
Sviluppo embrionale
Sviluppo embrionale
di Conrad H. Waddington, Antonio García-Bellido
SVILUPPO EMBRIONALE
Biologia dello sviluppo di Conrad H. Waddington
Sommario: 1. Introduzione. 2. I concetti della biologia dello sviluppo: a) concetti generali; b) le basi dello sviluppo; c) cenni storici; d) concetti di controllo e integrazione. 3. Sviluppo ed evoluzione: a) mutazione e selezione; b) assimilazione genetica ed ereditarietà dei caratteri acquisiti. □ Bibliografia.
1. Introduzione
Si dice comunemente che tra tutti i grandi problemi classici della biologia i meno compresi sono quelli inerenti alla biologia dello sviluppo. Recentemente alcuni biologi molecolari, forti delle loro scoperte concernenti gli schemi dei processi di controllo delle attività geniche nei Batteri, hanno suggerito che il problema dello sviluppo e del differenziamento degli organismi superiori debba essere affrontato in tali termini. Forse questo ottimismo deriva dalla mancanza di una chiara visione teorica della situazione. Solo quando la teoria è adeguata è possibile porsi correttamente i problemi. Attualmente vi sono parecchi punti della biologia dello sviluppo nei quali non ci è possibile ottenere che un modesto risultato; tuttavia siamo abbastanza vicini alla formulazione di una teoria soddisfacente per intuire che i problemi reali che sorgono saranno di gran lunga più difficili da risolvere di quanto spesso ritengano i meno esperti del campo.
Ragionando in termini di biologia dello sviluppo, bisogna considerare diversi livelli di teorizzazione. Forse quattro livelli sono sufficienti per poter procedere: a) metateorie, che hanno lo scopo di decidere su quali argomenti è vantaggioso teorizzare; b) teorie vere e proprie, che cercano di descrivere in termini generali la struttura logica dei problemi scelti e di definire concetti che forniscano un appropriato linguaggio con il quale poter discutere i problemi stessi; c) ipotesi generali, che specificano i tipi di meccanismi invocati per produrre queste strutture logiche; d) ipotesi particolari, che descrivono in quali modi i vari elementi di questo insieme di meccanismi possibili sono implicati nei casi particolari.
2. I concetti della biologia dello sviluppo
a) Concetti generali.
Il termine ‛sviluppo' può essere usato, nel suo senso più generale, in riferimento a qualsiasi processo caratterizzato da un cambiamento progressivo. In questo senso le più moderne teorie filosofiche postulano che tutte le entità esistenti, sia nel mondo fisico sia in quello biologico, sono caratterizzate da un qualche tipo di sviluppo. Tali punti di vista riportano agli inizi della filosofia. Tra i filosofi presocratici della Grecia ionica, cinquecento anni prima di Cristo, alcuni, come Eraclito, esprimevano l'idea che l'essenza di tutte le cose naturali fosse il divenire o cambiamento. Altri, al contrario (Democrito fu forse il primo), ritenevano che il mondo fosse costituito da combinazioni varianti di atomi, particelle queste inalterabili e non soggette a cambiamento o sviluppo. La scienza europea del primo periodo dopo il Rinascimento può essere considerata come dominata da queste vedute atomistiche, che raggiunsero il massimo sviluppo nel periodo che va dalle leggi della fisica di Newton alla teoria atomica della fisica proposta da Dalton, verso la metà del XIX secolo. Questo modo di interpretare la realtà non fu mai in facile accordo con le osservazioni compiute dai biologi, e negli ultimi cento anni una serie di scoperte nel campo delle scienze fisiche hanno riportato l'opinione dei filosofi verso la teoria eraclitea, che dava la massima importanza al divenire e allo sviluppo. Si è dimostrato che l'atomo, che a Dalton sembrava inalterabile, è scindibile e che mantiene la sua identità solo attraverso processi di interazione tra le particelle subatomiche che lo compongono. Queste stesse particelle devono essere considerate, sotto certi aspetti, non come ‛cose', ma come onde, vale a dire come processi. La teoria della relatività di Einstein mostra ancora che non è possibile assumere un concetto di spazio senza tempo nel quale si trovino cose che non cambiano: tempo e spazio sono uniti in una continuità spazio-tempo, continuità che implica che tutte le cose sono coinvolte nel tempo, cioè sono in sviluppo.
Gli organismi viventi mostrano ai biologi processi di cambiamento di tipi assai svariati. Sebbene tutti questi cambiamenti possano essere in qualche modo considerati come sviluppo, i biologi ritengono conveniente classificarli in un certo numero di ampie categorie, e usare il termine sviluppo solo per una di queste. In questo senso convenzionale, lo sviluppo biologico puo essere definito come una serie di cambiamenti progressivi e non ripetitivi che avvengono durante la vita di un organismo. Il nocciolo di questa definizione sta nel contrapporre lo sviluppo sia ai cambiamenti puramente ripetitivi che sono propri del funzionamento della macchina vivente, come l'ingestione, la digestione e l'assimilazione degli alimenti, l'assorbimento e l'utilizzazione dell'ossigeno e cosi via (cambiamenti questi che vanno sotto il nome di ‛metabolismo'), sia ai cambiamenti a più lungo termine che sono anch'essi non ripetitivi, ma che coinvolgono una sequenza di generazioni di individui, e che vanno sotto il nome di evoluzione. Come nel caso di definizioni più formali, queste distinzioni non possono essere conservate nel loro senso più completo quando siano applicate a un mondo reale. Ad esempio, negli organismi viventi molto semplici, quali i Virus e anche i Batteri, è difficile operare una distinzione tra metabolismo e sviluppo, dato che l'attività metabolica di una particella virale consiste in poco più dello sviluppo di una o più nuove particelle. E ancora, in certi casi il concetto di organismo singolo con una storia definita può essere difficilmente applicabile, come, ad esempio, nelle piante che si riproducono per divisione vegetativa, in cui cioè da un frammento di una pianta si sviluppa un'altra pianta completa. Qui è difficile interpretare la distinzione tra sviluppo ed evoluzione. Comunque, le possibilità di discussione che sorgono da questi casi speciali non sono sufficienti in alcun modo a invalidare l'uso generale, nell'ampio campo della biologia, delle distinzioni convenzionalmente fatte.
b) Le basi dello sviluppo
Come è noto, tutti gli organismi, compresi i più semplici come i Virus, sono costituiti da due componenti. La distinzione tra questi due componenti è stata sottolineata per la prima volta da A. Weismann alla fine del XIX secolo. Egli li chiamò ‛plasma germinale' e ‛soma'. Il primo è l'elemento essenziale che viene trasferito da una generazione alla successiva, per esempio nelle uova e nello sperma degli animali; il soma è tutto il corpo, che si viene a costituire con lo sviluppo dell'animale. In termini più moderni, il plasma germinale di Weismann è stato identificato con il DNA ( acido desossiribonucleico) che porta, codificate nella complessa struttura della sua molecola, le istruzioni necessarie per la sintesi degli altri componenti dell'organismo e per la loro organizzazione in appropriate strutture. L'insieme di tutti gli altri componenti (proteine, grassi, carboidrati, ecc.), organizzati in modo da formare un organismo con funzione metabolica, costituisce il soma (cioè il corpo). Con il termine ‛sviluppo biologico' noi intendiamo proprio quel processo di trasferimento delle istruzioni contenute nel DNA che costituisce il genotipo. Bisogna comunque ricordare che, in ogni organismo, le istruzioni possono essere trasferite solo mediante un appropriato meccanismo operativo. Negli organismi biologici la prima fase di questo meccanismo è attuata dalla cellula che porta le istruzioni alla generazione successiva: cioè negli animali e nelle piante dall'uovo fecondato, nei Virus dalla cellula infettata. Negli organismi che hanno più di un minimo grado di complessità, questo meccanismo operativo si modifica appena le istruzioni genetiche entrano gradualmente in opera e nuovi meccanismi di sintesi proteica vengono posti in condizione di funzionare. Il problema fondamentale della biologia dello sviluppo è di capire le interconnessioni che avvengono tra le istruzioni genetiche e i meccanismi attraverso i quali queste istruzioni vengono realizzate.
Nel linguaggio genetico il termine ‛genotipo' è usato per indicare le istruzioni ereditarie che vengono trasmesse da una generazione all'altra, mentre si chiama ‛fenotipo' l'organismo prodotto dall'elaborazione di queste istruzioni. Si può dire, quindi, che lo sviluppo biologico consiste nella produzione di ‛fenotipi'. La formazione del fenotipo di un organismo di una data generazione dipende dal funzionamento di una parte dei fenotipo della precedente (cioè la cellula uovo), nell'ambito del meccanismo che dà inizio all'interpretazione delle istruzioni contenute nel genotipo del nuovo organismo.
c) Cenni storici
Il tentativo di stabilire una scienza causale dello sviluppo embriologico costituisce una branca relativamente recente della biologia. Nell'ultima parte del XIX secolo, numerosi biologi europei (e principalmente tedeschi) misero in evidenza l'importanza di raggiungere alcune conoscenze causali dei processi di sviluppo. I più eminenti tra essi furono W. His, W. Roux e H. A. E. Driesch. I meccanismi da essi studiati erano necessariamente molto ipotetici, dal momento che era straordinariamente difficile condurre esperimenti che fossero realmente in grado di mettere in evidenza qualche interazione causale. I risultati sperimentali ottenuti dai primi ricercatori rimasero essenzialmente descrizioni fenomenologiche: cioè portarono, per così dire, alla luce del giorno certi fenomeni, senza cogliere i processi causali che li avevano determinati. Sia Roux che Driesch, ad esempio, misero a punto esperimenti nei quali asportavano o uccidevano uno dei due blastomeri prodotti dall'uovo fecondato. Quando Roux compì questa prova con un uovo di rana, il blastomero restante sviluppò mezzo embrione, esattamente ciò che sarebbe avvenuto se l'esperimento non fosse mai stato effettuato. In realtà ciò non mostrava nulla che non si sapesse già. Quando Driesch fece un esperimento simile con un uovo di riccio di mare, ciascuna metà dell'uovo sviluppò una larva completa. Questo era un nuovo sorprendente fenomeno, ma anche in questo caso la semplice osservazione non offriva alcun appiglio per la sua spiegazione.
Nei primissimi anni del nuovo secolo fiorirono molte attività in questo campo, peraltro senza troppi progressi. Una fiorente scuola americana di ricercatori studiò i primi stadi di sviluppo delle uova di Invertebrati. Diretti da E. G. Conklin, essi descrissero in dettaglio le ‛genealogie cellulari' attraverso le quali le varie parti di un uovo in divisione formano i vari organi della larva. Inoltre, cosa più importante, Conklin e i suoi collaboratori descrissero varie regioni di citoplasma identificabili all'interno dell'uovo fecondato, regioni che si dimostrarono necessarie per lo sviluppo di particolari organi o tessuti. Non era del tutto chiaro come questi ‛ooplasmi' agissero nè quali fossero le caratteristiche operative veramente importanti delle regioni distinguibili al microscopio per il colore o per altre proprietà superficiali. Comunque fu raggiunta una conclusione fondamentale. In Inghilterra Boycott e Diver studiarono la genetica di una chiocciola, Limnaea peregra, della quale esistono due forme differenti, una con avvolgimento a spirale sinistrorso e l'altra con avvolgimento destrorso. Questi autori mostrarono che le due forme, topologicamente distinte, possono essere riconosciute fin dai primissimi stadi di divisione e che questa proprietà dipende quasi certamente dalla distribuzione delle regioni ooplasmatiche nell'uovo al momento della fecondazione. Boycott e Diver trovarono molto problematica la spiegazione di questa ereditarietà, ma A. Sturtevant chiari che i loro risultati erano facilmente interpretabili se si suppone che la direzione di avvolgimento dipenda dalla struttura della cellula uovo da cui l'individuo si sviluppa, e che questa struttura sia a sua volta determinata non dai geni contenuti nell'uovo fecondato ma piuttosto dai geni contenuti nell'ovario materno nel quale l'uovo raggiunge la maturità. Questa osservazione fu ripresa da Th. H. Morgan come prova conclusiva per una teoria generale, cioè che l'ordinamento spaziale degli ooplasmi dentro una cellula uovo sia in definitiva dipendente da geni, quelli dell'organismo materno, e non, come avrebbero voluto gli oppositori della genetica, un fenomeno indipendente da aggiungere alla genetica, prima di ottenere una completa spiegazione dei fattori di base da cui dipende lo sviluppo.
Un altro lavoro, più isolato, ma comunque di grande importanza per la formazione delle moderne concezioni della biologia, fu quello esposto da D'Arcy W. Thompson nel suo libro Growth and form. In quest'opera l'autore tracciò un approccio del tutto nuovo ai problemi della morfologia, e dimostrò che la forma delle cellule e la forma di alcuni animali e piante a semplice organizzazione assomigliano spesso a quelle di entità fisiche e possono essere descritte con le stesse formulazioni matematiche.
Egli non arrivò però a concludere che le forme organiche e inorganiche sono state prodotte attraverso gli stessi processi fisici. Infatti quando metteva a confronto la forma di un polipo con quella dello schizzo formato da una goccia che cade in una pozza d'acqua, era ovvio che le forze fisiche implicate non erano nè le stesse nè molto simili. Tuttavia era uguale la descrizione matematica della forma. Il lavoro di D'Arcy Thompson è stato importante non tanto perchè ha rivelato nuove conoscenze dei processi che realizzano una forma organica, ma piuttosto perché ha messo in rilievo l'importanza e la possibilità di applicare formulazioni matematiche a tipi di processi biologici che prima erano stati trattati solo con metodi di senso comune. Il suo lavoro non ebbe un seguito immediato, ma rimase un'opera base, che fu ripresa solo dopo molti anni.
Il principale centro del progresso della biologia dello sviluppo nei primi venticinque anni del secolo si ritrova in Germania. La tradizione di un'analisi sperimentale dei processi dello sviluppo era stata fondata da autori come Roux e Th. Boveri. All'inizio del Novecento H. Spemann cominciò lo studio di embrioni di Anfibio. Il suo grande merito fu di arrivare a formulare problemi ben definiti circa i meccanismi causali, in termini di interazioni tra parti differenti di un sistema in sviluppo. Il suo primo successo fu la dimostrazione che lo sviluppo del cristallino dipende da una interazione tra il calice ottico, che si origina dal cervello, e il sovrastante ectoderma. Trapiantando il calice ottico in una diversa posizione, mostrò che tale formazione, quando viene in contatto con un ectoderma che si trovi in un opportuno stadio di competenza, può indurre l'ectoderma stesso, che si sarebbe sviluppato in maniera del tutto diversa se non si fosse stabilito questo contatto, a sviluppare un cristallino. Concluse perciò che il calice ottico ‛induce' il cristallino.
Alla fine degli anni venti, Spemann e una sua allieva, H. Mangold, riuscirono a generalizzare questo concetto, dimostrando che può anche venir applicato alle primissime fasi del differenziamento dell'uovo di Anfibio, cioè quando appaiono i principali organi assili (sistema neurale, notocorda, somiti, ecc.). Questi autori dimostrarono definitivamente che il mesoderma può indurre lo sviluppo di organi assili (piastra neurale) in un qualsiasi ectoderma in un certo stadio di competenza, anche se questo ectoderma nel corso dello sviluppo non avrebbe mai dato origine a questi organi. L'importanza di tale risultato fu di fornire il primo processo causale controllabile sperimentalmente, identificato nello studio dello sviluppo. Ponendo un induttore in una posizione adatta in un embrione, si può controllare il modo in cui i tessuti circostanti si sviluppano. Questa fu la porta che apri la via allo studio completo dei processi di controllo dello sviluppo.
Spemann stesso, nelle prime fasi del suo lavoro, definì le sue entità causali solo in termini operativi, senza peraltro fornire un modello concettuale che spiegasse la loro azione. I suoi induttori od ‛organizzatori' furono definiti come quelle parti di tessuto che, trapiantate in certe situazioni particolari, producono certi risultati, i quali possono essere descritti attraverso le caratteristiche morfologiche e istologiche degli organi prodotti. Prima della metà di questo secolo, non si fece alcun serio tentativo per analizzare le sequenze causali in termini più analitici, cioè per mezzo della biochimica o della genetica.
d) Concetti di controllo e integrazione
Una delle più singolari caratteristiche di tutti i sistemi di sviluppo è la tendenza a dare un normale risultato finale anche se vengono prodotte nel sistema, nei primi stadi di sviluppo, lesioni o anomalie. In molti casi, e forse nella maggior parte, possono essere completamente compensate solo le lesioni prodotte durante un limitato periodo dello sviluppo. Si dice che, in questo periodo, il sistema è capace di regolazione, che è il termine generalmente usato per descrivere il fenomeno di ripristino delle condizioni normali.
La regolazione dello sviluppo è spesso indicata col termine di ‛omeostasi', concetto usato nelle scienze fisiche e preso in prestito dalla fisiologia. È questa una proprietà dei sistemi che persistono in uno stato di equilibrio e che, se alterati, tendono a ristabilire l'equilibrio iniziale. Esempi ben noti di sistemi omeostatici in fisiologia sono il contenuto di CO2 o il pH del sangue. Tali modelli, comunque, non danno una descrizione molto adeguata dei fenomeni che accadono nei sistemi in sviluppo. Un sistema in sviluppo, a priori, non rimane mai in uno stato di equilibrio inalterato, durante un dato periodo di tempo. Al contrario, un sistema in sviluppo cambia continuamente nel tempo, muovendosi in una definita ‛traiettoria temporale' da uno stadio iniziale, quale l'uovo fecondato, e attraverso vari stadi larvali giunge allo stadio adulto e infine alla senescenza. La regolazione che ha luogo in tali sistemi non è un ritorno all'iniziale equilibrio stabile, come accade nell'omeostasi, ma piuttosto una regolazione verso qualche stadio successivo della traiettoria temporale. Il termine più adatto per descrivere ciò non è ‛omeostasi', che indica il ritorno a una condizione statica, ma ‛omeorresi', che indica il ritorno a una condizione dinamica (Waddington, 1957).
Un'altra importante caratteristica fenomenologica dello sviluppo è che lo stato finale non è omogeneo o unitario, ma può essere suddiviso in un numero diverso di organi e di tessuti. L'intera traiettoria temporale finale, perciò, è la risultante di un dato numero di traiettorie componenti, che conducono all'uno o all'altro risultato finale distinguibile negli stadi di sviluppo successivi. Una scoperta importante dei primi esperimenti condotti su sistemi in sviluppo fu la dimostrazione che, almeno in molti casi, le differenti traiettorie temporali divergono le une dalle altre piuttosto bruscamente in un breve periodo di sviluppo; questa divergenza si presenta di solito prima che segni di differenza si rendano visibili al microscopio o siano rilevabili con altri mezzi d'indagine. L'esempio più importante e più significativo fu fornito dagli studi di Spemann sullo sviluppo dell'uovo di Anfibio al momento della gastrulazione. A quest'epoca l'embrione assume la forma di una sfera di cellule, cava all'interno, il cui emisfero inferiore sarà spinto all'interno (invaginato) per dare origine a mesoderma ed entoderma. L'emisfero superiore rimarrà alla superficie, formando un area ricoprente l'intero embrione, della quale all'incirca un terzo si svilupperà in sistema nervoso e il resto in epidermide. Spemann dimostrò sperimentalmente che, durante il periodo in cui avvengono questi movimenti morfogenetici di invaginazione ed espansione, avviene un processo attraverso il quale una parte dell'emisfero superiore si differenzierà in tessuto neurale, mentre l'altra parte darà luogo allo sviluppo dell'epidermide. Questo processo di ‛determinazione' delle vie di sviluppo avviene piuttosto precocemente, quando le cellule delle due regioni sembrano a tutta prima identiche. Che la determinazione sia avvenuta può essere in realtà dimostrato solo sperimentalmente. Prima che questa abbia avuto luogo, ogni porzione dell'emisfero può dare ongine o a tessuto neurale o a epidermide. Si può dimostrare ciò, ad esempio, trapiantando piccoli frammenti da una regione all'altra dell'embrione. Dopo che la determinazione è avvenuta, le cellule che erano situate nella zona neurale daranno origine a tessuto neurale in pressoché tutte quelle condizioni in cui possono vivere, ed è estremamente difficile, se non impossibile, indurle a dare qualche cosa di diverso. La stessa cosa, mutatis mutandis, accade per le cellule situate nella zona dell'epidermide.
Appena fu data questa dimostrazione e ne fu generalmente riconosciuta l'importanza, divenne chiaro che una corretta teoria dello sviluppo deve tener conto non solo dei processi attraverso i quali un sistema in sviluppo procede lungo la sua caratteristica traiettoria temporale, ma anche della natura dei processi attraverso i quali le traiettorie si allontanano le une dalle altre, e sono fissate o determinate nelle cellule in sviluppo.
Qualsiasi teoria sulla determinazione deve tener conto del fatto che uno stato determinato può essere trasmesso attraverso parecchie generazioni cellulari. Ciò era già risultato chiaro dagli esperimenti di Spemann, ma recentemente un esempio ancora più chiaro è stato fornito dal lavoro di E. Hadorn sulle cellule dei dischi immaginali di Drosophila. Questi sono piccoli raggruppamenti di cellule che si separano dal resto dell'embrione nei primi stadi di sviluppo e persistono attraverso tutta la vita larvale, differenziandosi poi verso caratteristiche adulte quando vengono stimolati dagli ormoni secreti nel periodo della pupazione (v. ormoni negli invertebrati). Questi ormoni non si ritrovano più nel corpo dell'insetto adulto, e Hadorn dimostrò che le cellule immaginali trapiantate nella cavità del corpo di un adulto subiscono molte divisioni cellulari senza peraltro mostrare alcun segno di differenziamento nei tessuti specifici dei corrispondenti organi adulti. Tuttavia, se le cellule che hanno subito molte divisioni si ritrapiantano infine in una larva pronta a diventare pupa, e quindi sotto il controllo degli ormoni secreti nello stato di pupa, iniziano immediatamente a differenziarsi. In quasi tutti i casi, un tessuto derivato da una cellula che normalmente avrebbe dato origine a un occhio si svilupperà ora in un occhio, un tessuto derivato da una cellula che avrebbe dato origine a un'ala svilupperà un'ala, e così via. Cioè, i tessuti hanno conservato, attraverso numerose divisioni cellulari, la determinazione a dare origine a quell'organo adulto al cui sviluppo avrebbero dato luogo in presenza di ormoni della pupazione.
Il nostro problema è quello di stabilire quale forma assuma questa determinazione. Come abbiamo visto, Spemann è stato in grado di mostrare, nelle uova di Anfibio da lui studiate, che le cellule determinate a prendere la via del differenziamento neurale sono sottoposte all'azione di qualche fattore derivante da cellule del mesoderma con le quali vengono in contatto attraverso il processo di invaginazione. Comunque, questo fatto è ben lontano dal rivelarci quale tipo d'influenza esercitino queste cellule mesodermiche, e su quali elementi delle cellule sottoposte a tale stimolo agisca questa influenza.
Tentativi per identificare l'agente della determinazione che proviene dal mesoderma di Anfibio non hanno ancora fornito una chiara identificazione di una particolare sostanza attiva o di altri agenti. Questi esperimenti hanno comunque prodotto un'importante conclusione generale: cioè che il processo di determinazione può aver luogo solo in un determinato periodo dello sviluppo, durante il quale le cellule dell'emisfero superiore dell'uovo di Anfibio sono come sospese tra le due possibilità di svilupparsi in tessuto neurale o in epidermide. Si dice che in questo periodo le cellule sono ‛competenti' per l'uno o per l'altro tipo di sviluppo. Mentre le cellule si trovano in questo stato, e solamente in questo, un insieme di agenti esterni può deviarle nell'una o nell'altra delle possibili vie di sviluppo. Tale situazione differisce da quella nella quale le cellule sono neutre o prive di caratteristiche e necessitano di un agente esterno che trasmetta loro la proprietà di diventare tessuto nervoso o epidermide: questo sta a significare che le cellule capaci di reagire richiedono informazioni o istruzioni dall'esterno. Tale non è la situazione caratteristica dello sviluppo biologico: sia in organismi più evoluti, quali gli Anfibi, sia in situazioni molto semplici, come l'induzione di enzimi nei Batteri, si trova che gli agenti estranei agiscono solo da innesco, determinando l'uno o l'altro processo. Tutte le informazioni necessarie per questi processi sono già contenute nelle cellule interessate.
Nei primi trent'anni di questo secolo è stata compresa l'esistenza di tali fenomeni di sviluppo. In tale periodo i biologi non avevano però ancora una chiara nozione dei concetti fondamentali con i quali lo sviluppo doveva essere spiegato. Gli embriologi tentavano di esprimere le loro osservazioni con termini non ben definiti, quali ‛potenze', ‛sostanze organo-formative', oppure facendo riferimento a proprietà cellulari che sono senz'altro reali, ma ovviamente di per se stesse piuttosto complesse e di natura certamente secondaria, quali l'adesione cellulare, la capacità di assorbimento selettivo dalle membrane cellulari e così via. Soltanto gradualmente i biologi dello sviluppo hanno compreso l'importanza di dimostrare attraverso la genetica che quasi tutte le istruzioni necessarie per la costruzione di un nuovo organismo sono contenute nei due genotipi che si fondono con la fecondazione. L'ulteriore piccola quantità di informazione, contenuta per esempio nella struttura dell'uovo, è essa stessa un prodotto delle informazioni genetiche fornite dalla madre, che produce l'uovo. Subito dopo Th. H. Morgan comprese l'importante concetto generale che le unità fondamentali, con le quali deve essere interpretato lo sviluppo, sono i geni; però riuscì a sviluppare tale concetto solo in maniera piuttosto generale, senza fornire molti esempi ben analizzati di meccanismi genetici che agiscono nello sviluppo. Negli anni trenta R. B. Goldschmidt e C. H. Waddington fornirono un'analisi soddisfacente di tali meccanismi. Poco dopo, la teoria secondo la quale geni specifici controllano la sintesi di particolari proteine ebbe un sostanziale e convincente sostegno dal lavoro di Beadle e Tatum e di altri autori sulla sintesi di enzimi in Neurospora. Senza dilungarci a descrivere lo sviluppo di questo lavoro, che appartiene ora alla biologia molecolare, si può sicuramente dire che questi esperimenti hanno ormai persuaso tutti i biologi che le unità essenziali dello sviluppo sono i geni. Il problema fondamentale della teoria dello sviluppo è, ora, capire come queste unità interagiscono l'una con l'altra per formare meccanismi più complessi, che determinano i comportamenti cellulari o tessutali con i quali ci appaiono i diversi tipi di sistemi in sviluppo.
Come abbiamo visto, uno dei caratteristici tipi di comportamento a questo livello più complesso è la costruzione di traiettorie temporali stabilizzate. Nello sviluppo di un dato tessuto od organo, quale il sistema neurale dei Vertebrati, un grandissimo numero di geni deve essere impegnato nel controllo della sintesi di particolari proteine. In certi casi, ad esempio nella formazione delle ali di Drosophila, è stata definitivamente dimostrata l'attività di almeno trenta o quaranta geni; ma certamente ne sono implicati molti di più, non si sa ancora se parecchie decine, parecchie centinaia od oltre. Comunque, si deve considerare che l'azione di tutti questi geni forma come una rete di interconnessioni, implicando parecchi tipi di meccanismi di controllo a feedback e di altri circuiti di interazione. Questi meccanismi fanno si che il sistema determini un risultato finale nel quale molte componenti sono presenti in concentrazioni stabilite precisamente. Inoltre il processo dello sviluppo che porta a questo risultato finale deve essere ‛tamponato o stabilizzato' nel senso che, se il processo viene allontanato in uno stadio precoce dal suo normale corso, tende all'omeorresi e si riporta ad alcuni stadi successivi della normale traiettoria. Queste traiettorie temporali tamponate sono state chiamate chreods (creodi; Waddington, 1957-1967), parola che significa ‛via obbligatoria'. La comprensione che le unità fondamentali dello sviluppo sono i geni e che un creode implica l'azione di decine, se non centinaia, di geni, ci porta a immaginare i creodi come strutture in uno spazio multidimensionale. Un matematico francese, R. Thom (1970), ha trattato questo concetto in termini di topologia analitica ed è cosi riuscito a sviluppare una teoria dei possibili tipi di creodi ramificati, una classe di fenomeni che chiama ‛catastrofi'. Egli pretende di aver dimostrato che sette, e soltanto sette, sono i tipi di creodi che possono esistere in un mondo definito da tre dimensioni di spazio e una di tempo. La comprensione che lo sviluppo biologico è fondamentalmente un'espressione di attività controllate di geni ha risolto finalmente una delle più antiche dispute filosofiche circa la natura dello sviluppo quella tra preformismo ed epigenesi. Il primo postulava che, all'inizio dello sviluppo, per esempio nell'uovo fecondato, il sistema contenesse già formato ciascun organo che doveva solo manifestarsi con lo sviluppo. La teoria dell'epigenesi, per contro, supponeva che le forme che si manifestavano successivamente venissero prodotte durante il corso dello sviluppo non già da un germe preesistente, bensì dall'interazione dei costituenti più semplici dell'uovo.
3. Sviluppo ed evoluzione
a) Mutazione e selezione.
Si ritiene ormai concordemente che l'evoluzione venga realizzata dal processo darwiniano dipendente dalla mutazione e dalla selezione naturale. L'esposizione di questa tesi, tuttavia, tende a trascurare il fatto che, mentre la mutazione avviene nel genotipo, la selezione naturale agisce solo a livello del fenotipo. Ne consegue che la teoria dell'evoluzione richiede, come sua parte essenziale, una considerazione dei processi dello sviluppo o dei processi epigenetici attraverso i quali il genotipo viene tradotto nel fenotipo.
Un organismo in sviluppo è soggetto alla selezione naturale esercitata dalle sue particolari condizioni ambientali. L'ambiente non è lo stesso per tutti gli individui di una popolazione e non rimane necessariamente lo stesso attraverso periodi di tempo di evoluzione. In realtà possiamo considerare un organismo come qualcosa che deve venire in contatto con cambiamenti ambientali imprevedibili. L'evoluzione adotta, per venire incontro a questa situazione, due tipi fondamentalmente diversi di meccanismi di sviluppo, in varie proporzioni per i differenti organismi. Uno, forse il più ovvio, è l'acquisizione da parte dell'organismo di un alto grado di capacità, tale da permettere al suo sviluppo di essere modificato dalle circostanze ambientali in modo da far aumentare la sua idoneità biologica (fitness) in quell'ambiente: questo è il meccanismo dell'aumento continuo dell'adattabilità. È probabilmente esatto affermare che tutti gli organismi mostrano una certa capacità di adattamento ai loro ambienti, sia a breve termine (fisiologica), sia a più lungo termine (di sviluppo). Comunque nella maggior parte degli organismi, e in modo particolare nella maggior parte degli organismi superiori, è considerevolmente presente un meccanismo evoluzionistico alternativo a quello descritto. Questo meccanismo alternativo consiste nel costruire creodi di sviluppo ben tamponati o canalizzati, che portano a un risultato finale relativamente invariato in relazione ad ambienti molto diversi. Il secondo meccanismo è probabilmente usato in situazioni nelle quali l'ambiente cambia in maniera marcata durante la vita dell'animale; un'adattabilità troppo elevata alle primitive condizioni ambientali indurrebbe forse l'organismo a sviluppare forme che potrebbero essere meno adatte, in un periodo successivo della sua vita, alle cambiate condizioni ambientali.
Sia questa o no la ragione principale per l'evoluzione di sistemi di sviluppo canalizzati, è comunque un dato di fatto che è molto diffuso un grado elevato di canalizzazione. Non si trovano facilmente esempi in cui la forma di un animale sia strettamente dipendente dalle primitive condizioni ambientali, sebbene questo si osservi ad esempio nelle piante e, per fare un altro esempio, in certi crostacei d'acqua dolce nei quali quasi ogni stagno o effimera massa d'acqua determina la comparsa di una corrispondente modificazione dello sviluppo. Molto più frequentemente, comunque, si ha a che fare con situazioni come quella del topo domestico, nel quale si ritrova uno stato finale di sviluppo sempre uguale, sia che abbia vissuto in ambiente tropicale, sia in un freddo magazzino di deposito. L'unico effetto che si può rilevare, derivato da queste due diverse situazioni, è un leggero aumento di lunghezza della coda nell'ambiente caldo, dove è necessario un meccanismo più efficiente per dissipare il calore del corpo.
Questo tipo di canalizzazione dello sviluppo restringe di molto gli effetti fenotipici che possono essere prodotti dalle mutazioni geniche. In particolare, molte nuove mutazioni che avvengono in singola dose in un organismo diploide non risulta che causino alcuna alterazione del fenotipo. Questa è l'espressione più generale del fenomeno della dominanza dell'allele di tipo selvaggio. Come la presente discussione dovrebbe chiarire, questa non deve essere considerata come una relazione che implichi soltanto l'allele normale e l'allele mutato di un particolare locus, ma è piuttosto il risultato di una generale canalizzazione dello sviluppo nella quale anche molti altri loci genici hanno una parte importante.
b) Assimilazione genetica ed ereditarietà dei caratteri acquisiti.
Una controversia molto lunga nel campo dell'evoluzione riguardava il problema se le modificazioni fenotipiche causate da ambienti anormali potessero venire ereditate, nel senso che tali modificazioni si sarebbero trovate nelle generazioni successive anche in assenza dello stress iniziale dovuto all'ambiente. L'ipotesi che esse potessero essere ereditate fu prospettata da J. B. Lamarck nel XVIII secolo, ed è generalmente conosciuta come ‛ereditarietà dei caratteri acquisiti'. Fino a poco tempo fa essa aveva sostenitori tra i biologi, alcuni dei quali la usavano per contrastare le teorie darwiniane dell'evoluzione. Recentemente le opinioni su questo argomento si sono sviluppate in due diverse direzioni. In primo luogo è stato rilevato che la distinzione tra caratteri ereditati e acquisiti è arbitraria poiché tutti i caratteri sono in qualche modo ereditati, in quanto dipendono dal genotipo dell'organismo, e sono anche in qualche modo acquisiti, dal momento che lo sviluppo è influenzato anche dall'ambiente circostante. Ciò tuttavia è in certa misura cavilloso, dato che quello che stiamo discutendo è se le modificazioni dello sviluppo prodotte da ambienti anormali siano o no presenti nei discendenti di individui modificati, quando non è più presente l'ambiente anormale. L'ipotesi lamarckiana suggerisce che vi sia una qualche proprietà biologica intrinseca in grado di render conto di questo fatto, senza ricorrere al meccanismo darwiniano della selezione.
In secondo luogo è stato rilevato che la combinazione dell'adattabilità e della canalizzazione nello sviluppo ci permette di spiegare tale fenomeno in termini darwiniani piuttosto che lamarckiani. L'ambiente anormale che agisce durante lo sviluppo può riuscire a modificare anche un sistema di sviluppo ben canalizzato. Se la modificazione è di tipo adattativo, e aumenta l'idoneità biologica degli individui nei confronti dell'ambiente anormale, sarà favorita dalla selezione naturale. Lo sviluppo di individui selezionati potrebbe, comunque, mostrare anche alcune proprietà di canalizzazione, cioè di invarianza verso i cambiamenti ambientali. Questa invarianza potrebbe essere sufficiente a evitare che la discendenza di individui selezionati ritorni completamente al fenotipo originale, anche se viene allontanata dall'ambiente anormale. Dopo che la selezione per una modificazione adattativa in un ambiente anormale ha agito per molte generazioni, può essere prodotta una forma la cui canalizzazione è sufficientemente forte per mantenere il nuovo fenotipo quasi inalterato quando si ristabiliscono le condizioni ambientali precedenti la comparsa dell'alterazione. Questo processo, che è stato dimostrato in numerosi esperimenti di laboratorio, è conosciuto come assimilazione genetica (Waddington, 1964). Esso produce esattamente gli stessi risultati messi in evidenza dai sostenitori dell'eredità dei caratteri acquisiti di Lamarck, ma con un meccanismo darwiniano ortodosso che agisce su sistemi di sviluppo che hanno le comuni proprietà della canalizzazione e dell'adattabilità. (V. anche embriologia, eredità biologica, evoluzione, gene e genetica).
Bibliografia
Goldschmidt, R. B., Physiological genetics, New York 1938.
Kuhn, A., Entwicklungsphysiologie, Berlin-Heidelberg-New York 19652.
Morgan, Th. H., Embryology and genetics, New York 1934.
Needham, J., Biochemistry and morphogenesis, London 1942.
Pflugfelder, O., Lehrbuch der Entwicklungsgeschichte und Entwicklungsphysiologie der Tiere, Stuttgart-Hohenheim 1970.
Spemann, H., Embryonic development and induction, New Haven 1938.
Thompson, D'Arcy, W., Growth and form, London 19423.
Waddington, C. H., Organizers and genes, London 1941.
Waddington, C.H., Principles of embryology, London 1956.
Waddington, C. H., New patterns in genetics and development, New York 1962.
Analisi del differenziamento di Antonio García-Bellido
Sommario: 1. Introduzione. 2. Archetipi e adattamento: a) anatomia comparata; b) fattori ereditari. 3. Blastemi e morfogeni: a) morfologia sperimentale; b) gradienti; c) interazioni embrionali; d) campi morfogenetici; e) campi di posizione; f) movimenti morfogenetici; g) riconoscimento cellulare. 4. Cellule e geni: a) genealogie cellulari; b) compartimenti e limitazioni nella discendenza cellulare; c) il genoma attivo; d) mosaici genetici; e) regolazione genica. 5. Molecole e informazione: a) l'approccio riduzionista; b) determinismo e vincoli; c) vincoli molecolari e genetici. 6. Considerazioni conclusive. □ Bibliografia.
1. Introduzione
Lo sviluppo è una delle più complesse manifestazioni della vita. Tale complessità si esprime nella molteplicità delle forme che compaiono sia nel corso della vita degli individui (ontogenesi) sia nelle varie specie (filogenesi). Il compito specifico della biologia dello sviluppo è quello di descrivere e spiegare come si determinino tali differenze; per la biologia, anzi, lo sviluppo ha avuto la funzione di canone interpretativo. L'osservazione che i caratteri morfologici dei genitori venivano trasmessi fedelmente alla prole ha dato ongine allo studio dell'ereditarietà, e la somiglianza tra individui di razze e specie diverse ha portato alla nozione di evoluzione. Verso la metà del secolo scorso la cellula fu identificata come l'unità costruttiva fondamentale dello sviluppo di piante e animali, mentre nella seconda metà del secolo i cromosomi furono riconosciuti come gli unici organelli cellulari in grado di replicarsi essi sono ugualmente distribuiti nelle cellule figlie e vengono trasmessi alla prole da ambedue i gameti. All'inizio del nostro secolo fu stabilita la correlazione fra trasmissione di varianti ereditarie e di segmenti cromosomici; grazie a ciò l'ereditarietà diviene il nesso causale tra sviluppo ed evoluzione.
L'isolamento e la caratterizzazione dei componenti chimici della cellula iniziarono nella prima metà del secolo scorso e terminarono con l'identificazione, avvenuta al volgere del secolo, dei principali tipi di entità molecolari tra questi solo le proteine, sotto forma di enzimi responsabili sia delle modificazioni metaboliche sia delle componenti strutturali e degli organelli cellulari, mostrano un grado di variazione sufficiente a permettere una correlazione con l'eterogeneità tessutale e i fenomeni evolutivi. Verso la metà del nostro secolo è stato possibile spiegare la specificità delle proteine sulla base della specifica sequenza lineare dei loro componenti, gli amminoacidi. Negli anni cinquanta sono state comprese le proprietà di replicazione del DNA - un polimero lineare di nucleotidi componente costante dei cromosomi - e poco dopo si è riusciti a chiarire la natura di ‛stampo' del DNA, che, tramite la sequenza dei suoi nucleotidi, determina quella degli amminoacidi che costituiscono le proteine è stato in tal modo possibile spiegare le basi molecolari della mutazione. Eredità, sviluppo ed evoluzione sono stati così ricondotti a proprietà delle molecole.
Ma in che modo tali proprietà molecolari riescono a condizionare e determinare la diversità morfogenetica? Come possono assicurare la costanza dello sviluppo di una particolare specie e nello stesso tempo la variabilità tra le specie? L'approccio riduzionista, o analitico, è riuscito a definire gli elementi della variazione. Il problema operativo che la biologia dello sviluppo deve affrontare oggi è quello di scoprire fino a che punto è possibile prevedere le interazioni tra le molecole, e quindi la loro funzione, partendo dalla conoscenza degli elementi che le compongono. In tal modo si riuscirebbe a capire il mondo morfologico in quanto sottosistema del mondo possibile definito dalle proprietà delle molecole. Altrimenti, esistono leggi o limitazioni, che regolano i livelli di organizzazione al di sopra di quello molecolare, che non possono essere previste puramente sulla base della conoscenza delle proprietà degli elementi componenti le molecole ? Qualunque risposta a questo interrogativo richiede una descrizione, classificazione e analisi causale dei processi coinvolti nello sviluppo come quelle portate avanti nei classici studi di biologia dello sviluppo. In questo articolo sarà seguito l'indirizzo storico, in modo da mostrare come i metodi usati, le idee e gli interessi prevalenti abbiano condizionato l'attuale comprensione della fenomenologia dello sviluppo.
2. Archetipi e adattamento
a) Anatomia comparata.
Il XX secolo ha ereditato dai secoli precedenti una gran quantità non solo di informazioni sui reali processi implicati nello sviluppo, ma anche di ricerche epistemologiche sui modi di classificare e spiegare la diversità morfologica. La prima metà del XIX secolo è stata dominata dalla ‛morfologia razionale', che ha tentato di trovare delle proprietà invariabili (leggi della forma) che spiegassero la stabilità interna degli aspetti morfologici specifici e la costanza di talune caratteristiche morfologiche comuni a specie diverse. Il principio di correlazione di Cuvier cercò di dare una spiegazione funzionale della morfologia attraverso l'adattamento reciproco delle varie parti a formare un tutto integrato. E. Geoffroy Saint-Hilaire spiegò le variazioni morfologiche tra le specie come il risultato di modificazioni secondarie di un sottostante piano di organizzazione (Bauplan) comune. La relazione topologica tra le parti permise di distinguere le omologie e le analogie delle variazioni morfologiche. Le diverse appendici degli organismi metamerici ci appaiono come il risultato di modificazioni secondarie di una comune organizzazione seriale. L'anatomia comparata ci aiuta a scoprire l'organizzazione essenziale o di base (gli ‛archetipi') comune al maggior numero possibile di specie, in modo da stabilire una classificazione naturale (tassonomia) delle forme viventi. In gran parte i diversi ‛tipi' definiti in tal modo corrispondono agli attuali phyla della tassonomia moderna. I reperti fossili e i dati sullo sviluppo in specie con complessi cicli vitali (metamorfosi, metagenesi, ecc.) aiutarono a chiarire i problemi di analogia' e ‛affinità' (appartenenza allo stesso gruppo di classificazione, ‛tipo'). Queste astrazioni formali portarono da una parte a proporre leggi interne che ‛organizzano' e danno stabilità alle forme, e dall'altra a proporre la derivazione di forme particolari da altre più primitive. J. B. Lamarck suggerì specificamente che queste variazioni fossero filogenetiche, causate cioè dall'adattamento ai particolari ambienti in cui le specie vivono.
Verso la metà del XIX secolo i biologi cominciarono a interessarsi al problema della generazione, a porre la questione storica dell'origine. La morfologia fu sostituita dalla morfogenesi. Analisi particolareggiate dei processi dello sviluppo portarono, in questo periodo, a scartare due nozioni dominanti della biologia precedente: una è la nozione di preformazione, che considera lo sviluppo come la semplice amplificazione di strutture preesistenti nell'uovo o nello spermatozoo. I lavori descrittivi confermarono le conclusioni raggiunte da K. F. Wolff, e cioè che lo sviluppo è accompagnato dalla comparsa di strutture e organizzazioni nuove derivanti da materiale indifferenziato (epigenesi). La scoperta della divisione cellulare e il conseguente principio generale che le cellule derivano solo da cellule preesistenti (R. L. K. Virchow) portarono a scartare l'idea della generazione spontanea delle forme viventi. L'apparente contraddizione tra queste due nozioni ha travagliato la biologia fino all'avvento della biologia molecolare.
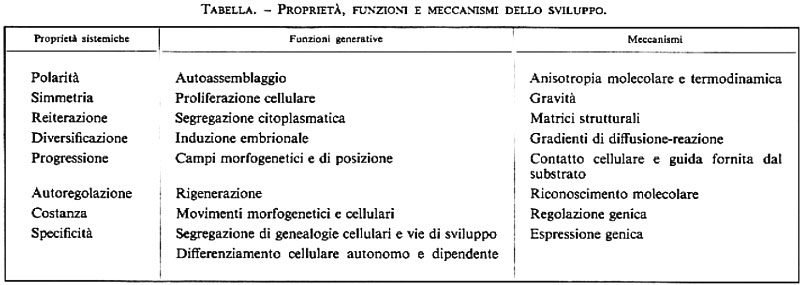
L'analisi comparata dei processi dello sviluppo permette di mettere in evidenza le proprietà fondamentali che lo regolano (v. tabella). Le categorie di organizzazione che ancora attendono di essere spiegate in termini evolutivi, genetici e molecolari sono: la polarità, presente a tutti i livelli di organizzazione, in base alla quale vi è una distinzione tra testa e coda, o stelo e radici, o, nelle uova, tra polo animale e polo vegetativo; la simmetria, che può suddividere l'embrione lungo uno o più piani, come nei Vertebrati, negli Echinodermi e nelle Fanerogame, o determinare un asse generativo a spirale, come nelle lumache; la reiterazione, cioè la ripetizione secondo uno schema spaziale di elementi identici od omologhi, quali ciglia, piume o peli, singoli organi o anche interi segmenti (metamerismo). Il metamerismo può presentarsi come reiterazioni identiche (come la segmentazione degli Anellidi) o essere modificato secondariamente (come negli Artropodi), e i segmenti possono essere in numero preciso e costante (come negli Insetti) oppure indeterminato (è il caso della strobilazione dei Cestodi).
b) Fattori ereditari
Verso la seconda metà del secolo scorso il successo dei principi causali o meccanicisti in fisica e in chimica influenzò il pensiero biologico. La teoria dell'evoluzione di Ch. Darwin tentò di dare una spiegazione causale alla nozione di archetipo: questa fu sostituita da quella di progenitore comune, che forniva anche una spiegazione del concetto di adattamento come risultato della selezione naturale. Così la diversità morfologica divenne una conseguenza della variazione di fattori ereditari, che possono essere diversi per quantità o qualità anche tra gli individui di una stessa popolazione; le combinazioni di fattori che forniscono la massima idoneità (fitness) sono quelle che divengono le più comuni nella discendenza. Pertanto, differenze geografiche ed ecologiche favoriscono nuove combinazioni di fattori, ed è così che possono originarsi nuove specie. Questo processo, andato avanti durante tempi lunghissimi e attraverso mutamenti geologici, può aver dato origine alla diversità morfologica osservabile attualmente. Le specie derivano da altre specie e possiamo stabilire una sequenza a ritroso fino a forme sempre più primitive; i reperti fossili ci possono fornire gli anelli mancanti fra le specie attualmente distinte dal punto di vista morfologico.
Anche i dati dell'embriologia forniscono un ottimo sostegno alla teoria dell'evoluzione. E. Haeckel stabilì i primi alberi genealogici (filogenie) che mettono in correlazione i gruppi animali con forme sempre più primitive sulla base di progenitori comuni; il suo modo di procedere faceva ancora uso di forme astratte, o archetipi, per identificare queste forme primitive, distinguendo i tratti palingenetici, più primitivi e più vicini alla struttura fondamentale (Bauplan) dei gruppi, da quelli cenogenetici, risultanti cioè da un adattamento secondario o sussidiario. In realtà, la morfologia comparata degli embrioni appartenenti allo stesso phylum mette in evidenza corrispondenze di forme palingenetiche con forme adulte di organismi più primitivi, tanto più evidenti quanto più è precoce lo stadio embrionale analizzato (legge di ricapitolazione di Haeckel). Questa legge, pur con le necessarie limitazioni, non soltanto aveva un valore pragmatico per l'individuazione di alberi filogenetici, ma forniva altresì la prima spiegazione razionale dello sviluppo, che sembra seguire un programma interno che si oppone ai mutamenti, sembra cioè dominato da una sorta di inerzia evolutiva.
La teoria dell'evoluzione basata sulla trasmissione di combinazioni di fattori ereditari è la prima teoria unificante che spiega il classico paradosso di costanza e variazione. Diviene di primaria importanza sapere quale sia la natura di questi fattori, come essi variino, come vengano trasmessi alla prole e come esercitino i loro effetti sullo sviluppo. La chiara distinzione stabilita da Weismann tra soma e linea germinale rese possibile l'analisi di questo problema. Secondo Weismann, lo sviluppo procede dalla originaria cellula uovo per diversificazioni di genealogie cellulari; le diverse parti dell'embrione, pertanto, sono il risultato della segregazione in tempi e localizzazioni diversi di cellule madri diverse, in maniera analoga a quanto avviene per le ramificazioni che avvengono nella speciazione nel corso dell'evoluzione. In queste segregazioni le varie parti divengono indipendenti anche per ciò che riguarda il loro sviluppo successivo; ad esempio, a uno stadio precoce dello sviluppo viene separata la linea germinale che darà origine ai gameti: essa contiene tutte le informazioni che verranno espresse in modo differenziale nel soma, pur rappresentando una linea indipendente e non essendo dal soma stesso influenzata. Questo è il primo dogma della biologia, formalmente equivalente al dogma centrale della moderna biologia molecolare che stabilisce l'irreversibilità del flusso di informazioni nel senso DNA → proteine.
Durante gli ultimi venti anni del secolo scorso fu compreso il meccanismo della mitosi, che assicura una uguale distribuzione dei cromosomi nelle cellule figlie (W. Flemming, W. Roux). Fu accuratamente descritto anche il processo di divisione riduzionale della meiosi in base al quale si originano gameti aploidi, aventi, cioè, metà dei cromosomi presenti nelle cellule mitotiche diploidi (O. Hertwig, A. Weismann, Th. H. Boveri), e il processo di fecondazione che risulta dalla fusione dei due gameti aploidi dei genitori. I cromosomi divennero così i candidati più ovvii a sostenere il ruolo di trasportatori dei fattori ereditari, prima ancora che, nel 1900, fosse riscoperta l'opera di G. Mendel.
Verso il volgere del secolo, i cromosomi divennero il punto focale degli studi sia embriologici sia tassonomici. Il fatto che il cariotipo, cioè il loro numero, la loro grandezza e la loro forma, fosse variabile tra specie diverse, ma costante all'interno di una stessa specie, fu di aiuto per la costruzione di alberi filogenetici, che in molti casi confermarono le relazioni filogenetiche stabilite sulla base dei caratteri morfologici; infatti, le differenze ontogenetiche riscontrabili all'interno di una data specie, come le differenze sessuali, e i cambiamenti morfologici che avvengono in organismi con cicli vitali complessi, erano talvolta associati con diversità del cariotipo. Ad esempio, le differenze sessuali sono spesso correlate con differenze morfologiche dei cromosomi sessuali. La partenogenesi negli animali o il suo equivalente nelle piante (apomissi) dà origine a femmine (come negli Afidi) o maschi (come nelle Api) che sono in genere molto diversi dai normali individui sessuati sia da un punto di vista morfologico che embriologico: tali processi sono determinati da una anormale segregazione dei cromosomi durante la meiosi. In modo analogo, rilevanti differenze sistemiche tra forme in metagenesi (quali idroidi e medusoidi nei Celenterati) sono associate alle fasi sessuata e asessuata; nelle piante l'eteromorfismo delle generazioni gametofitiche e sporofitiche è associato con una costituzione cromosomica rispettivamente aploide e diploide. Tuttavia, in altri casi di sviluppo discontinuo, come il polimorfismo degli organismi sociali (polipi o Insetti) o la metamorfosi dei Crostacei e degli Insetti, le differenze morfologiche non hanno potuto essere messe in relazione a differenze cromosomiche. I fattori ereditari responsabili di queste morfologie alternative tra individui di una stessa specie devono perciò risiedere in entità ancora più piccole dei cromosomi. La morfologia comparata ha raggiunto il massimo grado di riduzione atomistica, e già il Weismann aveva inquadrato il problema con sufficiente chiarezza: le strutture al di sotto del livello cromosomico sono entità chimiche.
3. Blastemi e morfogeni
a) Morfologia sperimentale
La morfologia sperimentale si è sviluppata durante l'ultimo quarto del XIX secolo. Il problema dell'origine e della genesi degli organismi che la morfologia comparata voleva risolvere è ora impostato diversamente, e lo scopo è quello di scoprire le cause reali che determinano i processi dello sviluppo. La rivista ‟Archiv für Entwicklungsmechanik der Organismen", fondata da Roux nel 1883, contiene nel suo numero iniziale un manifesto delle intenzioni. Per poter capire lo sviluppo è necessario sconvolgerlo, in modo da scoprire le basi meccaniche dei processi ontogenetici. Le tecniche dell'embriologia sperimentale consistono nella separazione di parti, cellule o territori embrionali (blastemi), e nello studio del loro sviluppo in espianti o dopo trapianto in altre parti dell'embrione. Dopo che fu chiarita la dipendenza morfogenetica delle parti, gli esperimenti mirarono a isolare i componenti chimici (morfogeni) responsabili di tali interazioni. Le spiegazioni causali vengono in un primo tempo definite in termini di forze, di ioni, di reazioni fisicochimiche che avvengono nell'ambito di gradienti o campi, e in seguito come risultato della specificità di particolari molecole.
Grazie all'interpretazione di due esperimenti classici fu possibile definire due concezioni o atteggiamenti relativi alla comprensione dello sviluppo. Ambedue gli esperimenti avevano lo scopo di chiarire l'effetto dell'ablazione di uno dei due blastomeri risultanti dalla prima divisione zigotica. Roux effettuò l'esperimento sull'uovo di rana, pungendo e quindi uccidendo un blastomero; H. A. E. Driesch invece, separò i due blastomeri di un uovo di riccio di mare. I risultati ottenuti furono assolutamente diversi nel primo caso si sviluppò metà embrione, nel secondo due embrioni di dimensioni più piccole. Dal primo esperimento Roux dedusse l'esistenza di determinanti diversi (genetici) per le diverse parti dell'embrione che vengono segregati, a guisa di mosaico, durante lo sviluppo, e ciò fornì il supporto sperimentale al concetto di differenziamento somatico e a quello di segregazione somatica contrapposto a quello di segregazione della linea germinale, formulato da Weismann. Secondo Driesch, invece, il suo esperimento doveva essere interpretato nel senso che lo sviluppo esprime una proprietà della vita, l'autoregolazione, che non è possibile ridurre in termini meccanicistici o atomistici. Esperimenti successivi dimostrarono che anche nella rana, se i blastomeri vengono separati invece che uccisi, si determina uno sviluppo regolativo. La mancanza di regolazione nel caso che un blastomero venga ucciso può essere causata dal fatto che questo, prima di entrare in citolisi, fornisce al blastomero intatto che continua a proliferare le informazioni normali.
Le interpretazioni di questi esperimenti, in termini applicabili alla morfogenesi in generale, erano tuttavia premature, in quanto esse corrispondevano a due atteggiamenti diversi ancora riscontrabili tra i biologi dello sviluppo. La concezione di Roux era meccanicistica e riduzionistica, quella di Driesch era olistica e strutturalistica, e portava a porre una maggiore enfasi, nell'ambito della morfogenesi, su quei fenomeni epigenetici che avvengono a un livello ben lontano da quello genetico. Secondo la visione di Roux e Weismann, lo sviluppo era un fenomeno decentralizzato nel quale ogni parte dell'embrione era libera di esprimere la propria informazione genetica, mentre secondo Driesch lo sviluppo era il risultato di un insieme integrato e interdipendente.
Esperimenti successivi effettuati sullo stesso tipo di organismi mostrarono che si può sviluppare un embrione intero da ognuno dei primi quattro blastomeri che risultano dai piani di divisione lungo l'asse animale-vegetativo, mentre si ottengono embrioni anormali da blastomeri ottenuti da divisioni che attraversano tale asse. Questa scoperta fu interpretata come la dimostrazione che in questo organismo il citoplasma dell'uovo è isotropo intorno all'asse, ma anisotropo lungo di esso. In altri organismi (Spiralia) la totipotenza dei blastomeri separati viene limitata già nella prima divisione, oppure (Mammiferi) viene mantenuta anche dopo molte mitosi lungo qualunque piano di divisione. In effetti, i due tipi di sviluppo, a ‛mosaico' o ‛regolativo', corrispondono a differenze che insorgono gradualmente e ambedue possono essere riscontrati in processi di sviluppo diversi nello stesso organismo.
L'esistenza di fenomeni regolativi nello sviluppo ha reso possibile la sperimentazione, ma ha reso sempre più difficile la loro spiegazione in termini meccanicistici o atomistici. Gli esperimenti di trapianto di blastemi in posizioni ectopiche nell'embrione hanno portato a operare una distinzione tra autodifferenziamento (sviluppo secondo l'origine) e differenziamento dipendente (sviluppo secondo la posizione). Trapiantando lo stesso blastema a stadi di sviluppo diversi è stato possibile effettuare una ulteriore distinzione tra potenzialità prospettica (l'inventano di ciò che un territorio può generare in seguito a manipolazioni sperimentali) e potenza reale (ciò che realmente accade in situ). I territori sono determinati in un particolare senso (sviluppo a seconda dell'origine) molto prima di esprimere un reale differenziamento; pertanto lo sviluppo fu interpretato come un processo di restrizione progressiva della potenzialità prospettica alla potenza reale. Queste nozioni segnarono il corso della moderna fisiologia dello sviluppo.
Roux e Weismann consideravano lo sviluppo come il risultato della segregazione di fattori determinanti, presenti nei cromosomi, durante la divisione cellulare, ma tale concezione venne presto scartata. Gli esempi che suffragavano il loro punto di vista quali la diminuzione di cromatina riscontrabile in Ascaris durante la divisione delle cellule somatiche, l'anormale morfogenesi associata con la generazione di blastomeri aneuploidi causata da fecondazioni multiple, le anormalità morfologiche negli ibridi interspecifici riscontrate da Boveri, o le varianti morfologiche causate da mutazioni vennero considerati eccezioni irrilevanti, dovute a perturbazioni disgregative aspecifiche, oppure furono ritenuti piccole variazioni sussidiarie che non avevano alcun effetto sull'organizzazione di base. Anche Th. H. Morgan, fondatore della genetica moderna e studioso di embriologia, dovette conformarsi a queste interpretazioni (v. Morgan, 1934). Per la maggior parte degli embriologi, lo sviluppo divenne, dall'inizio del secolo, un problema epigenetico i fattori genetici rispondono alle condizioni definite dalle segregazioni citoplasmatiche che si verificano durante la mitosi o dalle interazioni cellulari. Secondo questa concezione, il genoma era visto come una entità invariabile, fondamentalmente preposta a basilari processi cellulari e metabolici. D'altra parte, i complessi cambiamenti e movimenti citoplasmatici riscontrabili dopo la fecondazione, gli effetti della centrifugazione sullo sviluppo dell'uovo, i casi di avanzato sviluppo che si verificano in uova attivate partenogeneticamente ed enucleate, e lo sviluppo a mosaico suggerivano che i determinanti morfogenetici fossero localizzati nel citoplasma; la segregazione di tali determinanti nelle cellule era interpretata come la principale fonte di diversità morfogenetica.
b) Gradienti
Gli esperimenti su sistemi regolativi, basati sulla separazione e combinazione di parti, o su trapianti, suggerirono che i fattori morfogenetici determinanti fossero distribuiti nell'embrione lungo gradienti continui, o ‛campi' bidimensionali. Così nel riccio di mare, ad esempio, mentre gruppi di blastomeri provenienti dal polo animale danno luogo negli espianti a strutture per la maggior parte dorsali e quelli provenienti dal polo vegetativo a strutture ventrali, una combinazione di blastomeri presi a livelli diversi lungo l'asse animale-vegetativo dà origine a embrioni completi (Hörstadius, 1935). È pertanto probabile che i determinanti morfogenetici siano distribuiti lungo questo asse come due gradienti opposti aventi concentrazioni massime nei poli animale e vegetativo (v. fig. 1). Vale a dire che i determinanti morfogenetici non vengono distribuiti in maniera discontinua a seguito della divisione cellulare, ma anzi essi dirigono la morfogenesi tramite processi di interazione cellulare, mediati dal contatto cellulare o da diffusione chimica. Secondo tale concetto, la cellula perde la propria individualità e diviene o una barriera contro la diffusione o un elemento di connessione o di costruzione in un sistema di organizzazione sopracellulare.
c) Interazioni embrionali
Le interazioni cellulari possono essere il mezzo attraverso cui si genera la diversità. Osservazioni dirette avevano mostrato che, nello sviluppo degli Anfibi, la blastula originaria risultante dalla segmentazione dei blastomeri generava gli organi interni (gastrula) per invaginazione di materiale esterno verso l'interno della blastula attraverso il blastoporo. In tal modo lo strato cellulare continuo della blastula si trasforma dando luogo a tre strati concentrici - ectoderma, mesoderma ed entoderma - dai quali derivano in seguito tutti i tessuti dell'embrione. Inoltre, la collocazione del blastoporo stabiliva un asse embrionale che definiva un piano di simmetria bilaterale. Ai poli opposti di tale asse si trovano il futuro ano e la futura bocca, e lungo di esso l'ectoderma formerà un solco invaginato che in seguito si chiude per formare il tubo neurale (v. embriologia). Spemann e Mangold (v., 1924), effettuando degli esperimenti di trapianto, scoprirono che, ponendo del materiale mesodermico presuntivo ottenuto dal labbro dorsale del blastoporo sotto l'ectoderma ventrale, si determinava la comparsa ectopica di un nuovo asse embrionale. Se veniva usato materiale embrionale ottenuto da specie diverse di Anfibi con diversa pigmentazione citoplasmatica, si riuscivano a distinguere i derivati dell'ospite da quelli dell'impianto nell'asse ectopico. I derivati dell'ectoderma avevano esclusivamente origine dall'ospite, mentre cellule derivanti sia dall'ospite sia dall'impianto contribuivano a formare le strutture mesodermiche ed entodermiche. Il nuovo asse embrionale aveva una configurazione normale. Il territorio che dava questo tipo di risposta fu denominato da Spemann ‛organizzatore'; esso pareva preposto a due funzioni: una, l'‛induzione complementare', cambiava il destino presuntivo dell'ectoderma sovrastante da epidermide ventrale a epidermide dorsale e tubo neurale; l'altra, l'‛induzione assimilativa', consisteva nella auto-organizzazione dei derivati dello strato germinale interno in un tutto integrato.
Interazioni dello stesso tipo furono in seguito riscontrate in stadi successivi dello sviluppo degli Anfibi nella formazione di parti del sistema nervoso, delle vescicole otiche e ottiche, della bocca, pelle, ghiandole a secrezione interna, ecc. (v. fig. 2), e furono riscontrate anche in altri tipi di organismi, sia Vertebrati sia Invertebrati.
L'induzione embrionale è stata la prima operazione generativa che ha permesso di stabilire la sequenza dei processi di sviluppo secondo un ordine causale. I ripiegamenti multipli e i mutamenti delle posizioni relative dei blastemi dell'embrione determinano l'esposizione delle cellule a nuovi contatti, cambiandone i rapporti di contiguità, e rendono così possibili nuove interazioni induttive (v. fig. 2). L'induzione complementare è soggetta a svariate limitazioni, come quella di essere polarizzata o non reciproca: nel caso dei Vertebrati, è il mesoderma, o i suoi derivati, ad agire da induttore sull'ectoderma, mentre negli Insetti avviene il contrario. Il tessuto bersaglio pone delle limitazioni sia spaziali sia temporali (competenza); lo stesso territorio passa attraverso stadi durante i quali o non risponde all'induttore, o risponde dando origine a strutture diverse via via che il tempo passa e lo sviluppo progredisce. Anche territori appartenenti allo stesso tessuto competente danno risposte variabili a seconda della posizione che occupano. Inoltre, le strutture che si differenziano dal tessuto competente sono specie-specifiche. Quando sono state effettuate delle combinazioni di derma ed epidermide in tessuti embrionali di Rettili, Uccelli e Mammiferi, i derivati dell'epidermide hanno dato origine a scaglie, piume e peli dello stesso tipo di quelli della classe del donatore dell'epidermide (v. Sengel e Dhouailly, 1977). Esiste, tuttavia, anche un certo grado di specificità che dipende dall'origine dei territori induttori: effettuando analoghe combinazioni di derma ed epidermide nell'ambito di una stessa classe, lo schema di distribuzione di scaglie, piume e peli corrisponde alla classe e alla regione corporea del donatore da cui proviene il derma. Allo stesso modo, regioni diverse lungo l'asse antero-posteriore del mesoderma induttore risultano tanto più efficienti nell'indurre regioni neurali quanto più vicine si trovano alle posizioni normali corrispondenti.
Poco dopo che fu scoperto il fenomeno dell'induzione complementare, gli studiosi cercarono di identificare la natura chimico-fisica dei fattori inducenti. Nel 1944 Holtfreter (v., 1951) trovò, con esperimenti in vitro, che blastemi denaturati per mezzo del calore, provenienti dall'organizzatore naturale o da molti altri tessuti, o perfino agenti citolitici applicati al tessuto competente, riuscivano a promuovere una risposta induttiva anche se incompleta e disorganizzata. Usando dei sistemi modello, i ricercatori hanno tentato di restringere il numero delle sostanze implicate nel processo di induzione frapponendo dei filtri Millipore tra l'induttore e i blastemi competenti (v. Grobstein, 1961); nel fenomeno dell'induzione sembra che siano implicati proteine diffusibili, alcuni RNA e perfino i contatti tra membrane. L'attuale comprensione dei complessi fenomeni di induzione embrionale si deve al lavoro di Saxen e Toivonen (v. Saxen e Karkinen-Jaaskelainen, 1981). La specificità espressa dal tessuto rispondente, tuttavia, avvalora l'ipotesi che il ruolo dell'induttore sia quello di evocare o avviare certe risposte predeterminate del tessuto competente (v. Waddington, 1941).
d) Campi morfogenetici
La seconda funzione associata all'azione dell'organizzatore, e cioè l'‛induzione assimilativa', determina l'auto-organizzazione, o ‛regionalizzazione', di territori di cellule aventi diversa origine in un sistema integrato. In questo caso l'induttore opera come un centro organizzativo, in cui cellule provenienti sia dall'ospite sia dal donatore interagiscono in modo tale da determinare l'organizzazione finale. Durante lo sviluppo, interazioni reciproche di questo tipo sono comunissime. Le proprietà auto-organizzative di questi sistemi corrispondono a ciò che viene definito ‛campo morfogenetico'. In questi campi, le potenzialità morfogenetiche presuntive sono distribuite lungo un gradiente radiale che ha un valore massimo al centro (v. fig. 3A); nei Vertebrati, per esempio, il territorio embrionale che può dare origine agli arti è molto più esteso di quello che normalmente si differenzia realmente in tal senso. Trapianti ottenuti da una regione qualsiasi di questo territorio presuntivo sono in grado di dar luogo alla formazione degli arti, ma tale potenzialità viene progressivamente ristretta a zone sempre più vicine a quelle della localizzazione presuntiva, via via che lo sviluppo progredisce. La formalizzazione di queste proprietà ha portato a formulare la teoria dei campi, per mezzo della quale si cerca di spiegare molti processi ontogenetici in termini di diffusione chimico-fisica e di meccanismi di reazione (Turing; v. Weiss, 1939). Le cellule che si trovano in campi morfogenetici hanno valori scalari di concentrazione di morfogeni e sono dotate di polarità rispetto al centro. L'ablazione di parti di un campo morfogenetico determina un riaggiustamento della parte rimanente in modo da rigenerare il campo; combinando insieme due campi si ottiene la loro fusione in un unico campo nuovo. I campi morfogenetici possono in un secondo tempo dividersi in sottocampi e originare deformazioni anisotrope per interazione con altri campi o possono acquisire nuove polarità. Esperimenti di trapianto hanno mostrato che gli abbozzi di arti hanno in un primo tempo una polarità cefalo-caudale, che diviene in seguito dorso-ventrale (R. G. Harrison, 1925). La suddivisione dell'abbozzo degli arti lungo un asse prossimo-distale è forse il risultato di una suddivisione in campi morfogenetici concentrici.
Benché i campi morfogenetici siano stati caratterizzati con chiarezza in molti sistemi ontogenetici, sappiamo poco dei meccanismi cellulari che vi sono coinvolti. Il sistema meglio studiato è quello del plasmodio di aggregazione dell'ameba Dictyostelium (v. Loomis, 1975). È noto che l'aggregazione delle amebe migratorie in corpi fruttiferi è mediata da gradienti di concentrazione di un prodotto cellulare, l'‛acrasina', che è stato caratterizzato come AMP ciclico in alcune specie, e come acetilcolina in altre. Le amebe si concentrano lungo un gradiente da esse stesse generato, la cui concentrazione è modulata dalle cellule stesse che esercitano un controllo sull'attività di certi enzimi capaci di degradare il prodotto che funge da segnale (fosfodiesterasi o acetilcolinesterasi; v. Gerisch e Hess, 1974). Che cosa determini il cessare dell'aggregazione e l'inizio della formazione del cono e del corpo fruttifero e la sua successiva suddivisione in regioni, e tuttavia ancora ignoto (v. embriologia).
e) Campi di posizione
Nei classici campi morfogenetici le irregolarità della configurazione finale sono dovute all'interazione delle cellule attraverso un processo sia continuo sia sequenziale; tuttavia, in alcuni sistemi ontogenetici appaiono in modo discontinuo e simultaneo delle irregolantà nel campo originariamente omogeneo. Esempi di ciò sono rappresentati dallo schema di distribuzione delle ossa della mano, dalla distribuzione delle piume o dei peli nella pelle o dalla disposizione dei diversi tipi di neuroni negli strati della corteccia cerebrale. I processi che danno origine a questi schemi morfologici hanno reso necessario supporre l'esistenza di un'altra funzione generativa. Queste irregolarità si possono generare in modo autonomo relativamente al tessuto nel quale esse appaiono, oppure essere determinate dal tessuto sottostante, come abbiamo visto nel caso dello schema di distribuzione di piume e peli negli esperimenti di combinazione di derma ed epidermide. I sistemi più semplici e meglio studiati sono quelli nei quali lo schema viene originato in modo autonomo in un unico monostrato di cellule, come ad esempio nella formazione dei modelli di distribuzione cuticolare negli Insetti (v. fig. 3B). In questi modelli, i diversi elementi cuticolari (setole, sensilli e tricomi) si formano come unici derivati di cellule epidermiche, che mantengono una posizione relativa costante e caratteristica per le varie parti del corpo. Esperimenti di trapianto di frammenti di epidermide, che mantengono o mutano la propria polarità nei confronti del tessuto ospite, nello stesso o in diversi livelli di un particolare segmento, suggeriscono l'esistenza di un campo di posizione sottostante. Questi esperimenti mostrano che il frammento trapiantato mantiene la propria polarità originaria in relazione all'asse corporeo, come pure un ‛valore di posizione' in relazione a certi confini, come i bordi visibili dei segmenti. Un trapianto che non rispetti queste coordinate spaziali determina un riaggiustamento tra il tessuto dell'impianto e l'ospite, o una rotazione dell'impianto (o il suo riassorbimento, nel caso che sia innestato con una differenza di angolazione di 180°) e un accrescimento intercalare in modo da ripristinare i corretti valori di posizione e di polarità. I trapianti effettuati tra diversi segmenti del corpo o delle appendici suggeriscono che i loro campi di posizione siano omologhi; si ha una reazione ospite-donatore solo nel caso che i valori di posizione e la polarità non corrispondano (Stumpf, 1956; Locke, 1959; Bohn, 1974). L'ablazione di parte del campo di posizione può portare alla perdita dei corrispondenti elementi della struttura, se viene impedita la proliferazione cellulare. Tuttavia, nella maggior parte dei casi di regolazione della struttura, il blastema rimanente può determinare il rimodellamento e la rispecificazione della struttura che viene così rigenerata (morfallassi), oppure la proliferazione cellulare intercalare (epimorfosi) può ripristinare le distanze originali tra i rimanenti elementi della struttura.
Nonostante che la maggior parte della fenomenologia a favore del concetto di campo di posizione derivi da esperimenti di rigenerazione, è probabile che tali campi siano realmente operanti nello sviluppo normale. La più esplicita formulazione del ruolo che l'informazione sulla posizione gioca nella morfogenesi e nella formazione delle strutture si deve a Wolpert (v., 1969). Secondo la teoria dell'informazione fornita dalla posizione, ogni cellula viene in primo luogo definita dalla sua posizione in un campo, come nel caso del campo morfogenetico; questa posizione è a sua volta definita da valori scalari in un sistema di coordinate spaziali, cartesiane o polari (quest'ultimo è il caso delle appendici); tali valori potrebbero corrispondere a gradienti di concentrazione di morfogeni specifici, a differenze di fase tra onde oscillanti o a più generiche funzioni chimico-fisiche di diffusione-reazione. La cellula registra in modo autonomo i segnali provenienti dai campi di posizione e vi risponde a seconda della propria costituzione genetica e della propria storia ontogenetica. Il differenziamento localizzato di una cellula e i confini dei campi di posizione sono definiti da valori di soglia imposti dal meccanismo genetico che risponde; pertanto possono originarsi delle discontinuità dello schema in conseguenza di risposte genetiche a un campo di posizione invariante e monotonico (v. fig. 3B). In un primo tempo i campi di posizione sono piccoli e contengono poche cellule, al massimo un centinaio; tuttavia, la successiva proliferazione può portare a una suddivisione dei campi originari, dovuta all'interpretazione genetica e alla generazione di nuove discontinuità di soglia tra campi confinanti. In teoria, gli stessi meccanismi di segnalazione possono venire usati più volte, ed essere anzi semplici e universali. La teoria dell'informazione posizionale è una teoria unificatrice che combina tutte le proprietà dei campi morfogenetici con la specificità della risposta determinata dalla costituzione genetica. Purtroppo, però, i tentativi di caratterizzare le basi chimico-fisiche dei segnali legati alla posizione e i sistemi genetici specifici che rispondono a questi segnali non hanno finora avuto successo. I veri meccanismi cellulari operanti nella specificazione della posizione rimarranno ignoti finché non si sarà analizzato in profondità qualche sistema ontogenetico. Come riescono le cellule a rispondere simultaneamente, e tuttavia in modo indipendente, a molti parametri coordinati ? Le cellule agiscono come relè o sono assolutamente permeabili alla trasmissione delle informazioni provenienti dalla posizione ? Le discontinuità di gradiente sono generate dalla specificazione genetica o la precedono ? Come si determina la forma dei campi di posizione? In qual modo i campi riescono a suddividersi in un numero costante di sottocampi, come nel caso del numero dei segmenti nell'embrione degli Insetti? Come si ottiene la regolazione nei Ciliati monocellulari (Frankel, 1975)?
f) Movimenti morfogenetici.
Durante lo sviluppo può essere ottenuta una diversità spaziale modificando la posizione relativa dei territori; questi cambiamenti sono il risultato di movimenti morfogenetici che interessano blastemi interi o singole cellule. I meccanismi comunemente impiegati per generare l'organizzazione tridimensionale sono i ripiegamenti, che possono avere la forma di evaginazioni o invaginazioni ad esempio, il cordomesoderma risulta dall'invaginazione all'interno del blastocele dei territori dorsali della blastula. L'applicazione di coloranti vitali al territorio del cordomesoderma presuntivo ha reso possibile seguirne il processo di invaginazione (W. Vogt, 1925). Tuttavia, non è ancora stato possibile determinare con certezza la causa di questo processo. La proliferazione cellulare con la conseguente espansione del territorio può essere la causa meccanica dell'invaginazione, ma potrebbero essere operanti anche dei cambiamenti locali della forma delle cellule dovuti a mutamenti del citoscheletro. Tramite espianti del labbro dorsale del blastoporo, è stato possibile dimostrare che i cambiamenti di forma delle cellule che avvengono durante il processo di invaginazione e il successivo differenziamento sono una caratteristica autonoma delle cellule del cordomesoderma presuntivo (v. Holtfreter, 1943). È pertanto possibile che specifici movimenti morfogenetici cellulari siano una conseguenza di una precedente specificazione cellulare (determinazione) che definisce il loro comportamento successivo. Anche esperimenti in vitro effettuati per studiare i processi associati alla formazione di tubi e cavità (quali il tubo neurale o le cavità celomatiche nel mesoderma somitico e splancnico) hanno mostrato movimenti e ripiegamenti autonomi che riflettono una precedente specificazione cellulare.
Nei Vertebrati, un tipo molto comune di movimento morfogenetico è quello della migrazione di cellule in piccoli gruppi o singolarmente. Per esempio, le cellule che daranno origine a melanoblasti e al sistema nervoso simpatico hanno origine nella cresta neurale e migrano attraverso i somiti e la splancnopleura disponendosi secondo schemi spaziali specifici. E ancora incerto se il differenziamento finale delle cellule della cresta neurale sia già determinato quando questa viene abbandonata, o se sia una conseguenza delle vie che esse seguono e della posizione che raggiungono durante la migrazione. Esperimenti di trapianto di blastemi provenienti da donatori aventi pigmentazione diversa indicano che il differenziamento dei melanociti dipende dalla loro origine, ma la distribuzione dipende dalla loro posizione (Twitty, 1945; v. Rawles, 1948). Il substrato sul quale le cellule migrano può fornire delle indicazioni che guidano le cellule verso la loro destinazione finale; tali indicazioni possono essere di natura chimica e agire per diffusione da una certa distanza (chemiotassi) o di natura fisica (struttura e natura chimico-fisica del substrato) e agire per contatto locale. Lo studio della migrazione delle cellule endoteliali, che formeranno la rete di capillari vascolari, e della migrazione delle fibre nervose ha fatto supporre che nella formazione delle strutture siano operanti ambedue i meccanismi di guida, chimico e per contatto (v. Hamburger e Levi-Montalcini, 1950; v. Sperry, 1963).
Un caso speciale di migrazione cellulare è quello grazie al quale si origina la disposizione tridimensionale dei neuroni nel cervelletto e nella corteccia. La disposizione multistratificata risulta dalla delaminazione delle cellule secondo un piano perpendicolare al monostrato epiteliale onginano (v. fig. 5D). Studi condotti su topi allofenici (che si originano per aggregazione di blastomeri provenienti da ceppi geneticamente diversi di topi e differiscono nella morfologia dei neuroni) indicano che la posizione assunta nella disposizione finale dipende in gran parte dagli schemi specifici di migrazione dei diversi tipi cellulari; durante la migrazione, le interazioni tra neuroblasti e glia determinano lo schema di connessione neuronale e il differenziamento finale, ma la specificita cellulare è probabilmente definita nel momento in cui le cellule si trovano nell'epitelio bidimensionale originario (v. Caviness e altri, 1981).
Lo studio dei processi morfogenetici associati ai movimenti del blastema e alla migrazione cellulare suggerisce che l'organizzazione tridimensionale sia una conseguenza della specificazione cellulare che avviene quando le cellule si trovano in monostrati bidimensionali; l'espressione di caratteristiche proprietà cellulari di comportamento si estrinseca nei movimenti che rispecchiano quella specificazione.
g) Riconoscimento cellulare
Lo studio dettagliato dei processi morfogenetici ha portato in primo piano il ruolo svolto dalle specifiche proprietà delle cellule nella determinazione della loro organizzazione spaziale. Ci si è domandati fino a che punto le proprietà dei blastemi riflettano le proprietà delle singole cellule. All'inizio del secolo, nel 1907, E. B. Wilson aveva mostrato che cellule dissociate da spugne adulte erano in grado di riassociarsi ricostituendo delle spugne funzionali; mescolando cellule di due specie diverse, esse si separavano le une dalle altre ricostituendo i due tipi originari. Solo molto più tardi Holtfreter esegui lo stesso tipo di esperimento di riaggregazione usando tessuti embrionali di Anfibio, e trovò (1939) che frammenti di mesoderma e di ectoderma provenienti dalla gastrula, mescolati e coltivati in vitro, erano in grado di separarsi gli uni dagli altri e di organizzarsi secondo la configurazione relativa che si ritrova normalmente nell'embrione. Gli stessi esperimenti, effettuati mescolando cellule dissociate da entoderma, mesoderma ed ectoderma provenienti da donatori con pigmentazione diversa, rivelarono l'esistenza di specifiche proprietà del tessuto (‛affinità') che determinavano la stessa organizzazione spaziale e lo stesso differenziamento cellulare che si riscontra in embrioni in stadi avanzati di sviluppo (v. Townes e Holtfreter, 1955; v. fig. 4A). Analoghe proprietà di auto-organizzazione furono in seguito riscontrate in molti esperimenti di dissociazione e di aggregazione cellulare di particolari organi embrionali. La natura chimico-fisica di queste attività cellulari specifiche è ancora oggetto di discussione. Le affinità potrebbero rispecchiare proprietà differenziali di adesione-coesione (v. Steinberg, 1963) o l'esistenza di marcatori molecolari specifici, presenti sulla superficie cellulare, che determinerebbero le proprietà di riconoscimento e di associazione delle cellule stesse (v. Moscona, 1962).
Esperimenti di dissociazione e mescolamento cellulare eseguiti usando dischi immaginali di Drosophila indicano chiaramente l'esistenza di proprietà di riconoscimento estremamente specifiche da parte delle cellule (v. fig.4B). I dischi immaginali sono monostrati di cellule dell'epidermide, derivate dall'ectoderma embrionale, che durante la metamorfosi si differenzieranno dando origine alle diverse parti del corpo e alle configurazioni cuticolari dell'adulto. L'uso di marcatori genetici consente di identificare l'origine delle singole cellule nei nuovi aggregati che si formano. Mescolando cellule embrionali di dischi immaginali diversi, o anche frammenti dello stesso tipo di disco, le cellule selezionano e ricostruiscono in monostrato l'organizzazione del disco o della regione da cui provengono. Invece, cellule provenienti da un disco dello stesso tipo e della stessa regione ricostruiscono il profilo come un mosaico, cioè mischiandosi a cellule aventi marcatori genetici diversi. Inoltre, mescolanze di cellule provenienti da dischi immaginali di individui normali e di mutanti omeotici, che mostrano trasformazioni segmentali (v. cap. 4, È d), mettono in evidenza le caratteristiche di riconoscimento del differenziamento finale e non quelle definite dall'origine segmentale del disco (v. García-Bellido, 1972). Questi risultati indicano che il riconoscimento cellulare è una delle caratteristiche che vengono definite durante il differenziamento cellulare, e anche che le differenze genetiche (forse espresse in configurazioni molecolari specifiche), insieme ai valori differenziali di posizione e al diverso grado di adesione cellulare, hanno un ruolo rilevante nei meccanismi di riconoscimento cellulare.
4. Cellule e geni
a) Genealogie cellulari
Anche se fin dalla metà del XIX secolo è stato riconosciuto che la cellula è l'unità costruttiva dello sviluppo, la scoperta della sua specificità morfogenetica è molto più recente. Ci si è resi conto, infatti, che molti processi morfogenetici sono dovuti al comportamento di singole cellule, e che questo comportamento è a sua volta determinato dall'espressione autonoma del genoma cellulare attivo. In realtà, i primi esperimenti dell'embriologia furono effettuati durante la segmentazione, nel corso della quale le singole cellule possono essere facilmente identificate e manipolate. Tuttavia, la grande complessità strutturale dell'uovo e il ruolo primario svolto dal genoma materno nel determinare i primi processi dello sviluppo non permettevano di comprendere lo sviluppo stesso in termini di cellule e geni. La nozione di genealogia o discendenza cellulare deriva dagli studi effettuati in embrioni con sviluppo a mosaico. Durante l'osservazione diretta e in esperimenti di ablazione fu possibile mettere in luce in questo tipo di embrioni il ruolo svolto dai piani di divisione nel suddividere il citoplasma dell'uovo e nel definire le diverse vie di sviluppo (Conklin, 1905; v. Wilson, 19253). I diversi blastomeri acquisiscono presto un significato prospettico discreto, che è espresso per eredità cellulare in tipi e localizzazioni cellulari diversi. Divenne chiaro in seguito che diverse modalità di proliferazione cellulare negli animali e nelle piante hanno una funzione nella morfogenesi e nel differenziamento la divisione asimmetrica, in base alla quale il citoplasma non viene diviso egualmente sia dal punto di vista della quantità sia da quello della polarità; le divisioni cambiali, che da una cellula madre generano una serie di cellule figlie identiche; la proliferazione intercalare, che fa si che si riempiano dei territori aventi una determinata estensione o che causa delle invaginazioni localizzate, ecc. Lo studio delle discendenze cellulari, d'altra parte, mostra che lo sviluppo si attua per mezzo della segregazione di famiglie di tessuti aventi ciascuno un repertorio particolare di tipi di differenziamento, come quelli che derivano dai principali strati germinativi, cioè ectoderma, mesoderma ed entoderma. Il ruolo della cellula diviene ovvio quando il processo di differenziamento cellulare può essere definito in termini non solo morfologici ma anche biochimici. Secondo tale visione riduzionistica, lo sviluppo è il risultato di una segregazione discontinua di tipi cellulari definiti non solo in base al loro destino presuntivo, ma anche al loro differenziamento finale. Fino a che punto la posizione e l'organizzazione generale sono conseguenza della specificità genetica delle singole cellule?
L'analisi dettagliata delle discendenze cellulari richiede l'uso di marcatori cellulari, che per l'osservazione diretta possono essere morfologici o spaziali, o possono derivare dalla marcatura di cellule progenitrici con coloranti vitali, enzimi o marcatori fluorescenti, oppure ancora possono consistere in variazioni genetiche. Queste ultime, per la loro indelebilità, hanno particolarmente contribuito alla comprensione di tali processi. I mosaici che si originano nei topi in seguito a mescolamento di blastomeri (Mintz, 1965; v. Mintz, 1971), o in Drosophila mediante manipolazioni cr0mosomiche durante lo sviluppo (v. Stern, 1954), hanno fornito i risultati più importanti di questo tipo di studi. I mosaici genetici, in Drosophila, possono essere il risultato di una perdita, spontanea o indotta, di uno dei cromosomi sessuali nelle prime divisioni di segmentazione, dando luogo a cellule XX (femmina) o X0 (maschio). Se il cromosoma mancante porta alleli normali, e l'altro cromosoma X, invece, porta dei mutanti che fungono da marcatori cellulari, questi ultimi marcheranno tutte le cellule X0 della progenie. La distribuzione dei territori mutanti (X0) rispetto a quella dei territori normali (XX) può rivelare la posizione relativa degli abbozzi quando l'embrione è in uno stadio precoce, il numero di cellule che daranno luogo all'abbozzo e può mettere in luce anche l'esistenza di movimenti dei blastemi e migrazioni cellulari durante lo sviluppo. I mosaici genetici, in Drosophila, possono venire originati anche durante il periodo della proliferazione tramite ricombinazioni mitotiche (v. fig. 5A), consistenti nell'interscambio di segmenti cromosomici durante l'anafase del ciclo mitotico. Questo interscambio, che avviene nelle cellule eterozigoti rispetto ai mutanti che fungono da marcatori cellulari, può dare origine a una cellula figlia del tipo selvaggio e a una cellula mutante omozigote. La progenie di quest'ultima, cioè un ‛clone' cellulare, esprimerà il fenotipo mutante nel territorio a cui darà origine. La ricombinazione mitotica può venire indotta da agenti esterni in qualunque periodo dello sviluppo. Dallo studio dei cloni che ne risultano possiamo dedurre alcuni parametri quantitativi della proliferazione cellulare (numero di cellule nell'abbozzo, tipo della successiva proliferazione cellulare, orientamenti mitotici preferenziali, velocità di divisione, ecc.), in qualunque parte e momento di qualunque sistema ontogenetico. La forma dei cloni, la loro distribuzione spaziale e gli elementi della configurazione (pattern) di cui sono portatori può fornirci delle indicazioni sul modo in cui si forma tale pattern.
b) Compartimenti e limitazioni nella discendenza cellulare
In Drosophila, i cloni che hanno avuto inizio durante la segmentazione hanno nell'adulto l'aspetto di territori compatti che includono una delle due strutture adiacenti presenti nella cuticola adulta; ciò indica che i nuclei di segmentazione sono totipotenti e che la successiva proliferazione non è ancora determinata per ciò che riguarda il pattern finale. Tuttavia, i cloni che hanno avuto inizio durante lo stadio del blastoderma includono strutture che non si estendono oltre un particolare segmento; nell'ambito di ogni segmento la proliferazione è ancora indeterminata, perché i cloni possono includere due strutture adiacenti qualsiasi, interne a un segmento, ma non possono attraversare i confini del segmento stesso. I cloni che hanno avuto inizio a uno stadio successivo non possono estendersi che a un semisegmento, e cioè a una qualunque delle due regioni, anteriore o posteriore, che derivano dal segmento. Nell'ala, i cloni anteriori possono includere qualunque parte del settore anteriore dell'ala stessa, ma non oltrepassano mai la linea virtuale che, estendendosi attraverso tutti i derivati cuticolari del segmento alare (mesotoracico), suddivide l'ala nei settori anteriore e posteriore. Linee omologhe suddividono altri segmenti in due regioni, una anteriore e una posteriore, chiamate ‛compartimenti'. I cloni che hanno inizio ancora più tardi mostrano limitazioni maggiori e sono confinati alle parti dorsali o a quelle ventrali del segmento. Nel disco immaginale dell'ala (disco mesotoracico dorsale) i cloni che si originano più tardi sono limitati a uno dei compartimenti, dorsale o ventrale, e ai compartimenti prossimale (notum) e distale (lamina alare) secondo linee di demarcazione costanti. In tal modo, durante la proliferazione cellulare, l'originario abbozzo alare si suddivide secondo una partizione binaria in due, quattro e otto compartimenti (v. fig. 7). I confini dei compartimenti, che sono definiti dai limiti comuni dei cloni nei diversi individui, sono fissi e passano attraverso elementi caratteristici costanti del pattern che sarebbe altrimenti continuo. È probabile che il riconoscimento cellulare operi per tutto il corso dello sviluppo a mantenere separate le cellule dei compartimenti vicini. All'interno dei compartimenti i cloni sono largamente sovrapposti, il che indica che la proliferazione degli elementi particolari del pattern è ancora indeterminata. Così lo sviluppo dei segmenti e dei dischi immaginali di Drosophila avviene per passi successivi di determinazione, che causano la segregazione topografica di una popolazione cellulare originariamente omogenea e danno origine a due sottopopolazioni omogenee (policlonali; v. García-Bellido e altri, 1976). A questo punto esse acquisiscono un diverso significato prospettico che si esprime nel pattern finale seguendo un repertorio costante di patterns specifici. Le segregazioni omologhe hanno luogo in segmenti diversi e in compartimenti definiti in precedenza, in modo tale che la diversità finale può essere spiegata formalmente tramite una combinazione di decisioni omologhe che influenzano discendenze cellulari separate. Esempi di segregazione clonale omologa per la definizione di segmenti e compartimenti sono stati riscontrati anche in altri insetti (Lawrence, 1981; v. Crick e Lawrence, 1975).
La segregazione dei compartimenti durante lo sviluppo avviene in gruppi limitati di cellule (20-50); in questo periodo non esistono differenze istologiche tra cellule di compartimenti diversi né si riscontra alcuna caratteristica morfologica lungo i confini dei compartimenti. Tuttavia in esperimenti di mescolamento cellulare (v. cap. 3, È g) è stato possibile mettere in evidenza che cellule appartenenti a compartimenti diversi hanno diverse affinità cellulari, il che fa presumere che vi sia un differenziamento molecolare prima che sia visibile il differenziamento strutturale finale. La crescita indeterminata dei doni all'interno dei compartimenti fa pensare alle proprietà regolative dei campi morfogenetici, mentre i confini dei compartimenti potrebbero coincidere coi confini di discontinuità che sono necessari come punti di riferimento nel caso dei campi di posizione (v. cap. 3, È e).
Durante lo sviluppo dei dischi immaginali ha luogo anche un altro tipo di segregazione clonale, che influenza le singole cellule. I doni si restringono a elementi cuticolari specifici contenenti o setole o tricomi (v. fig. 6B) molte divisioni cellulari prima del differenziamento finale che avviene alla metamorfosi. I doni delle setole di regola contengono un unico organo delle setole (e hanno origine da due divisioni cellulari differenziali), mentre i cloni dei tricomi (derivati di singole cellule epidermiche) contengono parecchi elementi. Nel periodo in cui avvengono tali limitazioni clonali non si riscontra alcuna differenza istologica tra i tipi cellulari presuntivi, anche se in esperimenti di aggregazione cellulare le cellule madri delle setole si comportano come se fossero già determinate.
Segregazioni clonali dello stesso tipo sono state recentemente descritte in altri organismi. Durante stadi precoci di sviluppo del topo si riscontra una segregazione policlonale e binaria (v. Gardner, 1978), tramite la quale la morula appena formata si suddivide prima in trofoblasto e massa cellulare interna, e in seguito ognuna di queste dà origine al trofoblasto polare e murale e a ectoderma ed entoderma primitivi. Questi ultimi, dopo ulteriori suddivisioni, daranno origine all'embrione. Segregazioni clonali omologhe si verificano durante la produzione dei diversi tipi di cellule ematiche nei Mammiferi; in questo caso le comuni cellule precursori del mesoderma splancnico danno origine alle linee di differenziazione granulocitica, eritrocitica e linfoide (v. Metcalf e Moore, 1971). Analoghe segregazioni clonali avvengono durante il differenziamento dei fibroblasti in mioblasti e condroblasti (v. Holtzer, 1970). In altri invertebrati con sviluppo a mosaico, le segregazioni monoclonali sono più comuni di quelle policlonali (v. fig. 6A). Nella sanguisuga (Hirudinidae), ripetute segregazioni clonali, secondo un sistema quasi cambiale, separano i precursori (teloblasti) dei vari tipi di neuroni dai derivati epidermici e mesodermici in segmenti successivi (v. Weisblat e altri, 1980). Nel laboratorio di Brenner è stata studiata in modo approfondito la discendenza cellulare invariante della larva del nematode Cenorhabditis, in cui eventi di segregazione omologa in discendenze diverse generano elementi simili in parti diverse dell'organismo. La discendenza è sottoposta a restrizioni tali che l'ablazione di una sola cellula causa l'assenza di tutta la discendenza prospettica (Sulson, 1981). Analogamente, lo studio delle chimere nelle piante vascolari ha mostrato che la regolarità nella segregazione di epidermide, parenchima fondamentale e derivati vascolari deriva sempre da strati cellulari diversi del meristema, presenti nel cono di crescita dello stelo, in seguito a divisioni periclinali e anticlinali (v. Dermen, 1953).
c) Il genoma attivo
L'esistenza delle discendenze cellulari e la regolarità e discontinuità con cui compaiono i diversi tipi cellulari durante lo sviluppo suggeriscono che nel genoma vi sia un certo repertorio di risposte geniche prestabilite. Nonostante la scoperta della natura particolata dell'ereditarietà, avvenuta all'inizio di questo secolo, il ruolo del genoma nello sviluppo è rimasto oscuro fino a poco tempo fa. Il ruolo morfogenetico dominante del nucleo rispetto a quello del citoplasma è stato messo in luce dagli esperimenti di incroci interspecifici, di ibridi merogonici e di trapianti nucleari, effettuati all'inizio del secolo. I classici esperimenti di trapianto nucleare tra specie diverse dell'alga monocellulare Acetabularia (Hämmerling, 1932; v. Hämmerling, 1953), hanno stabilito con chiarezza il ruolo del nucleo nella morfogenesi. Gli esperimenti di fecondazione multipla effettuati da Boveri sul riccio di mare, risultanti in anormalità dovute alla presenza di blastomeri aneuploidi, suggerirono che funzioni specifiche dello sviluppo fossero localizzate in cromosomi specifici. L'effetto di mutazioni particolari su specifici aspetti dello sviluppo suggeriva l'esistenza di fattori genetici specifici che determinavano specifiche funzioni. Il principale interesse dei genetisti dello sviluppo fu pertanto quello di identificare queste funzioni per riuscire a capire la natura dei fattori ereditari. Si riuscì a dimostrare che alcune mutazioni che avevano effetto su determinati processi metabolici erano associate a difetti enzimatici (Garrod), ma la spiegazione in termini di funzionamento difettoso delle anormalità morfologiche causate dalle mutazioni risultò più ardua.
Negli anni trenta e quaranta una grande mole di lavoro fu dedicata al tentativo di collegare direttamente le anormalità morfologiche a una disfunzione genetica. Esperimenti su topi (v. Grüneberg, 19522) e su Drosophila (v. Hadorn, 1955) permisero di individuare delle genealogie di effetti morfologici e fisiologici causati da fattori letali. Nei casi meglio studiati, tali genealogie misero in luce alterazioni metaboliche riconducibili a enzimi anormali o a insufficienze ormonali, come anche a cambiamenti strutturali di certi tipi cellulari. Tuttavia, geni mutanti diversi potevano essere responsabili di effetti secondari simili, e alleli diversi dello stesso gene potevano determinare effetti diversi. La comprensione, in termini molecolari, di due esempi classici risultò paradigmatica delle difficoltà insite nell'approccio genetico al problema dello sviluppo. L'anemia a cellule falciformi, una sindrome estremamente specifica da un punto di vista sia anatomico che fisiologico, fu ricondotta a una molecola globinica anormale (v. Pauling e altri, 1949). Fu inoltre dimostrato che le anormalità comportamentali e anatomiche specifiche del gatto siamese erano una conseguenza del difettoso funzionamento dell'enzima tirosinasi, collegabile al metabolismo dei pigmenti (v. Guillery e altri, 1974). Naturalmente, gli effetti di queste mutazioni non erano una conseguenza del cattivo funzionamento di geni specificamente responsabili del processo dello sviluppo che risultava alterato, ma piuttosto della perturbazione di interazioni altamente integrate; il loro fenotipo specifico rifletteva semplicemente l'esistenza di tali interazioni. Le mutazioni potevano aiutare a mettere in luce queste interazioni, anche se non è ancora sicuro che esistano realmente dei geni propriamente morfogenetici. Lo sviluppo, infatti, potrebbe essere spiegato come una serie di reazioni metaboliche che determinano diversi equilibri, ognuno dei quali è caratteristico di un grado particolare di differenziamento morfologico o biochimico (v. Goldschmidt, 1938; Waddington, 1957). La natura di queste reazioni a catena fu chiarita in termini molecolari dalla scoperta della regolazione enzimatica a retroazione e della regolazione genica nei Batteri.
d) Mosaici genetici
Il quadro di questa situazione è stato in parte modificato dalla possibilità di studiare mutazioni clonali in mosaici genetici. L'esistenza di mutazioni che fungono da marcatori e che si esprimono nelle singole cellule ci permette di allargare il problema dell'autonomia cellulare ad altri tipi di mutazioni (v. Stern, 1954; v. fig. 6B). Accoppiando nello stesso braccio cromosomico mutazioni morfogenetiche con mutazioni che fungono da marcatori cellulari, è possibile studiare nei cloni l'effetto dei territori mutanti sui territori normali circostanti e viceversa. In tal modo, anche mutazioni organismiche letali (che coprono il 90% circa dei 5.000 geni totali) possono essere studiate in Drosophila direttamente al livello cellulare. Lo studio delle mutazioni che influenzano il differenziamento degli elementi cuticolari ha messo in evidenza un comportamento cellulare autonomo. Analogamente, la maggior parte delle mutazioni che influenzano gli enzimi implicati nelle vie metaboliche, del tipo di quelle che determinano i diversi pigmenti dell'occhio, sono autonome rispetto alle cellule. Lo studio dei letali nei cloni ha mostrato che circa il 90% di essi sono vitali nell'epidermide addominale. In altri tessuti la proliferazione cellulare dipende dai geni in modo più rigido. Solo una piccola frazione (10% circa) dei letali vitali nei cloni ha effetti morfogenetici. Il loro fenotipo corrisponde a cambiamenti delle proprietà di adesione delle cellule, alla velocità di proliferazione, agli orientamenti mitotici, ecc., e il loro studio potrà mettere in luce le proprietà di comportamento cellulare che hanno importanza nella morfogenesi. Nella morfogenesi, tuttavia, i processi cellulari autonomi sembrano influenzati da alleli mutanti solo in una piccola frazione del genoma (v. García-Bellido e Ripoll, 1978).
Lo studio in cloni di mutazioni a effetto sistemico ci ha permesso di identificare i geni implicati nella specificazione delle genealogie cellulari. Le mutazioni sistemiche modificano lo schema di distribuzione degli elementi all'interno del sistema, oppure determinano delle trasformazioni da un sistema all'altro senza influenzare il normale differenziamento cellulare. Appartengono a questo secondo tipo le mutazioni che trasformano il fenotipo del sesso genetico in quello del sesso opposto (mutazioni di differenziamento sessuale), o sostituiscono la disposizione e la morfologia dei derivati di un segmento con quelli di un altro segmento (mutazioni omeotiche).
Mutanti omeotici in Drosophila sono noti da lungo tempo. Alleli vitali di un determinato locus possono variare sia nell'entità della trasformazione (espressività) sia per ciò che riguarda le regioni del segmento implicato (specificità); inoltre, è possibile riprodurre trasformazioni simili a quelle causate dalla mutazione impiegando agenti esterni durante lo sviluppo di individui ‛selvaggi' (fenocopie). Questa fenomenologia ha portato a interpretare i fenotipi mutanti in termini di perturbazioni secondarie del normale processo di accrescimento (Waddington, 1957). Tuttavia, la possibilità di studiare gli alleli più estremi (mutazioni silenti), anche se letali, in cloni da ricombinazione mitotica ha permesso di attribuire numerose funzioni specifiche per lo sviluppo ai corrispondenti geni selvaggi. Gli effetti prodotti dagli alleli di un locus, anche dai più estremi, sono limitati a un particolare segmento o compartimento; ad esempio, nel complesso genico bithorax, singoli loci mutanti trasformano segmenti addominali e toracici specifici nel segmento anteriore immediatamente adiacente, e la delezione totale del complesso determina la trasformazione di tutti questi segmenti nel segmento mesotoracico (v. Lewis, 1978). Nei doni di ricombinazione mitotica tali mutanti mostrano un comportamento cellulare autonomo; possiamo da ciò dedurre che la proprietà sistemica del segmento si esprime in ogni singola cellula.
I mosaici derivati da cloni mutanti, che vanno incontro a trasformazione, e le cellule normali circostanti mostrano una sostituzione topologica puntuale dei patterns corrispondenti, il che fa pensare che entrambi i sistemi condividano campi di posizione omologhi. Cloni precoci (grandi) e tardivi (piccoli) mostrano la trasformazione omeotica; ciò indica che la funzione dell'allele selvaggio è necessaria nelle singole cellule per tutto il periodo della proliferazione. Tale funzione consiste nella specificazione di una particolare via segmentale diversa rispetto a una alternativa via ‛archetipica' che è quella del segmento mesotoracico - alla quale le cellule revertono in caso di mutazione. In effetti, mutazioni del complesso che portano a un eccesso di funzione normale causano trasformazioni reciproche, cioè i segmenti anteriori si trasformano in segmenti posteriori. Pertanto, le mutazioni modificano non solo la configurazione finale dell'adulto, ma anche quella dei segmenti dell'embrione, il numero delle cellule originali dell'abbozzo, la sua dinamica di proliferazione e le affinità delle cellule; in pratica, modificano tutti i parametri dello sviluppo del segmento implicato, dallo stadio di blastoderma in poi. Il differenziamento di una particolare via segmentale si verifica quindi allo stato blastodermico, che precede di molte divisioni cellulari il differenziamento finale e visibile del segmento specifico.
In Drosophila sono noti sistemi genetici analoghi a quello del complesso bithorax ad esempio, mutazioni nel complesso antennapedia modificano la specificazione segmentale dei segmenti cefalici e protoracici in mesotorace, e quelle del locus engrailed trasformano la specificazione dei compartimenti posteriori nei corrispondenti compartimenti antenori dello stesso segmento. Combinazioni di due mutanti, quali engrailed e uno qualunque dei complessi bithorax o antennapedia, determinano una doppia trasformazione, che riguarda la specificazione sia del segmento che del compartimento. L'analisi clonale di questi mutanti ha messo in evidenza lo stesso comportamento autonomo delle cellule, ed esperimenti di mescolamento cellulare hanno mostrato dei cambiamenti corrispondenti dell'affinità cellulare.
La specificità del locus della trasformazione omeotica, la sua restrizione ai futuri segmenti o compartimenti, e l'effetto della mutazione a livello cellulare indicano che i corrispondenti geni selvaggi determinano la specificazione di tutte le cellule di un compartimento al momento della segregazione della loro discendenza. Il fatto che nella condizione mutante la via dello sviluppo si sposti verso una via alternativa precisa suggerisce che nella segregazione un particolare gene omeotico sia attivo nei confronti di una discendenza e inattivo nei confronti di quella alternativa. Poichè l'effetto della combinazione di due mutanti è additivo, si può presumere che le segregazioni progressive siano specificate da un gruppo combinatorio di attività che i diversi geni omeotici esplicano o meno, cioè un ‛indirizzo codificato' che definisce le particolari vie di sviluppo (v. fig. 7). Come tali geni riescano a determinare questa specificazione è ancora ignoto, ma è molto probabile che essi agiscano selezionando altri geni presenti nel genoma che inviano i loro segnali genetici sotto forma di proprietà di comportamento della cellula, le quali determinano la configurazione del pattern finale e la morfologia dei diversi segmenti e compartimenti (García-Bellido, 1975). Mutanti simili, probabilmente omologhi ai mutanti omeotici di Drosophila, sono noti anche in altri insetti. Pure in Cenorhabditis si stanno scoprendo delle mutazioni che influenzano corrispondenti dicotomie in famiglie di discendenze. I limiti imposti dall'analisi genetica potrebbero aver impedito la scoperta di analoghi tipi di mutazioni anche in altri organismi.
e) Regolazione genica
La scelta tra le alternative che si presentano al momento della segregazione delle discendenze viene effettuata sulla base di coordinate anatomiche invarianti; l'analisi clonale ha mostrato che nello sviluppo normale la specificazione ‛anteriore' avviene nelle cellule che si trovano nella parte anteriore del segmento. È possibile che per effettuare queste scelte le cellule ricevano dei segnali posizionali o che rispondano a interazioni cellulari polarizzate. Le analisi dei mutanti omeotici, effettuate sia dal punto di vista genetico che da quello dello sviluppo, e quella delle loro fenocopie suggeriscono la presenza di interazioni regolative tra i loro alleli selvaggi. Nell'attivazione dei geni del complesso bithorax sembra siano implicati anche i prodotti di altri geni. Lo studio di combinazioni di mutanti e di diverse dosi dei loro alleli selvaggi, oltre che l'analisi clonale, ha portato a ritenere che i prodotti di due loei particolari definiscano in modo specifico lo stato di attivazione o inattivazione dei geni bithorax. Uno di questi loci sembra che produca, in modo analogo ai geni regolatori dei Batteri, un repressore che riconosce i geni bithorax impedendo che essi si esprimano; l'altro pare implicato nella produzione o distribuzione di un induttore (un metabolita nel caso della regolazione batterica) che riconosce e inattiva il repressore. Concentrazioni variabili dei prodotti dei due geni lungo l'asse antero-posteriore allo stadio di blastoderma determinano la distribuzione del repressore libero secondo un gradiente che a sua volta determina un'attivazione differenziale dei geni del complesso bithorax lungo l'asse dell'embrione e una specificazione segmentale differenziale. Questi stessi due geni regolatori sembra siano coinvolti anche nella regolazione dei geni del complesso antennapedia. Spetterà all'analisi molecolare confermare, modificare o escludere questo modello basato su dati genetici; tuttavia, nella sua forma attuale, esso ci aiuta a raffigurarci i processi implicati nella produzione dei segnali di posizione e nella loro registrazione nel genoma in termini genetici, e a chiarire i compiti da essi svolti nel portare a termine lo sviluppo. Questo modello stabilisce in modo esplicito l'esistenza di una gerarchia di interazioni genetiche che hanno a che fare con la specificazione delle vie di sviluppo: geni ‛attivatori' determinano l'attività di geni ‛selettori' che dipendono da segnali esterni; questi ultimi geni avrebbero effetto su gruppi di altri geni, i ‛realizzatori', che convertirebbero i segnali genetici dei ‛selettori' in specifici comportamenti cellulari (v. fig. 7). In questo modello, le vie di sviluppo vengono definite dalla scelta sequenziale tra alternative genetiche nelle discendenze cellulari. Geni specifici, aventi un ruolo analogo a quello degli enzimi nelle vie metaboliche, sarebbero comparsi durante l'evoluzione per facilitare o favorire la scelta di specifiche alternative di sviluppo.
Gerarchie genetiche dello stesso tipo sembrano essere presenti anche in altre vie di sviluppo. In Drosophila, nel corso del differenziamento delle caratteristiche sessuali dimorfiche, il principale meccanismo che determina il sesso, cioè il rapporto cromosoma sessuale/autosoma, è legato allo stato di attivazione-inattivazione di un gene legato al cromosoma sessuale. La sua attività determina quella di altri loci autosomici che si trovano più in basso in questa gerarchia; mutazioni in tali loci causano una completa reversione sessuale a livello cellulare. L'attività di uno di questi loci (double sex) sembra determinare in ultima analisi l'attività di sottogruppi di geni, maschili o femminili, che regolano il differenziamento sessuale. Mutazioni a livello di questo locus causano la comparsa di caratteristiche sessuali intermedie nelle singole cellule. In maniera analoga a ciò che accade nel caso dei geni omeotici, mutamenti nella costituzione genetica dei loci che determinano il sesso mostrano, nei mosaici genetici che si generano durante lo sviluppo, trasformazioni cellulari autonome (v. Baker e Ridge, 1980).
Anche la specificazione del pattern cuticolare sembra essere sotto il controllo di geni specifici. Lo schema di disposizione delle setole è il risultato di interazioni cellulari a breve distanza tra precursori cellulari delle setole, che hanno luogo abbastanza precocemente durante la proliferazione degli abbozzi epidermici. La densità e la distribuzione degli elementi nella configurazione finale dipendono dal grado di attività dei geni del complesso achaete-scute, determinato a sua volta dall'attività di geni regolatori che influenzano le interazioni cellulari. Come in altri casi discussi in precedenza, tuttavia, il differenziamento finale delle setole è un evento cellulare autonomo, che rispecchia in modo diretto la costituzione genetica delle singole cellule rispetto a questi geni (v. García-Bellido, 1981).
È probabile che l'azione regolativa dei geni sia operante anche in altri sistemi di sviluppo. Ibridi di cellule somatiche, ottenuti per fusione di tipi cellulari diversi o aventi diversa origine genetica, hanno mostrato che l'attività genetica, misurata in base alla comparsa o all'eliminazione di particolari prodotti genici, può essere modulata da prodotti diffusibili di origine nucleare (v. Harris, 1970; v. Ephrussi, 1972). Cambiamenti di determinazione improvvisi (‛transdeterminazione') in colture di dischi immaginali di Drosophila, che determinano la comparsa di strutture alari in abbozzi del disco originariamente dell'arto, rivelano l'esistenza di sottogruppi discreti di geni alternativi. È presumibile che tali mutamenti transdeterminativi siano una conseguenza di una mutata attività di geni regolatori (omeotici) (v. Hadorn, 1967). Analogamente, la comparsa di tipi o tessuti cellulari discreti nei corpi embrioidi riscontrata nei teratocarcinomi del topo suggerisce l'esistenza di programmi di sviluppo discontinui (Stevens; v. Jacob, 1977). Questi fenomeni, anche in mancanza di riscontri a livello molecolare o genetico, indicano pure che, similmente a ciò che accade nei Batteri, la regolazione genica e il differenziamento cellulare non dipendono da cambiamenti irreversibili nel genoma. Il fatto che nuclei di tessuti differenziati siano capaci di determinare un normale sviluppo di uova enucleate di rana (v. Gurdon, 1974), o che cellule isolate da tessuti fruttiferi possano produrre una nuova pianta, (v. Steward, 1958) rappresentano casi estremi di reversibilità del genoma attivo. Sembra pertanto che il genoma degli organismi in via di sviluppo sia un sistema aperto avente stati stazionari diversi che sono determinati da una regolazione genica reversibile. Questa interpretazione contrasta col concetto che considera lo sviluppo come una cascata irreversibile di eventi molecolari che risultano da cambiamenti strutturali o da conversioni metaboliche operate dalle reazioni enzimatiche.
La possibilità di visualizzare l'attività di trascrizione nei rigonfiamenti (puffs) dei cromosomi politenici (v. genetica: Citogenetica) ha permesso di identificare i processi della regolazione genica a un livello vicino a quello molecolare. Gli schemi di attività genica che è possibile visualizzare in questo modo sono specifici sia del tessuto che dello stadio di sviluppo (v. Beermann, 1952) e possono essere modulati da cambiamenti delle condizioni sperimentali in colture cellulari in vitro; ad esempio, l'applicazione di shock termici e l'uso di ormoni possono dar luogo a puffs aventi configurazioni specifiche. Inoltre, specifici ormoni determinano particolari schemi di attività genica in particolari sequenze, il che indica l'esistenza di meccanismi specifici di regolazione tra i gruppi di geni. Studi biochimici hanno permesso di caratterizzare le condizioni fisiologiche e cellulari di questa regolazione almeno in queste condizioni l'attività genica sembra reversibile (v. Ashburner, 1975).
In termini genetici il concetto secondo cui lo sviluppo è il risultato dello svolgimento di un programma può essere ricondotto a quello secondo il quale esso è la conseguenza dell'esistenza di interazioni tra gruppi di geni specifici. Nella selezione di tali gruppi sembra che le interazioni di posizione, a breve e lunga distanza (contatto cellulare, morfogeni e ormoni diffusibili), abbiano la funzione di innescare dall'esterno una risposta definita in modo specifico dal repertorio genetico della singola cellula. Il genoma appare pertanto come un archivio o registro delle funzioni genetiche. Se vie di sviluppo del tipo di quelle riscontrate in Drosophila sono una caratteristica generale dello sviluppo embrionale, tali istruzioni dovrebbero essere codificate nei geni che selezionano in modo specifico e controllano l'attività di sottogruppi di funzioni geniche.
5. Molecole e informazione
a) L'approccio riduzionista.
Verso la metà del nostro secolo, con l'approccio riduzionista si è giunti al livello molecolare nella spiegazione dei fenomeni biologici, si è cercato, cioè, di definire le proprietà e gli eventi dello sviluppo in termini di meccanismi (v. meccanicismo e vitalismo). La complessità dell'organizzazione può essere ridotta a combinazioni di elementi più semplici; ad esempio, organelli cellulari come membrane, ribosomi e cromosomi possono essere visti come strutture composte da elementi molecolari discreti, gli enzimi, e le proteine strutturali come specifiche sequenze di amminoacidi. I metaboliti (come gli zuccheri), gli acidi nucleici e perfino gli amminoacidi vengono prodotti da attività enzimatiche che rimuovono dalle molecole certi atomi o ne aggiungono altri presenti nel mezzo circostante. Anche il problema dello sviluppo può essere riproposto in termini molecolari come si origina la specificità della struttura molecolare e come tale struttura determina la sua funzione?
La scoperta da parte di J. D. Watson e F. H. C. Crick nel 1953 della struttura e delle proprietà del DNA facilitò la risposta al primo quesito. Enzimi e proteine sono in genere sintetizzati come una sequenza di amminoacidi su uno stampo di sequenze nucleotidiche omologhe presenti nel DNA. Negli anni sessanta la biologia molecolare ha descritto in dettaglio questo processo di trasformazione isomorfica (v. genetica). La sequenza lineare dei nucleotidi del DNA viene copiata in una sequenza di nucleotidi dell'RNA (trascrizione), che viene letta come una specifica sequenza di amminoacidi (traduzione). Alterazioni nella sequenza del DNA (mutazioni) si convertono in errori di lettura, cioè in proteine errate. La configurazione tridimensionale delle proteine è dovuta al ripiegamento della catena polipeptidica, determinato dalle cariche ioniche dei gruppi liberi degli amminoacidi o dalla presenza di legami covalenti tra di essi. Pertanto, la sequenza lineare specifica condiziona la configurazione tridimensionale. La struttura terziaria, tuttavia, ha un certo grado di libertà può essere modificata in modo reversibile dall'ambiente ionico, dalle interazioni della proteina coi suoi substrati nel caso degli enzimi o dal grado di ossidazione del suo gruppo prostetico, come nel caso dell'emoglobina. Cambiamenti di configurazione di questo tipo avvengono quando, per l'interazione di proteine diverse fra di loro o con altre molecole, si formano dei complessi, quali enzimi polimerici, ribosomi o membrane.
Per ciò che riguarda l'informazione, la struttura terziaria di una proteina e la sua associazione quaternaria dipendono strettamente dalla sequenza dei suoi amminoacidi, e pertanto tali strutture possono essere considerate esse stesse come ‛codificate' nel DNA. Il processo che genera queste strutture è detto ‛autoassemblaggio', e sembra essere implicato nel controllo di livelli di organizzazione che vanno oltre quello degli organelli cellulari. Un processo di questo tipo è operante, ad esempio, nella morfogenesi di alcuni virus (v. fig. 8). Gli studi effettuati dai gruppi di Kellenberger e di R. S. Edgar sul batteriofago T4 di E. coli hanno mostrato che la morfologia del capside (nelle sue parti di testa, collo e coda) dipende fondamentalmente dall'associazione spontanea di 40 proteine diverse codificate da geni diversi; ad esempio, la testa risulta dall'associazione di un numero finito di proteine identiche aventi simmetria e proprietà di combinazione tali che possono formare solo un icosaedro regolare. Analogamente, la reiterata associazione di proteine simili genera il collo, e quella di altre proteine genera la struttura della coda. Piccole alterazioni post-traduzionali di polipeptidi nascenti possono essere necessarie per rendere possibile la combinazione di parti diverse del capside (v. Edgar e Wood, 1966). Processi simili di autoassemblaggio sono alla base della morfogenesi di altri virus. Ci sono delle analogie tra questi processi e ciò che avviene in sistemi di sviluppo più complessi, come per esempio le simmetrie intrinseche, la reiterazione e l'associazione combinatoria di elementi.
L'analisi genetica della morfogenesi virale ha mostrato che l'assemblaggio è estremamente sensibile alle mutazioni. Questi sistemi dipendono tanto strettamente dalle complesse interazioni steriche che la maggior parte delle sostituzioni amminoacidiche, in una qualsiasi delle proteine costruttive, porta al crollo della struttura. La variazione e quindi l'evoluzione di questi sistemi è morfologicamente, e perciò geneticamente, sottoposta a vincoli. Solo un limitato numero di combinazioni è compatibile con la stabilità strutturale.
b) Determinismo e vincoli.
Fino a che punto l'autoassemblaggio regola l'organizzazione di strutture complesse come cromosomi, ribosomi, mitocondri e membrane ? Fino a che punto la differenza morfologica tra tipi cellulari, come ad esempio quella tra fibroblasti e cellule epiteliali, dipende dai componenti del loro citoscheletro? Secondo l'approccio riduzionista, la struttura e la funzione finali sono determinate dalle proprietà dell'interazione degli elementi componenti e possono essere spiegate in questi termini. Le variazioni dell'evoluzione deriverebbero quindi dallo sfruttamento dei gradi di libertà di tali interazioni, pur entro i limiti stabiliti dai vincoli strutturali. Questo punto di vista suggerisce che lo sviluppo dovrebbe essere studiato cominciando dal fondo, cioè dalla descrizione dettagliata degli elementi implicati nelle interazioni. Già a questo livello di organizzazione compare il primo problema epistemologico dell'approccio riduzionista. La funzione di una molecola è definita dalle sue interazioni, che sono a loro volta conseguenza della struttura degli elementi implicati; ma la conoscenza di tutte le strutture esistenti e dei loro possibili cambiamenti di configurazione non è sufficiente per prevedere quelle interazioni. Secondo un punto di vista alternativo, la plasticità dell'interazione, che aumenta col crescere del livello organizzativo, è tanto grande che i vincoli vengono imposti dalla funzione in maniera sempre più indipendente dalla natura degli elementi componenti. La diversità tra questi indirizzi interpretativi a livello molecolare è parimenti riscontrata nella interpretazione dei livelli più elevati di organizzazione nel corso dello sviluppo.
L'analisi molecolare ci permette di definire la sequenza lineare degli elementi e la struttura sia delle proteine che del DNA. Dallo studio comparato di queste molecole in organismi diversi, è possibile chiarire quali proprietà non possono variare e quali invece possono, e fino a che punto, durante l'evoluzione. Alcuni complessi proteici hanno composizione e configurazione molto costanti lungo tutta la scala filogenetica; è questo il caso degli istoni associati al DNA in tutti i cromosomi degli eucarioti e delle subunità maggiori e minori dei ribosomi, presenti sia nei procarioti che negli eucarioti. La costante configurazione quaternaria di questi complessi è probabilmente dovuta ai vincoli imposti dall'associazione dei diversi istoni fra di loro e col DNA, e, nel caso dei ribosomi, dall'associazione delle proteme che li compongono fra di loro e con rRNA, 5sRNA, RNA diversi e mRNA. È perciò sorprendente che, nonostante che la configurazione quaternaria, e in gran parte anche quella terziaria, delle proteine si sia mantenuta costante, le sequenze amminoacidiche nella stessa proteina possano essere diverse nei vari organismi. Anche lo studio delle variazioni delle sequenze amminoacidiche negli enzimi ha mostrato l'esistenza di regioni variabili e di altre costanti, associate, queste ultime, al dominio proteico implicato nella reazione enzimatica. Nelle regioni che possono variare, la variazione può consistere in piccole sostituzioni amminoacidiche, o in cambiamenti radicali nella sequenza e nella lunghezza della catena peptidica. Nei diversi tipi di tRNA, che adattano la sequenza codificante dell'mRNA ad amminoacidi particolari per la formazione dei peptidi, la regione dell'anticodone è ovviamente costante e specifica per gli amminoacidi, mentre il resto della molecola può variare grandemente, sia nella sequenza primaria sia nella configurazione secondaria, a seconda dei diversi tipi di tRNA. I tRNA devono riconoscere gli enzimi attivatori specifici che associano i diversi amminoacidi ai tRNA. Ma la configurazione terziaria (determinata tramite cristallografia a raggi X) è stranamente molto simile in tutti i tRNA, probabilmente a causa della loro interazione con la comune superficie ribosomiale sulla quale si forma il legame peptidico. Nell'ambito delle strutture del tRNA, sembra che la selezione abbia operato al livello funzionale delle interazioni molecolari, ma abbia permesso una considerevole variazione dell'organizzazione della sequenza al livello strutturale.
La diversità esistente tra i tRNA rappresenta un altro modello degli eventi legati all'evoluzione che non possono essere predetti sulla base di considerazioni strutturali. La corrispondenza riscontrata tra l'anticodone del tRNA e il particolare amminoacido che lega il tRNA non può essere spiegata in termini di struttura. Si può presumere che questa corrispondenza si sia verificata casualmente e sia stata poi fissata e conservata tra molte altre statisticamente possibili per selezione naturale. Spiegazioni di questo tipo possono rendere conto della prevalente presenza in tutti gli organismi di amminoacidi L e di zuccheri D, invece che delle loro forme alternative, aventi asimmetria ottica opposta. Nonostante che sia le forme D sia quelle L si trasformino spontaneamente le une nelle altre, e si trovino ambedue in natura, ciascuna forma richiede un enzima diverso. Gli organismi ne possiedono uno solo; la scelta di questo da parte degli organismi primitivi può essere stata un evento casuale perpetuatosi per selezione naturale. Il ruolo svolto dalla mutazione, altro evento casuale che produce diversità molecolare, è ormai stabilito con sicurezza.
Il fatto che un rigido determinismo non sia riuscito a spiegare la complessità in termini causali ha determinato uno spostamento dell'interesse verso la ricerca della natura dei vincoli che determinano l'organizzazione discontinua. É tuttavia scoraggiante il fatto che via via che aumentano i dati comparativi sulle molecole, le leggi che regolano l'organizzazione sopramolecolare diventino sempre più difficili da capire. Nel DNA esistono sequenze uniche, probabilmente regioni codificanti le proteine, e sequenze ripetitive; queste ultime compaiono in famiglie che possono variare, anche in specie correlate, per numero, lunghezza della parte che si ripete, sequenza e distribuzione nel genoma. La loro presenza nella maggior parte degli eucarioti fa pensare che svolgano una precisa funzione (v. Britten, 1982); in alcuni casi esse codificano specifici tipi di RNA di funzione nota (rRNA, 5sRNA, ecc.). Le altre sequenze non vengono neppure trascritte, e potrebbero rappresentare regioni strutturali per funzioni cromosomiche meccaniche, oppure artefatti dell'evoluzione, che non sono direttamente esposte alla selezione naturale a livello di specie, ma vengono utilizzate come strumenti di trasposizione genica, cioè per la riorganizzazione del genoma. Nella trascrizione anche sequenze uniche potrebbero non contenere informazioni, dato che appaiono interrotte, a breve distanza, da codoni di interruzione o privi di significato (non sense) in tutte le possibili chiavi di lettura. Le regioni del DNA che codificano per le proteine possono essere presenti in forma singola o ripetersi molte volte con una disposizione in serie o dispersa nel genoma. L'analisi delle mutazioni, tuttavia, ha mostrato che solo una di queste copie viene usata nella trascrizione; anche in questo caso, l'organizzazione del genoma può essere spiegata solo a posteriori, come fonte di variazione genetica. Famiglie di proteine aventi funzione analoga (miosine, actine, ecc.) sembrano originate dalla duplicazione genica che, attraverso graduali modifiche e diversificazioni della stessa sequenza nucleotidica, sarebbe responsabile della comparsa di nuove funzioni.
In termini molecolari l'evoluzione è basata sulla mutazione, e la variazione molecolare viene mantenuta per tutto il tempo in cui essa resta compatibile con l'interazione molecolare. Tale interazione è la funzione che viene percepita dalla selezione naturale. La variazione rende possibile un aumento del numero e dei tipi di interazioni, che si attuano attraverso modifiche delle copie di geni e attraverso combinazioni diverse dei prodotti genici che stabiliscono relazioni sempre più complesse. Alla complessità è collegato un aumento del grado di libertà di queste interazioni, e la selezione deve operare a livelli di organizzazione sempre più alti, ben lontani da quello della struttura molecolare. In questo processo sembra che l'evoluzione abbia favorito meccanismi di regolazione e di adattabilità piuttosto che meccanismi che determinano la rigidità e la precisione della morfogenesi.
c) Vincoli molecolari e genetici
L'analisi molecolare ha messo in luce la straordinaria diversità funzionale che può essere ottenuta per mezzo di associazioni, combinazioni reiterate e interazioni tra le molecole. Pertanto, il problema che i biologi dello sviluppo devono risolvere è quello di spiegare perché questa continua fonte di variazioni si risolve in tipi discontinui di morfologie, ai vari livelli di organizzazione. Le soluzioni morfologiche discontinue potrebbero essere il risultato di errori di campionatura effettuati dall'evoluzione, oppure potrebbero riflettere l'esistenza di vincoli funzionali. La scoperta di questi vincoli è uno dei principali obiettivi che si pone la moderna biologia dello sviluppo. A livello molecolare, due sono le limitazioni che si ritrovano in tutti i processi morfogenetici e nell'evoluzione: il riconoscimento molecolare e l'inerzia genetica.
Il riconoscimento molecolare sta alla base dei processi multipli di interazione, come la percezione degli odori durante l'accoppiamento degli organismi bisessuali, il riconoscimento cellulare nel corso dei processi morfogenetici, le reazioni enzimatiche e l'autoassemblaggio dei complessi molecolari, ed è operante anche nella regolazione genica. L'associazione di proteine specifiche, che sono prodotte dai geni regolatori, con sequenze di DNA o con configurazioni strutturali specifiche in vicinanza di un gene strutturale, determina l'attività di trascrizione di tale gene. Divengono cosi possibili delle combinazioni spaziali e temporali di sottogruppi di funzioni geniche. Nei Batteri sono presenti enzimi costitutivi ed enzimi che possono essere indotti da metaboliti presenti nel mezzo esterno; questi ultimi vengono trascritti solo quando la concentrazione intracellulare di questi metaboliti raggiunge determinati valori. Il prodotto proteico di un gene regolatore riconosce i metaboliti in modo specifico e si lega a essi; ciò determina un cambiamento di configurazione in seguito al quale esso diventa incapace di riconoscere la sequenza regolativa cis del gene strutturale per l'enzima, che in tal modo resta esposto all'azione della RNA-polimerasi e può essere trascritto. Se il metabolita manca, la stessa proteina regolativa riconosce la regione regolativa cis, vi si associa e impedisce in tal modo la trascrizione. Il riconoscimento molecolare operato dalle proteine regolative, pertanto, dipende da cambiamenti di configurazione (effetto allosterico) modulati dai metaboliti in uno dei domini della molecola proteica, e dal DNA nell'altro (v. Jacob e Monod, 1961). Cambiando le costanti di affinità proteina-DNA-metabolita, le stesse interazioni determinano un controllo negativo per repressione (la trascrizione ha luogo in assenza della proteina regolativa) o un controllo positivo per attivazione (la trascrizione ha luogo in seguito all'associazione della proteina regolativa, eventualmente legata all'induttore, con il DNA del gene strutturale).
I tipi di controllo, negativo o positivo, sono specifici dei diversi sistemi regolativi. I geni regolatori possono controllare uno o, più spesso, diversi geni strutturali (batterie) collocati uno vicino all'altro o dispersi nel genoma. Dati genetici e molecolari indicano che il numero dei geni presenti nei Batteri varia tra duemila e quattromila; solo una piccola parte di questi sono regolatori, mentre un maggior numero si trova sotto controllo regolativo e la maggioranza è di tipo costitutivo.
Come abbiamo visto nel cap. 4, È e, vi sono molte ragioni per pensare che la regolazione genica sia operante anche nel differenziamento cellulare degli organismi pluricellulari. È probabile che questa regolazione si attui attraverso queste stesse proprietà di riconoscimento molecolare. Sequenze canoniche, in vicinanza della parte terminale 5′ della regione strutturale codificante riconosciuta dagli enzimi (come la RNA-polimerasi II), sono state riscontrate in molti geni di organismi diversi. Non sappiamo ancora se la regione regolativa cis dei geni strutturali consenta il riconoscimento molecolare da parte di proteine regolative per mezzo di specifiche sequenze di DNA, come si verifica nei Batteri, oppure per mezzo di una configurazione della cromatina che solo indirettamente dipende da particolari sequenze del DNA. I processi molecolari implicati nella regolazione genica sono stati definiti in modo migliore nei sistemi ormone-dipendenti nei Vertebrati e negli Insetti. Nel caso della regolazione della trascrizione genica operata dall'ecdisone negli Insetti, il tessuto bersaglio dell'ormone produce un recettore proteico che lega l'ormone presente alla membrana cellulare e lo trasporta nel nucleo. Un'associazione del complesso ormone-recettore proteico coi loci che rispondono all'ormone è stata osservata nei cromosomi politenici di Drosophila (v. Gronemeyer e Pongs, 1980). Tuttavia, il fatto che sia necessaria la presenza dello specifico recettore dell'ormone nel tessuto bersaglio fa dubitare che l'azione ormonale possa essere considerata un esempio di regolazione genica; gli ormoni potrebbero essere semplicemente dei modulatori della trascrizione di geni scelti in precedenza attraverso il reale meccanismo di differenziamento.
Un fenomeno scoperto di recente e anch'esso basato sul riconoscimento molecolare potrebbe essere implicato nella regolazione genica a livello post-trascrizionale. Tale fenomeno, chiamato splicing (‛rottura e saldatura') dell'RNA, è stato riscontrato in animali, piante e nei loro virus (v. Crick, 1979). L'RNA primario che risulta dalla trascrizione viene sottoposto a un procedimento in base al quale certe sequenze intercalari (introni) vengono rimosse, mentre le sequenze rimanenti (esoni) vengono legate a formare un unico RNA maturo, per opera di enzimi specifici. Il numero e la lunghezza degli introni dello stesso gene possono essere diversi nei diversi organismi. Gli introni sembrano essere una caratteristica genica tanto più diffusa quanto più si sale lungo la scala filogenetica. Il ruolo dello splicing dei geni nell'evoluzione e nello sviluppo al presente ci sfugge, ma ci sono dati che sembrano indicare che esso possa essere utilizzato nella regolazione genica. Nei geni di alcuni virus lo splicing differenziale dei trascritti dello stesso gene puo originare proteine diverse ed è possibile che tale fenomeno sia operante anche nel differenziamento cellulare. È probabile che questo sia ciò che accade in alcuni complessi genici di Drosophila, come i complessi bithorax e achaete-scute (v. cap. 4; v. Hogness e altri, 1981). Lo splicing dei geni, a differenza dello splicing dell'RNA, potrebbe essere responsabile della diversità delle molecole correlate durante lo sviluppo. Nel caso della produzione dei diversi tipi di immunoglobuline, la traslocazione del DNA ha luogo tra sequenze specifiche in modo tale che combinazioni specifiche di regioni lunghe e corte, costanti e variabili, delle diverse immunoglobuline abbiano luogo seguendo un ordine e una sequenza precisi durante il differenziamento degli immunociti (v. Tonegawa e altri, 1977).
Processi di regolazione dell'espressione genica che avvengono gradualmente in seguito a trascrizione genica sono noti da lungo tempo negli organismi multicellulari. Il controllo della regolazione enzimatica esercitato dai prodotti finali è un meccanismo molto diffuso nei processi energetici e metabolici, ed è basato sulle proprietà allosteriche degli enzimi. Il presupposto necessario per la specifica strutturazione finale di proteine nascenti, allo scopo di ottenere residui funzionali, che si verifica in molti processi di differenziamento cellulare, è il riconoscimento molecolare; probabilmente anche il riconoscimento cellulare è mediato dal riconoscimento molecolare. Molecole secrete nel mezzo o associate alle membrane sono probabilmente operanti nella segregazione di popolazioni cellulari e nelle migrazioni morfogenetiche cellulari.
In alcune proteine gli esoni corrispondono a domini specifici di attività della proteina finale. Ciò apre la possibilità che nuove funzioni abbiano origine in seguito a una traslocazione del DNA delle sequenze esoniche in nuove localizzazioni. In tal modo si potrebbero formare nuovi geni combinando vari frammenti, e nuove proteine che contengono combinazioni diverse di funzioni già sperimentate. La trasposizione dei geni nel genoma e la sostituzione e dispersione delle sequenze intermedie ripetitive, che, come risulta da studi recenti, avvengono rapidamente all'interno delle popolazioni, sono un fatto largamente diffuso in piante e animali. Il riconoscimento molecolare fornisce in tal modo al genoma una enorme quantità di variazione e di funzioni combinatorie (v. Davidson, 1982; v. Britten, 1982).
Il riconoscimento molecolare che permette un aumento della diversità è al contempo una forte limitazione per la variazione. Nuove interazioni geniche e nuove funzioni proteiche richiedono il cambiamento simultaneo di elementi che interagiscono, e questi vincoli reciproci aumentano via via che cresce il numero degli elementi implicati. In tal modo la variazione dovuta a mutazione viene mantenuta entro i limiti compatibili con la funzione. Questa può essere la spiegazione della sorprendente osservazione che il numero di geni è rimasto relativamente limitato nonostante la straordinaria complessità raggiunta nell'evoluzione morfologica. Calcoli genetici, basati sulla frequenza di loci vitali per intervallo cromosomico (v. Lefevre, 1 974), e dati molecolari, basati sul numero di specie diverse di mRNA che si manifestano durante lo sviluppo, hanno permesso di attribuire a Drosophila e ad altri insetti un numero di geni variante tra cinquemila e diecimila. Gli stessi dati molecolari suggeriscono che i geni del topo non siano più di cinquantamila (Bishop e altri, 1978); questo numero rappresenta un incremento al massimo di un ordine di grandezza rispetto al numero di geni calcolato per i Batteri. È probabile che la complessità morfologica si ottenga per nuove combinazioni piuttosto che per aggiunta di nuove funzioni.
L'esistenza di tipi correlati di differenziamento cellulare, di segmenti omologhi in organismi metamerici e di genealogie cellulari omologhe in parti diverse dell'organismo suggerisce l'esistenza di raggruppamenti distinti di geni. L'analisi delle mutazioni ha mostrato che i seguenti fenomeni potrebbero essere il risultato di modeste alterazioni rispetto a un programma genetico fondamentalmente costante modificazioni secondarie di segmenti rispetto a un archetipo generale; nuove configurazioni spaziali causate da cambiamenti nella distribuzione degli stessi elementi; variazioni nella velocità relativa di sviluppo degli organi (eterocronismo) che causano la spettacolare diversità morfologica finale; infine, differenze morfologiche allometriche tra specie correlate. L'esistenza di programmi che si ripetono nello spazio rende conto di modifiche localizzate in vie di sviluppo altrimenti simili. Ciò permette che nello sviluppo si verifichi il fenomeno dell'adattamento di parti a funzioni specifiche, chiamato da Lamarck ‛selezione a mosaico'. Viceversa, l'esistenza di tipi morfologici irriducibili (Cuvier), i moderni phyla, suggerisce che vi siano modelli costruttivi diversi e discontinui. L'anatomia comparata mostra che gli organismi più primitivi di un dato phylum sono più regolari e reiterativi dei loro derivati filogenetici. Se i tipi originali rappresentino soluzioni alternative e integrate a combinazioni di interazioni geniche o se rappresentino dei campionamenti casuali tra i molti possibili tipi intermedi che avrebbero potuto riempire lo spazio morfologico, potrà essere soltanto oggetto di discussione teorica. Tuttavia, qualunque sia stata la loro origine, i principi costruttivi sono rimasti fondamentalmente gli stessi durante tutto il corso della successiva evoluzione del phylum. Può darsi che questa invarianza sia il risultato di una inerzia genetica che impedisce il verificarsi di fenomeni genetici fondamentali nello sviluppo di forme derivate. È probabile che l'attuale diversità morfologica sia dovuta a modifiche localizzate rispetto alla sottostante omogeneità di base.
6. Considerazioni conclusive
Seguendo l'indirizzo riduzionista di Weismann si è riusciti a identificare la natura molecolare dei fattori ereditari; lo scopo era quello di scoprire al livello molecolare non solo le fondamentali componenti strutturali degli organismi, ma anche le istruzioni che regolano le loro interazioni (v. Jacob, 1 970), cioè gli ‛algoritmi genetici' che determinano i diversi processi dello sviluppo embrionale.
L'autoassemblaggio è il fenomeno più gravido di conseguenze, quello in base al quale viene determinata la morfologia direttamente dall'informazione genetica. Con l'aumentare del grado di libertà delle interazioni molecolari, ai livelli di organizzazione più alti, le proprietà molecolari ( cioè il riconoscimento molecolare) rappresentano un fattore limitante piuttosto che una causa dei fenomeni. A meno che non esistano dei sottogruppi di ‛geni morfogenetici' o regolatori che favoriscano o accelerino determinate interazioni molecolari nelle vie di sviluppo, alla stregua degli enzimi nel caso delle vie metaboliche, non si può sperare di trovare nel genoma una descrizione dello sviluppo.
L'analisi molecolare, d'altra parte, ha messo in evidenza l'enorme versatilità del genoma nel modificare le funzioni, nel generarne di nuove, nell'effettuare nuove combinazioni di funzioni attraverso l'evoluzione, e questa versatilità è tale da poter rendere conto di infinite soluzioni morfologiche. Pertanto, la discontinuità che si rileva nello spazio morfologico non potrebbe essere dovuta alle limitazioni imposte nell'ambito del dominio epigenetico? Fino a che punto limitazioni di ordine fisico, ecologico, comportamentale e anche ontogenetiche impongono regole o leggi ai più alti livelli di organizzazione? Questo è l'atteggiamento epistemologico dello strutturalismo biologico, una moderna estensione della spiegazione teleologica che Driesch dava dello sviluppo, alla quale egli dette il nome di ‛entelechia' (Stent, 1974; Webster e Goodwin, 1982). Per gli strutturalisti, il significato di qualsiasi struttura, nell'ambito del comportamento, o della linguistica, o della chimica, è dato dalla sua funzione o dal contesto nel quale opera piuttosto che dai veri e propri componenti della struttura. In biologia, la selezione naturale è solo in grado di percepire le funzioni; essa darà origine a morfologie convergenti nonostante divergenze evolutive e variazioni di pattern, effettuando un rimescolamento di elementi invarianti. Per gli strutturalisti, il significato o la funzione precede e condiziona l'evoluzione morfologica; è il movimento a spiegare la comparsa di flagelli o appendici, è la percezione visiva a determinare le connessioni neurali specifiche, ed è il diverso comportamento sessuale a essere responsabile dei caratteri sessuali secondari. Questo modo di pensare pone l'accento sulla ricerca di leggi di trasformazione che rendano conto della comparsa di strutture particolari, rendano conto cioè del modo in cui un comportamento specifico possa esser fatto risalire a particolari reti di connessioni neurali, la morfogenesi alle interazioni cellulari, la specificità molecolare a combinazioni di elementi isomerici. In questo modo l'analisi spesso procede dai più alti ai più bassi livelli di organizzazione. Il valore euristico di questo approccio risiede nel fatto che esso dà uguale rilevanza a tutti i livelli organizzativi e ci stimola a ricercare leggi che siano operanti in ognuno di essi.
La moderna biologia dello sviluppo sta ancora dibattendo il vecchio problema della scelta tra il determinismo di Weismann e il finalismo di Driesch. Da allora è stato compiuto un solo passo di rilevante importanza concettuale la diversità morfologica è stabile e riproducibile perché è in qualche modo rappresentata nel genoma e solo così la variazione può essere incorporata nell'evoluzione. Pertanto, la comprensione della variazione morfologica nello sviluppo e nell'evoluzione può realizzarsi solo attraverso la comprensione del linguaggio genetico. La biologia dello sviluppo necessita ora di una grammatica genetica.
Bibliografia
Ashburner, M., Puffing patterns in Drosophila, in Handbook of genetics (a cura di R. King), vol. III, New York 1975, pp. 793-811.
Baker, B. S., Ridge, K. A., Sex and the single cell. I. On the action of major loci affecting sex determination in Drosophila melanogaster, in ‟Genetics", 1980, XCIV, pp. 383-423.
Beermann, W., Chromomerenkonstanz und spezifische Modifikation der Chromosomenstrukturen in der Entwicklung und Organdifferenzierung von Chironomus tentans, in ‟Chromosoma", 1952, V, pp. 139-198.
Bishop, J. O., Beckmann, J. S., Campo, M. S., Hastie, N.D., Izquierdo, M., Perlman, S., DNA-RNA hybridization, in ‟Philosophical transactions of the Royal Society" (Series B), 1975, CCLXXII, pp. 147-157.
Britten, R. J., Genomic alterations in evolution, in Evolution and development (a cura di J. T. Bonner), Berlin 1982, pp. 41-64.
Caviness, V. S., Pinto-Lordo, M., Evrard, P., The development of laminated pattern in the mammalian neocortex, in Morphogenesis and pattern formation (a cura di T. G. Connelly), New York 1981, pp. 103-126.
Crick, F. H. C., Split genes and RNA splicing, in ‟Science", 1979, CCIV, pp. 264-270.
Crick, F. H. C., Lawrence, P. A., Compartments and polyclones in Insect development, in ‟Science", 1975, CLXXXIX, pp. 340-347.
Davidson, E. H., Evolutionary change in genomic regulatory organization: speculation on the origins of novel biological structures, in Evolution and development (a cura di J. T. Bonner), Berlin 1982, pp. 65-84.
Dermen, H., Periclinal cytochimeras and origin of tissues in stem and leaf of peach, in ‟American journal of botany", 1953, XL, pp. 154-168.
Edgar, R. S., Wood, W. B., Morphologies of phage T4 in extracts of mutant infected cells, in ‟Proceedings of the National Academy of Sciences", 1966, LV, pp. 498-505.
Ephrussi, B., Hybridization of somatic cells, Princeton 1972.
García-Bellido, A., Pattern formation in imaginal discs, in Results and problems in cell differentiation, vol. V (a cura di H. Ursprung e R. Nüthiger), Berlin 1972, pp. 59-91.
García-Bellido, A., Homeotic and atavic mutations in Insects, in ‟American zoologist", 1977, XVII, pp. 613-629.
García Bellido, A., From the gene to the pattern: chaeta differentiation, in Cellular controls in differentiation (a cura di C. W. Lloyd e D. A. Rees), New York 1981, pp. 257-304.
García-Bellido, A., Ripoll, P., Cell lineage and differentiation in Drosophila, in Results and problems in cell differentiation, vol. IV (a cura di W. J. Gehring), Berlin 1978, pp. 119-156.
García-Bellido, A., Ripoll, P., Morata, G., Developmental compartmentalization in the dorsal mesothoracic disc of Drosophila, in ‟Developmental biology", 1976, XLVIII, pp. 132-147.
Gardner, R. L., The relationship between cell lineage and differentiation in the early mouse embryo, in Genetic mosaics and cell differentiation (a cura di W. J. Gehring), Berlin 1978, pp. 205-241.
Gerisch, G., Hess, B., Cyclic-AMP-controlled oscillations in suspended Dictyostelium cells: their relation to morphogenetic cell interactions, in ‟Proceedings of the national Academy of Sciences", 1974, LXXI, pp. 2118-2122.
Goldschmidt, R. B., Physiological genetics, New York 1938.
Grobstein, C., Cell contact in relation to embryonic induction, in ‟Experimental cell research", 1961, VIII, suppl., pp. 234-245.
Gronemeyer, H., Pongs, O., Localization of ecolysterone on polytene chromosomes of Drosophila melanogaster, in ‟Proceedings of the National Academy of Sciences", 1980, LXXVII, pp. 2108-2112.
Grüneberg, H., The genetics of the mouse, den Haag 19522.
Guillery, R. W., Casagrande, V. A., Oberdorfer, M. D., Congenitally abnormal vision in siamese cats, in ‟Nature", 1974, CCLII, pp. 195-199.
Gurdon, J. B., The control of gene expression in animal development, Oxford 1974.
Hadorn, E., Lethalfaktoren in ihrer Bedeutung für Erbpathologie und Genphysiologie der Entwicklung, Stuttgart 1955.
Hadorn, E., Dynamics of determination, in ‟Symposia of the Society for Experimental Biology", 1967, XXV, pp. 85-104.
Hamburger, V., Levi-Montalcini, R., Some aspects of neuroembryology, in Genetic neurology (a cura di P. Weiss), Chicago 1950.
Hämmerling, J., Nucleo-cytoplasmic relationships in the development of Acetabularia, in ‟International review of cytology", 1953, II, pp. 475-498.
Harris, N., Cell fusion, Oxford 1970.
Hogness, D. S., Bender, N. W., Akam, M. E., Saint, R. B., Spierer, P., Molecular organization and expression in the bithorax gene complex of Drosophila, in ‟Journal of supramolecular structure and cell biochemistry", 1981, V, suppl., pp. 385 ss.
Holtfreter, J., Gewebeaffinität, ein Mittel der embryonalen Formbildung, in ‟Archiv für experimentelle Zellforschung", 1939, XXIII, pp. 169-209.
Holtfreter, J., A study of the mechanism of gastrulation. I., in ‟Journal of experimental zoology", 1943, XCIV, pp. 261-318.
Holtfreter, J., Some aspects of embryonic induction, in ‟Growth", 1951, X, pp. 117-152.
Holtzer, H., Myogenesis, in Cell differentiation (a cura di O. Shjeide e J. De Vellis), Princeton 1970, pp. 476-503.
Hörstadius, S., The mechanics of sea urchin development studied by operative methods, in ‟Biological reviews", 1939, XIV, pp. 132-179.
Jacob, F., La logique du vivant. Une histoire de l'hérédité, Paris 1970 (r. it.: La logica del vivente. Storia dell'ereditarietà, Torino 19722).
Jacob, F., Mouse teratocarcinoma and embryonic antigens, in ‟Immunology", 1977, XXXIII, pp. 3-32.
Jacob, F., Monod, J., Genetic regulatory mechanisms in the synthesis of proteins, in ‟Journal of molecular biology", 1961, III, pp. 318-356.
Lefevre, G. Jr., The relationship between genes and polytene chromosome bands, in ‟Annual review of genetics", 1974, VIII, pp. 57-62.
Lewis, E. B., A gene complex controlling segmentation in Drosophila, in ‟Nature", 1978, CCLXXVI, pp. 565-570.
Loomis, W. F., Dictyostelium discoideum: a developmental system, New York 1975.
Metcalf, D., Moore, M. A. S., Haemopoietic cells, Amsterdam 1971.
Mintz, B., Clonal basis of mammalian differentiation, in Control mechanisms of growth and differentiation (a cura di D. D. Davies e M. Balls), London 1971, pp. 345-370.
Morgan, Th. H., Embryology and genetics, New York 1934.
Moscona, A. A., Analysis of cell recombination in experimental synthesis of tissues in vitro, in ‟Journal of cellular and comparative physiology", 1962, LX, suppl. 1, pp. 65-80.
Pauling, L. H., Itano, A., Finger, S. J., Wells, I. C., Sickle-cell anemia, a molecular disease, in ‟Science", 1949, CX, pp. 543-548.
Rawles, M. F., Origin of melanophores and their role in development of color patterns in Vertebrates, in ‟Physiological reviews", 1948, XXVIII, pp. 383-408.
Saxen, L., Karkinen-Jaaskelainen, M., Biology and pathology of embryonic induction, in Morphogenesis and pattern formation (a cura di T. G. Connelly), New York 1981, pp. 21-48.
Sengel, P., Dhoually, D., Tissue interactions in amniote skin development, in Cell interactions in development (a cura di M. Karkinen-Jaaskelainen, L. Saxen e L. Weiss), London 1977, pp. 153-169.
Spemann, H., Mangold, H., Über Induktion von Embryonalanlagen durch Transplantation artfremder Organisatoren, in ‟Wilhelm Roux Archiv für Entwicklungsmechanik der Organismen", 1924, C, pp. 599-638.
Sperry, R. W., Chemoaffinity in the orderly growth of nerve fiber patterns and connection, in ‟Proceedings of the National Academy of Sciences", 1963, L, pp. 703-710.
Steinberg, M. S., Reconstruction of tissues by dissociated cells, in ‟Science", 1963, CXLI, pp. 401-408.
Stent, G., Explicit and implicit semantic content of the genetic information, in Foundational problems in the special sciences (a cura di A. Butts e P. Hintikka), Dordrecht 1977, pp. 131-149.
Stern, C., Two or three bristles, in ‟American scientist", 1954, XLII, pp. 213-247.
Steward, F. C., Growth and organized development of cultured cells. III. Interpretation of the growth from free cell to carrot plant, in ‟American journal of botany", 1958, XLV, pp. 709-713.
Tonegawa, S., Brack, C., Hozumi, N., Pirrota, V., Organisation of immunoglobulin genes, in ‟Cold Spring Harbour symposia on quantitative biology", 1977, XLII, pp. 921-931.
Townes, P. L., Holtfreter, J., Directed movements and selective adhesion of embryonic amphibian cells, in ‟Journal of experimental zoology", 1955, CXXVIII, pp. 53-120.
Waddington, C. H., Organizers and genes, London 1941.
Webster, G., Goodwin, B. C., The origin of species: a structuralist approach, in ‟Journal of social and biological structures", 1982, V, pp. 15-47.
Weisblat, D. A., Harper, G., Stent, G., Sawyer, R. T., Embryonic cell lineages in the nervous system of the glossiphonid leech Helobdella triserialis, in ‟Developmental biology", 1980, LXXVI, pp. 58-78.
Weiss, P., Principles in development, New York 1939.
Wilson, F. B., The cell in development and heredity, New York 19253.
Wilson, H. V., On some phenomena of coalescence and regeneration in Sponges, in ‟Journal of experimental zoology", 1907, V, pp. 245-258.
Wolpert, L., Positional information and the spatial pattern of cellular differentiation, in ‟Journal of theoretical biology", 1969, XXV, pp. 1-47.