
Tatto
Tatto
Il tatto fa parte della sensibilità somatica generale insieme alla sensibilità termica, o termocezione, alla sensibilità dolorifica, o nocicezione, e alla sensibilità relativa alla posizione, all'orientamento e ai movimenti del corpo e delle sue parti, o propriocezione. Il tatto può essere definito come il complesso di sensazioni causate da un contatto diretto fra la superficie del corpo e oggetti esterni, oppure fra due o più parti del corpo stesso. Queste sensazioni sono distinguibili per la loro qualità e per i meccanismi anatomofisiologici che coinvolgono dalle sensazioni termiche e dolorose che possono derivare dal contatto materiale fra il corpo e gli agenti fisici e chimici esterni. Il tatto serve alla percezione cosciente sia del mondo esterno sia del proprio corpo, e partecipa, in modo generalmente inconscio, anche alla regolazione dei movimenti.
Tuttavia il tatto considerato come un sistema di senso a sé stante è un'astrazione. Nella vita ordinaria la conoscenza del mondo esterno si fonda sempre sull'integrazione di molteplici informazioni fornite da diverse modalità sensoriali, e lo stesso vale per la conoscenza del proprio corpo. La forma di un oggetto sconosciuto viene ordinariamente percepita mediante la vista, ma per decidere se l'oggetto è liscio o ruvido, molle o duro, pesante o leggero, caldo o freddo, tagliente o smusso sono necessarie informazioni di contatto diretto fornite non solo dal tatto, ma anche da altre modalità della sensibilità somatica. Il riconoscimento alla cieca tramite il cosiddetto 'tatto attivo', cioè palpando, manipolando e soppesando un oggetto, implica l'attivazione non solo di recettori tattili ma anche di altri recettori. I movimenti di esplorazione manuale servono indubbiamente a stimolare in modo efficace il senso del tatto, ma allo stesso tempo attivano anche vari recettori di strutture profonde (muscoli, articolazioni, tendini), generando informazioni non tattili ma propriocettive che contribuiscono al riconoscimento delle dimensioni e di altre proprietà strutturali dell'oggetto.
Ugualmente utile al riconoscimento di un oggetto è l'identificazione della sua temperatura apparente, che dipende dalla conducibilità termica del materiale da cui esso è costituito (per es., legno o metallo); tale identificazione avviene per azione dei termocettori cutanei e non dei recettori tattili. I recettori tattili della cavità orale cooperano inscindibilmente con quelli del gusto e dell'olfatto e con i recettori termici per il riconoscimento e la deglutizione delle sostanze alimentari, fornendo informazioni sulla forma, le dimensioni e le caratteristiche di superficie del contenuto della cavità stessa. Infine, anche i nocicettori cutanei possono intervenire nel processo di esplorazione per contatto rilevando potenzialità lesive di oggetti taglienti o pungenti. Quando si parla di riconoscimento tattile di un oggetto nella vita quotidiana o anche nell'esame clinico, si allude quindi a una prestazione non solo tattile, ma multisensoriale, più appropriatamente denominata 'stereognosi' o 'percezione aptica'.
I recettori tattili
Le sensazioni tattili prendono origine principalmente dalla cute, ma anche da mucose esposte all'esterno, come quelle della cavità orale o degli organi genitali esterni oppure la congiuntiva dell'occhio. Gli stimoli atti a generare sensazioni tattili sono deformazioni meccaniche non dolorose della cute o delle mucose, misurabili fisicamente come variazioni locali della pressione o della tensione. I sensori veri e propri di queste variazioni sono i terminali di fibre nervose funzionanti come meccanocettori dotati della capacità di trasdurre gli stimoli tattili in segnali elettrici, cioè in potenziali d'azione destinati a raggiungere il sistema nervoso centrale. La trasduzione si basa sull'azione deformante degli stimoli tattili trasmessi attraverso i tessuti cutanei o le mucose alla membrana dei suddetti terminali. Nella membrana si aprono allora dei canali ionici che consentono la diffusione intracellulare di cationi (specialmente sodio), con conseguente depolarizzazione della membrana stessa e generazione di potenziali d'azione al raggiungimento della soglia. I terminali delle fibre nervose al servizio del tatto sono sistematicamente associati a strutture cellulari specializzate non nervose che ne modulano la risposta meccanocettiva agli stimoli. Il complesso formato da uno o più terminali nervosi e dalle strutture cellulari associate costituisce un recettore tattile.
La cute glabra, che nella specie umana occupa tipicamente la superficie palmare delle mani, le labbra e la pianta dei piedi, contiene quattro tipi di recettori tattili, due superficiali (corpuscoli di Meissner e di Merkel) e due profondi (corpuscoli di Pacini e di Ruffini). I corpuscoli di Meissner, ubicati all'apice delle papille dermiche e ancorati da fibrille collagene alle cellule basali dell'epidermide, sono costituiti da strati di cellule epiteliali piatte fra i quali si infilano i terminali di più assoni (da due a sei) seguendo percorsi tortuosi. I corpuscoli di Merkel sono situati in grappoli nel derma interpapillare alla giunzione fra derma ed epidermide; ciascuno di essi è costituito da una cellula epiteliale che ingloba un terminale nervoso configurato a disco in una struttura semirigida avente l'apparenza di una sinapsi. Dividendosi ripetutamente nel derma, uno stesso assone può dare origine fino a cento terminali che innervano altrettanti corpuscoli di Merkel adiacenti e raggruppati in un'area di circa 5 mm2. I corpuscoli di Ruffini, disposti nella profondità del derma e strettamente connessi alle sue componenti collagene, sono strutture fusiformi incapsulate, nelle quali fibre connettivali e ramificazioni terminali nervose si incrociano ripetutamente ad angolo retto.
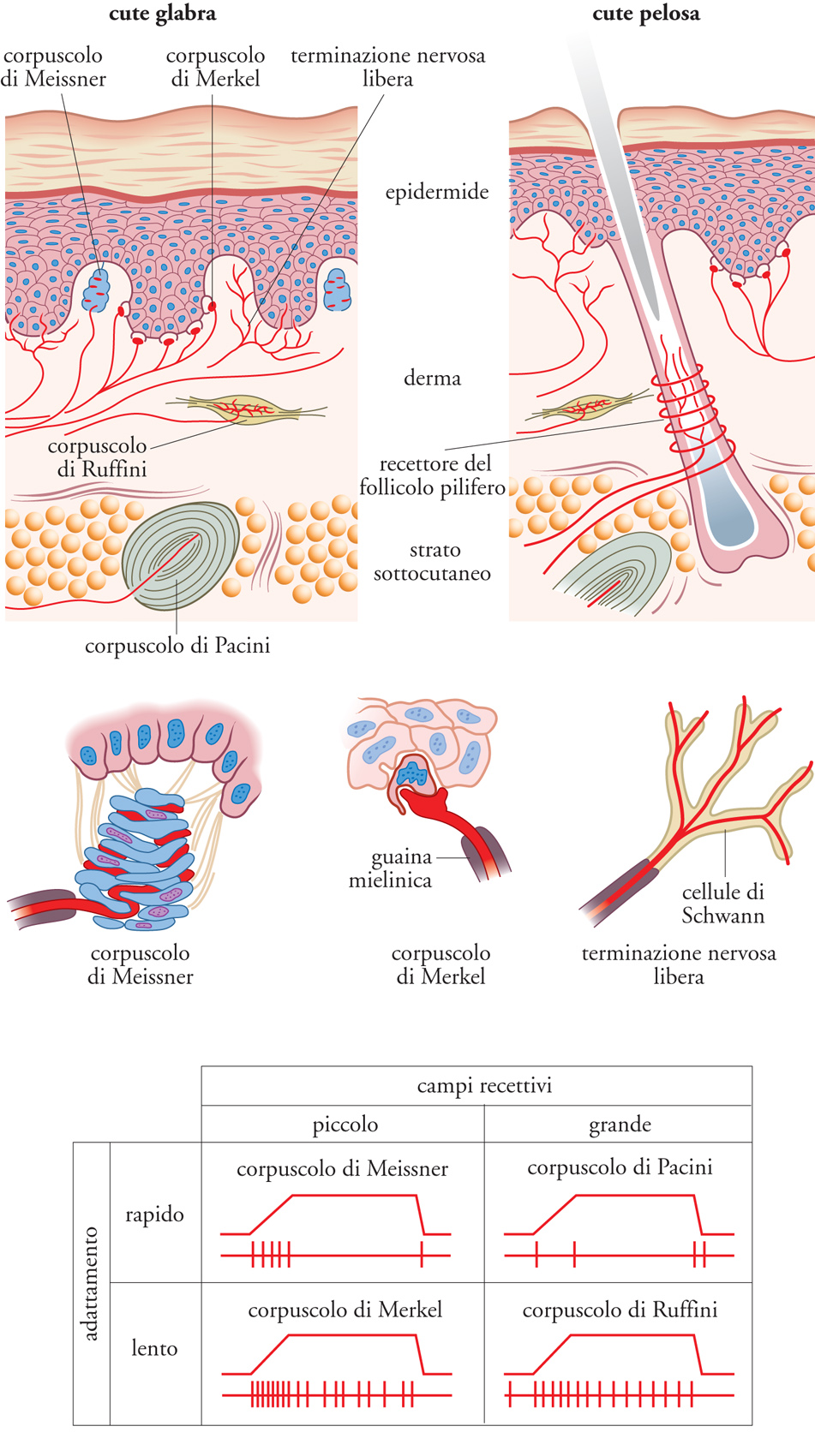
Il derma profondo e soprattutto lo strato sottocutaneo sono la sede dei corpuscoli di Pacini, in ciascuno dei quali un singolo terminale nervoso è avvolto da numerose lamelle concentriche (fino a settanta), formate da cellule connettivali allungate disposte come le foglie nel bulbo di una cipolla. Nella cute pelosa sono presenti corpuscoli di Merkel, di Ruffini, di Pacini, ma non corpuscoli di Meissner; in compenso sono presenti recettori dei follicoli piliferi, costituiti da terminali assonici giustapposti parallelamente alla guaina della radice del pelo o avvolti ripetutamente attorno a essa. Nella superficie palmare della mano i recettori più superficiali (di Meissner e di Merkel) sono assai più numerosi di quelli profondi. Nella cute dei polpastrelli il numero di ciascuno dei due tipi di recettori superficiali è pari a circa 100/cm2; rispetto a questo valore, la densità di entrambi i tipi si dimezza nella cute alla base del dito e si riduce ulteriormente a circa 20/cm2 sul palmo. I recettori profondi (di Ruffini e di Pacini) sono meno numerosi (densità pari a circa 20/ cm2 per entrambi i tipi), senza notevoli variazioni fra polpastrelli e palmo della mano. In altre parti della cute la densità dei recettori tattili è molto inferiore rispetto a quella delle regioni palmari. La fig. 2, in alto e al centro, fornisce uno schema dei recettori tattili presenti nella cute glabra e in quella pelosa.
Poiché dà origine a più terminali, una singola fibra nervosa proveniente dalla cute convoglia in genere informazioni da più recettori tattili, ma tutti dello stesso tipo. Pertanto in linea di principio le sensazioni tattili sono riconducibili alle attività di quattro canali afferenti paralleli, corrispondenti ai quattro tipi di recettori. Registrando i potenziali d'azione lungo una fibra afferente durante vari tipi di stimolazione tattile, è possibile individuare la selettività di risposta di un determinato tipo di recettore (e quindi del canale afferente corrispondente) e accertare le relazioni fra diversi parametri dello stimolo e le caratteristiche della risposta. Inoltre la stimolazione elettrica di singole fibre afferenti identificate nell'uomo permette di ottenere descrizioni soggettive delle sensazioni tattili riferibili alle attività dei diversi canali afferenti. Si distinguono anzitutto recettori tattili a lento e a rapido adattamento. A fronte di una deformazione della cute invariante nel tempo, la risposta delle fibre collegate con recettori a lento adattamento persiste, seppur con una modesta riduzione temporale della sua intensità, durante tutto il periodo di stimolazione. Per contro, le fibre collegate con recettori a rapido adattamento non rispondono continuativamente, ma solo all'inizio e alla fine di tale stimolazione.
I corpuscoli di Merkel e di Ruffini sono classificabili come recettori a lento adattamento, mentre quelli di Meissner e di Pacini sono classificabili come recettori a rapido adattamento (fig. 2 in basso). L'adattamento rapido è attribuibile al fatto che le componenti non nervose del corpuscolo di Meissner e del corpuscolo di Pacini trasmettono ai loro terminali nervosi solo le variazioni della pressione esercitata sulla cute, agendo d'altro canto come uno schermo che protegge i terminali stessi da una deformazione cutanea statica. A dimostrazione dell'importanza delle strutture cellulari accessorie per l'adattamento, il terminale nervoso di un corpuscolo di Pacini si rivela capace di rispondere in maniera continuativa a una pressione statica diretta soltanto quando venga denudato dei suoi rivestimenti connettivali. Un'altra differenza fra i diversi tipi di recettori è costituita dalle dimensioni del campo recettivo di ciascuna fibra afferente, definito come l'area di cute la cui deformazione attiva la fibra stessa. Le fibre in rapporto con i recettori superficiali hanno campi recettivi piccoli, con diametro trasverso inferiore a 5 mm, e margini netti. Al contrario, i recettori profondi hanno campi recettivi grandi (per es., tutta la superficie palmare di un dito o metà del palmo) e margini sfumati.
Specializzazioni funzionali dei recettori tattili
I corpuscoli di Merkel sono selettivamente sensibili a deformazioni locali della cute causate dal contatto con punte, spigoli e superfici curve. Essi sono fondamentali per la percezione della forma e delle caratteristiche di superficie (rugosa o liscia, piatta o curva) degli stimoli tattili e quindi per il riconoscimento non visivo degli oggetti. Usando i polpastrelli delle tre dita intermedie, si può discriminare una superficie piatta da una concava o convessa con un raggio di curvatura di 2 m, una prestazione interamente attribuibile ai recettori di Merkel. I corpuscoli di Meissner sono sensibili a vibrazioni di bassa frequenza generate dal contatto attivo diretto con gli oggetti e hanno grande importanza per il controllo della forza con cui un oggetto viene afferrato per essere sollevato. La forza della presa digitale o manuale varia automaticamente non solo con il peso, ma anche in funzione della forza di attrito fra cute e oggetto, forza che dipende dalle caratteristiche di superficie dell'oggetto stesso. Un oggetto liscio viene afferrato per il sollevamento con una forza prensile maggiore di quella con la quale viene afferrato un oggetto rugoso di pari peso, meno scivoloso.
L'informazione tattile necessaria per questa regolazione della forza prensile è abolita dall'applicazione sulla superficie cutanea di un anestetico locale che inattiva selettivamente o primariamente proprio i recettori di Meissner a causa della loro massima vicinanza alla superficie stessa.
I corpuscoli di Pacini sono straordinariamente sensibili alle vibrazioni ad alta frequenza applicate tangenzialmente alla cute, tanto che i più sensibili possono essere attivati da ampiezze di vibrazione pari a soli 10 nm. Si ritiene che essi siano utilizzati quando si eseguono esplorazioni tattili a distanza dal corpo con l'intermediazione di oggetti tenuti in mano, come per esempio il bastone di un cieco. Le sensazioni tattili così generate, che di fatto hanno carattere prevalentemente vibratorio, sono localizzate soggettivamente non sulla mano che tiene il bastone, ma sulla punta del bastone stesso, che entra così a far parte dello schema corporeo. I corpuscoli di Ruffini sono attivati da tensioni tangenziali nella cute che probabilmente contribuiscono alla propriocezione della mano in movimento tramite la segnalazione continua delle variazioni locali di tensione nella cute palmare deformata dai movimenti manuali stessi. La stimolazione elettrica delle fibre connesse con i recettori di Ruffini non dà origine a sensazioni tattili, e pertanto si ritiene che anche la stimolazione naturale di questi recettori non partecipi all'esperienza cosciente, bensì al controllo afferente della postura e dei movimenti della mano. Infine i recettori associati ai peli sono attivati dalla flessione dei peli stessi, che agisce deformando il terminale nervoso aderente al follicolo. Pertanto la stimolazione di questi recettori avviene, con corrispondenti sensazioni tattili, anche in assenza di contatto diretto fra stimolo e superficie cutanea. Poiché il comportamento dei recettori associati ai peli è per vari aspetti simile a quello dei corpuscoli di Meissner, essi vengono considerati come i recettori superficiali a rapido adattamento della cute pelosa.
Psicofisica del tatto
Il senso del tatto può essere analizzato quantitativamente e in isolamento da altri sistemi di senso nelle condizioni artificiali del laboratorio sperimentale o dell'esame clinico. Le abilità tattili comunemente esaminate sono la soglia di sensibilità per il rilevamento di stimoli tattili, la localizzazione alla cieca degli stimoli stessi, l'acuità o capacità di risoluzione tattile e la sensibilità vibratoria, o pallestesia, che include sia il semplice rilevamento sia la discriminazione di stimoli vibratori della cute.
I dati psicofisici ed elettrofisiologici ottenuti da esperimenti condotti su soggetti umani e su macachi indicano che la cute, analogamente alla retina, possiede un sistema relativamente poco sensibile ma con alta capacità di risoluzione spaziale e un sistema relativamente molto sensibile ma con bassa capacità di risoluzione spaziale. I recettori di quest'ultimo sistema sono i corpuscoli di Meissner, in grado di rispondere ad avvallamenti della cute di pochi μm di profondità, ma incapaci di risolvere la posizione di uno stimolo entro i loro campi recettivi, aventi un diametro trasverso di 3÷5 mm. Un singolo potenziale d'azione convogliato da una fibra connessa con un recettore di Meissner riesce a evocare una sensazione di toccamento, mentre una stimolazione elettrica ripetitiva della stessa fibra causa una sensazione di vibrazione a bassa frequenza. I recettori di Merkel sono quattro volte meno sensibili di quelli di Meissner, ma la loro elevata capacità di risolvere la posizione dello stimolo entro il campo recettivo consente le massime prestazioni di acuità tattile. L'acuità tattile può essere misurata come la minima distanza percepibile fra due stimoli tattili, oppure con la lettura tattile di lettere in rilievo di varie dimensioni impresse o mosse sulla cute, oppure ancora con la discriminazione tattile dell'orientamento spaziale di griglie a fini barre parallele separate da distanze variabili. Con questi vari metodi la massima capacità di risoluzione spaziale si riscontra sulle labbra (0,5 mm) e sui polpastrelli (1 mm); essa diminuisce progressivamente con l'età per la perdita fisiologica di recettori tattili, tanto che negli ottantenni la soglia di risoluzione spaziale sui polpastrelli si innalza fino a 3 mm. La stimolazione elettrica ripetitiva di una fibra connessa con i recettori di Merkel causa una sensazione di contatto ma non una sensazione vibratoria.
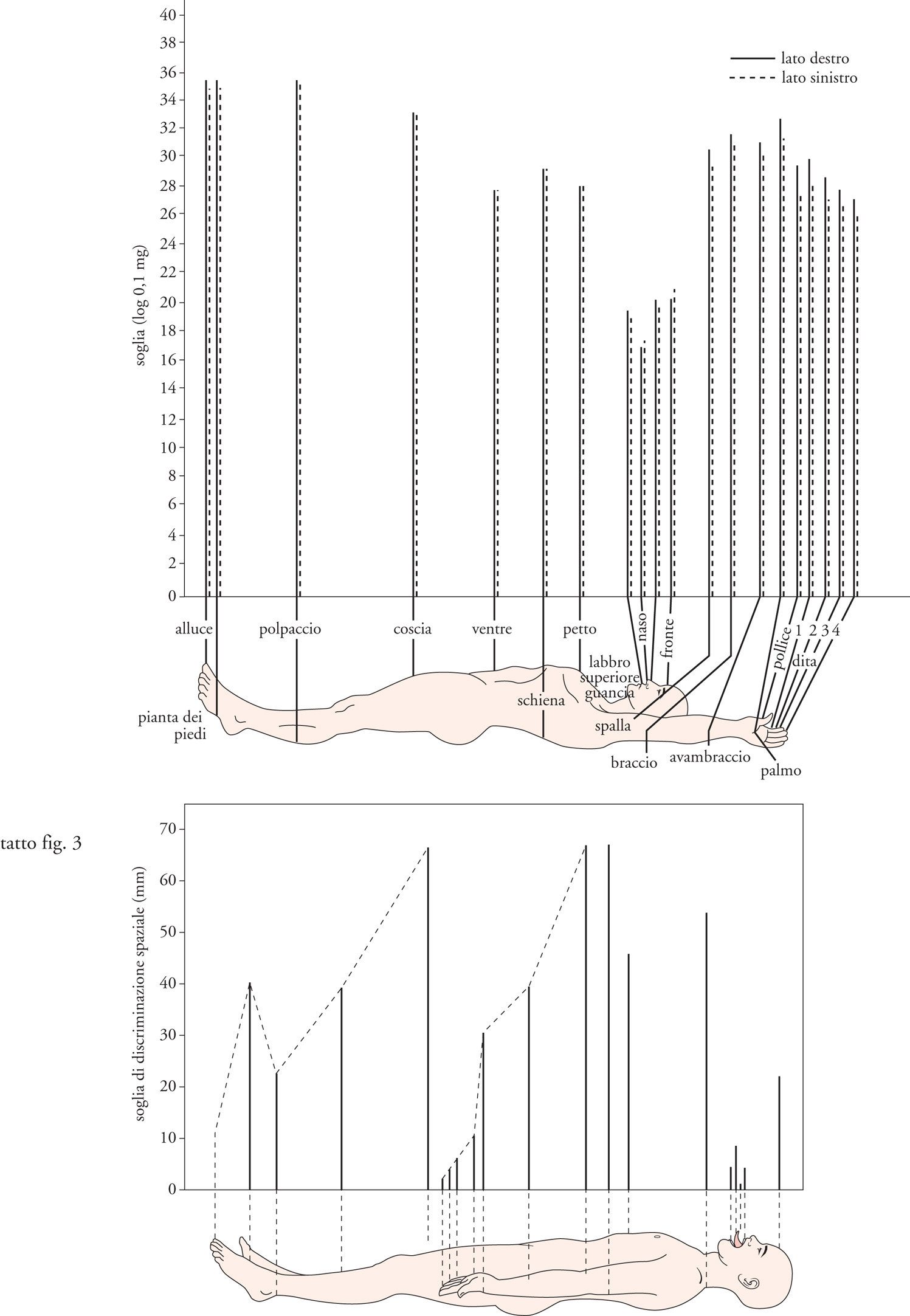
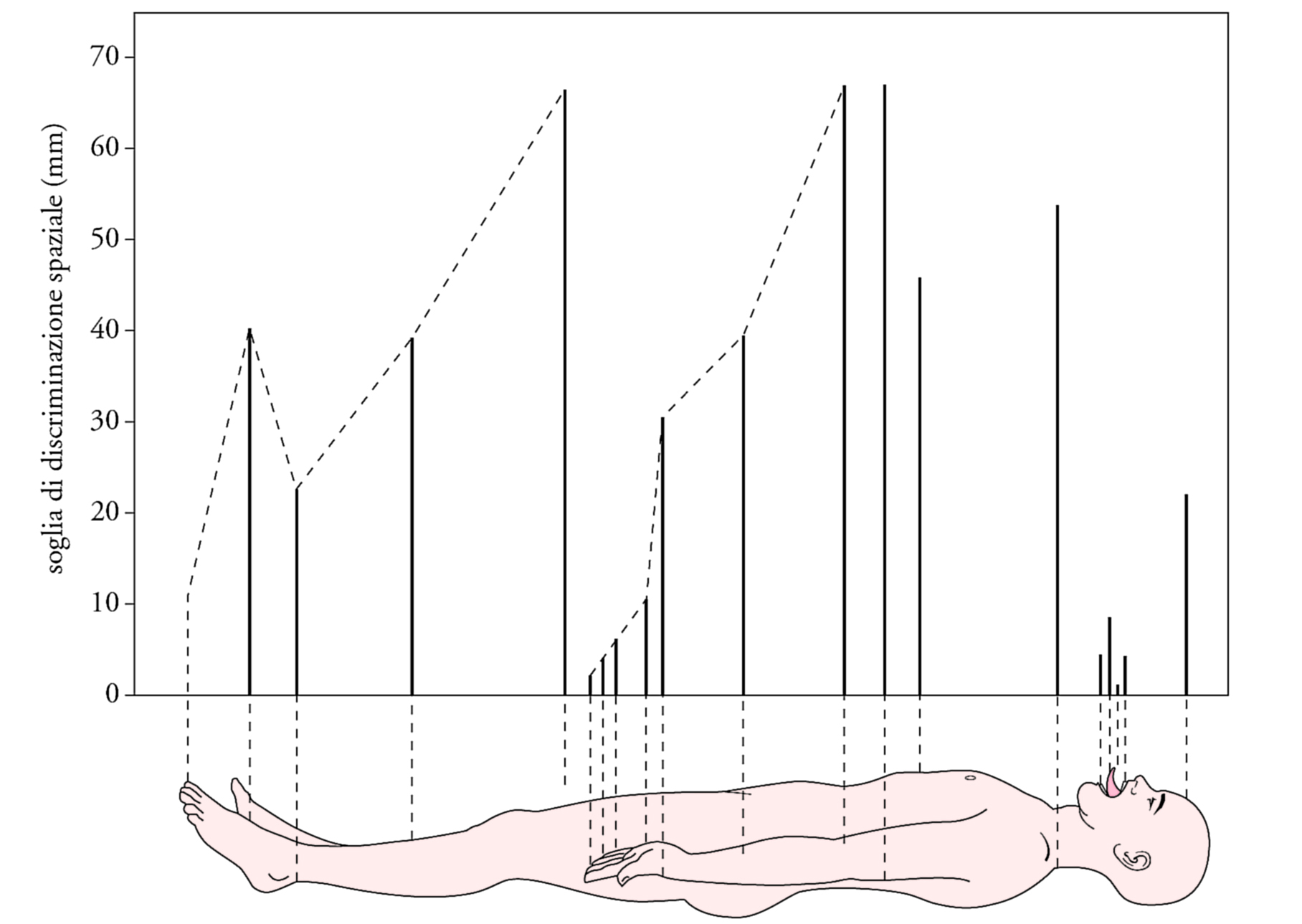
Le figg. 3 e 4 mettono a confronto le variazioni in diverse zone cutanee della soglia per il rilevamento degli stimoli tattili con quelle dell'acuità tattile. Le differenti variazioni regionali fra le due funzioni sono spiegate al meglio dall'esistenza di due sistemi recettoriali differenziati, uno (Meissner) specializzato nella sensibilità e l'altro (Merkel) nell'acuità tattile. Tradizionalmente l'esame clinico del senso tattile vibratorio si esegue applicando alla cute il piede di un diapason oscillante a diverse frequenze. Si distingue una sensazione di fluttuazione a bassa frequenza (massima sensibilità attorno ai 50 Hz) da una sensazione vibratoria ad alta frequenza (massima sensibilità attorno ai 250 Hz). La sensazione di fluttuazione è dovuta ai recettori di Meissner, come dimostra la sua selettiva abolizione in seguito ad anestesia della superficie cutanea. La sensazione di vibrazione ad alta frequenza, non abolita dall'anestesia cutanea superficiale, è dovuta invece alla stimolazione dei recettori di Pacini. Alle rispettive frequenze ottimali, la soglia di ampiezza per il rilevamento di uno stimolo vibratorio è circa mille volte più bassa per i recettori di Pacini (10 nm) che per quelli di Meissner (10 μm). Sulla cute glabra della mano è possibile discriminare ampiezze o frequenze dello stimolo vibratorio che differiscono fra di loro solo del 5÷10%.
Vie e centri nervosi del tatto
I terminali dei recettori del tatto appartengono a fibre nervose rivestite di mielina che perdono questa guaina in vicinanza della loro terminazione nella cute o nelle mucose. Si tratta di fibre con un diametro di 5÷15 μm e una velocità di conduzione di 35÷75 msec, ciascuna costituita dalla branca periferica dell'assone a T di neuroni tattili di prim'ordine. Fra questi neuroni, quelli che convogliano le informazioni relative alla sensibilità tattile dalla nuca ai piedi hanno sede nei gangli radicolari spinali, mentre quelli che convogliano le informazioni relative alla sensibilità tattile della faccia e della cavità orofaringea risiedono nei gangli di senso di quattro nervi cranici (trigemino, facciale, glossofaringeo e vago). Le fibre formate dalle branche periferiche degli assoni dei neuroni tattili dei gangli radicolari decorrono nelle radici posteriori del midollo, nei nervi spinali e infine nei nervi cutanei, mentre le fibre dei neuroni dei gangli encefalici decorrono nei rami cutanei o mucosi dei rispettivi nervi cranici.
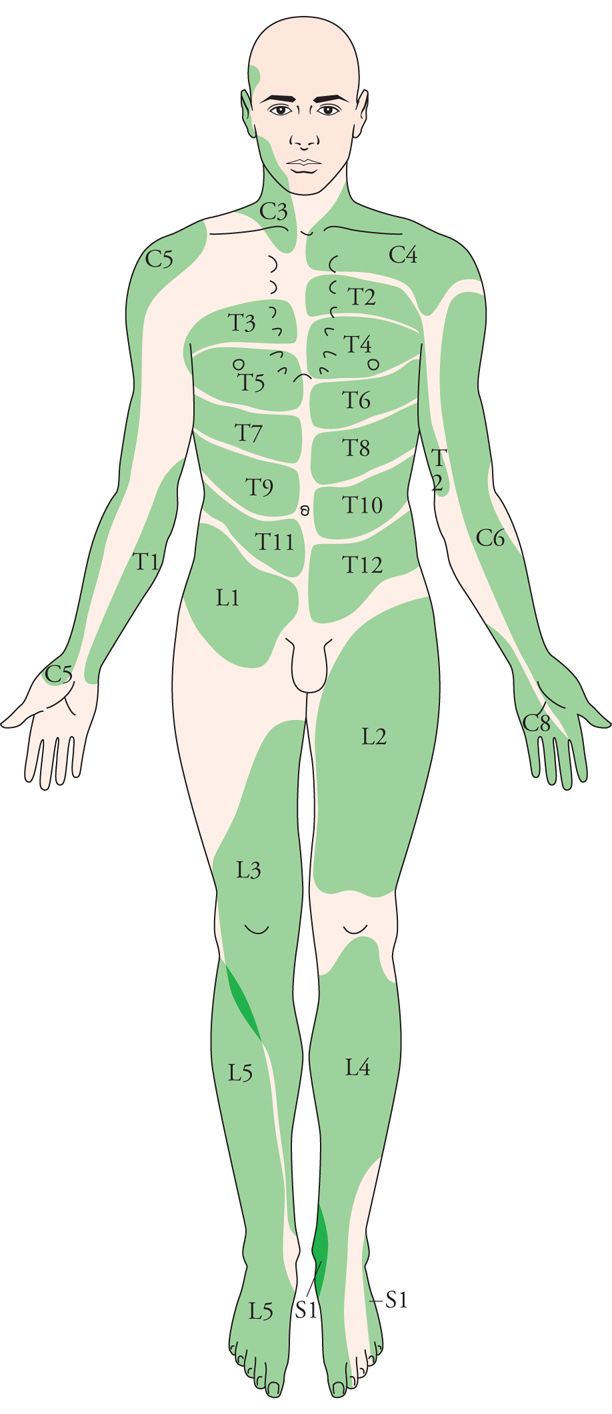
Le fibre tattili afferenti che decorrono in ciascuna radice dorsale del midollo spinale portano informazioni da una striscia di cute detta 'dermatomero'. A ciascuna radice corrisponde un dermatomero, e la sezione o l'applicazione di un anestetico locale a una singola radice produce la perdita della sensibilità tattile, dolorosa e termica nel dermatomero corrispondente. Va però rilevato che i confini fra i dermatomeri non sono netti, per l'esistenza di zone di sovrapposizione e per il fatto che la posizione dei confini varia a seconda della sensibilità esaminata (fig. 5). I dermatomeri della faccia corrispondono alle tre branche del nervo trigemino, la sensibilità tattile della cute del padiglione auricolare e del meato acustico esterno è servita da rami del facciale e del vago, quella della cavità orofaringea da rami del trigemino, del glossofaringeo e del vago. Le fibre costituite dalle branche centrali dei neuroni tattili spinali di prim'ordine penetrano nel midollo e ascendono nel cordone posteriore dello stesso lato assieme a fibre provenienti da recettori profondi per la propriocezione (fusi neuromuscolari, recettori tendinei e articolari). In una sezione cervicale del midollo si può constatare che le fibre provenienti dai segmenti sacrali e lombari occupano la parte mediale del cordone, formando il fascicolo gracile, mentre quelle provenienti dai segmenti toracico e cervicale decorrono più lateralmente e formano il fascicolo cuneato. Alcuni rami collaterali delle fibre dei due fascicoli terminano nel corno dorsale del midollo, ma i rami principali sono destinati a raggiungere il tronco dell'encefalo per terminare nei nuclei omonimi del midollo allungato dello stesso lato. Da questi nuclei parte un sistema di fibre, detto lemnisco mediale, che incrociandosi poco dopo la sua origine raggiunge il nucleo ventro-postero-laterale del talamo. A sua volta tale nucleo proietta alla corteccia della circonvoluzione postcentrale del lobo parietale dello stesso lato, che contiene le aree riceventi primarie per la sensibilità somatica, tatto incluso.

La rappresentazione del tatto nella corteccia cerebrale è caratterizzata da una somatotopia particolarmente manifesta nell'homunculus sensitivo, mappabile con vari metodi sulla circonvoluzione postcentrale. La stimolazione elettrica di un determinato punto di questa corteccia evoca, in pazienti sottoposti a interventi neurochirurgici in anestesia locale, sensazioni tattili riferibili a un determinato punto della superficie corporea del lato opposto, secondo la disposizione ordinata rappresentata nella fig. 6. Nell'homunculus sensitivo, la grandezza sproporzionata della faccia e delle mani rispetto al resto del corpo riflette la maggiore densità dei recettori tattili in queste regioni somatiche. In realtà la circonvoluzione postcentrale contiene quattro aree distinte in base alla costituzione cellulare (citoarchitettonica), disposte in una sequenza anteroposteriore e denominate 3a, 3b, 1 e 2 nella classificazione di Brodmann. In ciascuna di queste aree è presente una mappa ordinata della metà controlaterale del corpo. L'area 3a riceve informazioni dai fusi neuromuscolari ed è quindi considerata un'area non tattile ma propriocettiva, mentre le altre tre aree partecipano in diversa maniera alla rappresentazione del tatto. Le aree 3b e 2 sono esclusivamente tattili, e di fatto l'area 3b viene considerata la prima stazione corticale del senso del tatto. In ciascuna regione della mappa somatotopica di quest'area alcuni neuroni rispondono selettivamente a stimoli dei recettori tattili a rapido adattamento (Meissner e Pacini), mentre altri rispondono selettivamente ai recettori tattili a lento adattamento (Merkel). Non sono state descritte risposte corticali attribuibili ai recettori di Ruffini. I neuroni caratterizzati dai due tipi di risposta in base all'adattamento sono organizzati in aggregati verticali con diametro trasverso di circa 0,6 mm, che si alternano ordinatamente nello spessore della corteccia, a dimostrazione del fatto che in quest'area le proiezioni da differenti recettori tattili vengono mantenute segregate. Nelle aree 1 e 2 avviene già un'integrazione dei segnali trasmessi dai vari tipi di recettori somatici, grazie alla convergenza di informazioni ricevute dalle aree 3a e 3b. I neuroni dell'area 2 rispondono a stimoli combinati dei recettori tattili a rapido adattamento (Meissner e Pacini), e quelli dell'area 1 a stimoli combinati di recettori tattili e propriocettori muscolari, articolari e tendinei, fornendo presumibilmente una prima base per la stereognosi. Ulteriori elaborazioni dei segnali tattili avvengono in altre regioni corticali, grazie alle proiezioni delle aree della circonvoluzione postcentrale ad altre aree parietali, all'insula e al lobo frontale. Le mappe corticali sensitive (come anche quelle motorie) non sono organizzazioni stabili, ma possono modificarsi considerevolmente nel corso della vita per azione dell'esperienza. Nelle mappe tattili corticali, la rappresentazione di un dito si estende a spese della rappresentazione di altre regioni somatiche se quel dito viene impiegato a lungo in compiti di discriminazione tattile. Riorganizzazioni delle mappe corticali per la sensibilità somatica spiegano anche il fenomeno dell'arto fantasma, consistente in un complesso di sensazioni spesso dolorose, ma anche tattili, localizzate soggettivamente in una parte del corpo inesistente perché amputata, o privata dei nervi di senso. Le lesioni selettive delle vie tattili centrali o delle aree corticali con funzioni tattili producono vari disturbi della sensibilità tattile, soprattutto a carico dei suoi aspetti più fini come la localizzazione precisa e la capacità discriminativa. Va però sottolineato che lesioni anche estese delle vie tattili, per esempio una sezione totale dei cordoni posteriori del midollo spinale, o una lesione delle aree della circonvoluzione postcentrale da un lato, non aboliscono mai completamente la sensibilità tattile nelle regioni somatiche corrispondenti, dato che persiste un'imperfetta capacità di rilevare stimoli e di localizzarli, sia pure grossolanamente. Ciò suggerisce che esistano proiezioni centrali del senso del tatto che decorrono non nei cordoni posteriori, ma nel sistema spinotalamico assieme alle fibre della nocicezione e della termocezione. Esistono inoltre proiezioni non crociate che portano alla corteccia di un lato informazioni tattili provenienti dalla metà del tronco e dalle regioni prossimali degli arti dello stesso lato.
Bibliografia
Berlucchi, Aglioti 1997: Berlucchi, Giovanni - Aglioti, Salvatore M., The body in the brain: neural bases of corporeal awareness, "Trends in neurosciences", 20, 1997, pp. 560-564.
Hsiao 2003: Hsiao, Steven S. - Yoshioka, Takashi - Johnson, Kenneth O., Neural basis for somesthesis, in: Encyclopedia of cognitive science, edited by Lynn Nadel, London, Nature Publishing Group-Macmillan, 2003, IV, pp. 92-96.
Johnson 2001: Johnson, Kenneth O., The roles and functions of cutaneous mechanoreceptors, "Current opinion in neurobiology", 11, 2001, pp. 455-461.
Klatzky, Lederman 2003: Klatzky, Robert L. - Lederman, Susan J., Haptic perception, in: Encyclopedia of cognitive science, edited by Lynn Nadel, London, Nature Publishing Group-Macmillan, 2003, III, pp. 508-512.
Mountcastle 2005: Mountcastle, Vernon B., The sensory hand: neural mechanisms of somatic sensation, Cambridge (Mass.), Harvard University Press, 2005.