
Virus
Virus
Il termine 'virus' proviene dal latino e significa 'veleno'. I virus sono formazioni biologiche elementari, le più piccole e semplici strutture biologiche presenti in natura, alle quali manca qualsiasi organizzazione cellulare e qualsiasi meccanismo produttore di energia. A differenza dei batteri, i virus non sono visibili al microscopio ottico. Essi sono costretti, per riprodursi, a vivere all'interno delle cellule: sono pertanto obbligati a comportarsi come parassiti intracellulari. Se i virus sono stati identificati solo di recente, le malattie virali sono note da tempo, avendo accompagnato la storia dell'umanità sin dalla nascita delle prime civiltà. Al contrario, i virus non costituirono, probabilmente, un grande problema per l'uomo primitivo, in particolare nel Paleolitico, quando gli Ominidi cacciatori-raccoglitori vivevano in piccoli gruppi, isolati gli uni dagli altri, e si spostavano sovente. Con l'avvento del Neolitico, l'uomo si trasformò in agricoltore-allevatore, diventando stanziale. Di conseguenza, si andarono formando gruppi sempre più grandi, finché non si giunse alla costituzione delle prime civiltà fluviali (quella assiro-babilonese, egiziana, cinese, indiana, ecc.) e alla nascita delle prime grandi città. Fu a questo punto che le malattie virali cominciarono a dilagare, sino a diventare endemiche, potendo essere sostenute da una popolazione sufficientemente numerosa e tenuta insieme da una densa rete di comunicazioni.
Prototipi di malattie come il vaiolo o il morbillo, definite 'malattie della civilizzazione' dallo storico William McNeill, originarono dal bestiame non più di mille anni prima della comparsa di tali aggregazioni umane, e iniziarono a comportarsi come le attuali malattie dell'infanzia, in Mesopotamia, duemila anni prima della nascita di Cristo. Nelle aree abitate dai Sumeri, già dal 3000 a.C. erano presenti una dozzina di città che contavano 30.000÷50.000 abitanti, in stretta comunicazione fra loro, tali da formare un'aggregazione umana di circa mezzo milione di individui.
Le malattie virali hanno segnato profondamente la storia dell'umanità. È a tal riguardo paradigmatico ciò che accadde quando Hernando Cortez e i suoi uomini vennero attaccati e sconfitti dagli Aztechi a Tenochtitlan, nel 1520. Invece di essere inseguiti e uccisi, gli Spagnoli poterono riorganizzarsi, trovando anche alleati nel campo avverso, per poi assediare e distruggere la città e il tempio che si trovava al suo centro. Quello che accadde ai quattrocento Spagnoli fuggiti da Tenochtitlan sembrava inspiegabile. È però noto che, durante la cosiddetta noche trista, ovvero la notte della sconfitta degli Spagnoli, il vaiolo scoppiò nella città, e lo stesso nipote di Montezuma, che aveva condotto le truppe azteche alla vittoria, ne morì. Gli effetti di una malattia virale su una popolazione vergine, come quella degli Aztechi nel XVI sec., possono essere devastanti; diversamente, gli Spagnoli, essendo vissuti in una società in cui il vaiolo si comportava come una malattia endemica dell'infanzia, difficilmente sarebbero arrivati all'età adulta senza esserne stati affetti durante i primi anni di vita. La resistenza acquisita dagli Spagnoli nei confronti del vaiolo dovette apparire agli indigeni come un evento numinoso, sancendo la vittoria del Vecchio sul Nuovo mondo.
L'identificazione dei virus
L'esistenza di particolari agenti patogeni dell'uomo (nonché degli animali e delle piante) che, a differenza di altri microrganismi già noti, quali batteri o Protozoi, non erano visibili al microscopio ottico e non crescevano sui normali mezzi nutritivi, venne dimostrata negli ultimi decenni del XIX secolo. Louis Pasteur, nel 1881, riprodusse la rabbia in cani e conigli mediante inoculazione di tessuto nervoso (batteriologicamente sterile e privo di microbi evidenziabili col microscopio ottico) appartenente ad animali deceduti di quella malattia. Non essendo stato in grado di individuare l'agente causale, egli ipotizzò, con grande intuito, che esso fosse troppo piccolo per essere visibile. Nel 1892, il botanico russo Dimitri Ivanovsky riuscì a infettare piante sane con il succo, filtrato per candela (e perciò privo di microrganismi relativamente grandi), di piante affette dal 'mosaico del tabacco', una malattia virale che fa appunto assumere alle foglie di tabacco l'aspetto di un mosaico; è per questo che Ivanovsky viene ora ricordato come lo scopritore dei virus. Nel 1898, Friedrich Löffler e Paul Frosch riprodussero l'afta epizootica (nota agli anglosassoni come foot and mouth disease) in bovini sani, mediante inoculazione di linfa aftosa di bovini malati, e Giuseppe Sanarelli dimostrò che era possibile trasmettere la mixomatosi a conigli sani inoculando filtrati batteriologicamente sterili. Finalmente, nel 1899, Martinus Willem Beijerinck dimostrò che era possibile trasmettere il mosaico del tabacco in serie, da pianta a pianta, per mezzo di filtrati, e designò come contagium vivum fluidum l'agente infettivo in grado di attraversare i minuti pori delle candele e di riprodursi indefinitamente, passando da un organismo all'altro. Nacque così, sul finire del secolo, il concetto di 'agente infettivo filtrabile', meglio noto, oggi, come virus.
Si entrava, lentamente, in una nuova era. Prima del secolo scorso, solo un'attenta osservazione clinica permetteva di distinguere un caso di vaiolo dalla varicella o il morbillo dalla rosolia, e solo in seguito al lavoro di Rudolph Virchow si era riusciti a identificare il substrato patologico di alcune malattie infettive di origine virale. Bisogna arrivare agli anni Venti del XX sec. per assistere all'identificazione del primo virus umano, il virus della febbre gialla, da parte di Walter Reed. Questa fu seguita rapidamente da altre scoperte e, alla fine degli anni Trenta, una serie di virus era già stata identificata: virus oncogeni (potenzialmente in grado di provocare tumori nell'uomo e/o in alcuni animali), batteriofagi (in grado di infettare i batteri, conferendo loro, talvolta, specifiche proprietà), i virus dell'influenza e della parotite, e diversi arbovirus (trasmessi da Artropodi, in particolare da Insetti). Nel 1940, Max Delbruck e Salvador Edward Luria posero le basi della genetica microbica e della biologia molecolare, e identificarono gli eventi più importanti del ciclo riproduttivo virale. Successivamente, gli studi di Oswald Avery, Alfred D. Hershey e Martha Chase dimostrarono che il materiale genetico dei batteriofagi era costituito da DNA. Quindi, alla fine degli anni Quaranta, John F. Enders riuscì a far crescere il poliovirus su colture di tessuto, favorendo lo sviluppo di vaccini inattivati con formalina (da parte di Jonas Edward Salk) e di vaccini vivi attenuati (messi a punto da Albert Bruce Sabin). Si entrava così nell'era della moderna virologia.
Gli ulteriori avanzamenti tecnologici, con lo sviluppo della microscopia elettronica, dell'ultracentrifugazione e delle tecniche per l'elettroforesi di acidi nucleici e proteine, gettarono le fondamenta per una chiara definizione della struttura dei virus, e per un'analisi biochimica dettagliata delle componenti dei virioni (le particelle virali complete e potenzialmente infettanti) e delle loro proprietà. La cristallografia a raggi X ha poi favorito la definizione strutturale dei virus a un livello vicino a quello atomico. Infine, le tecniche di biologia molecolare hanno permesso di sequenziare l'intero genoma di diversi virus, mentre l'uso di anticorpi monoclonali ha consentito di identificare con grande precisione specifici domini sulle proteine virali. Queste conquiste hanno favorito lo sviluppo di tecniche diagnostiche e il disegno di efficaci terapie antivirali. Oggi, grazie a metodiche quali la PCR (Polimerase chain reaction), siamo in grado di amplificare anche piccole quantità di acido nucleico virale e di facilitarne la successiva identificazione. Per esempio, la PCR può permettere la diagnosi precoce di un'infezione virale, senza che sia necessario attendere la comparsa degli anticorpi, come avveniva quando erano disponibili soltanto le tecniche sierologiche.
I virus emergenti
L'espressione 'virus emergenti' si riferisce sia a virus che compaiono per la prima volta nella popolazione umana sia a virus che, apparentemente scomparsi, 'riemergono' dopo molti anni. Dalla fine degli anni Sessanta del Novecento, anche in conseguenza del miglioramento delle tecniche diagnostiche e della capacità di identificare nuovi virus, abbiamo assistito alla comparsa di una serie di nuove infezioni virali nell'uomo: le terribili febbri emorragiche africane, dovute a filovirus quali Marburg ed Ebola, vennero identificate per la prima volta, rispettivamente, nel 1967 e nel 1976; l'AIDS (Acquired immune deficiency syndrome) fece invece la sua prima apparizione nel 1981, ma ora si stima che il suo agente causale, l'HIV (Human immunodeficiency virus), fosse presente nella popolazione umana, o almeno in remote tribù africane, già negli anni Trenta; la SARS (Severe acute respiratory syndrome), dovuta a un nuovo coronavirus umano, fu identificata a Hong Kong all'inizio del 2003, ma era comparsa nella provincia cinese del Guandong sin dal novembre 2002. Queste malattie virali emergenti hanno tutte un serbatoio naturale di tipo animale: lo scimpanzé per l'HIV, lo zibetto o il pipistrello per il coronavirus della SARS, ecc. I serbatoi naturali, od 'ospiti definitivi', sono caratterizzati dal fatto che non sviluppano malattia pur producendo una quantità notevole di particelle virali. Quando l'uomo viene in contatto con animali infetti può, in particolari condizioni, infettarsi. Se il virus, mutando, è in grado di adattarsi all'uomo, allora diventerà capace di trasmettersi da persona a persona, determinando probabilmente la comparsa di un piccolo outbreak (focolaio epidemico), o anche una vera e propria epidemia di grandi dimensioni (quando l'epidemia si diffonde nella maggior parte del globo si parla di pandemia).
Un fenomeno particolare di emergenza e riemergenza di infezioni virali è rappresentato dai virus influenzali. Questi, infatti, mutano in continuazione, determinando ogni anno epidemie di dimensioni variabili. Dal momento che il sistema immunitario delle persone che hanno sviluppato l'influenza in anni precedenti (anche l'anno prima) non riconosce le proteine di superficie (importanti antigeni virali) a causa delle mutazioni intercorse nel frattempo, si crea continuamente una situazione di suscettibilità, seppure solo parziale, ai nuovi ceppi. Di tanto in tanto, parti del genoma del virus influenzale umano possono essere scambiate con quelle di virus influenzali animali; in particolare, ciò avviene nel maiale, che è recettivo sia ai virus umani sia a quelli dei volatili migratori (oche e anatre), che rappresentano il vero serbatoio dei virus influenzali. Il cosiddetto 'passaggio di specie', ovverosia l'adattamento del virus a una specie diversa (e, per quanto ci riguarda, all'uomo), ha determinato nel secolo scorso tre pandemie: la 'spagnola' (1918-1919), l''asiatica' (1957-1958) e la 'Hong Kong' (1968-1969). Nel 1997, poi, per la prima volta, si è documentato il passaggio di un terribile virus influenzale aviario, il cosiddetto H5N1, direttamente da un volatile (gallo) all'uomo. L'estensione e la persistenza dell'infezione nei volatili selvatici asiatici, e i conseguenti focolai epidemici verificatisi nel pollame (quando l'infezione passa dai volatili selvatici a quelli domestici assume caratteristiche di elevata patogenicità), dal quale l'infezione può essere trasmessa all'uomo, hanno creato un grande allarme circa la possibilità che questo virus possa dar luogo, mutando e divenendo trasmissibile da persona a persona, a una nuova pandemia.
Esistono naturalmente alcuni fattori che facilitano la comparsa e la diffusione di nuove, emergenti infezioni da virus: (a) l'esplorazione di aree geografiche remote, precedentemente isolate, può portare l'uomo a contatto con nuovi serbatoi d'infezione; (b) i cambiamenti climatici e ambientali possono favorire il contatto con serbatoi animali (per es., particolari condizioni climatiche, che possono dare impulso alla migrazione di topi e altri roditori dai campi alle case, oppure lo sfruttamento delle foreste tropicali); (c) l'urbanizzazione può contribuire ad amplificare alcune infezioni virali concentrando persone che provengono da luoghi diversi in un'unica area densamente popolata; (d) l'aumentata esposizione a fattori iatrogeni, quali l'ospedalizzazione, l'eventuale uso di siringhe non sterili, il trapianto di organi animali, può aumentare il rischio di nuove infezioni virali per l'uomo; (e) i viaggi internazionali, e in particolare gli spostamenti veloci, possono diffondere nuove infezioni a distanza di breve tempo o anche contribuire all'introduzione di vecchie infezioni in aree precedentemente indenni.
Come è fatto e come si replica un virus
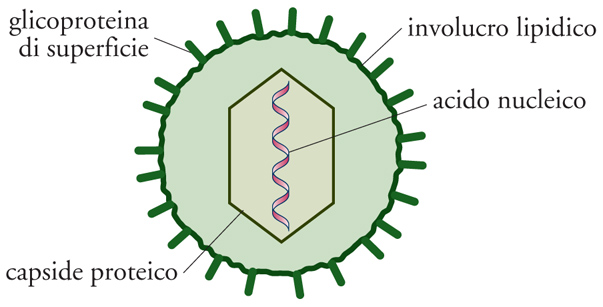
I virus hanno dimensioni, forma e struttura variabili. Quelli più piccoli, in genere di forma sferica, non arrivano a 20 nm. I più grandi possono superare in lunghezza il micron, ma non arrivano a 100 nm di larghezza. I virus di forma cilindrica hanno un diametro variabile in genere fra 60 e 250-300 nm. Il costituente essenziale di un virus è rappresentato dal nucleocapside, cioè dall'insieme di acido nucleico e capside proteico, mentre all'esterno può essere presente o meno un involucro lipidico. Il virus, nella sua forma completa, viene definito 'virione'.
L'informazione genetica dei virus è codificata dall'acido nucleico, DNA o RNA, di dimensioni variabili, a elica unica o doppia, di forma circolare o lineare. I virus a RNA lineare possono avere sia un singolo pezzo di acido nucleico sia un numero variabile di segmenti. L'acido nucleico virale è contenuto all'interno di una capsula proteica (capside), composta da molteplici subunità proteiche pressoché identiche (i cosiddetti 'capsomeri'). Il capside preserva il genoma virale dall'azione di agenti denaturanti esterni, e ne facilita l'ingresso all'interno delle cellule mediante un ancoraggio a specifici recettori presenti sulla superficie (per la precisione, sulla membrana plasmatica) della cellula dell'ospite. La combinazione dell'acido nucleico e del capside proteico che lo circonda viene definita 'nucleocapside'. Con l'eccezione dei virus più complessi, la struttura del capside virale presenta due possibili simmetrie: la simmetria elicoidale, in genere propria di virus a RNA, o la simmetria icosaedrica, assunta da virus sia a DNA sia a RNA.
In alcuni virus, il capside è circondato da un involucro lipidico o envelope, che viene acquisito quando il virus fuoriesce dalla membrana nucleare o citoplasmatica della cellula ospite. All'interno di tale doppio strato lipidico possono essere contenute glicoproteine di superficie, quali per esempio l'emoagglutinina o la neuramminidasi del virus influenzale. Queste proteine, che protrudono all'esterno della superficie del virus, fungono da veri e propri antigeni, cioè sono in grado di evocare una risposta anticorpale specifica. Sono anche presenti proteine della matrice, che sono localizzate nello strato interno dell'envelope e che lo connettono al capside. I virus possono contenere proteine ad attività enzimatica (quale, per es., la polimerasi), che gli permettono di iniziare la replicazione del proprio acido nucleico all'interno della cellula parassitata.
Innanzitutto, una volta entrato nell'organismo e raggiunto l'organo bersaglio, il virus deve penetrare all'interno di una cellula. Esso si attacca ai recettori cellulari, quindi penetra all'interno della cellula probabilmente mediante un meccanismo definito 'pinocitosi' (attraverso il quale la membrana cellulare ingloba il virus); infine avviene la denudazione dell'acido nucleico (il DNA o RNA virale perde il capside) e ha inizio la sua replicazione, nonché la sintesi delle proteine virali che andranno a costituire i nuovi virioni. Questi, una volta maturi, verranno liberati all'esterno della cellula, che (seppure non sempre andrà incontro alla morte, mentre i nuovi virioni saranno pronti a invadere nuove cellule e a iniziare un nuovo ciclo replicativo. (fig. 3)
Le proprietà dei virus
Fra le principali caratteristiche dei virus, oltre alle dimensioni, alla struttura e alla forma, alla composizione chimica e alla struttura antigene, è importante considerare la resistenza agli agenti chimici (disinfettanti) e fisici (calore). In particolare, sono stati estensivamente indagati gli effetti della temperatura, dei raggi ultravioletti, del pH e dell'etere, nonché di una serie di agenti chimici, sull'infettività dei virus. Ciò ha permesso di comprendere meglio le modalità di trasmissione indiretta di alcuni di essi, che richiedono una certa stabilità in determinate condizioni ambientali, e di acquisire utili informazioni sulla possibilità di interventi di inattivazione e sterilizzazione. In particolare, è importante correlare la suscettibilità dei virus a determinati agenti con la loro composizione strutturale. In genere, l'infettività virale viene annullata a 50÷60 °C per 30 min, anche se il virus dell'epatite B e i prioni sono più resistenti. È invece possibile conservare i virus mediante un raffreddamento a temperature inferiori a quelle del congelamento. I virus sono stabili a pH compreso fra 5 e 9, mentre vengono inattivati dall'esposizione ai raggi ultravioletti. La caratteristica più importante, anche per questioni di classificazione, è quella relativa alla sensibilità all'etere: in seguito al contatto vengono inattivati solo i virus che possiedono un involucro lipidico, quali per esempio gli herpesvirus e il virus dell'influenza, così come l'HIV.
L'effetto citopatico e il potere patogeno
I virus sono parassiti intracellulari obbligatori ed esplicano effetti citopatogeni, anche quando non danno manifestazioni cliniche apprezzabili. La riproduzione dei virus ha luogo solo all'interno e a spese delle cellule di un organismo ospite; queste possono esserne danneggiate in varia misura e presentare alterazioni lievi o gravi. Il complesso delle alterazioni procurate alle cellule ospiti si definisce 'effetto citopatico'. Le alterazioni citologiche dipendono sia dalla specie di virus infettante sia dal tipo di cellula infettata. Gli effetti citopatici osservati al microscopio ottico sono di diversi tipi, e comprendono sia alterazioni morfologiche cellulari che inclusioni.
Alterazioni morfologiche. Cellule arrotondate (da virus vaccinico), ingrandite (varicella), riunite in un sincizio (morbillo), con presenza o meno di vacuoli citoplasmatici; a volte, le cellule ospiti possono risultare del tutto distrutte. Nelle infezioni da oncovirus, infine, le cellule perdono la loro forma e la proprietà di rimanere monostratificate e si moltiplicano in strati sovrapposti.
Inclusioni (o corpi inclusi) da virus. Particolari formazioni presenti nel citoplasma o nel nucleo delle cellule ospiti, dell'ordine di grandezza del micron, rappresentano i centri di replicazione virale intracellulare, e possono essere di forma rotondeggiante, a pera, o del tutto irregolare; rispetto ai coloranti si comportano come materiale acidofilo o basofilo. Dall'esame al microscopio elettronico, si evince che si tratta di ammassi di particelle virali riunite insieme.
Inclusioni citoplasmatiche. Ne esiste una discreta varietà, dai corpi di Guarnieri, presenti nelle cellule della cute infettate da virus del vaiolo o del vaccino, ai corpi di Negri, grandi da 1 a 25 μ, che si trovano nelle cellule piramidali del corno di Ammone o in altre cellule nervose dell'uomo o di animali affetti da rabbia. I corpi di Henderson-Paterson, infine, si rinvengono negli strati basali dell'epidermide colpiti dal virus del mollusco contagioso.
Inclusioni nucleari di tipo A e B. Quelle di tipo A occupano gran parte del nucleo, e sono presenti nelle cellule parassitate da virus erpetici (corpi di Lipschuts) o dal virus della febbre gialla. Quelle di tipo B, che appaiono come piccoli granuli all'interno del nucleo, sono caratteristiche dei poliovirus. Il virus morbilloso determina inclusioni sia nel nucleo sia nel citoplasma.
I virus possono dar luogo anche ad alterazioni cromosomiche: il virus della varicella/Herpes zoster blocca la mitosi nella metafase, rompendo alcuni cromosomi e determinando la formazione di micronuclei; l'Herpes simplex può provocare una divisione anomala dei nuclei; il virus del morbillo una frammentazione dei cromosomi; i virus oncogeni una serie di anormalità cromosomiche. Infine, l'effetto dei virus si estrinseca anche a diversi livelli, ed è possibile rilevare fini alterazioni delle minute strutture cellulari al microscopio elettronico, nonché modificazioni metaboliche indotte a carico della sintesi cellulare del DNA, dell'RNA e delle proteine cellulari.
I virus possono essere patogeni per una o più specie. Alcuni virus sono patogeni solo per l'uomo (per es., morbillo, poliomielite, parotite epidemica), mentre altri, come la rabbia, hanno uno spettro d'ospite più ampio, potendo infettare sia l'uomo sia il cane, il lupo, il gatto e altri animali. Le porte d'ingresso per i virus nell'organismo umano sono rappresentate principalmente dalle prime vie respiratorie. Essi si impiantano sovente a livello di tonsille ed epitelio nasale, trachea, bronchi e bronchioli, penetrando mediante processi noti col nome di 'adsorbimento' e 'viropessi'. La mucosa congiuntivale e il tratto intestinale rappresentano altre porte d'ingresso. I virus trasmessi dagli Artropodi, infine, penetrano attraverso la pelle mediante la puntura dell'insetto. Una volta entrati, alcuni virus tendono a invadere l'organismo e si diffondono attraverso le vie linfatiche ed ematiche (la viremia, o presenza di virus nel sangue, è un fenomeno comune); altri virus utilizzano, per diffondersi, il sistema nervoso periferico.
Con le modalità descritte, i virus raggiungono i tessuti e gli organi bersaglio, sulla base di quello che viene definito il loro 'tropismo': distinguiamo così virus dermotropi (il cui bersaglio è la pelle), neurotropi, epatotropi, enterotropi, neurotropi. Modificazioni di tale tropismo vengono utilizzate per mettere a punto vaccini: attraverso passaggi in serie su uova fecondate o su colture di cellule si ottiene, per esempio, un ceppo vaccinale di virus della poliomielite enterotropo ma non neurotropo, cioè in grado di localizzarsi a livello intestinale ma non di raggiungere il sistema nervoso centrale e causare le paralisi che hanno reso tristemente famosa la malattia. Lo spettro clinico dell'infezione virale è ampio: in alcuni casi essa può decorrere in maniera del tutto asintomatica (resterà la sola risposta anticorpale, la 'cicatrice immunologica', a documentare l'avvenuta esposizione al virus), mentre in altri si manifestano i sintomi della malattia, con varie sfumature di gravità. A questo proposito, si definisce come 'potere patogeno' di un agente virale la sua capacità di provocare malattia, mentre per 'virulenza' intendiamo l'intensità del potere patogeno. Quest'ultima è una caratteristica propria di ogni specifico virus, per cui il rapporto fra il numero di casi di malattia (o di malattia grave) e il numero di persone infette varia a seconda del virus considerato.
La classificazione dei virus
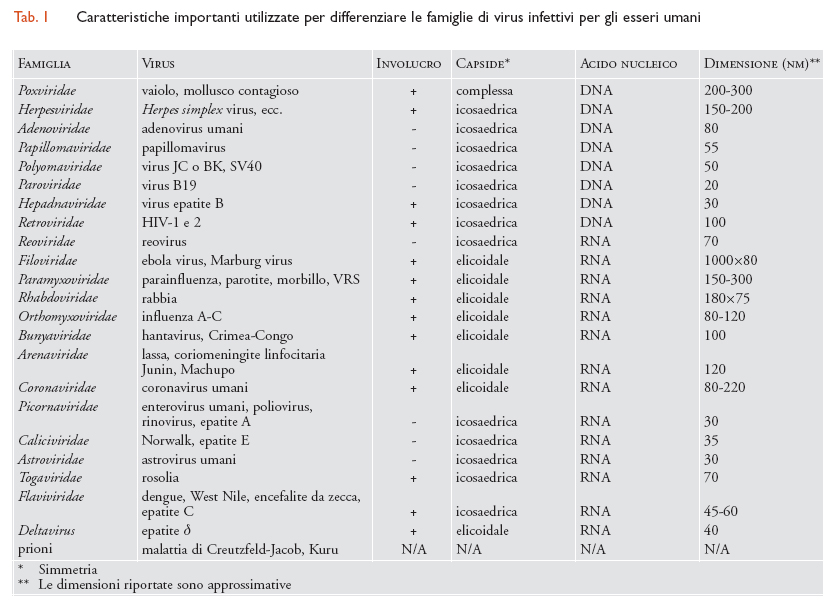
La prima classificazione dei virus quali entità distinte rispetto agli altri microrganismi si basava sulla loro capacità di passare attraverso filtri costituiti da membrane con pori di piccole dimensioni: tali pori lasciavano passare i virus (agenti filtrabili) ma non gli organismi più grandi, quali per esempio i batteri. Successivamente, per classificare i virus, vennero utilizzate alcune proprietà patogenetiche, quali per esempio il tropismo per specifici organi, o caratteristiche epidemiologiche (il termine 'arbovirus', o arthropod borne virus, definisce i virus trasmessi da Artropodi). Dagli anni Cinquanta del Novecento cominciarono a entrare in uso criteri morfologici e fisico-chimici, per poi arrivare, più recentemente, a definire i virus sulla base delle proprietà dell'acido nucleico. Attualmente, i sistemi di classificazione sono basati su: (a) tipo e struttura del loro acido nucleico e strategia di replicazione; (b) tipo di simmetria del capside virale (elicoidale o icosaedrico); (c) presenza o assenza di involucro lipidico. La microscopia elettronica, in particolare, ha permesso l'identificazione di caratteristiche dei virioni quali la dimensione, la forma, la simmetria, le caratteristiche della superficie virale, la presenza o assenza di involucro lipidico, il sito e il metodo di assemblaggio. Per classificare i virus possono anche essere utilizzate altre caratteristiche, quali la sensibilità ad agenti fisici o chimici (tab. 1).
I metodi diagnostici
I saggi utilizzati per la diagnosi di malattie virali possono essere raggruppati in tre categorie principali: (a) identificazione diretta; (b) isolamento virale; (c) sierologia. Nel primo caso il materiale clinico viene direttamente esaminato per valutare la presenza di particelle virali, antigeni virali, o acidi nucleici virali. A tal fine si possono usare diverse tecniche, quali la microscopia elettronica, l'esame istologico per il rilievo di corpi inclusi di natura virale, l'immunofluorescenza o metodi immunoenzimatici per l'identificazione di antigeni virali, o tecniche molecolari (per es., la PCR) per l'evidenziazione diretta di parti del genoma virale. Nel secondo caso il materiale prelevato dalla persona infetta viene inoculato in colture cellulari, uova o animali da esperimento, nel tentativo di far replicare e quindi identificare il virus, in base alla comparsa di un effetto citopatico (alterazioni nella morfologia cellulare), neutralizzazione mediante anticorpi specifici, ecc.
Infine la diagnosi sierologica si ottiene andando a rilevare nel siero del paziente gli anticorpi diretti contro il virus sospettato di essere la causa della malattia in atto. Dal momento che molte malattie infettive sono piuttosto comuni, una semplice positività sierologica non permette di esporre diagnosi, in quanto la presenza di anticorpi potrebbe essere dovuta a un'infezione contratta in passato. È per questo motivo che occorre eseguire due prelievi di sangue successivi (ad almeno due settimane di distanza l'uno dall'altro), il primo nella fase acuta della malattia e il secondo durante la convalescenza: la diagnosi di infezione viene posta in caso di sieroconversione (comparsa di anticorpi specifici in una persona precedentemente negativa) o di aumento del titolo anticorpale di almeno quattro volte. Infatti nella maggior parte delle malattie infettive ad andamento acuto, gli anticorpi tendono a comparire circa 15÷20 giorni dopo l'inizio dei sintomi. In alcuni casi, la diagnosi può essere posta valutando in un solo campione di siero la presenza di particolari anticorpi, le cosiddette IgM (Immunoglobuline M), che vengono prodotte precocemente, durante l'infezione primaria, e tendono a diminuire, fino a scomparire, nel giro di pochi mesi. A differenza delle metodiche di identificazione diretta, che sono in grado di rilevare l'infezione nelle sue prime fasi, le tecniche sierologiche, con l'eccezione della rilevazione delle IgM, permettono una diagnosi accurata ma tardiva.
Come l'organismo si difende dai virus
L'organismo può difendersi dall'attacco dei virus in diversi modi. Esiste innanzitutto una prima linea difensiva, che consiste nell'interferenza virale. Questo meccanismo di difesa immunitaria aspecifica rende le cellule dell'ospite resistenti a una seconda infezione da parte di un virus differente o dello stesso virus. Esso si basa su una competizione fra due virus diversi per uno specifico sito di replicazione, oppure sulla necessità di utilizzare, per la replicazione stessa, un sistema enzimatico già usato da un altro virus. Il più importante meccanismo di interferenza è rappresentato dall'interferone, un insieme di proteine prodotte da cellule infettate che, reagendo con cellule non ancora contagiate, le rendono più resistenti all'infezione virale. Le difese immunitarie specifiche sono però il vero baluardo contro le infezioni. L'organismo può difendersi dai virus, così come anche dai batteri o da altri insulti esterni, producendo anticorpi (generati dalle plasmacellule, che sono dei linfociti B attivati), ossia dei veri e propri proiettili che vanno a colpire specificamente gli antigeni di un determinato virus. Gli anticorpi, prodotti in seguito a infezione naturale o a vaccinazione, proteggono l'individuo da una seconda infezione da parte dello stesso virus. L'altro meccanismo è quello dell'immunità cellulo-mediata, che riconosce come attore principale il linfocita T, e che consiste in una risposta diretta all'infezione virale, non mediata dalla produzione di anticorpi.
Difese non naturali: i farmaci antivirali
Le infezioni virali sono particolarmente temibili perché i virus, contrariamente ai batteri, non sono sensibili agli antibiotici. Purtroppo, fino a pochi anni fa, esistevano pochissimi farmaci antivirali. Il primo di una certa efficacia fu l'amantadina, utilizzata per l'influenza. Essa, insieme a un prodotto simile, la rimantadina, fa parte di un gruppo di molecole attive contro la proteina M2 del virus influenzale. È disponibile oggi un altro gruppo di farmaci antinfluenzali, gli inibitori della neuramminidasi (zanamivir e oseltamivir), una delle proteine di superficie del virus; essi possono attenuare i sintomi e abbreviare di circa un giorno la durata della malattia; inoltre, se somministrati tempestivamente, possono diminuire il rischio di sviluppare la malattia in persone esposte al contagio. Ma il vero e proprio boom nella ricerca e nello sviluppo di farmaci antivirali si è avuto con l'HIV. Dapprima l'introduzione della monoterapia con zidovudina, un analogo nucleotidico inibitore della trascrittasi inversa (ossia un farmaco in grado di bloccare la replicazione virale), determinò benefici limitati nella terapia dell'infezione da HIV. A metà degli anni Novanta del Novecento, lo sviluppo di nuovi farmaci, e in particolare di quelli appartenenti alla classe degli inibitori della proteasi, diede inizio all'era della terapia combinata. I cosiddetti 'cocktail' di farmaci antiretrovirali (ossia combinazioni di almeno tre molecole) hanno determinato un rilevante aumento della sopravvivenza delle persone sieropositive e un sensibile miglioramento della loro qualità di vita.
Infine, è da ricordare come sostanze immunoregolatrici, per esempio l'interferone, trovino uso corrente, in combinazione con farmaci antivirali quali la lamivudina o la ribavirina, nella terapia delle epatiti da virus B e C. Purtroppo, i farmaci antivirali possono generare resistenze. Infatti i virus, specialmente quelli a RNA come l'HIV o i virus influenzali, tendono a mutare piuttosto velocemente. Se viene somministrato un farmaco antivirale, questo tende a selezionare quei ceppi virali che, a seguito di mutazioni casuali, hanno acquisito la proprietà di resistere al farmaco stesso. Il rischio di acquisizione di resistenze aumenta soprattutto in caso di somministrazione di dosi inadeguate (sottodosaggio) o di mancata aderenza alla terapia (cioè quando, specie in caso di trattamento prolungato nel tempo, i farmaci non vengono presi con costanza). Si tratta di un problema rilevante per la terapia antivirale, in particolar modo quando è limitato il numero di farmaci efficaci disponibili.
La prevenzione delle malattie virali: i vaccini
Il medico inglese Edward Jenner, sul finire del Settecento, notò che i mungitori sviluppavano una malattia dei bovini nota come 'vaccino' (causata da un virus 'imparentato', cioè simile ma non uguale, a quello del vaiolo umano) ma non il vaiolo. In base a questa osservazione mise a punto una tecnica di inoculazione del vaiolo vaccino, dimostrando la sua efficacia protettiva nei confronti del ben più pericoloso vaiolo umano. Un bambino di otto anni, James Phipps, fu il primo a essere vaccinato, nel maggio del 1796; all'inizio di luglio, Jenner gli inoculò del pus di un malato di vaiolo, ma Phipps non si ammalò, mostrando poi, in seguito ad altre prove, di essere completamente protetto. Fu questo il primo vero grande successo della vaccinologia e, più specificamente, della guerra contro una temibile malattia infettiva. Passarono molti anni prima che, nel 1966, l'Organizzazione Mondiale della Sanità (OMS) avviasse un ambizioso programma con l'obiettivo di immunizzare tutte le persone che erano state a contatto con individui infetti. Nell'ottobre del 1977, in Somalia, venne diagnosticato l'ultimo caso 'naturale' (ovvero non verificatosi in seguito a incidente in un laboratorio di ricerca) di vaiolo: Ali Maolin fu l'ultima persona contagiata da un altro essere umano a sviluppare la malattia. Dal momento che questo virus non ha serbatoi animali e si trasmette soltanto per contagio interumano, nel 1979 il mondo venne dichiarato libero dal vaiolo. Rimane questo, a oggi, l'unico esempio di una malattia infettiva eradicata mediante la vaccinazione.
Nel 1885, Louis Pasteur salvò la vita a un ragazzo di nome Joseph Meister, morso da un cane affetto da rabbia, grazie a un vaccino sperimentale da lui messo a punto. Fu sempre Pasteur a coniare il termine 'vaccinazione', per onorare l'idea illuminante di Jenner. Un'altra terribile malattia virale, la poliomielite, è stata messa sotto controllo in gran parte del mondo grazie ai successi della vaccinazione. Il vaccino Salk, costituito da virus inattivato (cioè ucciso), venne introdotto per primo negli anni Cinquanta del Novecento, e quindi seguito dal vaccino Sabin, costituito da virus attenuato (ovverosia vivente ma non in grado di provocare malattia a livello neurologico). Con campagne di vaccinazione mirate, coordinate dall'OMS, si è quasi raggiunto l'obiettivo di eradicare la poliomielite. I vaccini virali utilizzano in genere ceppi di virus vivo attenuato, virus inattivato, o anche subunità virali (componenti del virus non infettanti) o antigeni virali sintetizzati in laboratorio con tecniche di ingegneria genetica. Il principio su cui si basa la vaccinologia è quello di introdurre antigeni virali, sotto varia forma, che stimolino le risposte immunitarie dell'organismo senza riprodurre la malattia. Una persona vaccinata è quindi in grado, grazie a specifiche risposte umorali (gli anticorpi) o cellulari (immunità cellulo-mediata), di respingere l'attacco dei virus.
La vaccinologia è oggi una scienza in rapida espansione, sia dal punto di vista dello spettro di malattie infettive contro le quali sono ormai disponibili vaccini efficaci, sia dal punto di vista delle tecniche utilizzate nell'allestimento dei vaccini virali. Alcuni dei moderni vaccini sono in grado di prevenire anche determinati tipi di tumori conseguenti a infezioni virali croniche. L'esempio più eclatante è quello del vaccino contro il virus dell'epatite B, che previene non solo l'infezione acuta, ma anche le sue possibili conseguenze a lungo termine, quali la cirrosi epatica e il carcinoma epatocellulare (un tumore del fegato). Recentemente si è avuta evidenza di efficacia da parte di un vaccino contro diversi oncogeni del virus del papilloma umano (HPV), considerato la causa necessaria del tumore della cervice uterina, cioè il tumore del collo dell'utero. È quindi probabile che vaccini contro l'HPV vengano presto messi in commercio e utilizzati in campagne vaccinali, specialmente nei Paesi maggiormente colpiti.
Un discorso a parte merita infine il vaccino antinfluenzale. Come è noto, i virus dell'influenza tendono a mutare molto rapidamente. Questo è il motivo della riemergenza di continue epidemie, ma anche della necessità di modificare la composizione del vaccino di anno in anno. Pertanto, la prevenzione di questa malattia infettiva, specie nelle persone a rischio di complicanze, necessita della ripetizione della vaccinazione ogni anno all'inizio dell'autunno. I vaccini rappresentano tuttora la maggiore risorsa che possediamo per combattere le malattie virali, essendo limitato il numero di farmaci antivirali a nostra disposizione. I progressi tecnologici alla base dello sviluppo e della produzione di nuovi vaccini sono in tal senso essenziali. Le malattie da virus sono fra le più antiche, avendo accompagnato l'uomo sin dalla nascita delle grandi città. Le piccole entità chiamate virus, che molti non considerano neanche esseri viventi veri e propri, non essendo dotate della capacità di vivere e riprodursi al di fuori della cellula ospite, restano tuttavia una delle principali minacce per l'umanità. Anzi, nuovi virus, cosiddetti 'emergenti', vengono introdotti nella popolazione umana, seminando spesso malattia e morte. Nonostante gli indubbi successi ottenuti con l'introduzione di vaccini sempre più efficaci e ben tollerati, le malattie virali sono ben lungi dall'essere sotto controllo. In particolare, a differenza di quanto è accaduto per i batteri, molti dei quali causano patologie controllabili con i normali antibiotici, per la maggior parte delle malattie da virus non esiste terapia efficace. Nonostante i recenti successi della ricerca, in particolare la messa a punto di diverse classi di farmaci antiretrovirali attivi contro l'HIV, il nostro bagaglio complessivo di farmaci antivirali resta tuttora limitato. Ciò induce a ritenere cha la lotta contro le malattie virali sarà ancora lunga e si rivelerà una vera e propria sfida per il futuro dell'umanità.
Bibliografia
Büchen-Osmond 2003: Büchen-Osmond, Cornelia, Taxonomy and classification of viruses, in: Manual of clinical microbiology, 8. ed., edited by Patrick R. Murray, Washington D.C.-Oxford, Blackwell, 2003
(www.ncbi.nlm.nih.gov/ICTVdb/MCM8.pdf).
Evans, Kaslow 1997: Evans, Alfred S. - Kaslow, Richard A., Viral infections of humans. Epidemiology and control, 4. ed., New York-London, Plenum Medical Book, 1997.
Knipe, Howley 2001: Knipe, David M. - Howley, Peter M., Fields virology, 4. ed., London-Philadelphia, Lippincott Williams & Wilkins, 2001, 2 v.
McNeill 1982: McNeill, William H., La peste nella storia, Torino, Einaudi, 1982 (ed. orig.: Plagues and peoples, Garden City, Anchor Press/Doubleday, 1976).
Morse 1993: Morse, Stephen S., Emerging viruses, New York-Oxford, Oxford University Press, 1993.