
Stress e strategie di risposta
Stress e strategie di risposta
Affrontiamo in questo saggio il complesso e dibattuto problema di definire lo stress. AI contrario di altre teorie più tradizionali, che non hanno preso in considerazione il concetto di stimolo o quello di risposta, abbiamo suggerito che lo stress è uno stato mentale creato dal nostro cervello. L'ipotesi che viene proposta è che lo stress sia una risposta del sistema nervoso centrale (SNC) a stimoli che rappresentano incertezza. Le reazioni allo stress sono regolate dall'SNC e possono variare in base alle esperienze individuali e al patrimonio genetico. Le risposte più immediate allo stress sono caratterizzate da cambiamenti nel comportamento e nell'attività del sistema nervoso autonomo, del sistema endocrino e di quello immunitario. Tali manifestazioni, tuttavia, sono correlate e interdipendenti e possono comparire in molte situazioni differenti. Secondo la teoria presentata in questo saggio le risposte da sole non definiscono uno stimolo come stressante. Viene inoltre chiarito il concetto di coping, che rappresenta una riduzione dell'incertezza derivante dalla combinazione di controllo, previsione o feedback.
Introduzione
Finora non esiste una definizione precisa di stress, che abbia un significato operativo, e che venga accettata universalmente. Sebbene vi sia una copiosa letteratura sulle conseguenze biologiche e psicologiche di una moltitudine di situazioni che sono state classificate come stress, tra gli addetti ai lavori non c'è ancora un accordo sulla definizione più calzante per questo concetto.
In molte occasioni ciò ha fatto sì che si abbandonasse il problema della definizione, focalizzandosi sulle specifiche risposte psicobiologiche a un determinato stimolo. l problemi legati alla necessità di definire lo stress diventano chiari quando si esamina l'evoluzione del concetto. Storicamente, le due principali figure a cui si deve l'uso corrente del termine stress sono W.E. Cannon e H. Selye. Cannon enfatizzò l'attivazione del sistema nervoso autonomo in risposta allo stress. Nella sua famosa descrizione della risposta allo stress come flight-fight (fuga-combattimento) egli dimostrò l'importanza della secrezione di adrenalina e noradrenalina, ormoni del sistema nervoso simpatico, rilasciati l'uno dalla porzione midollare del surrene, l'altro dalle terminazioni simpatiche. Per Cannon, lo stress veniva definito in base agli stimoli necessari per ottenere queste risposte fisiologiche. Selye (1956) fu il primo studioso a far entrare il sistema endocrino nella definizione di stress. Nella ormai famosa (nel bene e nel male) serie di esperimenti che hanno portato alla formulazione di una delle teorie più solide nella ricerca sullo stress, la sindrome generale di adattamento, egli descrisse una triade di risposte che ipotizzava costituissero lo stress stesso. Queste includevano l'ipertrofia delle ghiandole surrenali, le ulcerazioni della parete dello stomaco e l'involuzione del timo. Questa triade di risposte implicava: il coinvolgimento del sistema endocrino, con un aumento nella secrezione di adrenocorticotropina (ACTH, Adrenocorticotropic Hormone) dall'ipofisi e di corticosterone dalle ghiandole surrenali; un aumento nell'attività del sistema nervoso autonomo, che si manifestava nelle ulcerazioni dello stomaco; l'attivazione del sistema immunitario (involuzione del timo). Nel 1956 Selye enunciò che lo stress includeva tutte queste manifestazioni e che era la presenza di questo complesso di risposte a definido come tale. Inoltre, secondo Selye, queste risposte si manifestavano in maniera aspecifica in seguito a qualsiasi tipo di stimolazione, indipendentemente dalla natura dello stimolo stesso. Perciò, in uno dei primi tentativi di definire lo stress in termini operativi, esso venne identificato in base al tipo di risposta che era in grado di provocare.
Nel 1975 J.W. Mason presentò un'altra versione del concetto di stress che mise in discussione la dottrina enunciata da Selye. Mason enfatizzò il ruolo cruciale degli "approcci psicologici messi in atto nelle reazioni emotive o di attivazione a seguito di eventi pericolosi o spiacevoli che si possono manifestare nella vita quotidiana". Egli riteneva che la cosiddetta non specificità delle risposte endocrine allo stress fosse dovuta alla componente emozionale legata al fatto di sperimentare stimoli capaci di indurre stress. Perciò la risposta non specifica descritta da Selye è principalmente di natura comportamentale o psicologica e "i processi interpretativi alla base delle risposte fisiologiche aspecifiche probabilmente coinvolgono un livello di funzionalità dell'SNC più elevato di quanto non si fosse precedentemente supposto". Nel 1979 J.W. Hennessye S. Levine ipotizzarono che la componente endocrina della risposta allo stress, l'asse ipotalamo- ipofisisurrene, fosse un indicatore sensibile dello stato di allerta dell'organismo e che perciò la sua risposta riflettesse un'aumentata attivazione emozionale. Questa teoria rappresentava una parziale estensione della posizione presentata da Mason. Grazie alle teorie di Mason e Levine, dunque, il cervello divenne una componente essenziale del concetto di stress.

Per molti aspetti, Mason e Selye cercavano di analizzare lo stress da due angolature molto diverse. La teoria di Selye si basava su un'intensa serie di studi che utilizzava procedure sperimentali altamente invasive e di lungo termine. È molto improbabile che questo tipo di studi possa essere ripetuto considerate le nuove regole sulla sperimentazione animale. Contrariamente a quelli di Selye, molti dei paradigmi sperimentali utilizzati da Mason erano caratterizzati dell'esposizione acuta a stimoli stressanti. Esiste ora una vasta letteratura scientifica in grado di documentare che la neurobiologia dello stress varia notevolmente a seconda che si operi in condizioni acute o croniche. I problemi che caratterizzano questo campo di studi sono stati enunciati forse nel modo più efficace da B.T. Engel (1985) quando, nella sua severa critica allo stress, notò come questo termine fosse utilizzato in molte accezioni differenti: come verbo, "stressare", come nome proprio, "stress", e come complemento di termine, la risposta "allo stress". In base a ciò lo stress è stato definito come lo stimolo (input), la risposta (output) o qualche variabile intermedia. Seguendo un approccio un po' differente, Levine e H. Ursin (1991), nel tentativo di dame una definizione, descrissero lo stress come formato da tre componenti: uno stimolo (lo stressare), l'elaborazione dello stimolo da parte del cervello (lo stress) e la risposta, il risultato dell'elaborazione (la risposta allo stress). Secondo Levine e Ursin qualsiasi definizione di stress deve includere tutte e tre queste componenti (fig. 2). Questo approccio non è completamente soddisfacente poiché, se esaminiamo le componenti di questa triade, la conclusione finale è che, nell'analisi del concetto di stress, ciò che assume un'importanza critica è quella centrale, il 'processore'. In questo saggio cercheremo di definire lo stress come uno stato creato dal processore, il sistema nervoso centrale (SNC). Il punto focale è che lo stress dipende dalla risposta dell'SNC a una data stimolazione e che la risposta o il complesso di risposte specifiche a esso generalmente associate sono regolate dall'SNC. Per difendere l'ipotesi appena enunciata si rende necessario discutere tanto la variabile (o le variabili) in entrata che le componenti della risposta allo stress. Ci auguriamo che la cruciale importanza dell'SNC si manifesti al momento dell'esame delle relazioni tra tutte le componenti che abbiamo associato con lo stress.
Lo stimolo: lo stressare
A un'analisi superficiale, i diversi stimoli classificati come capaci di indurre una risposta allo stress non possiedono ovvie caratteristiche fisiche in comune. L'elenco di tali stimoli stressanti include sia quelli cosiddetti fisici (per esempio, la simulazione di un intervento chirurgico, l'emorragia, lo shock elettrico, il rumore intenso, l'iniezione di sostanze tossiche) sia stimoli classificati come puramente di natura psicologica (per esempio, la frustrazione, la separazione tra madre e prole, la separazione da gruppi di conspecifici, un aumento nella quantità di lavoro, la perdita degli affetti). Questa lista non pretende in alcun modo di essere onnicomprensiva. Poiché tutti questi stimoli sono in grado di attivare una o più risposte associate allo stress, gli studiosi hanno cercato di individuare alcune proprietà comuni a questi elementi che spieghino la loro capacità di provocare risposte molto simili. Mason e Levine hanno affermato in diverse occasioni che l'elemento comune a tutti quegli stimoli definiti stressanti è di natura psicologica o emozionaIe. Mason ha dimostrato che se uno stimolo termico viene presentato improvvisamente si ha un'attivazione dell'asse ipotalamo-ipofisi-surrene, mentre se il soggetto sperimentale viene esposto allo stesso aumento di temperatura in maniera graduale non si assiste ad alcuna risposta. Sulla base di questo e di altri studi egli concluse che la differenza tra questi risultati era la risposta emozionale associata all'improvviso cambiamento di temperatura (da quella ambientale a una più elevata). Levine, nel suo tentativo di trovare fattori tramite i quali raggruppare i diversi stimoli che provocano una risposta allo stress, ha affermato che la loro caratteristica comune era una mancanza di informazione (incertezza). L'incertezza potrebbe derivare sia dall'assenza totale di informazione, sia dall'incapacità a espletare un'azione richiesta, quale ricevere un rinforzo positivo o evitame uno negativo. Anche l'assenza o la perdita di controllo e la mancanza di predizione sono in grado di produrre un'incertezza che, processata dal cervello, mette in moto gli eventi neurobiologici che risultano nella risposta allo stress.
Si è tentato in molti modi di discriminare tra diversi tipi di stress quali, per esempio, tra stress neurogenico e quello psicogenico, stress fisico e quello psicologico e così via. Il problema con ciascuna di queste classificazioni è la difficoltà di separare in maniera chiara le componenti fisiche e psicologiche da molti, cosiddetti, stimoli fisici stressanti. Alcuni dati di neuroanatomia sembrerebbero avvalorare l'ipotesi che non tutti gli stimoli attivino le stesse vie nervose. Esistono, tuttavia, stimoli puramente psicologici che non contengono alcuna componente fisica. Perciò è possibile identificare alcuni elementi comuni che rendono queste situazioni psicologiche stressanti.
Secondo Levine e Ursin, la visione contemporanea dello stress può essere ben caratterizzata da un modello impositivo. Questa visione deriva principalmente da una voluminosa serie di studi in cui, per indurre una risposta allo stress, venivano usati stimoli estremamente traumatici. Esistono inoltre alcune specifiche situazioni psicologiche in grado di adattarsi al modello impositivo, quali per esempio, la sconfitta. Tuttavia, da un punto di vista psicologico, è stato ipotizzato (Levine e Ursin, 1991) che stimoli in grado di indurre stress possiedano la proprietà di rappresentare l'assenza di componenti critiche nell'ambiente. Con ciò si intende la mancanza di informazione (incertezza o non prevedibilità) relativamente alla possibilità di ottenere un risultato positivo o di evitame uno negativo. Perciò gli stimoli stressanti, o i segnali che li rappresentano, indicano la mancanza di qualcosa che viene ritenuta estremamente importante per l'organismo. Dunque, è possibile classificare gli stimoli stressanti non solo in termini di manifestazione di un evento, ma anche in termini di omissione o di assenza di componenti critiche dall'ambiente. La somiglianza tra questa visione e la nozione precedentemente enunciata di incertezza dovrebbe essere evidente. Molti stimoli stressanti possono essere descritti in questo modo, per esempio, la frustrazione, la perdita della madre, la separazione dai conspecifici, la perdita del lavoro e così via. Noi proponiamo che, forse, insita in tutti gli eventi psicologici che vengono visti come stress c'è l'omissione o l'assenza di informazione critica (incertezza) che mette in atto una cascata di risposte regolate dal cervello.
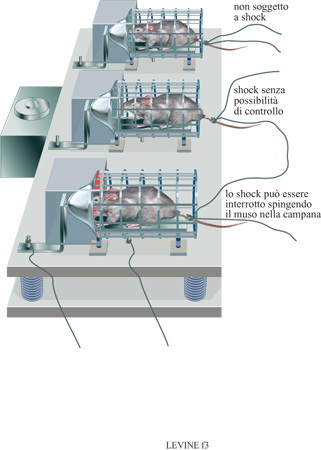
Questo campo di studi deve molto a J.M. Weiss (1972) per il suo lavoro pionieristico sul coping. È grazie a questi studi che sono state identificate alcune variabili di cruciale importanza per la comprensione della regolazione della risposta a eventi stressanti. Gli esperimenti di Weiss hanno dimostrato come una risposta fisiologica (ulcera allo stomaco) a uno stimolo fisico ben definito quale lo shock elettrico, può essere modificata se si permette all'animale di esercitare qualche forma di controllo su di esso (v. anche il saggio di G. Mason, in questo volume). Ciò può avvenire, per esempio, dando all'animale la possibilità di regolare la lunghezza o l'intensità dello shock (controllo), fomendogli indicazioni sull'inizio o sulla fine dello stimolo (prevedibilità), o informandolo sull'efficacia di una sua eventuale risposta (feedback). In questi esperimenti Weiss utilizzò ciò che viene definito un disegno triadico. Un gruppo di animali sperimentali poteva porre fine allo shock eseguendo l'appropriata risposta operante, mentre un altro gruppo di soggetti sperimentali riceveva la stessa stimolazione con lo shock, ma non poteva mettere fine allo stimolo. l soggetti di un ulteriore gruppo sperimentale venivano posti nell'apparecchiatura senza ricevere alcuno shock (fig. 3). l risultati di questi studi hanno dimostrato come gli animali che non avevano avuto la possibilità di porre fine allo shock presentavano le ulcerazioni più evidenti, nonostante fossero stati sottoposti a una stimolazione di intensità simile a quella degli altri gruppi sperimentali. In esperimenti successivi Weiss presentò agli animali un segnale sonoro che precedeva, perciò predicendolo, l'inizio dello shock. Se confrontati con soggetti non esposti allo stimolo uditivo (non prevedibilità), quelli che avevano udito il segnale presentavano un numero minore di ulcere. Inoltre, egli dimostrò che anche un segnale presentato successivamente a una risposta operante positiva era in grado di prevenire, almeno in parte, le ulcerazioni allo stomaco (feedback).
Queste tre variabili principali, controllo, feedback e prevedibilità, hanno alcuni elementi in comune, in particolare, la capacità di ridurre o eliminare l'incertezza. Il controllo fornisce all'organismo la capacità di eliminare, o almeno regolare, la durata e l'intensità dello stimolo. La prevedibilità, per definizione, dovrebbe avere lo scopo di ridurre l'incertezza. Anche il feedback può ridurre l'incertezza poiché esso fornisce informazioni sull'efficacia e sul successo della risposta. Possiamo ipotizzare che i processi cognitivi e comportamentali che riducono l'incertezza determinino la diminuzione o l'eliminazione di una o più risposte fisiologiche a un evento stressante. Tali modificazioni della risposta allo stress risultanti da una, o da una combinazione, delle variabili esaminate rappresenterebbero, secondo questo approccio, il coping. Forse il più potente dei meccanismi che costituiscono il coping è il controllo. Sebbene negli esperimenti originali la variabile dipendente fosse rappresentata dalle ulcere allo stomaco, si è visto che altri sistemi fisiologici sono influenzati dal concetto di controllo. Questi includono il sistema nervoso autonomo (SNA), i processi neurochimici, le risposte neuroendocrine e nervose. Inoltre, gli effetti del controllo sono stati dimostrati in numerose specie di mammiferi quali ratti, cani, primati non umani e nell'uomo. È importante sottolineare che il concetto di coping utilizzato da Weiss e altri è molto diverso dall'approccio di R.S. Lazarus e S. Folkman (1984). Essi definirono il coping in questo modo: "il coping ha a che vedere con i tentativi di risolvere un problema che un individuo mette in atto quando le richieste che gli vengono sottoposte sono molto importanti per il suo essere, come in una situazione di estremo pericolo o in una particolarmente favorevole, e quando queste richieste attingono alle sue risorse adattative". Tale defmizione implica molti concetti. In primo luogo, enfatizza l'importanza del contesto emozionale nel coping; in secondo luogo, permette l'inclusione sia del lato negativo (o stress) delle emozioni sia del loro lato positivo quale potenziale mezzo di realizzazione o gratificazione. Infine, dà risalto a quelle prove che non sono di routine o automatizzate, cioè quelle degli esiti incerti e che approssimano ai limiti delle capacità adattative individuali.
Ciò che appare chiaro dalla defmizione di coping proposta da Lazarus è che alcuni processi di coping possono aumentare il rischio di assumere un comportamento non adattativo, mentre altri potrebbero migliorare l'adattamento e ridurre il rischio di esiti negativi. Perciò questa definizione di coping contempla la possibilità di adottare sia le strategie di coping efficaci sia quelle che non lo sono. Al contrario, nella teoria proposta da Weiss non può esservi un coping inefficace. Il coping è definito come una risposta con esito positivo che si basa per lo più sulla riduzione degli indici patologici o fisiologici.
Se si assume che lo stress è prodotto dall'incertezza generata dall'assenza di adeguata informazione o di controllo, si pone immediatamente il problema del modo in cui tale incertezza venga percepita ed elaborata. Non è possibile predire gli effetti di un determinato stimolo sull'organismo senza sapere in che modo tale stimolo, in una determinata situazione, viene elaborato dal cervello di ciascun individuo. Ciò infatti dipende, almeno in parte, dall'informazione relativa a quello stimolo e a quella situazione in esso immagazzinata. In altre parole, l'elaborazione di un evento come incerto dipende anche dalle precedenti esperienze. Inoltre, per determinare se una situazione si presenta come incerta, l'organismo ha bisogno di un comparatore: l'unico capace di questa funzione è il sistema nervoso centrale.
Elaborazione dello stimolo: lo stress
L'attivazione fisiologica in risposta all'incertezza è una delle componenti centrali del modello presentato da E.N. Sokolov (1960). Sebbene il modello di Sokolov fosse un tentativo di spiegare il processo di abituazione, una sua interpretazione più ampia potrebbe anche spiegare perché l'incertezza determini le risposte solitamente associate allo stress. Quando il soggetto sperimentale viene esposto a uno stimolo improvviso, ne consegue una risposta di orientamento. Lo stimolo è in grado di evocare risposte sia comportamentali sia fisiologiche. Le componenti fisiologiche della risposta di orientamento includono l'attivazione dell'encefalo, misurata originariamente come un aumento dell'attività elettrica, modifiche nell'attività del sistema nervoso autonomo e un aumento nell'attività neuroendocrina. Recentemente, tecniche più raffinate come la misurazione dell'attivazione di geni precoci, quali per esempio c-fos, hanno dimostrato che, in seguito a stimoli inaspettati, si ha un'attivazione generalizzata a livello del sistema nervoso centrale. L'espressione di questi proto-oncogeni è stata riconosciuta come un indicatore dell'attivazione funzionale dell'attività nervosa in seguito a diverse situazioni, incluso lo stress. Se lo stimolo viene presentato frequentemente, la maggior parte di queste reazioni diminuisce gradualmente per poi scomparire del tutto.
Il modello di Sokolov si basa su un comparatore in cui stimoli o situazioni nuove vengono confrontati dal sistema nervoso centrale con una rappresentazione di eventi precedenti. Questo processo di comparazione genera aspettative. La violazione di tali aspettative porta a una risposta di attivazione (stress). Perciò l'incertezza può essere vista come violazione di un'aspettativa. Secondo questa prospettiva, la valutazione degli stimoli in entrata da parte dell'individuo è cruciale per determinare se essi provocheranno o meno una risposta di stress. La valutazione dello stimolo, più che lo stimolo di per sé, è sotto il controllo del sistema nervoso centrale. Non sorprende, dunque, che due eventi identici producano, sia nell'uomo sia nelle altre specie animali risposte allo stress di grandezza molto variabile e attivazioni nel sistema di risposta. Il destino di ciascuno stimolo dipende da fattori centrali, ovvero dallo stato del processore primario, il cervello. Nella maggior parte dei casi, perciò, l'attività di base dell'SNC consiste nell'attuare una qualche forma di comparazione. Ciò sembra essere vero per risposte semplici quali il riflesso di orientamento (Sokolov, 1960). La novità può essere individuata solo in seguito a una valutazione del segnale che ha luogo nel cervello. Ciò è vero anche nel caso dell'incertezza. L'informazione ha accesso a strutture del sistema nervoso centrale che la confrontano con quella precedentemente immagazzinata. Questi confronti possono essere considerati come l'aspettativa.
R.C. Bolles (1972) ha ipotizzato che vi siano diversi tipi di aspettative. Queste sono, nell'ordine, aspettative rispetto allo stimolo e rispetto all'esito della risposta. Le aspettative rispetto allo stimolo hanno a che vedere con l'immagazzinamento di informazioni sulle relazioni temporali tra stimoli (si veda il condizionamento classico). Le aspettative rispetto all'esito della risposta derivano dall'apprendimento delle conseguenze di specifiche azioni. Tali aspettative riguardano gli esiti della risposta, e sono in relazione con il condizionamento strumentale. Poiché il coping può essere inquadrato nel contesto delle aspettative sugli esiti della risposta, esso è il risultato di un processo di apprendimento. Un essere vivente si aspetta che l'esito sia positivo con un alto livello di probabilità, e questa aspettativa riduce nell'individuo la sua risposta allo stress.
L'ottenimento di esiti di risposta positivi dipende da alcuni meccanismi di coping descritti precedentemente: il controllo, la capacità di agire attivamente in presenza di stimoli positivi o negativi e di imparare le conseguenze di tali atti. Se la risposta ha un esito positivo, quale l'ottenere un rinforzo positivo o l'essere in grado di sfuggire o di evitare un evento fastidioso, il cervello immagazzinerà quest'informazione come un' aspettativa dell'esito positivo. Per questo tipo di apprendimento è necessario non solo che tali eventi positivi accadano, ma anche che il cervello sia capace di codificare questi risultati. L'incapacità al coping, e di conseguenza, l'incapacità a registrare gli esiti positivi di una risposta, porta ad alti livelli di stress. Lo stabilirsi di risposte con esito positivo riduce l'attivazione fisiologica sia nell'uomo (Ursin, 1988) sia nell'animale. Perciò, eventi puramente psicologici, quali l'assenza di informazione (novità), il non essere all'altezza delle aspettative, o l'omissione (frustrazione) di un rinforzo positivo precedentemente atteso, sembrano essere elementi critici in grado di provocare stress.
Differenze individuali
Poiché in questo tentativo di defmire lo stress il fattore cruciale è rappresentato dalla variabile da elaborare, è importante sottolineare che alcuni eventi possono modificare permanentemente lo stato cerebrale di un organismo con la conseguenza che due stimoli, apparentemente identici, possono dare luogo a risposte differenti, sia da un punto di vista comportamentale sia fisiologico. Di conseguenza, non è possibile identificare tutti i fattori che potrebbero essere alla base delle differenze individuali. È stato proposto che alcune di queste differenze abbiano una base genetica (Suomi, 1987). Tuttavia, molte evidenze sperimentali mostrano che alcune esperienze avute nelle fasi di sviluppo del cervello modificano il processo di elaborazione dell'informazione. È stato, infatti, ampiamente documentato che le esperienze precoci (prenatali e postnatali) possono modificare in maniera permanente la struttura e la funzionalità cerebrale (Meaney et al., 1993). In passato si è assistito a un rinnovato interesse per questo campo di studi con la messa a punto di nuovi paradigmi sperimentali.
La manipolazione che è stata studiata più estesamente durante l'ontogenesi è quella frequentemente chiamata early handling o manipolazione precoce. Questo termine in effetti non è molto appropriato. La manipolazione sperimentale consiste semplicemente in un breve periodo di separazione dalla madre. È stato dimostrato come tale procedura modifichi gli esiti dell'esposizione allo stress nel soggetto adulto. Le risposte comportamentali, endocrine, neurochimiche e immunologiche a stimoli identici risultano molto diverse in adulti manipolati precocemente. In generale, gli effetti della manipolazione precoce si riflettono in una diminuzione generalizzata nella risposta allo stress. Recentemente si è appurato che altre forme di esperienze precoci producono soggetti adulti con una risposta allo stress molto marcata. Lunghi periodi di separazione materna (3 ore giornaliere, dal secondo al quattordicesimo giorno postnatale) sono un esempio di una manipolazione che determina nel soggetto adulto una risposta allo stress accentuata, sia dal punto di vista comportamentale sia endocrino. Anche le esperienze avute in età adulta possono causare modificazioni a lungo termine dell'SNC. Al contrario delle esperienze precoci, questi cambiamenti sembrerebbero essere reversibili. L'esposizione a uno shock inevitabile e non anticipabile, per esempio, fa sì che il soggetto diventi iperreattivo agli stimoli e mostri risposte esagerate. Tali risposte anormali non sembrano dipendere dal tipo di paradigma sperimentale e sono generalizzabili a numerosi stimoli. Un esempio eclatante degli effetti dell'esposizione a stimoli avversivi non anticipabili e incontrollabili è il fenomeno della learned helplessness (impotenza appresa).
Le differenze individuali nella risposta allo stress possono anche dipendere dal contesto in cui lo stimolo capace di indurre stress viene presentato. Nell'uomo, è stato dimostrato che la presenza di un supporto sociale aumenta la capacità di sopportare alcuni eventi stressanti (Isaacson e Janzon, 1986). Simili effetti di riduzione dello stress a carico delle interazioni sociali sono stati dimostrati anche nei primati non umani che vivono in strutture sociali complesse (Levine, 1993). Per riassumere, il punto di vista presentato finora si basa sulla capacità cerebrale di elaborare informazione riguardo la possibilità di ottenere esiti positivi o di evitare quelli negativi. Gli stimoli in grado di indurre stress agiscono grazie a una mancanza di informazione (incertezza). I meccanismi di coping sono efficaci se sono in grado di fornire elementi informativi tali da ridurre l'incertezza.
La risposta allo stress
La risposta generica a stimoli in grado di generare stress è definita risposta allo stress. La risposta allo stress dipende dal modo in cui il cervello percepisce gli stimoli come stress, e attiva specifici sistemi neuronali che portano a una cascata di risposte. Si presume che questi eventi siano la conseguenza dell'attivazione di uno o più meccanismi neuronali capaci di produrre un esito misurabile, di natura comportamentale e corrispondente a una o più risposte fisiologiche; raramente durante lo stress viene attivato un solo sistema fisiologico. Nella situazione più comune, quando sperimenta eventi che vengono designati come stress, l'organismo mette in atto una serie di risposte. Sistemi diversi possono dar luogo a risposte con dinamiche differenti, quali per esempio, diversi profili temporali, ma caratterizzate da emissione sincrona. Perciò il numero di risposte che si osservano è limitato solo dalla capacità del ricercatore di misurarle. Lo stesso concetto può essere esposto affermando che l'attivazione nervosa raramente avrà come risultato un solo tipo di risposta. Nella maggior parte dei casi, il ricercatore concentrerà la propria attenzione sul sistema che ritiene essere il più interessante e che ha mostrato di essere un indicatore affidabile dello stress. La letteratura scientifica è ricca di esempi sulle molte facce che la risposta allo stress può assumere.
La biologia molecolare ha fornito una vasta gamma di nuove metodologie per studiare la neurobiologia dello stress. Un primo effetto è che le nostre conoscenze sono aumentate enormemente, almeno quanto la nostra confusione. Ciò che sembra chiaro è che tutti i sistemi nervosi coinvolti nella regolazione delle risposte sono in comunicazione costante tra loro. Per fare solo alcuni esempi, i neuropeptidi, che rivestono un ruolo diretto nella regolazione del sistema ipotalamo-ipofisi-surrene, hanno anche la capacità di influenzare il sistema nervoso autonomo e il comportamento. Per esempio, i principali neurotrasmettitori della famiglia delle catecolammine rispondono tutti agli stessi stimoli, sia pure ognuno in un modo diverso. Molti di questi neurotrasmettitori sono coinvolti in maniera diretta nella regolazione della secrezione endocrina. Le relazioni reciproche tra i sistemi endocrino, autonomo e immunitario sono molto ben documentate. Il problema di questo campo d'indagine è che, finora non conosciamo il vocabolario alla base di tale dialogo, per non parlare della grammatica.
A questo punto non vi è nessuna ragione a priori per selezionare un indicatore specifico della risposta allo stress quale solo e unico marcatore dello stress. Un buon marcatore dello stress è quello che funziona in un determinato contesto sperimentale. Esso deve essere affidabile, misurabile e sensibile. Tuttavia, nessuna particolare risposta fisiologica che risponda a tutti questi requisiti è più valida di un'altra. Ci sono alcune importanti considerazioni da fare. Sebbene i diversi stimoli possano venire regolati in maniera confrontabile da fattori psicologici, non si può dare per scontato che vi sia una relazione di causa-effetto con le diverse risposte. Il controllo, per esempio, può ridurre le ulcere allo stomaco, e contemporaneamente l'entità della secrezione dei glucocorticoidi. Tuttavia, i livelli di glucocorticoidi non sono la causa delle ulcere, che dipendono invece dalla mancanza di controllo. I maggiori sistemi fisiologici che sono stati utilizzati come indicatori dello stress sono il sistema nervoso centrale, l'asse lobo limbico-ipotalamo-ipofisi-surrene e il sistema immunitario.
Il sistema nervoso autonomo
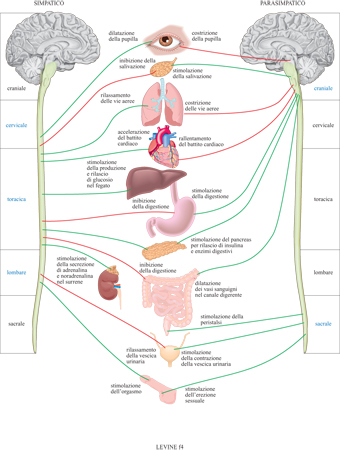
Il sistema nervoso autonomo è uno dei primi sistemi fisiologici di cui è stato dimostrato il ruolo nella risposta allo stress: esso ha il compito di regolare molte delle risposte periferiche associate allo stress. Il cervello regola l'azione dell'SNA tramite proiezioni neuronali che dall'encefalo afferiscono alla colonna dorsale e agli organi periferici (fig. 4). Il sistema nervoso autonomo è suddiviso in due sistemi apparentemente opposti: il sistema nervoso simpatico e quello parasimpatico. Il primo attiva molte delle risposte alle situazioni definite d'emergenza. Queste includono i cambiamenti nella frequenza cardiaca, un'aumentata sudorazione, variazioni nella conduttanza della pelle, un aumento della pressione sanguigna e della motilità gastrica, la secrezione di noradrenalina dalle terminazioni nervose simpatiche e di adrenalina dalla porzione midollare del surrene.
Le risposte dell'SNA sono in gran parte involontarie, sebbene ci sia qualche evidenza sperimentale che il biofeedback (procedura di apprendimento del controllo volontario di risposte viscerali) possa indurre qualche regolazione volontaria di questo sistema. Le varie componenti del sistema nervoso simpatico vengono attivate per preparare il corpo a rispondere in modo appropriato alle emergenze reali o percepite come tali. Esse sono coinvolte nella reazione di combattimento-fuga. Poiché queste reazioni vengono regolate da connessioni neuronali dirette, esse sono molto rapide. Alcune delle risposte di questo sistema utilizzano come mediatori chimici la noradrenalina e l'adrenalina. Poiché il rilascio di questi neurotrasmettitori dai nervi simpatici e dalla porzione midollare della ghiandola surrenale è anch'esso regolato da un'innervazione diretta, tale processo deve essere molto veloce. Il sistema nervoso parasimpatico gioca in generale un ruolo opposto, mediando le attività vegetative. Per esempio, l'attivazione della componente parasimpatica del sistema nervoso autonomo, subito dopo un pasto abbondante, determina uno stato di calma e di letargia.
Le proiezioni simpatiche e parasimpatiche dall'encefalo raggiungono i loro specifici organi di innervazione e, una volta attivate, producono su di essi risultati opposti. Chiaramente l'attivazione simultanea di entrambi i rami del sistema nervoso autonomo sarebbe controproducente; si otterrebbe una situazione analoga a quella in cui si mettesse il piede contemporaneamente sull'acceleratore e sul freno. Affinché ciò non accada le regioni cerebrali che attivano la componente simpatica durante lo stress allo stesso tempo inibiscono quella parasimpatica.
Il sistema endocrino
L'ipofisi, tradizionalmente considerata come la ghiandola principale, seceme la maggior parte degli ormoni peptidici che regolano la secrezione delle ghiandole periferiche. Questi ormoni ipofisari prendono il loro nome dagli organi bersaglio; per esempio, l'ormone di stimolazione tireotropo TSH viene così chiamato perché la sua azione è quella di stimolare il rilascio e la sintesi di tiroxina dalla ghiandola tiroide. Noi concentreremo la nostra attenzione sull'ormone ipofisario adrenocorticotropina (ACTH), che regola la secrezione di glucocorticoidi da parte della ghiandola surrenaIe. Esistono delle differenze di specie nel tipo di glucocorticoide che viene secreto dalla ghiandola surrenale. Nei roditori, il glucocorticoide principale è il corticosterone, mentre nei canidi, nei primati e nell 'uomo, il glucocorticovide più potente è il cortisolo. L'adrenocorticotropina e il corticosterone, o il cortisolo, sono considerati i principali ormoni dello stress.
Nel 1948 G.W. Harris presentò una teoria che avrebbe rivoluzionato il mondo dell'endocrinologia. Il tema centrale era elegante nella sua semplicità. Sebbene in questa discussione ci interesseremo solamente della regolazione degli ormoni dello stress, i principi stabiliti da Harris possono essere applicati alla maggior parte delle ghiandole endocrine del corpo.
Harris postulò che l'ipofisi fosse un organo dipendente, a sua volta regolato da peptidi sintetizzati e immagazzinati nel cervello e rilasciati e trasportati attraverso i vasi portali che mettono in connessione anatomica l'ipofisi con il cervello. Se la sintesi di questi neuropeptidi venisse bloccata, mediante lesioni o un altro tipo di danno dell'ipotalamo, o se il loro trasporto fosse interrotto, per esempio, interrompendo il flusso sanguigno dall'ipotalamo all'ipofisi, questa ghiandola non sarebbe più in grado di rilasciare i suoi ormoni, inattivandosi.
Questa teoria ha gettato le fondamenta per la creazione della neuroendocrinologia. La teoria di Harris si riferiva specificamente al controllo di quegli ormoni secreti dall'ipofisi anteriore. E. Scharrer (1963) aveva precedentemente dimostrato che la vasopressina, un ormone dell'ipofisi posteriore, viene sintetizzata nel cervello per essere poi direttamente trasportata all'ipofisi posteriore. Originariamente, Harris aveva postulato che per ciascun ormone ipofisario vi fosse un corrispondente ormone di rilascio sintetizzato nell'ipotalamo, una piccola struttura vicino alla base del cervello. Oggi sappiamo che l'ipotalamo contiene un insieme di fattori, sia di rilascio sia inibitori, che regolano l'ipofisi, la quale a sua volta controlla la secrezione delle ghiandole periferiche. In alcuni casi il cervello blocca il rilascio di un ormone ipofisario mediante un singolo fattore inibitore. Più spesso, gli ormoni ipofisari sono regolati dall'azione coordinata sia di fattori di rilascio sia di fattori inibitori. Per complicare ulteriormente il quadro, la secrezione di almeno uno dei fattori di rilascio ipotalamici più critici coinvolto nella risposta allo stress, l'ormone di rilascio della corticotropina, dipende da una vasta gamma di stimoli afferenti da altre strutture nervose. Sono state descritte alcune delle vie nervose multiple che attivano il rilascio di corticotropina (Sawchenko, 1991).
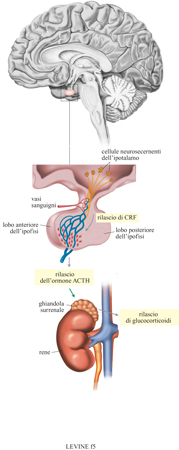
Abbiamo già illustrato due dei più importanti ormoni dello stress, l'adrenalina e la noradrenalina. Nella periferia, gli altri due ormoni importanti sono l'adrenocorticotropina e il corticosterone. L'attivazione di questi ormoni avviene a partire dagli impulsi nervosi che stimolano il rilascio di corticotropina dall'ipotalamo nel sistema portale per poi raggiungere l'ipofisi. La corticotropina agisce, quindi, sulle cellule ipofisarie aumentando la sintesi e la secrezione di adrenocorticotropina che, a sua volta, stimola le cellule della corteccia surrenale a sintetizzare e rilasciare il corticosterone (fig. 5). Quest'ultimo e le secrezioni del sistema simpatico sono insieme responsabili della maggior parte degli eventi che si susseguono nel corpo in seguito a stress. Come nel caso di molti processi fisiologici, esistono alcuni meccanismi in grado di inibire questa cascata di eventi. Il meccanismo principale è costituito dalla capacità del corticosterone di inibire la secrezione di corticotropina e di adrenocorticotropina agendo con feedback negativo su alcune strutture del sistema nervoso centrale quali l'ippocampo, l'ipotalamo e l'ipofisi.
Questa descrizione del sistema endocrino non evidenzia il fatto che esistono ancora molti aspetti oscuri del meccanismo che porta allo stress. Per esempio, altri neuropeptidi sono coinvolti nella risposta allo stress; è stato infatti dimostrato come un importante neuropeptide, la vasopressina, agisca in maniera sinergica con la corticotropina. Inoltre, sebbene le risposte allo stress solitamente coinvolgano gli ormoni che abbiamo menzionato, le dinamiche della risposta possono variare a seconda del tipo di stimolo o dell'intensità dello stimolo stesso. Diversamente dall'attivazione acuta, la presentazione cronica di stimoli in grado di provocare stress modifica il pattern degli ormoni rilasciati. In generale, la maggior parte degli stimoli stressanti provoca i cambiamenti ormonali che abbiamo sottolineato, tra questi sicuramente quelli relativi alle componenti glucocorticoide e simpatica. Tuttavia, la rapidità e l'intensità delle modificazioni del rilascio ormonale possono dipendere dallo stimolo. È come se diversi stimoli producessero una diversa 'firma ormonale', specifica per quel tale stimolo, e che può anche differire in soggetti diversi.
Il sistema immunitario
Sebbene gli aspetti essenziali del sistema nervoso autonomo e del sistema endocrino possano essere descritti efficacemente, per quanto in maniera succinta e semplificata, con il sistema immunitario il compito è più arduo. Ciò deriva dalla particolarità delle problematiche presentate da questo sistema. Negli anni passati abbiamo assistito all'emergere di una disciplina interamente nuova definita con vari epiteti: psicoimmunologia, psiconeuroimmunologia, psiconeuroendoimmunologia, neuroimmunomodulazione e così via. Parte del problema consiste nel fatto che, all'interno del sistema immunitario, avvengono diversi tipi di processi immunitari. Per esempio, abbiamo un'immunità mediata da cellule e un'immunità umorale. Inoltre, il funzionamento del sistema immunitario sembra essere molto diverso da quello degli altri sistemi (nervoso autonomo ed endocrino). Nel sistema nervoso autonomo e in quello endocrino, l'attivazione dell'ormone periferico è mediata centralmente. Al contrario, i prodotti del sistema immunitario possono, dalla periferia, attivare alcuni meccanismi nervosi. L'esempio più evidente di questo processo è rappresentato dalla relazione tra le citochine e il rilascio di corticotropina, campo di ricerca che ha recentemente ricevuto notevole attenzione. È stato dimostrato che alcune cito chine prodotte dai macrofagi, in particolare l'interleuchina-l, agiscono direttamente sull'ipotalamo per indurre la secrezione dell'ormone di rilascio della corticotropina che, a sua volta, determina un aumento della secrezione dei glucocorticoidi che possono tamponare attivamente molte delle funzioni del sistema immunitario. Perciò la regolazione omeostatica del sistema immunitario avviene attraverso meccanismi diversi da quelli che regolano l'attività endocrina.
La maggior parte delle recenti evidenze sperimentali indica che i sistemi endocrino, nervoso autonomo, immunitario e centrale interagiscono e rispondono a stimoli farmacologici e fisiologici in maniera coordinata. La presenza in tutti questi sistemi degli stessi ormoni neurotrasmettitori e recettori rafforza l'ipotesi che tra essi esista una comunicazione bidirezionale. Più in particolare le risposte allo stress da parte dei sistemi nervoso centrale, endocrino e immunitario forniscono un esempio di risposta integrata. Nel caso della relazione tra il sistema immunitario e quello endocrino, i due principali candidati del loro coordinamento (attraverso il circuito nervoso endocrino e immunitario) sono la corticotropina e l'interleuchina-l.
Tuttavia, se a un certo livello la complessità delle relazioni multiple tra i vari sistemi biologici è in qualche modo scoraggiante, la relazione tra stress e malattia è anche più difficile da chiarire. Esiste una vasta letteratura clinica che documenta come lo stress cronico possa avere profonde ripercussioni sulla salute. In un certo senso, lo stress ha preso il posto della peste del 20° secolo. Quante volte ci è capitato di sentire il nostro medico di famiglia attribuire l'uno o l'altro dei nostri sintomi inspiegabili allo stress? Nonostante alcuni risultati interessanti, rimane poco chiaro quanto lo stress, e in particolare lo stress cronico, renda un individuo più vulnerabile a malattie che, in circostanze normali, sarebbero efficacemente combattute dal sistema immunitario. Assumere che lo stress sia legato alla evoluzione del nostro stato di salute implica che questi effetti dipendano da una sua azione diretta sul sistema immunitario. Tuttavia, molti dei parametri del sistema immunitario studiati in laboratorio non possiedono alcuna relazione conosciuta con lo stress. Non è possibile negare che vi sia qualche tipo di relazione tra stress e salute e che lo stress cronico possa produrre molti effetti negativi sulla salute. Ma, come è stato elegantemente esposto da R.M. Sapolski (1994): "non tutti i bambini possono diventare presidenti, non possiamo porre fine alle guerre semplicemente tenendoci per mano e cantando canzoni popolari, e la fame non sparisce semplicemente immaginando un mondo in cui essa non esiste. Non tutto ciò che è malattia trova la sua causa nello stress, né è in nostro potere guarire noi stessi da tutti i peggiori incubi sulle malattie semplicemente riducendo lo stress e pensando in maniera positiva alla nostra salute. Volesse il cielo che fosse così. Vergogna per coloro che pretendono di sostenere questa visione delle cose".
Relazione tra comportamento e fisiologia
Abbiamo precedentemente affermato che non esiste alcuna ragione specifica perché un parametro di misura dello stress sia meglio di un altro, dando per scontato che quel dato parametro sia stato misurato correttamente. Tuttavia, accade spesso che, qualora si ottengano più misurazioni della risposta allo stress, queste non mostrano di coincidere. Tale discordanza si verifica frequentemente nel caso in cui si saggiano contemporaneamente risposte comportamentali e fisiologiche. Per esempio, quando scimmie scoiattolo, catturate nel loro ambiente naturale e allevate in cattività, vengono esposte singolarmente a un serpente posto sul tetto della loro gabbia in un recipiente di plexiglas, si possono osservare numerosi comportamenti che esprimono paura. La scimmia emette grida di allarme, aumenta la sua attività ed evita il recipiente con il serpente, standone il più lontano possibile. L'animale mostra anche un elevato aumento di cortisolo. Se il serpente viene presentato con le stesse modalità a soggetti che vivono in un gruppo sociale, anziché da soli, i comportamenti esibiti saranno sostanzialmente gli stessi, ma senza che si verifichino modificazioni nell'attività endocrina. Nella condizione di isolamento vi è concordanza tra comportamento e indici fisiologici di stress; nel gruppo sociale questi due parametri sembrano essere indipendenti.
Quando gli indici di stress comportamentali e fisiologici sono in accordo, il problema di interpretare gli eventi è minimo. La difficoltà si manifesta nel caso in cui vi sia una dissociazione o un disaccordo tra due o più delle variabili di risposta tradizionalmente utilizzate. Una discordanza implica che due variabili, che si assume siano legate causalmente, varino secondo direzioni differenti e che, perciò, non risultino collegate. Una vera dissociazione si manifesta nel caso in cui la risposta comportamentale varia mentre le risposte fisiologiche rimangono costanti, e viceversa. In molti casi ciò accade. Per citarne uno, nei piccoli di mammiferi lo stress è stato definito sulla base dell'espressione di un importante comportamento, la vocalizzazione, spesso definita come vocalizzazione di angoscia. Un aumento nella vocalizzazione, in seguito alla separazione dalla madre, è stato correlato linearmente allo stress provato dal soggetto. In una recente serie di studi si è osservato che, dopo la separazione dalla madre, le vocalizzazioni dei piccoli di scimmia erano inversamente correlate alla quantità di cortisolo presente nel sangue. Inizialmente si era ipotizzato che le vocalizzazioni fossero un atto strumentale, in grado di servire come risposta di coping, e quindi capaci di ridurre lo stress. In esperimenti successivi abbiamo manipolato le vocalizzazioni indotte dalla separazione mediante la somministrazione di agonisti o antagonisti degli oppiacei (morfina e naloxone). Sebbene la morfina riducesse le vocalizzazioni di angoscia, mentre il naloxone le aumentasse, la secrezione di cortisolo, così come il livello dei metaboliti della noradrenalina presenti nel liquido cerebrospinale, rimanevano identici in tutti i soggetti separati, indicando una vera e propria dissociazione tra queste grandezze. Si osserva una discordanza anche quando il comportamento altera la risposta fisiologica. L'aggressione indotta da shock ne è un esempio. Se un ratto viene sottoposto a shock elettrico (ponendolo su una griglia elettrificata) in presenza di un altro ratto, entrambe i membri della coppia assumono una postura di combattimento, spesso attaccandosi, e in alcuni casi ferendosi. Tuttavia, negli animali che combattono, la secrezione di adrenocorticotropina è significativamente inferiore a quella dei soggetti sottoposti alla stessa intensità di shock elettrico, ma senza la possibilità di combattere. Il combattimento indotto da shock è stato considerato come un esempio di comportamento di sostituzione. È stato teorizzato che i comportamenti di sostituzione potrebbero avere lo scopo di ridurre le risposte biologiche agli stimoli che le hanno scatenate. La polidipsia è un altro esempio di comportamento di sostituzione che riduce la secrezione di corticosterone provocata da un rinforzo presentato in maniera intermittente e con un lungo intervallo tra una presentazione e l'altra.
Le differenze trovate sperimentalmente tra gli indici di stress comportamentali e fisiologici possono avere una spiegazione teorica. P.J.I Groves e R.F. Thompson (1970) hanno distinto tra stimoli-risposte o sistemi motori con vie neuronali dirette, e sistemi di stato o di attivazione, con connessioni diffuse a livello del sistema nervoso centrale. Quando si induce una risposta motoria, vengono attivati sia il sistema stimolo-risposta sia i sistemi di eccitazione. Tuttavia questi due sistemi sembrano essere indipendenti. Quando si provoca ripetutamente una risposta motoria, l'ampiezza della risposta può diminuire (abituazione) o aumentare (sensibilizzazione). L'abituazione è fasica e avviene a livello del sistema motorio; la sensibilizzazione è tonica e si pensa sia regolata dal sistema che controlla l'attivazione. Poiché i sistemi stimolo-risposta e di eccitazione sembrano essere sensibili a parametri diversi di stimolazione (la frequenza modifica l'abituazione mentre l'intensità è più legata alla sensibilizzazione), è probabile che essi siano dissociati, particolarmente nel caso di una stimolazione ripetuta. Perciò, nei roditori, l'esposizione ripetuta a un ambiente nuovo porta a una diminuzione della risposta motoria, quale una riduzione nel comportamento difreezing ('immobilità'), così come a una riduzione della risposta di defecazione governata dal sistema autonomo. Al contrario, in seguito a stimolazione ripetuta, la secrezione del corticosterone non mostra alcuna riduzione, anche dopo numerose esposizioni.
Un altro esempio di dissociazione è quello in cui indici fisiologici differenti non si accordano tra loro. Un caso molto chiaro si è verificato in una lunga serie di studi in cui soggetti umani venivano sottoposti a un allenamento per il lancio con il paracadute. Tutti gli indici comportamentali e fisiologici dei soggetti erano alterati immediatamente prima e in seguito al primo lancio. Dopo una serie di lanci dalla torre di addestramento, sia le risposte comportamentali, sia la maggior parte di quelle ormonali, non erano più evidenti. Tuttavia, durante tutto il corso dell'addestramento perdurarono l'aumento sia dei livelli di testosterone e noradrenalina nelle urine sia della frequenza del battito cardiaco. Altri esempi di differenze direzionali nelle variazioni degli indici dei sistemi endocrino e nervoso autonomo sono state documentate sia negli animali sia nell'uomo.
Una conclusione che si può trarre da questi dati è che risposte fisiologiche diverse rispondono a parametri psicologici diversi, o vengono regolate diversamente a livello del sistema nervoso centrale. J.P. Henry e P.M. Stephens (1977) hanno anche suggerito che esista una specificità endocrina associata ai differenti moduli comportamentali in situazioni di stress sociale. Essi sostengono l'ipotesi che esistano un sistema dell'amigdala - forse legato alle catecolammine e all'attivazione del sistema nervoso simpatico - e un meccanismo di attivazione dell'ippocampo associati al sistema ipotalamo-ipofisi-surrene. È stato sufficientemente dimostrato che vi sono situazioni in grado di attivare le diverse componenti della risposta allo stress. Come faccia il cervello a organizzare e a determinare quale sistema debba attivarsi, e in quali circostanze, è uno degli interrogativi principali dei ricercatori che studiano la neurobiologia dello stress. Nei casi in cui si può ritenere che il comportamento funzioni come una risposta strumentale o di coping, non ci sono molti problemi interpretativi poiché la risposta comportamentale può essere considerata funzionale e adattativa. In questi casi, l'effetto che ci si aspetta è quello della discordanza dell'andamento dei parametri comportamentali e fisiologici. Quando vi è dissociazione, si preferisce accettare il parametro fisiologico come quello che riflette in maniera più diretta ciò che il cervello sta valutando come stress.
Conclusioni
In questo saggio ci siamo strettamente impegnati ancora una volta nel tentativo di definire lo stress affrontando i problemi ancora aperti che caratterizzano questo campo di studi. La definizione di stress che abbiamo proposto potrebbe non essere più soddisfacente delle altre. È intuitivo pensare che lo stress sia un concetto chiaro per chi lo prova. La nostra definizione implica che, affinché uno stimolo possa indurre il cervello a mettere in moto la gamma di risposte allo stress, è necessario che il sistema nervoso centrale riconosca in esso alcune caratteristiche peculiari (l'incertezza). Tale definizione implica che senza l'elaborazione da parte del cervello e l'attivazione di specifici meccanismi neuronali gli stimoli non hanno alcuna rilevanza per l'organismo. Inoltre, sebbene effetti di stimoli differenti possano essere gli stessi, quali il rilascio di ormoni dalle ghiandole surrenali e l'attivazione simpatica, non è la risposta a definire uno stimolo come stressante. Questa nozione è contraria a quella comunemente accettata. Gli ormoni del sistema nervoso simpatico, così come quelli prodotti dalle ghiandole surrenali, rappresentano le componenti principali dei meccanismi adattativi dell'organismo. Senza questi ormoni, che esplicano funzioni metaboliche vitali, il corpo non può fronteggiare efficacemente gli stimoli fisiologici che gli si possono presentare. A. Munck e collaboratori (1984) hanno ipotizzato che un aumento nella secrezione dei glucocorticoidi abbia l'effetto di sopprimere alcune reazioni acute a molti stimoli fisici. Ciò significa che la secrezione di questi ormoni è necessaria per regolare e ridurre le risposte iniziali, onde evitare che dilaghino e mettano a repentaglio l'organismo. La funzione di questi ormoni, perciò, dovrebbe essere considerata come un meccanismo omeostatico atto a ristabilire l'equilibrio fisiologico. Non è sorprendente, perciò, che tipi diversi di stimoli possano attivare vie neuronali distinte in grado di rispondere a tipi differenti di stimolazione. È giusto affermare che ogni stimolo in grado di attivare i neuroni che producono corticotropina debba essere considerato come stress? Il nostro punto di vista è molto vicino alla teoria presentata originariamente da Mason. Non sono le risposte o gli stimoli a definire lo stress, ma la componente emozionale che viene registrata ed elaborata dal cervello, quando a esso vengono presentate informazioni discrepanti.
Sia negli animali sia nell 'uomo l'incertezza, la mancanza di informazione e l'assenza o la perdita di controllo sono in grado di provocare risposte che corrispondono a uno stato di allarme. Al contrario, la presenza di informazione pertinente, di comportamenti che abbiano effetti positivi, o di azioni che portino a controllare le circostanze in cui ci si viene a trovare, riduce o migliora gli stati di allarme, con una conseguente riduzione delle risposte fisiologiche a essi associate. Le teorie che sono state presentate si discostano dai tradizionali approcci allo stress e pongono molte domande che possono e devono essere risolte, dal momento che le metodologie utilizzate per lo studio del cervello diventano ogni giorno più sofisticate.
Bibliografia citata
BOLLES, R.C. (1972) Reinforcement, expectancy, and learning. Psychol. Rev., 79, 394-404.
ENGEL, B.T. (1985) Stress is a noun! No, a verb! No, an adjective. In Stress and coping, a c. di Field T.M., Maccabe P.M., Schneiderian N., New Jersey, L. Erlbaum Associates, pp. 1-11.
GROVES, P.M., THOMPSON, R.F. (1970) Habituation: a dual-process theory. Psychol. Rev., 77, 419-450.
HARRIS, G.W. (1948) Electrical stimulation of the hypotalamus and the mechanism of neural control of the adenohypophysis. Physiol. Rev., 28, 139-179.
HENNESSY, J.W., LEVINE, S. (1979) Stress, arousal and the pituitary adrenal system: a psychoendocrine model. In Progress in psychobiology and physiological psychology, a c. di Sprague J.M., Epstein A.N., New York, Academic Press, pp. 133-178.
HENRY, J.P., STEPHENS, P.M. (1977) Stress, heaIth and the sodal environment. A sociobiologic approach to medicine. New York, Springer Verlag.
ISAACSON, S.O., JANZON, L., a c. di (1986) Sodal support-health and disease. Stockholm, Almqvist and Wiksell.
LAZARUS, R.S., FOLKMAN, S. (1984) Stress, appraisal, and coping. New Y ork, Springer Verlag.
LEVINE, S. (1993) The influence of social factors on the response to stress. Psychother. Psychosom., 60, 33-38.
LEVINE, S., URSIN, H. (1991) What is stress. In Stress, neurobioIogy and neuroendocrinology, a c. di Brown M.R., Koob G.F., Rivier C., New York, Marcel Dekker, pp. 3-23.
MASON, J.W. (1975) A historical view ofthe stress field. J. Rum. Stress, 1, 6-12, 22-36.
MEANEY, M.J., BHATNAGAR, S., LAROCQUE, S., SHANKS, N., SHARMA, S., SMYTHE, J., VIAU, V., PLOTSKY, P.M. (1993) IndividuaI differences in the hypothalemic-pituitary-adrenal stress responce and the hypothalamic CRF system. In Corticotropin reIeasing factor and cytokines: role in the stress response, a c. di Tache T., Rivier C., New York, New York Academy of Science, 697, 70-85.
MUNcK, A., GUYRE, P.M., HOLBROOK, N.J. (1984) Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr. Rev., 5, 25-44.
SAPOLSKY, R.M., a c. di (1994) Why zebras don't get ulcers: an updated guide to stress-related diseases, and coping. New York, W.H. Freeman and Co.
SAWCHENKO, P.E. (1991) The final common path: issues concerning the organization of centraI mechanismus controlling corticotropin secretion. In Stress: neurobiology and neuroendocrinology, a c. di Brown M.R., Koob G.F., Rivier C., New York, M. Dekker, pp. 55-72.
SCHARRER, E., SCHARRER, B. (1963) Neuroendocrinology. New York, Columbia University Press.
SELYE, H. (1956) The Stress of Life. New York, Mc Graw Hill.
SOKOLOV, E.N. (1960) Neuronal models and the orienting reflex. In The centraI nervous system and behavior, a c. di Brazier M.A.B., New York, Josiah Macy Jr. Foud., pp. 187-276.
SUOMI, S.J. (1987) Genetic and maternal contributions to individuaI differences in Rhesus monkey biobehavioral development. In Perinatal deveIopment: a psychobiological perspective, a c. di Krasnegor N.A., Blass E.M., Hofer M.A., Smotherman W.P., Orlando, Academic Press, pp. 397-419.
URSIN, H. (1988) Expectancy and activation: an attempt to systematize stress theory. In Neurobiological approaches to human disease, a c. di Hellhammer D., Florin I., Wiener H., Toronto, H. Huber, pp. 313-334.
WEISS, J.M. (1972) Influence of psychological variables on stress induced pathology. In Physiology, emotion, and psychosomatic illness, a c. di Porter R., Knight J., Amsterdam, Elsevier, pp. 253-265.