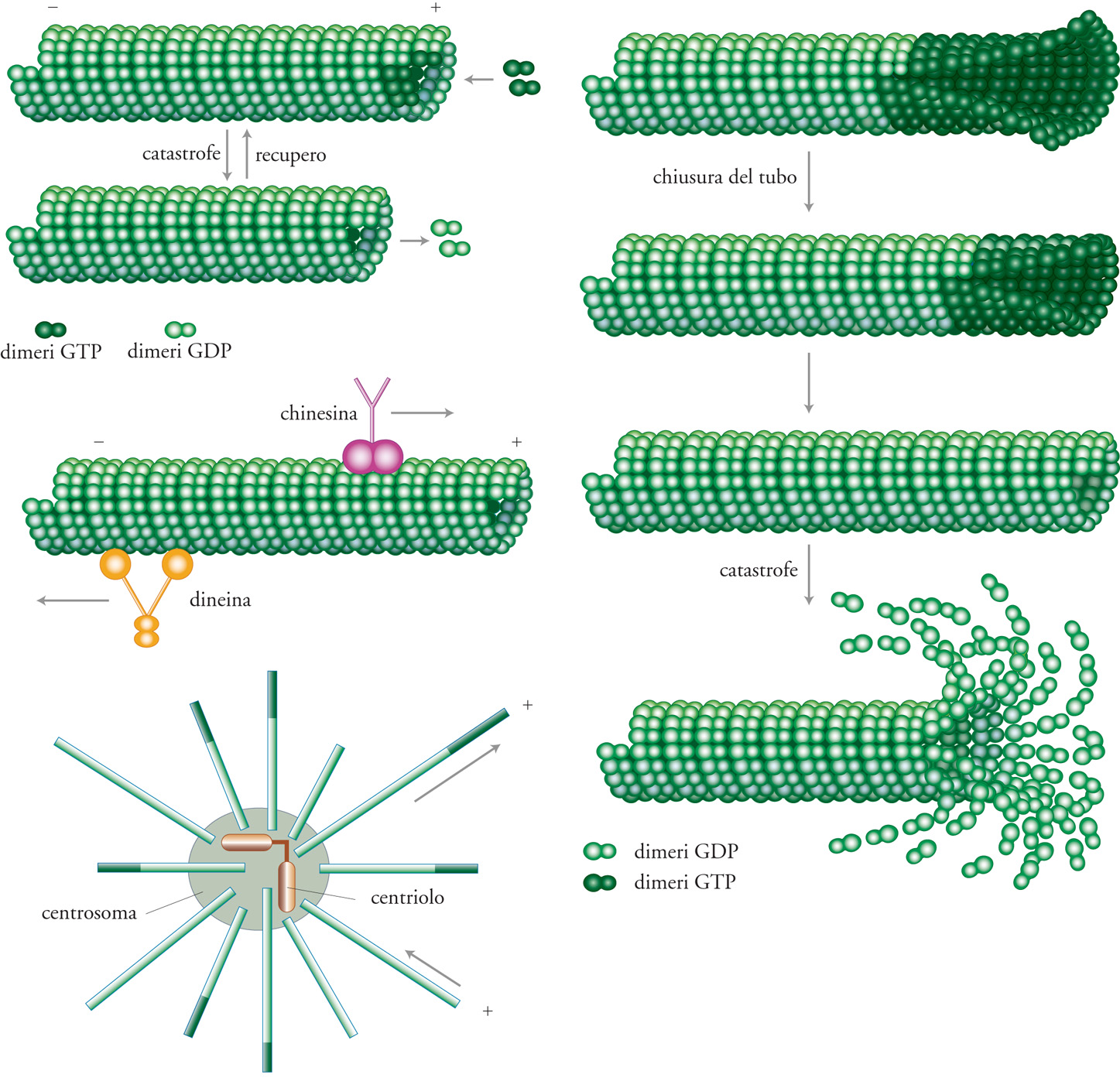
I microtubuli sono costituiti da dimeri di tubulina. La tubulina, come l'actina, ha una struttura asimmetrica, responsabile della polarità del polimero (fig. I.1). A differenza dei filamenti di actina, ciascun microtubulo è composto da un numero variabile di protofilamenti (in vivo generalmente tredici) disposti a formare un tubo cavo. Le molecole di tubulina sono dimeri, ciascun componente dei quali ha un sito di legame per il GTP. Uno dei due siti può scambiare il GTP e, durante l'assemblaggio, il GTP di questo sito viene idrolizzato a GDP. Come si verifica per l'actina, l'idrolisi non è richiesta per l'assemblaggio, ma per generare un certo grado di instabilità nel polimero. Il dimero di tubulina libero, con legato il GTP, mostra infatti un'affinità molto più grande per le subunità contenenti GTP rispetto ai dimeri contenenti GDP, situati all'estremità del polimero. Mentre, in assenza di proteine associate, i microfilamenti di actina sono altamente stabili, i microtubuli sono piuttosto instabili e mostrano un comportamento particolare, importante per le loro proprietà morfogenetiche, definito 'instabilità dinamica'.
Questa locuzione indica che, in certe condizioni, i microtubuli possono rapidamente oscillare tra fasi di crescita e di accorciamento (fig. I.2). Tale comportamento è importante perché può essere modulato da cambiamenti citoplasmatici locali. Si ritiene che i microtubuli diventino instabili una volta che, nelle subunità terminali del polimero, tutto il GTP è idrolizzato e sembra che ciò sia in relazione ai cambiamenti strutturali che si verificano nella parete dei microtubuli in crescita. L'estremità dinamica dei microtubuli è detta anche 'estremità positiva' (fig. I.1). L'instabilità dinamica del microtubulo è definita da quattro parametri misurabili con tecniche di videomicroscopia, che implicano l'uso di un microscopio munito di telecamera: la velocità di crescita, la velocità di riduzione, la frequenza di catastrofe (intesa come il passaggio dall'allungamento all'accorciamento) e la frequenza di recupero (inteso come il passaggio dall'accorciamento all'allungamento). Ciascun parametro può essere modulato indipendentemente e, di conseguenza, influire sul comportamento dei microtubuli.
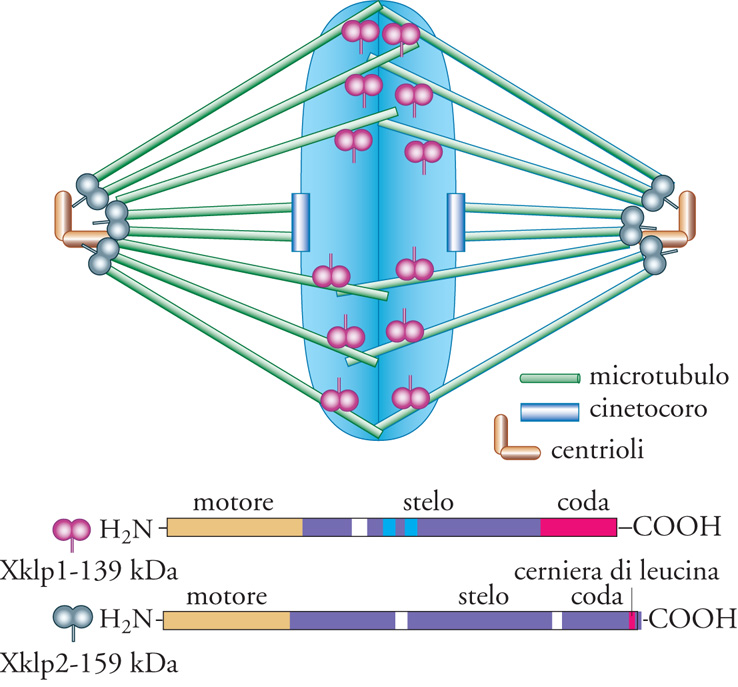
Lungo i microtubuli si muovono due tipi di motori: le proteine tipo dineina, che si spostano verso l'estremità negativa, e le proteine tipo chinesina, che si dirigono, salvo qualche eccezione, verso l'estremità positiva (fig. I.1). Entrambi i motori sono coinvolti in molti aspetti dell'organizzazione cellulare, dal trasporto delle vescicole alla membrana plasmatica o dei cromosomi durante la mitosi, all'organizzazione della disposizione dei microtubuli, sia durante l'interfase sia nel corso della mitosi. Questa varietà di funzioni è resa possibile dal fatto che, in aggiunta al dominio motore, operano domini bersaglio che indirizzano il motore verso carichi di tipo diverso. Esistono differenti gruppi di motori definiti per mezzo di sequenze specifiche presenti nel dominio motore, al di fuori delle regioni altamente conservate che costituiscono i domini leganti l'ATP e i microtubuli. Di solito i motori sono costituiti da un dominio globulare, unito a un peduncolo superavvolto, che consente la dimerizzazione. Il peduncolo è, a sua volta, connesso con una coda globulare che spesso contiene la sequenza che indirizza il motore verso il bersaglio. La fig. 8 mostra Xklp1 (Xenopus kinesin-like protein 1) e Xklp2 (Xenopus kinesin-like protein 2), due proteine tipo chinesina, indirizzate rispettivamente ai cromosomi e ai centrosomi. La Xklp1 è necessaria per il posizionamento dei cromosomi e per l'assemblaggio del fuso, mentre la Xklp2 è indispensabile per separare i poli del fuso durante la profase. La sequenza amminoacidica che determina la localizzazione della Xklp1 è tuttora oscura, mentre quella della Xklp2 è localizzata nella coda.
Organizzazione tridimensionale dei microtubuli
A differenza dei microfilamenti di actina, i microtubuli sono polimeri piuttosto rigidi. Mentre l'actina è utilizzata per modulare la consistenza del citoplasma nei processi contrattili e nel trasporto locale delle vescicole, i microtubuli sono coinvolti nell'organizzazione generale, cioè nella forma e nella polarità delle cellule. Lungo i microtubuli sono trasportati strutture e organelli diversi. Le modalità secondo cui i microtubuli sono organizzati tridimensionalmente e orientati nel citoplasma costituiscono eventi chiave nella morfogenesi cellulare. Un aspetto importante dell'organizzazione dei microtubuli in tutte le cellule riguarda il meccanismo tramite il quale essi sono orientati in modo coordinato. Si possono distinguere due modi secondo i quali i microtubuli possono essere orientati in arrangiamenti di polarità uniforme: essi sono organizzati da specifici centri di nucleazione, oppure per mezzo di motori che li smistano e li collegano in modo orientato.
Esistono diversi tipi di centri di nucleazione nelle cellule, ma negli eucarioti superiori i più comuni sono costituiti da un centriolo circondato da un addensamento di materiale in grado di organizzare i microtubuli (fig.I.1). I centrioli sono cilindri costituiti a loro volta da triplette di microtubuli. La loro vera funzione è ancora del tutto sconosciuta a eccezione del fatto che sono utilizzati come 'semi' per formare ciglia e flagelli. Dal momento che la crescita dei microtubuli è orientata con l'estremità positiva in testa, i centri di nucleazione formano astri di microtubuli con polarità omogenea, tutti con l'estremità negativa al centro e quella positiva in posizione periferica. Poiché i microtubuli possono essere soggetti a instabilità dinamica, la lunghezza di quelli organizzati da tali centri di nucleazione può variare da un valore virtualmente infinito, limitato soltanto dalla disponibilità di subunità libere di tubulina, a un valore stazionario, definito dallo stato dinamico della popolazione. Effettivamente, come già accennato, i parametri dell'instabilità dinamica dei microtubuli possono essere modulati, nella cellula, da fattori di regolazione tipo MAP (Microtubule associated proteins, proteine associate ai microtubuli) o da fattori destabilizzanti. La fosforilazione di tali fattori può inoltre regolare la loro attività; pertanto, si possono creare nella cellula condizioni in cui, come nel corso della mitosi, si formano astri brevi di microtubuli aventi una lunghezza media ben definita.
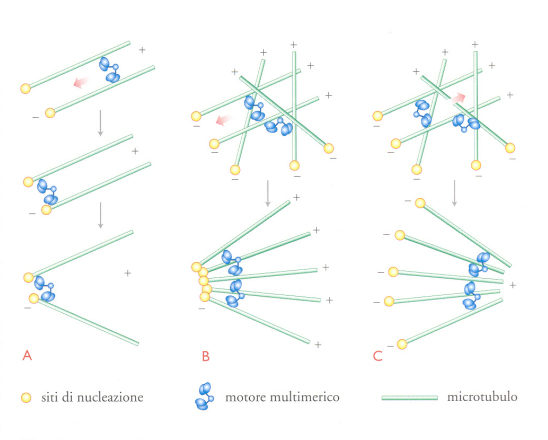
Secondo il meccanismo precedente, i microtubuli vengono organizzati in un astro a polarità uniforme dai centri di nucleazione e dalla regolazione della dinamica dei microtubuli. Effettivamente, per generare solo microtubuli ordinati dal centrosoma, le condizioni dovrebbero essere tali da escludere un assemblaggio spontaneo dei polimeri, in aggiunta a quello regolato dai centrosomi. Esiste un altro modo di produrre astri di microtubuli a polarità uniforme, in assenza di un preciso centro di nucleazione. Questo risultato può essere raggiunto tramite i motori (fig.9). Tale tipo di autoorganizzazione dei microtubuli in sistemi polarizzati, in assenza di centrosomi preformati, è stato osservato bene, durante l'interfase, in numerosi sistemi. Esperimenti condotti su estratti di uova mitotiche di Xenopus hanno anche dimostrato che microtubuli stabilizzati con un farmaco, il tassolo, si riorganizzano progressivamente, a partire da un'iniziale distribuzione casuale, in piccoli astri che contengono materiale centrosomico in posizione centrale. Poiché l'inattivazione del motore di dineina, diretto verso l'estremità negativa, blocca la formazione dell'astro, è evidente che i motori possono organizzare i microtubuli in astri. Come questo possa avvenire è illustrato nella fig.9. Se un motore plurisubunitario si lega a due microtubuli, si muove necessariamente fino alle loro estremità. Nel caso in cui più microtubuli siano coinvolti nel processo, viene costituito un astro a polarità uniforme. Mentre in vivo sono le estremità negative dei microtubuli a essere sempre raggruppate insieme al centro, in sistemi puri, composti da microtubuli e dal motore di chinesina diretto verso l'estremità positiva, è stata dimostrata la capacità dei motori unidirezionali di formare astri a polarità uniforme, in cui l'assemblaggio si verifica con le estremità positive dei microtubuli al centro. Tale meccanismo richiede che i motori interessati abbiano due proprietà: devono avere più teste motrici; non devono oltrepassare l'estremità negativa dei microtubuli o, almeno, ci deve essere un accumulo di motori o di fattori che si legano trasversalmente all'estremità negativa del microtubulo, in modo tale che lo stato di equilibrio osservato sia radiale e non si ottenga un fascio di fibre parallele.
Il fuso mitotico: un esempio di morfogenesi dovuta all'autoorganizzazione dei microtubuli
Il fuso mitotico costituisce l'apparato grazie al quale i cromosomi sono segregati nel corso della mitosi. La segregazione si verifica a causa della bipolarità del fuso, costituito da due gruppi di microtubuli di polarità opposta, disposti intorno ai cromosomi. Poiché i cromosomi sono attaccati tramite i cinetocori ai microtubuli, ognuno può essere trasportato da queste strutture proteiche che agiscono come funi in grado di guidarlo sino al polo verso il quale è rivolto e da cui trarrà origine la cellula figlia. Pertanto, la delucidazione dei principî in base ai quali i microtubuli si organizzano in un fuso bipolare può fornire un nuovo modello per la comprensione della morfogenesi cellulare in generale.
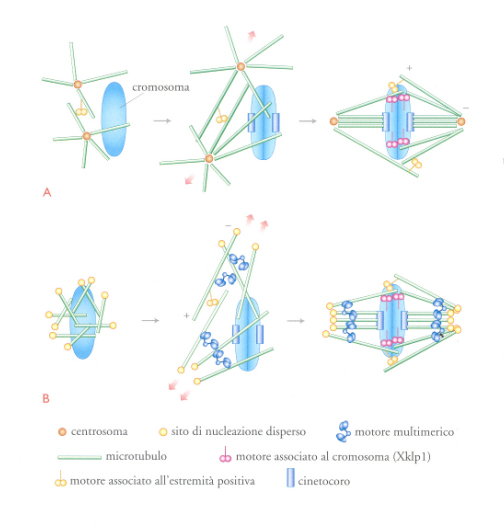
Le difficoltà incontrate nella comprensione del modo in cui si forma il fuso sono dovute al fatto che esso costituisce un apparato altamente dinamico: sebbene all'equilibrio possegga una lunghezza e una forma precise, i suoi microtubuli si rinnovano con un'emivita compresa tra pochi secondi e un minuto. Per lungo tempo si è ritenuto che l'orientamento uniforme dei microtubuli in ciascuna metà del fuso derivasse dalla loro uniforme polarità di crescita a partire dai centrosomi in direzione dei cromosomi. Effettivamente nelle cellule animali i centrosomi, una volta duplicati, migrano attorno al nucleo e dopo la disgregazione della membrana nucleare formano due astri di microtubuli che sembrano sondare lo spazio fino a che non vengono catturati dai cinetocori (fig.10A), secondo un processo denominato di 'ricerca e cattura'. Inoltre, i cromosomi esercitano un effetto locale sul citoplasma che dà luogo alla nucleazione e alla stabilizzazione dei microtubuli nelle loro vicinanze. Pertanto, nel fuso mitotico di una cellula animale esistono due popolazioni di microtubuli: quelli nucleati dai centrosomi che fanno parte degli astri presenti ai poli e quelli nucleati dai cromosomi che formano il fuso vero e proprio. L'effetto locale si esercita, in parte, attraverso due gradienti, uno dei quali è dovuto alla forma attivata di una piccola GTPasi denominata Ran e l'altro alla fosforilazione proteica. Entrambi favoriscono la dinamica e la nucleazione dei microtubuli intorno ai cromosomi. Il risultato è che i microtubuli si allungano preferenzialmente verso i cromosomi (fig. 10A) e vengono catturati dai cinetocori e dai motori presenti sui bracci dei cromosomi. Non appena i microtubuli dalle due metà del fuso entrano in contatto, si legano insieme attraverso interazioni antiparallele, che stabilizzano ulteriormente il fuso in una configurazione bipolare, con i poli disposti a 180° l'uno rispetto all'altro.
Nelle piante e nelle cellule in meiosi, i microtubuli vengono nucleati casualmente, in prossimità dei cromosomi, probabilmente da siti di nucleazione dispersi, non organizzati in un centrosoma (fig. 10B). Essi si allungano in prossimità dei cromosomi per effetto della stabilizzazione locale e vengono organizzati nei poli da motori di dineina orientati verso l'estremità negativa, secondo il meccanismo descritto nel paragrafo precedente. I microtubuli sono, poi, separati dai motori presenti sui bracci dei cromosomi, orientati verso l'estremità positiva, e da quelli che interagiscono con i microtubuli antiparalleli. Tali meccanismi separano queste strutture proteiche in due popolazioni antiparallele di polarità uniforme, generando un fuso bipolare, con i cromosomi localizzati all'equatore.
Secondo quanto appena esposto, sia i fusi meiotici sia quelli mitotici si formano secondo principî simili: la modulazione locale delle dinamiche dei microtubuli da parte dei cromosomi e la separazione dei microtubuli in una struttura bipolare da parte di motori che leggono la loro polarità. La presenza di centrosomi organizzati non è essenziale ai fini della bipolarità; questa è centrata e stabilizzata in prossimità dei cromosomi che creano nel citoplasma un'asimmetria locale. In un certo senso, il fuso mitotico si organizza all'interno di un 'campo morfogenetico', definito intorno ai cromosomi dalla localizzazione di enzimi che regolano l'attività delle proteine interessate alle dinamiche dei microtubuli, e quella dei motori, che muovono i microtubuli in modo coordinato. È probabile che principî simili guidino l'organizzazione del citoscheletro durante l'interfase, nel corso della diversificazione e della differenziazione cellulare. La differenza consiste nel fatto che altri enzimi, localizzati in particolari domini cellulari, controllano l'attività di altri motori e regolatori delle dinamiche dei microtubuli.
I microtubuli e le forme dei protisti
Le forme dei protisti sono determinate prevalentemente dall'arrangiamento dei microtubuli che intervengono anche nell'organizzazione delle ciglia sensoriali, di quelle vibratili e dei flagelli utilizzati per il movimento. Anche l'organizzazione della bocca è dovuta ai microtubuli. Considerando la formazione del fuso mitotico, si potrebbe pensare che i diversi arrangiamenti tridimensionali dei microtubuli siano determinati dalla coespressione di una combinazione di molecole che influiscono sulla dinamica, sulla posizione e sull'orientamento dei microtubuli stessi nella cellula, e che ne sia responsabile un particolare campo morfogenetico con enzimi localizzati. Occorre valutare l'origine dell'asimmetria intorno alla quale questo campo potrebbe essersi generato. È interessante notare come nei protisti l'arrangiamento dei microtubuli nello spazio non sia solo il risultato di un processo di autoassemblaggio basato sull'espressione di una combinazione di molecole nella stechiometria appropriata. È stata dimostrata, infatti, l'esistenza di una memoria strutturale che regola l'orientamento delle file di ciglia nella corteccia dei ciliati; in questo modo l'asimmetria viene trasmessa da una generazione alla successiva. Anche la determinazione degli assi antero-posteriore e dorso-ventrale di molti embrioni si basa, principalmente, sull'orientamento dei microtubuli e ciò è in parte determinato dai segnali trasmessi dalla madre all'ovocita. Proprio come i mitocondri e i cloroplasti, che negli eucarioti si sono diffusi in seguito a un evento simbiotico, la forma delle cellule può essere pertanto determinata, in parte, da proprietà che riguardano l'autoorganizzazione di molecole come la tubulina e, in parte, dalla trasmissione, da una generazione all'altra, di configurazioni specifiche, che, originatesi spontaneamente, si sono poi stabilizzate nel corso dell'evoluzione. Queste determinano un'asimmetria di tipo fisico intorno alla quale il citoscheletro può venire organizzato in modi specifici.
Dalla forma alla funzione: la dualità del citoscheletro
Quando si prendono in considerazione le modalità con cui si generano le forme a livello cellulare, appare chiaro che il legame tra forma e funzione è stabilito in larga misura dal citoscheletro. È anche evidente che la maggior parte delle funzioni correlate alla forma cellulare è eseguita da una specifica organizzazione del citoscheletro che, nello stesso tempo, determina la forma appropriata della cellula. Questa è la ragione per cui, per discutere del modo in cui forma e funzione sono correlate attraverso l'autoorganizzazione del citoscheletro, è interessante comparare il meccanismo di assemblaggio del fuso mitotico con quello che presiede all'organizzazione di forme cellulari così diverse quali quelle delle cellule epiteliali e muscolari. Senza dubbio, la differente organizzazione tridimensionale delle proteine del citoscheletro, nelle diverse cellule, dipende dall'espressione di gruppi eterogenei di proteine regolatrici, in grado di modulare le proprietà di autoorganizzazione dei microtubuli e dei microfilamenti. A loro volta, microtubuli e microfilamenti svolgono funzioni cellulari specifiche che dipendono dalla loro organizzazione tridimensionale. Si comincia quindi a comprendere come una specifica funzione sia strettamente legata alla forma della struttura cellulare che la svolge. Poiché le forme biologiche possono essere molto complesse, occorre scegliere un particolare aspetto della forma per collegarlo con una funzione specifica. Come sottolineato, l'asimmetria è la chiave che determina la forma e le funzioni cellulari. Nelle cellule essa viene definita principalmente dalla polarità dei microtubuli e dei microfilamenti. È questa la chiave che porta alla formazione di specifiche forme cellulari e all'organizzazione e distribuzione degli organelli nello spazio.
Il fuso mitotico
Si è visto come, durante la mitosi, i microtubuli vengano organizzati in una configurazione bipolare con una funzione evidente: distribuire i cromosomi negli opposti domini della cellula in divisione dai quali avranno origine le due cellule figlie. È interessante notare come la bipolarità del fuso sia generata proprio dai cromosomi, i quali determinano l'orientamento dei microtubuli tramite la modulazione locale delle dinamiche e della nucleazione di queste strutture proteiche e attraverso l'azione dei motori. L'apparato costruito in questo modo acquista la capacità di segregare i cromosomi e si origina, pertanto, dallo stesso oggetto che deve trasportare. Poiché il meccanismo di assemblaggio del fuso mitotico è ora sufficientemente conosciuto, siamo in grado di capire come il comportamento coordinato di una serie di componenti e molecole con intrinseche e specifiche proprietà fisiche possa produrre forme precise a loro volta in grado di adempiere una funzione specifica, come la segregazione dei cromosomi. Molti altri esempi riconducibili a questo fenomeno possono essere rinvenuti in diversi sistemi cellulari, di cui è possibile delineare i meccanismi tramite i quali vengono generate le varie forme cellulari e le funzioni a esse associate, sebbene essi non siano stati delucidati così bene come è avvenuto nel caso del fuso mitotico.
Le cellule epiteliali
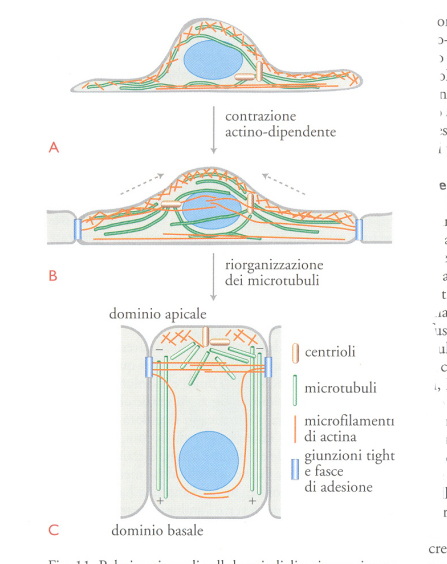
Per fronteggiare due ambienti molto diversi, le cellule epiteliali hanno la capacità di effettuare, tra questi, un trasporto regolato e orientato di fluidi e di soluti. Esse sono in grado di svolgere queste funzioni grazie alla divisione della loro membrana plasmatica in due domini, profondamente diversi per quanto riguarda la composizione lipidica e proteica. Tale divisione è ottenuta tramite le giunzioni tight tra cellule vicine, a livello delle membrane plasmatiche. Le giunzioni tight formano una barriera tra la superficie apicale, rivolta verso l'ambiente esterno, e la superficie basolaterale che poggia sulla membrana basale ed è bagnata dal liquido interstiziale. Nelle cellule epiteliali, i microtubuli mostrano un orientamento piuttosto uniforme con l'estremità positiva rivolta verso il dominio basolaterale, mentre alcuni microtubuli sono orientati in modo casuale nel dominio apicale (fig. 2A e fig. 11C). Consideriamo il modo in cui potrebbe verificarsi l'orientamento dei microtubuli (fig. 11). Quando le giunzioni si rompono, le cellule perdono la loro polarità; in queste condizioni i microtubuli assumono un'organizzazione assai diversa: si dispongono in modo radiale a partire da un punto poco definito, localizzato in prossimità del centro di nucleazione. Se le cellule sono in grado di ristabilire le giunzioni e polarizzarsi di nuovo, i microtubuli si riorganizzano e recuperano il loro orientamento originale (fig. 11A). Questo processo richiede il ristabilirsi delle giunzioni e la presenza dei microfilamenti di actina (fig. 11B).
Partendo dalle nostre conoscenze sui meccanismi molecolari, che sono alla base dell'organizzazione tridimensionale dei microtubuli, possiamo ipotizzare due modalità per il loro orientamento nelle cellule epiteliali. Nel primo, i centri di nucleazione potrebbero essere spostati dall'actina verso il dominio apicale delle cellule, orientando così la crescita di queste strutture proteiche (fig.11C). Nel secondo meccanismo potrebbero essere posizionati, nella corteccia cellulare a livello apicale, motori come la dineina orientati verso l'estremità negativa. In tal modo, le estremità positive dei microtubuli sarebbero spinte verso il dominio basolaterale delle cellule. Anche se mancano ancora prove sperimentali al riguardo, è assai probabile che questo secondo meccanismo si verifichi dal momento che nei neuroni la dineina corticale sembra interagire con i microtubuli per orientare il fuso mitotico e per trasportare queste strutture proteiche. Inoltre, secondo questa ipotesi, la dineina dovrebbe essere localizzata nella corteccia cellulare apicale e potrebbe essere legata ai filamenti corticali di actina.
Per l'origine dell'organizzazione delle cellule epiteliali, si potrebbe proporre, quindi, un modello sequenziale. In un primo momento le molecole di adesione cellulare inducono l'assemblaggio di una rete corticale di actina, all'interno della quale sono localizzati e ancorati i motori di dineina diretti verso l'estremità negativa dei microtubuli. Nel momento stesso in cui i filamenti di actina iniziano a contrarsi sotto l'azione della miosina, i microtubuli vengono spinti verso il basso e allineati dalla dineina corticale che si sposta verso la loro estremità negativa; i microtubuli sono, a loro volta, stabilizzati e resi piuttosto rigidi dalle MAP che li connettono tramite legami trasversali.
Le cellule epiteliali costituiscono un caso interessante, dal momento che la generazione della polarità richiede il trasporto orientato delle proteine di membrana lungo i microtubuli e questo trasporto costituisce anche la funzione principale dell'epitelio formato. Si stabilisce una specie di feedback positivo: segnali extracellulari comportano l'organizzazione dei microtubuli in un orientamento apico-basale che, a sua volta, determina il trasporto specifico delle proteine di membrana necessarie per stabilire la polarità di questo costituente cellulare; tale sistema continua poi a essere impiegato nel mantenimento dello stato stazionario. Come nel caso del fuso mitotico, un processo di autoorganizzazione è probabilmente all'origine di una funzione cellulare specifica.
Le cellule muscolari
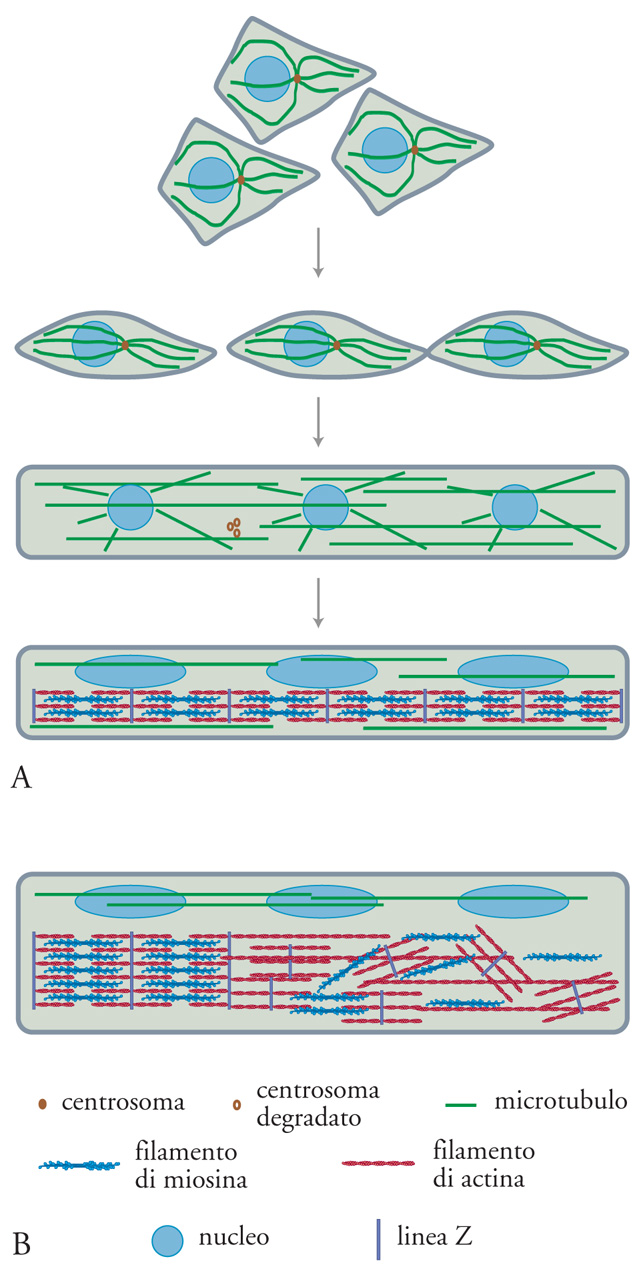
La morfogenesi delle fibre del muscolo scheletrico è particolarmente interessante in quanto consiste nell'allineamento e nella fusione di cellule singole, i mioblasti. Nell'embrione i mioblasti si differenziano in siti destinati a formare muscoli striati come i somiti, gli abbozzi di arti, ecc. I successivi eventi di fusione comportano la formazione di lunghi miotubi multinucleati. Prima che la fusione si verifichi, avviene l'ordinamento dei microtubuli a partire dai centri (fig. 12A), contenenti i centrioli circondati dal materiale di nucleazione dei microtubuli, localizzati vicino al nucleo del mioblasto. Subito dopo la fusione, i centrioli vengono distrutti e i microtubuli sembrano essere nucleati, almeno in parte, dalla membrana nucleare. In questa fase sono coinvolti due tipi di microtubuli: quelli che irradiano dai centri di nucleazione e una fitta popolazione di microtubuli allungati, paralleli all'asse longitudinale dei miotubi. Non è stata ancora chiarita l'organizzazione di tali microtubuli, per quanto riguarda la polarità e la lunghezza: è possibile che crescano a partire dalla membrana nucleare e che vengano organizzati da motori per tutta la lunghezza dei miotubi. Dopo la fusione, i nuclei vengono progressivamente rilocalizzati proprio sotto la membrana cellulare. A questo punto non si rileva alcuna organizzazione actomiosinica tipica delle miofibrille, anche se la maggior parte dei componenti necessari è in via di sintesi.
Le miofibrille sono formate da sarcomeri perfettamente allineati, costituiti da filamenti di actina antiparalleli, di lunghezza ben definita, e da filamenti bipolari di miosina. I primi, cioè i filamenti sottili, sono ancorati alla linea Z, costituita prevalentemente da α-actinina, che corrisponde alla linea di separazione tra un sarcomero e il successivo; i secondi, cioè i filamenti spessi, sono invece uniti a livello della linea M, o linea di mezzo del sarcomero. L'assemblaggio delle miofibrille apparentemente inizia non appena viene raggiunta, nel citoplasma, una concentrazione sufficiente delle proteine necessarie. Il problema è capire come vengano ordinati i filamenti sottili e quelli spessi, insieme alle proteine associate e a quelle di sostegno. Sappiamo che sia l'actina sia la miosina hanno la capacità di autoassemblarsi. In presenza di α-actinina e di altre proteine coinvolte nella sua formazione, probabilmente la linea Z inizia ad assemblare e a ordinare i microfilamenti di actina fino a che essi raggiungono la lunghezza tipica dello stato stazionario. Nello stesso tempo, si autoassemblano i filamenti spessi di miosina. Osservazioni strutturali, eseguite sia al microscopio elettronico sia con la tecnica dell'immunofluorescenza, suggeriscono una prima formazione di pre-miofibrille, in cui la spaziatura fra i sarcomeri è irregolare; esse sono rapidamente trasformate in miofibrille (fig. 12B).
Il problema è in che modo tutti questi componenti autoassemblati si allineino correttamente gli uni con gli altri. Una possibilità è che siano i numerosi microtubuli, presenti all'inizio della miogenesi, a garantire il corretto allineamento. In questo caso, alcuni dei componenti, come i filamenti spessi, potrebbero interagire con i microtubuli, eventualmente spostandosi lungo di essi, prima di essere posizionati opportunamente. Ciò implicherebbe il corretto orientamento dei microtubuli. Non è ancora noto come questo possa verificarsi, ma è chiaro che i microtubuli svolgono un ruolo essenziale nel dare la forma ai miotubi dopo la fusione dei mioblasti. L'ipotesi qui descritta è, al momento, tutta da verificare, ma potrebbe essere verificata facilmente con le tecniche attualmente disponibili. Una descrizione della miogenesi è stata riportata da Clara Franzini-Armstrong e Donald A. Fischman (1994). La contrazione muscolare è senza dubbio l'esempio più stupefacente del notevole potere morfogenetico della polarità dei polimeri. L'organizzazione antiparallela dei microfilamenti di ciascun sarcomero, unitamente all'organizzazione bipolare della molecola di miosina, dà luogo al più efficiente apparato in grado di produrre lavoro a partire da energia chimica. Anche in questo caso, le proprietà delle singole molecole che costituiscono tale apparato appaiono coinvolte nella sua stessa morfogenesi.
Si è dunque visto come forma e funzione cellulare siano correlate e come la forma possa determinare la funzione. Se si esamina il problema a livello molecolare, ci si rende conto di come l'organizzazione tridimensionale del citoscheletro determini, allo stesso tempo, una data forma e la funzione a essa associata. Nel fuso mitotico i cromosomi vengono separati dalla bipolarità del fuso; negli epiteli il trasporto orientato dei soluti dipende dall'orientamento dei microtubuli e nei muscoli la produzione di forza a livello macroscopico dipende dall'organizzazione specifica dell'actomiosina nei sarcomeri. Il vero problema è come si siano generate queste forme e come la differenziazione cellulare sia legata alla morfogenesi; in altri termini, quale sia il contributo dell'informazione genetica e quale quello dell'influenza della natura fisica dell'ambiente circostante sull'organizzazione tridimensionale delle proteine e dei lipidi. La funzione è il risultato dell'azione di entrambi questi fattori. È chiaro il contributo dell'informazione contenuta nelle proteine per l'organizzazione tridimensionale delle cellule: è, infatti, il folding (ripiegamento) delle proteine a generare molecole dotate di particolari proprietà fisiche, come gli enzimi e le proteine strutturali asimmetriche quali la tubulina e l'actina. Meno chiaro è il contributo dei fattori fisici dell'ambiente. Per esempio, l'organizzazione dei polimeri di actina e di tubulina dipende dalla loro forma allungata e dalle loro proprietà dinamiche che garantiscono potenzialità specifiche, quali l'instabilità dinamica e l'autoorganizzazione attraverso l'azione dei motori. Esistono due principî essenziali che controllano le forme attraverso il citoscheletro: la rottura della simmetria e l'autoorganizzazione; entrambi intervengono nei campi morfogenetici.
L'autoorganizzazione di polimeri, come i microfilamenti e i microtubuli, in arrangiamenti tridimensionali di forme diverse si basa, innanzi tutto, sulle loro proprietà dinamiche che non sono in equilibrio termodinamico e, secondariamente, sulla loro polarità. Sino a quando i polimeri si assemblano in un ambiente omogeneo, non può essere generata alcuna forma. Le forme possono essere generate solo se viene introdotta un'asimmetria relativa alle loro proprietà dinamiche o se i motori legati in modo asimmetrico alla cellula agganciano e caricano polimeri. La forma dipende ovviamente dalla natura e dalla combinazione dei fattori presenti nella cellula in un determinato momento. È in questo modo che i microtubuli possono essere organizzati diversamente durante la mitosi, nelle cellule epiteliali o nei miotubi, nel corso della miogenesi. Questa combinazione di fattori, insieme alle costrizioni di natura fisica imposte alle cellule durante la differenziazione, come la loro localizzazione sulla superficie dell'embrione, rappresentano ciò che viene definito campo morfogenetico. Tale campo non è un'astrazione; può, in linea di principio, essere definito per ciascun tipo di cellula, dando un nome ai fattori e alle forze applicate al citoscheletro e alle quali il citoscheletro reagisce.
In conclusione, si inizia a intravedere come l'informazione lineare immagazzinata nel genoma venga utilizzata per generare strutture tridimensionali nello spazio e nel tempo. Ne consegue che le strutture finali hanno funzioni chiare e definite come, per esempio, la segregazione cromosomica, la contrazione muscolare e il pensiero, sebbene all'origine di tale ordine ci sia un certo numero di interazioni di tipo stocastico. Tutto questo avviene grazie a un numero estremamente limitato di elementi base, cioè le proteine strutturali del citoscheletro, le quali interagiscono con una serie numerosissima di altri fattori. I principî alla base della morfogenesi cellulare sono relativamente pochi: è probabile che esista una sorta di codice morfogenetico che presto sarà decifrato. Si comincia a pensare che il dogma 'un gene una funzione' sia estremamente ingenuo. In effetti, dal momento che le forme giocano un ruolo fondamentale per la funzione cellulare e che gli stessi geni, come quelli per l'actina e la tubulina, generano centinaia di forme diverse, viene a perdersi la correlazione fra il gene singolo e la relativa funzione. Le funzioni scaturiscono dal comportamento coordinato di molti prodotti genici. Conoscere tutti i geni di un organismo, quindi, non significa affatto sapere in che modo esso è costruito. Per far ciò è necessario anche risolvere un problema complesso: capire come tutti i prodotti dei geni interagiscano in modo dinamico.
Bibliografia
Alberts 2002: Alberts, Bruce e altri, Molecular biology of the cell, 4. ed., New York, Garland Science, 2002 (1. ed.: London, Garland, 1989).
Baas 1999: Baas, Peter W., Microtubules and neuronal polarity: lessons from mitosis, "Neuron", 22, 1999, pp. 23-31.
Boleti 1996: Boleti, Haralabia - Karsenti, Eric - Vernos, Isabelle, Xklp2, a novel Xenopus centrosomal kinesin-like protein required for centrosome separation during mitosis, "Cell", 84, 1996, pp. 49-59.
Carlier 2003: Carlier, Marie-France - Clainche, Christophe Le - Wiesner, Sebastian - Pantaloni, Dominique, Actin-based motility: from molecules to movement, "Bioessays", 25, 2003, pp. 336-345.
Frankel 1984: Frankel, Joseph, Pattern formation in ciliated protozoa, New York, MacMillan, 1984.
Franzini-Armstrong, Fischman 1994: Franzini-Armstrong, Clara - Fischman, Donald A., Morphogenesis of skeletal muscle fibers, New York, McGraw-Hill, 1994.
Gerhart, Kirschner 1997: Gerhart, Jacquelyn - Kirschner, Marc W., Cells, embryos and evolution, Malden (Mass.)-Oxford, Blackwell Science, 1997.
Gilbert 1988: Gilbert, Scott F., Developmental biology, Sunderland (Mass.), Sinauers Associates, 1988.
Gurdon 1992: Gurdon, John B., The generation of diversity and pattern in animal development, "Cell", 68, 1992, pp. 185-199.
Hartwell 1999: Hartwell, Leland H. - Hopfield, John J. - Leibler, S. - Murray, Andrew W., From molecular to modular cell biology, "Nature", 402, 1999, pp. 47-52.
Hyatt 1996: Hyatt, B.A. - Lohr, J.L. - Yost, H. Joseph, Initiation of vertebrate left-right axis formation by maternal Vg1, "Nature", 384, 1996, pp. 63-65.
Hyman, Karsenti 1996: Hyman, Anthony A. - Karsenti, Eric, Morphogenetic properties of microtubules and mitotic spindle assembly, "Cell", 84, 1996, pp. 401-410.
Karsenti, Vernos 2001: Karsenti, Eric - Vernos, Isabel, The mitotic spindle: a self-made machine, "Science", 294, 2001, pp. 543-547.
Kerr 1999: Kerr, Jeffrey, Atlas of functional histology, London, Mosby Incorporated, 1999.
Kirschner, Mitchison 1986: Kirschner, Marc W. - Mitchison, Tim, Beyond self-assembly: from microtubules to morphogenesis, "Cell", 45, 1986, pp. 329-342.
Kirschner, Gerhart 1998: Kirschner, Marc W. - Gerhart, John, Evolvability, "Proceedings of the National Academy of Sciences USA", 95, 1998, pp. 8420-8427.
Knust 2002: Knust, E., Regulation of epithelial cell shape and polarity by cell-cell adhesion (Review), "Molecular membrane biology", 19, 2002, pp. 113-120.
Lafont, Simons 1996: Lafont, Frank - Simons, Kai, The role of microtubule based motors in the exocytic transport of polarized cells, "Seminars in cell & developmental biology", 7, 1996, pp. 343-355.
Lehman 1995: Lehman, Ruth, Establishment of embryonic polarity during Drosophila oogenesis, "Seminars in cell & developmental biology", 6, 1995, pp. 25-38.
Lengyel, Iwaki 2002: Lengyel, Judith A. - Iwaki, D.D., It takes guts: the Drosophila hindgut as a model system for organogenesis, "Developmental biology", 243, 2002, pp. 1-19.
Lubarsky, Krasnow 2003: Lubarsky, B. - Krasnow, M.A., Tube morphogenesis: making and shaping biological tubes, "Cell", 112, 2003, pp. 19-28.
Lyczak 2002: Lyczak, Rebecca - Gomes, José E. - Bowerman, Bruce, Heads or tails: cell polarity and axis formation in the early Caenorhabditis elegans embryo, "Developmental biology", 3, 2002, pp. 157-166.
Mays 1994: Mays, R.W. - Beck, Kenneth A. - Nelson, W. James, Organization and function of the cytoskeleton in polarized epithelial cells: a component of the protein sorting machinery, "Current opinion in cell biology ", 6, 1994, pp. 16-24.
Misteli 2001: Misteli, Tom, The concept of self-organization in cellular architecture, "The journal of cell biology", 155, 2001, pp. 181-185.
Moore, Endow 1996: Moore, Jonathan D. - Endow, Starin A., Kinesin-related proteins: a phylum of motors for microtubule based motility, "Bioessays", 18, 1996, pp. 207-219.
Mooseker, Cheney 1995: Mooseker, Marc S. - Cheney, Richard E., Unconventional myosins, "Annual review of the cell and developmental biology", 11, 1995, pp. 633-675.
Nédélec 1997: Nédélec, François - Surrey, Thomas - Maggs, A.C. - Leibler, Stanislas, Self-organization of microtubules and motors, "Nature", 389, 1997, pp. 305-308.
Nédélec 2003: Nédélec, François - Surrey, Thomas - Karsenti, Eric, Self-organization and forces in the microtubule cytoskeleton, "Current opinion in cell biology", 15, 2003, pp. 118-124.
Nelson 2003: Nelson, W. James, Adaptation of core mechanisms to generate cell polarity, "Nature", 422, 2003, pp. 766-774.
Pfeffer 2003: Pfeffer, Suzanne, Membrane domains in the secretory and endocytic pathways, "Cell", 112, 2003, pp. 507-517.
Pizon 2002: Pizon, Veronique e altri, Transient association of titin and myosin with microtubules in nascent myofibrils directed by the MURF2 RING-finger protein, "Journal of cell science", 115, 2002, pp. 4469-4482.
Simons, Toomre 2000: Simons, Kai - Toomre, Derek, Lipid rafts and signal transduction, "Nature reviews molecular cell biology", 1, 2000, pp. 31-39.
Surrey 2001: Surrey, Thomas - Nédélec, François - Leibler, Stanislas - Karsenti, Eric, Physical properties determining self-organization of motors and microtubules, "Science", 292, 2001, pp. 1167-1171.
Tassin 1985: Tassin, A.M. - Maro, B. - Bornens, M., Fate of microtubule organizing centers during in vitro myogenesis, "The journal of the cell biology", 100, 1985, pp. 35-46.
Thieffry, Sanchez 2002: Thieffry, Denis - Sanchez, Lucas, Alternative epigenetic states understood in terms of specific regulatory structures, "Annals of the New York Academy of Science", 981, 2002, pp. 135-153.
Verde 1992: Verde, Fulvia - Dogterom, M. - Stelzer, Ernst - Karsenti, Eric - Leibler, Stanislas, Control of microtubule dynamics and length by cyclin A and cyclin B dependent kinases in Xenopus egg extracts, "The journal of the cell biology", 118, 1992, pp. 1097-1108.
Vernos, Karsenti 1995: Vernos, Isabelle - Karsenti, Eric, Chromosomes take the lead in spindle assembly, "Trends in cell biology", 5, 1995, pp. 297-301.
Welch 1997: Welch, Matthew D. - Mallavarapu, Ameil - Rosenblatt, Jody - Mitchison, Tim J., Actin dynamics in vivo, "Current opinion in cell biology", 9, 1997, pp. 54-61.
Wittmann 2001: Wittmann, Torsten - Hyman, Anthony - Desai, Anshad, The spindle: a dynamic assembly of microtubules and motors, "Nature cell biology", 3, 2001, pp. 28-34.
Zernicka-Goetz 2002: Zernicka-Goetz, Magdalena, Patterning of the embryo: the first spatial decisions in the life of a mouse, "Development", 129, 2002, pp. 815-829.