
Specie e speciazione
Specie e speciazione
di Michael J. D. White
sommario: 1. Introduzione: a) concetti di specie e speciazione; b) variazioni delle specie. 2. Il processo di speciazione: a) meccanismi di isolamento riproduttivo; b) il concetto di specie in riferimento alle specie estinte e agli organismi a riproduzione asessuale. 3. Specie correlate. 4. Differenze genetiche fra specie: a) valutazione delle differenze genetiche; b) la divergenza evolutiva. 5. Fattori che influenzano le modalità di speciazione. 6. Speciazione cromosomica: a) zone parapatriche; b) speciazione invasiva e speciazione stasipatrica; c) speciazione simpatrica. 7. Specie e speciazione nelle piante: poliploidia. 8. Il problema della rettangolarità nella speciazione. □ Bibliografia.
1. Introduzione
a) Concetti di specie e speciazione
L'uomo primitivo creò, senza rendersene conto, il concetto di specie quando ravvisò l'esistenza in natura di differenti tipi di animali e piante, dando nomi a quelli che erano per lui di interesse, in quanto rappresentavano cibo o un nemico o semplicemente perché erano appariscenti.
Fino all'epoca di Linneo non furono fatti molti progressi, ma da quest'epoca in poi fu generalmente accettata l'idea che il mondo vivente esiste sotto forma di un vasto numero di specie che è utile raggruppare in generi, cosicché ogni specie possa essere designata inequivocabilmente da un nome latino binomiale (per es.: Homo sapiens, Pinus pinea). La scienza della tassonomia si interessa della delimitazione, della descrizione e della denominazione delle specie, mentre la sistematica si occupa del campo correlato e parzialmente sovrapposto della loro classificazione (v. sistematica biologica).
Fino alla comparsa, nel 1859, della fondamentale opera di Charles Darwin, L'origine delle specie, l'opinione generale era che ogni specie fosse stata creata indipendentemente dalle altre e che a tale fatto fosse dovuta la loro diversità. Dopo Darwin, però, gli scienziati si convinsero che le specie si erano originate mediante un processo evolutivo, cioè mediante modificazioni genetiche in popolazioni naturali di organismi esistiti in tempi passati. In linea molto generale si pensa che queste modificazioni siano state analoghe a quelle implicate nella produzione, in tempi storici, di nuove varietà o ibridi di piante e animali domestici da parte dell'uomo.
Si sono avute in passato numerose controversie a proposito della natura delle specie e dei dettagli del processo di ‛speciazione' mediante il quale hanno origine nuove specie nel corso dell'evoluzione. I primi concetti di specie erano essenzialmente morfologici e gli studiosi di tassonomia descrivevano le specie unicamente in base a differenze strutturali. Durante gli anni trenta venne accettato da tutti il concetto biologico di specie, in base al quale le specie costituite da organismi che si riproducono sessualmente erano definite come gruppi di individui che di norma si incrociano regolarmente, salvo che ciò non sia reso impossibile da impedimenti estrinseci come, per esempio, una separazione geografica. Questa definizione non esclude che specie distinte possano, occasionalmente, accoppiarsi e dare luogo a ibridi interspecifici: il punto fondamentale sta proprio nell'espressione ‛incrocio normale e regolare'. Questo fa degli organismi stessi i veri arbitri della differenziazione delle specie. Là dove il biologo è in grado di osservare in natura il suo materiale, può ottenere da questo le informazioni necessarie e assegnare nomi appropriati. In molti casi, tuttavia, chi si occupa di tassonomia studia in un museo materiale morto, raccolto da qualcun altro in una remota parte del mondo molti anni prima. In questo caso, egli può usare come criterio solo il suo giudizio personale per stabilire quali individui potrebbero formare un certo ‛gruppo riproduttivo A' e quali potrebbero costituire un ‛gruppo riproduttivo B'.
Il numero di specie attualmente viventi nel mondo non può assolutamente essere stabilito con sicurezza. Il numero di specie animali descritte si aggira attorno a 1.400.000, ma si sa che esistono molte specie non ancora descritte (per es. ne esistono alcuni esemplari nelle collezioni dei musei). Possiamo, inoltre, solo fare delle ipotesi sul numero di specie esistenti nelle foreste pluviali tropicali del mondo, nelle poco conosciute regioni dell'Asia centrale e meridionale e nelle profondità del mare, in quanto non sono mai state raccolte ed esaminate da scienziati. Vi sono problemi anche per quanto riguarda animali molto piccoli (Acari, Protozoi, e alcuni altri gruppi), dato che le specie sono molto numerose, ma pochi gli specialisti. In linea di massima è molto probabile che il numero di specie reali e distinte di animali sia dell'ordine di 10 milioni. Il numero di specie vegetali è certamente inferiore a un milione. Non può essere valutato con alcuna certezza il numero delle specie di tutti gli organismi vissuti nel passato e ora estinti; è tuttavia improbabile che il numero di specie esistite in una qualsiasi epoca abbia mai superato il numero di specie viventi all'inizio del nostro secolo. È certo però, che la velocità con la quale stanno attualmente scomparendo alcune specie è molto maggiore che in passato e che ciò è dovuto allo sfruttamento del pianeta da parte dell'uomo, che ha distrutto l'habitat di tali specie. Il numero di specie della maggior parte dei gruppi di organismi è quindi diminuito nelle ultime decine di anni e dobbiamo attenderci un ulteriore brusco calo nel prossimo secolo.
b) Variazioni delle specie
Gli individui di una specie non sono del tutto identici, in altre parole essi presentano una certa variabilità, che può essere sia continua sia discontinua. Variazioni discontinue si osservano quando vi sono tipi di individui che presentano marcate differenze senza che esistano soggetti con caratteristiche intermedie. Cosi in alcuni tipi comuni di chiocciola europea (Cepaea nemoralis, C. hortensis e Theba pisana) vi sono individui con differenti tipi di strie sulla conchiglia (una stria centrale, tre strie, cinque strie), oppure con conchiglie di colori differenti: gialle, marroni, rosa, ecc. Anche tra gli uomini è noto che esistono diversi tipi di variazioni genetiche non immediatamente evidenti, come ad esempio gli antigeni dei gruppi sanguigni.
La variazione continua non solo è sempre presente in relazione a caratteristiche metriche come la taglia, il peso, le proporzioni del corpo, ecc., ma si manifesta anche a proposito di molte caratteristiche biochimiche che regolano importanti processi fisiologici.
Sia la variazione continua sia quella discontinua si manifestano in due maniere differenti. Esse possono essere presenti in singole popolazioni di una determinata specie, quelle cioè che occupano una particolare zona. Tutte le popolazioni di una specie che si riproduce sessualmente, pertanto, sono dette ‛polimorfe'. Esiste tuttavia anche il fenomeno della variazione geografica. Ogni specie presente in una vasta zona geografica mostra alcune differenze correlate con la distanza. Questo fenomeno è noto a tutti se facciamo riferimento alla specie umana; infatti gli individui nativi dell'Africa sono per molti aspetti diversi da quelli dell'Asia orientale o dell'Europa centrale, e, in scala più piccola, ci sono differenze morfologiche, anche se minori, fra le popolazioni umane della Scandinavia e quelle della regione alpina, della Sicilia o di altre regioni d'Europa. Quindi nella maggior parte delle specie animali e vegetali possiamo individuare l'esistenza di numerose ‛razze geografiche' alle quali in passato si tendeva ad attribuire nomi latini; così nel caso del comune topo delle case, Mus musculus, gli zoologi distinguevano Mus musculus musculus, Mus musculus domesticus, Mus musculus brevirostris e altri, che venivano considerati ‛sottospecie'. Le sottospecie sono tuttavia essenzialmente soggettive e non possono essere definite nella stessa maniera oggettiva delle specie; pertanto attualmente gli studiosi di tassonomia sono generalmente contrari al riconoscimento di sottospecie formali designate con nomi latini. È comunque normale nel linguaggio comune usare per le razze geografiche denominazioni come ‛razza settentrionale', ‛razza montana', ‛razza mediterranea', ecc.
2. Il processo di speciazione
a) Meccanismi di isolamento riproduttivo
La speciazione è essenzialmente il processo in base al quale le differenze genetiche fra specie si determinano e si accumulano nel corso dell'evoluzione. Poiché si tratta di un processo graduale, dobbiamo aspettarci di trovare in natura un numero piuttosto elevato di casi di specie in statu nascendi, e in tali casi può essere molto difficile decidere se ci troviamo di fronte a una o due specie. Situazioni di tal genere hanno importanza particolare per i biologi che analizzano il meccanismo della speciazione.
Comunque, poiché la speciazione richiede di solito millenni per il suo completamento, è evidente che non può essere studiata sperimentalmente nella sua interezza. Possiamo tutt'al più compiere esperimenti su piccole parti dell'intero processo studiando, ad esempio, gli effetti della selezione naturale o artificiale su alcuni tipi di modificazione che sappiamo accompagnare la speciazione.
Il meccanismo di evoluzione biologica dovuto alla selezione naturale fu compreso nelle sue linee generali diversi decenni prima che il meccanismo di speciazione cominciasse a essere analizzato in dettaglio. Cosi scrisse William Bateson (v., 1922): ‟Anche se i suoi meccanismi sono abbastanza oscuri, l'evoluzione è un processo piuttosto evidente. Ma quella parte specifica ed essenziale della teoria dell'evoluzione che riguarda l'origine e la natura delle specie rimane completamente avvolta nel mistero". A questo R.A. Fisher (v., 1930) aggiunse: ‟Gli stretti legami genetici che uniscono le specie in un singolo corpo mettono in rilievo il problema della loro divisione, problema che implica complessità simili a quelle che si riscontrano in una discussione sulla divisione dei corpi celesti, poiché il tentativo di tracciare il corso degli eventi attraverso una serie di stadi di stabilità sembra richiedere in entrambi i casi una conoscenza più dettagliata di quella che può fornire lo studio degli stadi stabili stessi".
Un importante passo in avanti fu fatto da Dobzhansky (v., 1937), il quale individuò l'esistenza di ‛meccanismi di isolamento riproduttivo' che agiscono in modo da prevenire un'efficace ibridazione tra specie correlate che, altrimenti, arriverebbero alla fusione. Questi meccanismi intrinseci di isolamento, che hanno una base genetica, sono ovviamente differenti dall'isolamento geografico che impedisce l'incrocio tra specie le cui aree di distribuzione sono completamente separate e in natura non si sovrappongono.
L'ibridazione tra specie differenti può essere completamente impedita oppure può verificarsi, ma essere biologicamente inefficace. I meccanismi riproduttivi di isolamento possono pertanto distinguersi in ‛prezigotici', quando impediscono la formazione dell'ibrido, e in ‛postzigotici', quando riducono la vitalità e la fecondità degli ibridi. La situazione, in linea di principio, è la stessa per animali e piante, sebbene i dettagli siano ovviamente differenti. I meccanismi prezigotici comprendono l'‛isolamento ecologico' (le popolazioni occupano differenti habitat anche se si trovano nella stessa zona geografica), l'‛isolamento temporale' (nelle due popolazioni l'accoppiamento o la fioritura avvengono in stagioni differenti o in momenti differenti della giornata), l'‛isolamento etologico' (il corteggiamento o il comportamento precedente l'accoppiamento presso le due popolazioni sono differenti, cosicché l'attrazione sessuale tra esse è ridotta o assente), l'‛isolamento meccanico' (i genitali delle due popolazioni - o la struttura dei fiori nel caso delle piante - non sono morfologicamente compatibili, cosicché il passaggio di sperma o di polline è impedito) e infine l'‛isolamento gametico' (i gameti delle due popolazioni non si attraggono, oppure lo sperma o i tubuli pollinici dell'uno non sono vitali nei tratti genitali o negli stigmi dell'altro).
I meccanismi di isolamento postzigotico includono la ‛non vitalità dell'ibrido' (l'ibrido non sopravvive oltre un certo stadio, precedente la maturità sessuale), la ‛sterilità dell'ibrido' (le prime generazioni degli ibridi sono sterili, sia quando si accoppiano tra di loro sia quando si reincrociano con una delle generazioni parentali) e l'‛esaurimento dell'ibrido' (sebbene gli ibridi della prima generazione siano fertili, le successive generazioni presentano una ridotta vitalità o fecondità).
È evidente che tutti questi meccanismi di isolamento tendono a prevenire il flusso di materiale genetico da una specie all'altra. Ogni specie biologica possiede quindi un ‛pool di geni' indipendente. In pratica quasi tutte le specie completamente differenziate sembrano essere mantenute separate in natura da una combinazione di numerosi meccanismi di isolamento genetico. Se questi sono esclusivamente postzigotici, vi sarà uno spreco biologico di gameti che viene però evitato se esistono efficaci meccanismi di isolamento premeiotico.
L'acquisizione di meccanismi di isolamento riproduttivo è perciò un aspetto estremamente importante della speciazione, anzi secondo alcuni autori è in pratica il solo aspetto che deve essere preso in considerazione. La speciazione però implica di solito l'adattamento a un nuovo habitat, a una nuova fonte di cibo o a un nuovo tipo di vita, e tali cambiamenti sono ovviamente troppo importanti per poter essere trascurati.
Dopo che fu riconosciuta l'importanza dei meccanismi dell'isolamento riproduttivo nella speciazione, Mayr (v., 1942) sostenne che l'acquisizione di questi meccanismi era possibile solo in condizioni di isolamento geografico, cioè che l'estrinseco isolamento spaziale di una popolazione dal corpo principale della specie doveva necessariamente precedere l'acquisizione di nuovi meccanismi intrinseci di isolamento biologico aventi base genetica. Questo modello di ‛speciazione allopatrica' negava che la speciazione potesse avvenire anche ‛simpatricamente', cioè attraverso l'adattamento a due diversi habitat o fonti di nutrimento, presenti nella stessa area geografica.
L'opinione di Mayr sulla componente geografica nel processo di speciazione dominò per circa venticinque anni; attorno al 1967 fu però ripresa in esame e ora sembra che esistano diverse modalità di speciazione che differiscono sia per la componente geografica sia per la natura dei cambiamenti genetici intervenuti (v. White, 1978).
b) Il concetto di specie in riferimento alle specie estinte e agli organismi a riproduzione asessuale
Vi sono due situazioni nelle quali il concetto di specie biologica basato sull'isolamento genetico non può essere applicato. Una è il caso delle specie estinte, le specie della paleontologia. Queste specie fossili sono quindi ‛morfospecie', definite unicamente sulla base di differenze strutturali. Nel caso di popolazioni fossili contemporanee, le differenze morfologiche tra di esse possono essere valutate statisticamente e confrontate con le specie dello stesso gruppo ancora viventi (se ne esistono), definite sia secondo il criterio biologico sia su basi morfologiche. I paleontologi distinguono anche specie consecutive appartenenti alla stessa linea evolutiva e assegnano loro dei nomi; nella maggioranza dei casi, però, questa è una procedura arbitraria, perché l'evoluzione di queste specie nel corso del tempo (‛anagenesi') è un processo continuo e non vi è in genere alcun criterio obiettivo per decidere a quale punto una specie sfumi in un'altra, sebbene in qualche caso un'improvvisa trasformazione morfologica venga documentata da resti fossili.
L'altra situazione in cui il concetto di specie biologica, basato sulla riproduzione sessuale, è inapplicabile è quella degli organismi che si riproducono esclusivamente per riproduzione asessuale (riproduzione agamica, partenogenesi v. riproduzione). In pratica, tuttavia, la difficoltà causata da questa situazione non è eccessiva; gli studiosi di tassonomia hanno infatti attribuito nomi binomiali specifici a organismi in grado di riprodursi solo per partenogenesi o agamicamente e tali nomi sono stati accettati dai biologi e dai genetisti che li studiano. Una sorta di giustificazione logica permette di parlare di ‛specie' che si riproduce partenogeneticamente o agamicamente, quando si può dimostrare che la popolazione in questione ha tratto origine da organismi ancestrali che si riproducevano sessualmente. Intervengono serie difficoltà, però, nel caso di popolazioni, o gruppi di popolazioni, che si riproducono asessualmente e che si pensa abbiano avuto origini multiple. È probabile che la classificazione di organismi partenogenetici e asessuali causerà difficoltà sempre crescenti con l'approfondirsi della conoscenza di queste forme viventi.
3. Specie correlate
L'entità delle differenze morfologiche tra specie vicine è estremamente varia. Quando le differenze morfologiche sono molto piccole (e in alcuni casi sono addirittura irrilevabili), parliamo di specie correlate (o di specie criptiche). Molte ‛specie' della tassonomia classica si sono rivelate negli ultimi anni come ‛complessi' che comprendono diverse specie biologiche completamente differenti, mantenute separate tra loro da meccanismi di isolamento genetico, nonostante che sia quasi impossibile distinguerle una dall'altra mediante metodi tradizionali.
Un classico esempio di specie correlate è rappresentato da Drosophila pseudobscura e D. persimilis nel Nordamerica occidentale. Le femmine di queste due specie sono morfologicamente indistinguibili, mentre vi è una piccola differenza negli organi genitali dei maschi. Una sicura attribuzione all'una o all'altra specie è possibile con l'esame citologico dei cromosomi. In natura i loro areali si sovrappongono ampiamente, ma D. pseudobscura tende a localizzarsi preferenzialmente in ambienti più caldi e secchi. Gli ibridi maschi di prima generazione ottenuti mediante incroci sperimentali sono sterili, ma gli ibridi di sesso femminile possono essere reincrociati con le specie parentali; la vitalità e fecondità degli ibridi di reincrocio sono però ridotte. In condizioni naturali l'isolamento etologico sembra essere molto forte: Dobzhansky esaminò, nell'arco di 35 anni, circa 27.000 individui e trovò solo un caso certo di ibrido naturale. Nonostante questa ibridazione naturale occasionale, sembra che non ci sia trasferimento di materiale genetico da una specie all'altra (sicuramente a causa delle ridotte vitalità e fecondità degli ibridi). I loro pools di geni sono dunque completamente separati.
Queste mosche non sono di alcun interesse economico per l'uomo. In un certo numero di casi, tuttavia, si è potuto dimostrare che organismi di grande importanza per l'uomo in quanto vettori di malattie o insetti nocivi appartengono a complessi di specie correlate, e che altri membri dello stesso complesso sono invece innocui o hanno importanza molto minore. Se per tali casi ci basassimo sulla tassonomia classica rischieremmo di impiegare enormi somme di denaro per controllare specie che sono relativamente o niente affatto importanti sia per la salute umana che per il benessere economico. Un esempio di tal genere si ebbe nel 1975, quando si pensò che una mosca della frutta, Dacus dorsalis, un insetto asiatico nocivo per circa 250 specie di frutti, fosse stato introdotto nell'Australia settentrionale. Vi fu un notevole allarme finché si scoprì che la popolazione australiana era costituita da una specie correlata e fino allora sconosciuta, distinguibile da quella asiatica per la dimensione e la forma di un cromosoma e che viveva solo su due specie di piante spontanee senza importanza economica.
In Africa le zanzare del gruppo Anopheles gambiae sono estremamente importanti come vettori della malaria. Si è attualmente scoperto, tuttavia, che si tratta di un complesso di sei specie correlate, alcune delle quali pungono l'uomo abitualmente, altre praticamente mai e pertanto non hanno importanza come vettori di tale malattia. Queste specie possono essere distinte in base ai loro cromosomi, mentre le differenze morfologiche sono minime (v. Coluzzi e altri, 1979). Una situazione analoga si verifica nel caso del dittero Simulium damnosum, responsabile, in un'ampia zona dell'Africa centrale, della trasmissione della filariosi da oncocerca (cecità fluviale). Si tratta di un complesso di undici specie di cui solo alcune pungono l'uomo abitualmente: anch'esse sono distinguibili su basi cromosomiche, mentre le differenze morfologiche sono impercettibili.
Nei due casi citati l'ibridazione fra le specie correlate è scarsa o nulla. In altri casi, invece, vi può essere un elevato tasso di ibridazione, perché mancano i meccanismi di isolamento precedenti all'accoppiamento, ma ciò nonostante le specie mantengono i loro pool genetici separati. Così Bullini e altri (v., 1978) hanno dimostrato che il ben noto parassita intestinale dei cavalli Parascaris equorum (precedentemente noto come Ascaris megalocephala) è in realtà rappresentato da una coppia di specie: Parascaris equorum vero, con due coppie di cromosomi nel genoma, e Parascaris univalens, con una sola coppia. Uno studio elettroforetico dei due sistemi enzimatici ha messo in luce che le differenze genetiche tra le due specie sono notevoli, mentre le differenze morfologiche sono di scarsa entità. Di 2.238 individui studiati, 2.092 erano Parascaris univalens, 127 P. equorum e 19 ibridi naturali. Nonostante l'alta frequenza di ibridazione, lo studio elettroforetico ha dimostrato che le due specie hanno pool genetici ben diversi e che non vi è flusso genetico, forse a causa della sterilità degli ibridi.
4. Differenze genetiche fra specie
a) Valutazione delle differenze genetiche
Le differenze genetiche fra specie correlate sono di diversi tipi. Possono esserci, come già detto, marcate differenze (di numero, dimensione o forma) tra i cromosomi; possono anche esserci differenze nella struttura molecolare dei cromosomi, riguardanti in particolare la quantità e distribuzione delle sequenze nucleotidiche altamente ripetitive (DNA satelliti) che costituiscono l'eterocromatina visibile al microscopio ottico e che non sono organizzate come geni. Tra alcune specie si sono trovate differenze in un'altra frazione di DNA, e precisamente quella del DNA moderatamente ripetitivo. In conseguenza di tali mutamenti evolutivi nella quantità del DNA altamente e moderatamente ripetitivo, il quantitativo totale di DNA nel genoma di specie correlate (il cosiddetto valore C) può essere molto differente (v. tab. I).

Ci sono infine differenze alleliche nei loci dei geni strutturali responsabili della sintesi di un enzima o di altre molecole proteiche modificate. I vari tipi di differenze genetiche fra specie correlate di organismi devono essere misurati mediante parametri differenti e non esiste alcun modo soddisfacente di combinare tali parametri, che sono incommensurabili, in modo tale da fornire una misura generale della divergenza genetica.
Molti autori, usando metodi elettroforetici, hanno concentrato la loro attenzione esclusivamente sull'identificazione di differenze alleliche fra popolazioni, razze o specie, trascurando totalmente gli altri tipi di differenze genetiche che sono difficilmente quantificabili. Secondo la metodica di Nei (v., 1972) vengono usati due parametri. Il primo è l'‛identità genetica' (I), che è la valutazione della frazione dei loci genici rappresentati da alleli identici nelle due popolazioni; questo parametro varia, naturalmente, da uno (tutti gli alleli uguali) a zero (nessun allele identico). Il secondo è la ‛distanza genetica' (D), che è la valutazione del numero di mutamenti allelici che si sono avuti durante la divergenza evolutiva delle due popolazioni; essa varia da zero (nessun cambiamento, I=1) a infinito. D può essere maggiore di 1 poiché ogni locus genico, col progredire dell'evoluzione, può subire più di una sostituzione allelica. Il valore di D per la ‛distanza genetica' fra Parascaris equorum e P. univalens è 1,939. La relazione tra D e I è espressa dalla formula D=−loge I.
Ovviamente i valori di I e D saranno calcolati in base a dati su quanti più loci genici differenti possibile. Questo è necessario, perché vi sono alcuni loci altamente variabili la cui velocità di evoluzione è molto alta, mentre altri si evolvono molto più lentamente o sono del tutto stabili. Così Zimmerman e altri (v., 1978) hanno riscontrato che la distanza genetica fra popolazioni geografiche dei roditori Peromyscus boylii e P. pectoralis era 0,066 se calcolata solo in base ai loci che evolvono velocemente, mentre era solamente 0,004 se calcolata in base ai loci che evolvono più lentamente (valore medio D=0,03).
b) La divergenza evolutiva
Nei (v., 1975) ha suggerito la possibilità di calcolare in anni il tempo trascorso dalla divergenza evolutiva di due popolazioni mediante la formula: t=5×106 D. Si tratta ovviamente di una stima molto grossolana; Sarich (v., 1977) ha proposto, per i loci che evolvono rapidamente, di utilizzare la seguente formula: t=2,4×106 D. Sulla base di questi valori di t, la divergenza tra le specie del gruppo di Peromyscus boylii si sarebbe avuta 811.200 anni fa (se consideriamo i loci che evolvono rapidamente) o 9.690.000 anni fa (considerando quelli che evolvono lentamente). Il valore medio (5×106×0,334) sarebbe 1.670.000 anni fa. Poiché il valore di D precedentemente citato per Parascaris equorum e P. univalens (1,939) è un valore medio, basato sia sui loci che evolvono rapidamente sia su quelli che evolvono lentamente, possiamo, in linea di massima, accettare il metodo di stima di Nei e supporre che la divergenza evolutiva di queste specie sia avvenuta circa dieci milioni di anni fa.
Sarebbe comodo poter affermare che un particolare valore di D corrisponde al completamento del processo di speciazione; se tutti gli eventi connessi con la speciazione avvenissero con gli stessi meccanismi e nella stessa maniera ciò sarebbe possibile, ma certamente non è così. Nella tab. Il sono riuniti i dati sul gruppo di Drosophila willistoni e su due generi di Roditori. In questi tre casi il completamento del processo di speciazione sembra avvenire quando D raggiunge un valore compreso fra 0,3 e 0,4.

Tranne il caso particolare della speciazione per poliploidia (moltiplicazione del corredo cromosomico nel nucleo), la speciazione non è un processo istantaneo, ma si estende per un periodo di tempo misurabile in millenni. Bisogna pertanto distinguere fra gli eventi iniziali che causano il processo di speciazione e i cambiamenti successivi che avvengono col procedere della divergenza.
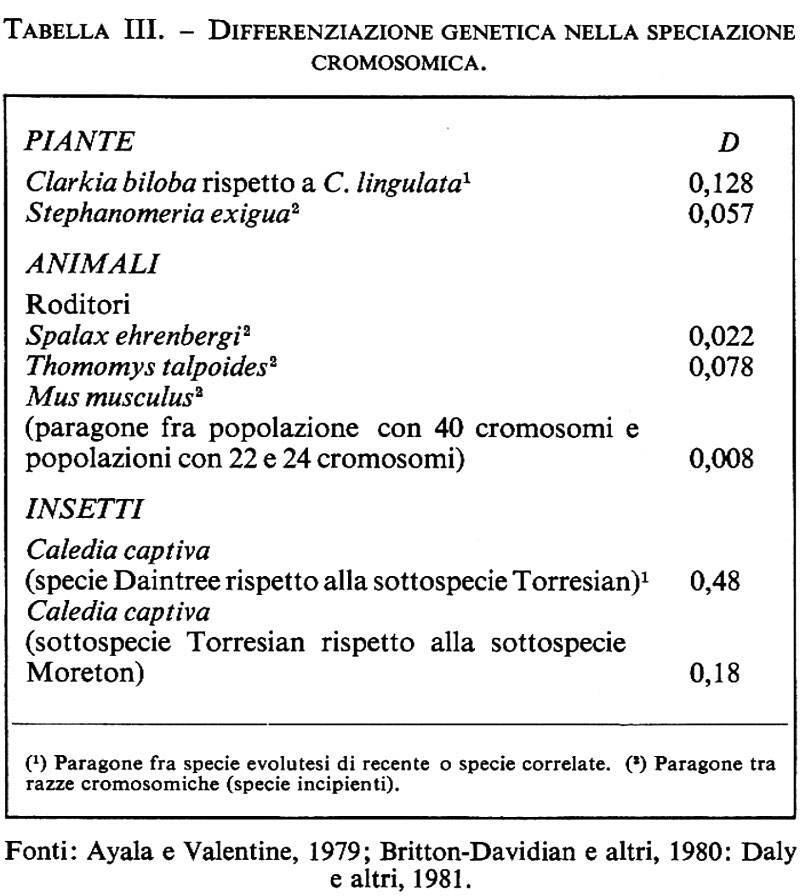
Per quanto riguarda gli animali (ma la cosa, in linea di massima, è estensibile alle specie vegetali) è chiaro che la maggior parte delle coppie di specie strettamente correlate presentano differenze che riguardano i cariotipi, in altre parole le modificazioni strutturali dei loro cromosomi risalgono al momento della loro separazione evolutiva. Nel genere Drosophila e in alcuni altri ditteri (Simulidae, Culicidae, ecc.) ci sono alcuni esempi di specie strettamente correlate, che sappiamo essere separate in quanto hanno caratteristiche morfologiche, biochimiche ed etologiche diverse, che sembrano avere lo stesso cariotipo. Tali specie sono dette ‛omosequenziali' (v. Wasserman, 1963; v. Carson e altri, 1970). La loro esistenza dimostra chiaramente che il riassestamento cromosomico non è una condizione sine qua non per la speciazione. Nondimeno tali specie rappresentano una bassissima percentuale del totale, e la situazione, molto più comune, di specie che differiscono nel cariotipo suggerisce chiaramente che i riordinamenti cromosomici sono spesso connessi con eventi di speciazione e si può quindi presumere che ne siano causa. Tale punto di vista è sostenuto dal fatto che capita frequentemente di trovare specie che consistono di un certo numero di ‛razze cromosomiche', e che differiscono nel cariotipo. In molti casi tali razze sembrano essere specie incipienti che hanno attraversato lo stadio di divergenza iniziale di cui abbiamo parlato, ma non hanno ancora subito il graduale processo di trasformazione genetica che porta al completamento del processo di speciazione. Nella tab. III diamo alcuni dati sul grado di differenziazione genetica (parametro D, secondo Nei) nel caso di razze cromosomiche o specie incipienti; nella maggioranza di tali casi è possibile vedere che, sebbene le differenze cromosomiche siano notevoli, la distanza genetica è molto piccola (minore di 0,2 e spesso anche di 0,1).
5. Fattori che influenzano le modalità di speciazione
Due fattori che hanno sicuramente avuto un ruolo preponderante nel determinare le modalità di speciazione caratteristiche di particolari gruppi di organismi sono il grado di specializzazione ecologica e il grado di ‛vagilità', termine tecnico col quale si indica il grado di mobilità mostrato da un organismo durante il corso del suo ciclo vitale; la vagilità può essere definita come la distanza media, in linea d'aria, fra il punto della Terra in cui un individuo viene al mondo (mediante fecondazione) e il punto in cui fa nascere la sua prole. Nel caso di organismi sedentari può trattarsi di pochi metri, mentre nel caso di specie marine o di quelle disperse dal vento può trattarsi di centinaia o addirittura di migliaia di chilometri.
La ‛specializzazione ecologica' è il grado di limitazione a cui sono soggette le specie a seconda dei particolari tipi di habitat. La sua forma più evidente è la ‛specializzazione trofica', che rappresenta il vincolo che lega una certa specie a un determinato tipo di cibo. Tra i casi più appariscenti vi sono quello degli insetti che possono vivere solamente su di uno specifico tipo di pianta-cibo e quello dei parassiti che infestano solo particolari specie di organismi.
6. Speciazione cromosomica
a) Zone parapatriche
Il fatto che le differenze cariotipiche esistenti tra specie correlate siano dovute, di solito, ad alcuni riassestamenti strutturali dei loro cromosomi ha naturalmente causato lo sviluppo di diverse ipotesi sulla ‛speciazione cromosomica'. Una caratteristica generale della speciazione cromosomica è quella di determinare una situazione tale per cui due razze cromosomicamente monomorfiche o due specie incipienti occupano areali del tutto separati tranne che per una limitata zona di sovrapposizione nella quale si riscontrano degli ibridi. Queste zone di sovrapposizione sono dette ‛parapatriche' e alcuni autorevoli scienziati hanno individuato una particolare categoria di ‛speciazione parapatrica', termine che tuttavia non sarà usato in questa sede poiché sembra riferito semplicemente a uno stato di fatto geografico che può aver avuto origine in vari modi differenti.
Zone parapatriche sono state descritte in particolar modo per organismi a ridotta vagilità, come cavallette senza ali, scarabei, isopodi, lucertole e piccoli roditori. Queste zone per organismi con vagilità molto limitata possono essere ridotte addirittura a poche centinaia di metri. Sembra che di solito nella zona di sovrapposizione avvenga un libero interincrocio. Si ritiene che la causa della scarsa estensione di queste zone sia identificabile nella combinazione di due fattori: la vagilità estremamente limitata di questi organismi e l'azione della forte selezione naturale contro gli ibridi (eterozigoti cromosomici) causata dalla loro minore fecondità o vitalità.
È stato a lungo discusso se le zone parapatriche siano da considerarsi primarie o secondarie, cioè se siano il risultato dell'espansione di popolazioni precedentemente allopatriche che sono così giunte in contatto. Secondo l'interpretazione qui adottata, la maggior parte delle zone parapatriche, e in particolare quelle in cui sono coinvolte razze cromosomiche, sono primarie; alcune, tuttavia, sono state riconosciute sicuramente come secondarie.
b) Speciazione invasiva e speciazione stasipatrica
Dal punto di vista geografico sembra vi siano due principali modelli di speciazione cromosomica: essi possono essere indicati come ‛invasivo' e ‛stasipatrico'. Nel caso della speciazione invasiva, una popolazione periferica di una specie va incontro a un riassestamento cromosomico che può essere un'inversione, una traslocazione o una fusione. Se questo fenomeno non viene eliminato per selezione naturale, dopo diverse generazioni si potrà trovare una piccola colonia della specie monomorfica rispetto al nuovo tipo di cromosoma (cioè ogni individuo è omozigote per esso). Se il nuovo omozigote è adattivamente superiore, potrà espandersi in un territorio precedentemente non occupato dalla specie. Se questo fenomeno si ripete parecchie volte di seguito, potremo avere un'intera catena di razze cromosomiche o specie incipienti che si estendono in modo lineare. Sembra che questo sia stato il caso dei ratti spinosi venezuelani della superspecie Proechimys guairae (v. Reig e altri, 1980), una catena di sei popolazioni (specie correlate o razze cromosomiche) con numeri cromosomici 2n=42, 44, 46, 48, 50, 62, distribuite in una sorta di ferro di cavallo nelle terre basse attorno all'estremità settentrionale della Cordigliera di Merida. Non è chiaro se la razza con 62 cromosomi sia la più antica (nel qual caso i cariotipi delle altre sarebbero derivati da una serie di fusioni cromosomiche), oppure se la popolazione ancestrale sia quella con 42 cromosomi (e in tal caso si sarebbe avuta una scissione cromosomica). È anche possibile che la popolazione ancestrale avesse 50 o 52 cromosomi e che l'evoluzione abbia progredito nei due sensi, cioè sia geograficamente, sia rispetto al numero cromosomico. È presumibile che le varie popolazioni di Proechimys si incontrino in strette zone di sovrapposizione che però non sono ancora state studiate, e non si sa con sicurezza se vi siano ibridi naturali in tali zone. Come in altri casi di speciazione cromosomica, la distanza genetica (D di Nei) fra le popolazioni del complesso di specie di Proechimys guairae è molto piccola e ciò fa supporre che queste popolazioni si siano allontanate dalla popolazione ancestrale tra i 50.000 e i 200.000 anni fa.
Il modello stasipatrico della speciazione cromosomica (v. White, 1968 e 1978) differisce dal modello invasivo solamente in quanto l'origine del riassestamento cromosomico è localizzata nel centro invece che nella periferia e la nuova razza cromosomica (potenzialmente una specie incipiente) si sviluppa entro il territorio occupato dalla forma ancestrale invece che all'esterno. Poiché i confini delle popolazioni di una specie generalmente cambiano col tempo, può essere difficoltoso distinguere i casi in cui la speciazione cromosomica è stata invasiva da quelli in cui ha seguito il modello stasipatrico.
In un primo momento si pensò che il modello stasipatrico fosse il principale meccanismo di speciazione, ma sicuramente non il solo, per le cavallette australiane senza ali della sottofamiglia delle Morabinae, insetti a bassissima vagilità. Sono state studiate molte zone parapatriche di ibridazione (‛zone di tensione' nella terminologia di Key: v., 1974) per il genere Vandiemenella; esse sono ampie circa 200-300 metri.
Altri esempi di evoluzione cromosomica che può essere invasiva o stasipatrica, o forse anche una combinazione fra le due, sono le dieci ‛razze' di insetto stecco Didymuria violescens descritte da Craddock (v., 1970 e 1975) e le varie specie di isopodi del genere Jaera viventi sulla costa atlantica dell'Europa (v. Bocquet, 1953; v. Lécher, 1967 e 1968). In Didymuria, che si nutre di foglie di eucalipto nelle grandi foreste dell'Australia orientale, il numero primitivo di cromosomi nel maschio era certamente 2n♂=39, ma in seguito a fusioni cromosomiche si sono originate razze o specie incipienti con 37, 35, 32, 31, 30, 28 e 26 cromosomi. Le razze con numero dispari di cromosomi (39, 37, 35, 31) hanno nel maschio un assetto cromosomico XO - in cui O indica l'assenza del cromosoma Y - mentre i maschi con numero pari di cromosomi (32, 30, 28, 26) hanno un assetto XY Il caso di Jaera sembra aver a che fare con l'adattamento di varie specie a differenti zone di costa, da quelle quasi terrestri a quelle più spiccatamente marine. Per molte coppie di specie è stata descritta l'ibridazione nelle strette zone di contatto o in zone più estese.
Alcuni evoluzionisti hanno trovato difficoltà ad accettare l'idea che la speciazione avvenga come conseguenza di un riassestamento cromosomico che diminuisce la fecondità o la vitalità (principalmente la prima) degli eterozigoti, perché pensano che tale riassestamento avrebbe avuto possibilità minime di sopravvivere alla selezione naturale. Un processo di questo tipo deve essere sicuramente considerato eccezionale in quanto la maggioranza dei riassestamenti cromosomici deleteri nell'eterozigote saranno eliminati dalla selezione naturale - ed è concepibile solamente in popolazioni di specie altamente frammentate in piccolissime colonie locali: in tali situazioni è comunque senz'altro possibile, specialmente se il nuovo riassestamento favorisce la ‛spinta meiotica' (per esempio se questo cromosoma modificato passa più facilmente, durante l'ovogenesi, nel nucleo dell'uovo piuttosto che in quello di un corpuscolo polare). Non è ancora stato provato con certezza che la spinta meiotica svolga un ruolo nella speciazione cromosomica, ma è probabile che si tratti di un fattore di una certa importanza.
Uno stadio molto precoce di speciazione cromosomica sembra si presenti in alcune popolazioni italiane del topo Mus musculus (v. Capanna e altri, 1977). Il cariotipo primitivo di tale specie, presente in tutta Europa e generalmente distinto in sottospecie geografiche, presenta 40 cromosomi a bastoncello. Tuttavia, in molte regioni italiane, come nella zona alpina, in Lombardia, in Abruzzo, in Molise e in Sicilia, esistono popolazioni in cui il numero dei cromosomi si è ridotto a 26, 24 o 22 per la presenza allo stato omozigote di 7, 8 o 9 cromosomi a due braccia derivati dalla fusione di due dei cromosomi originari. Queste popolazioni sembrano essere separate da quelle circostanti, con 40 cromosomi, da strette zone parapatriche in cui si formano ibridi. La fecondità degli ibridi è notevolmente ridotta da ‛incidenti' cromosomici alla meiosi.
Quale importanza ha il fatto che tutte queste fusioni cromosomiche si stabiliscano nelle popolazioni naturali? Queste popolazioni che presentano fusioni cromosomiche sono veramente specie incipienti? E se è cosi, quale stadio di speciazione hanno raggiunto? Attualmente non è possibile dare risposte precise a tali domande. È probabile che Mus musculus sia arrivato piuttosto recentemente in Europa dall'Asia, avendo ampiamente esteso la sua distribuzione in quanto commensale dell'uomo. È quindi probabile che le fusioni cromosomiche non abbiano più di qualche migliaio di anni. La vagilità di Mus musculus è certamente limitata e la sua popolazione è altamente frammentata, fattori questi che potrebbero favorire lo stabilirsi dei riassestamenti cromosomici purché fossero accompagnati da qualche vantaggio selettivo; ma la natura di tali vantaggi resta sconosciuta.
c) Speciazione simpatrica
I meccanismi cromosomici di speciazione che abbiamo descritto rientrano nella categoria che Mayr (v., 1963 e 1970) ha chiamato semigeografica, in opposizione al classico modello allopatrico di speciazione geografica, che prevede una completa separazione spaziale per le popolazioni in via di speciazione. All'estremo opposto si colloca la speciazione simpatrica, che non è geografica o è, al massimo, microgeografica. In passato tale modalità di speciazione è stata ritenuta impossibile da molti evoluzionisti; è stato comunque un argomento molto controverso.
Il concetto di speciazione simpatrica prevede che alcuni membri di una popolazione, adottando un nuovo tipo di cibo o un nuovo modello di vita, risultino, in un certo senso, isolati dal resto della popolazione all'interno della stessa area geografica. Un esempio ormai classico (anche se non si hanno elementi per affermare che la speciazione si sia completata) è rappresentato dalle mosche della frutta del genere Rhagoletis e in particolare Rhagoletis pomonella del Nordamerica, studiata da Bush (v., Modes of... e Sympatric..., 1975). Queste specie si riproducevano originariamente nel frutto del biancospino (Crataegus) negli Stati Uniti orientali, ma verso il 1864 fece la sua comparsa nella valle del fiume Hudson una razza di insetti che si nutriva delle mele che erano da poco state introdotte nella zona e gradualmente si diffuse da questo punto di origine in una grande area del Nordamerica orientale. Più recentemente sono comparse altre razze che si nutrono di prugne e di ciliegie.
Non vi è dubbio che l'origine di tali razze, che differiscono per l'ospite, sia sicuramente simpatrica. Il solo problema è se si tratti veramente di specie incipienti. Molte specie di Insetti, specialmente degli ordini dei Lepidotteri, Ditteri e Imenotteri, sono limitate esclusivamente a una singola specie di pianta ospite; analogamente anche molti tipi di vermi parassiti sono limitati a una sola specie di ospite. Sembra, perciò, che i cambiamenti della specializzazione trofica offrano ampie possibilità alla speciazione simpatrica. In alcuni casi può essere addirittura sufficiente un cambiamento da una parte a un'altra dello stesso ospite. Un esempio particolarmente significativo è stato descritto da Carson e Okada (v., 1980) ed è rappresentato dai moscerini del genere Drosophilella che vivono in Nuova Guinea. Si tratta di due specie correlate, una delle quali si riproduce sui fiori maschili e l'altra sui fiori femminili dell'infiorescenza ermafrodita di varie piante aroidi tropicali. Un'analoga coppia di specie è stata trovata nelle isole di Taiwan e di Okinawa. Sebbene molti dettagli di questo caso siano oscuri, a prima vista sembra che si tratti di un notevole esempio di speciazione simpatrica.
Il periodo di tempo necessario per il completamento della speciazione è, senz'altro, molto variabile. La prova più diretta proviene dall'arcipelago hawaiano, in cui l'età di ogni isola (tutte di origine vulcanica) è determinata con precisione dalla geologia. L'isola più recente, Hawaii, sorse dal mare circa 700.000 anni fa; in essa sono presenti un certo numero di specie endemiche di Drosophila, chiaramente originatesi dopo la colonizzazione da parte di insetti provenienti da isole più antiche (v. Carson, 1970). È quindi evidente che in 500.000-700.000 anni hanno potuto evolversi specie di Drosophila nettamente distinte.
7. Specie e speciazione nelle piante: poliploidia
Fra le piante la natura delle specie sembra essere essenzialmente la stessa che fra gli animali; vi sono però alcune differenze e, entro certi limiti, vi sono indubbie differenze nei meccanismi di speciazione. Molte specie di piante sono ‛poliploidi', vale a dire che nelle loro cellule vi sono più di due corredi cromosomici. Le specie animali, invece, sono quasi tutte ‛diploidi', hanno cioè due sorredi cromosomici nelle cellule somatiche e uno nei gameti (uova o spermatozoi). I poliploidi possono essere triploidi (3 corredi cromosomici), tetraploidi (4 corredi), pentaploidi (5), esaploidi (6), ecc.; i poliploidi i cui assetti cromosomici hanno numero dispari (triploidi e pentaploidi), per ciò che concerne la riproduzione sessuale sono necessariamente sterili. Invece i poliploidi con numero di assetti cromosomici pari (tetraploidi, esaploidi, ottoploidi) nella maggior parte dei casi hanno meiosi regolare e sono altamente fertili. Molte delle specie da raccolto più importanti - come il grano, l'avena, il sorgo, il riso e la canna da zucchero - sono poliploidi a numero di corredi cromosomici pari; altre, come il mais e l'orzo, sono diploidi.
Si pensava che vi fossero due tipi di poliploidi sostanzialmente diversi: autopoliploidi, aventi corredi cromosomici derivati tutti da una stessa specie ancestrale, e allopoliploidi, di origine ibrida, che deriverebbero i loro svariati corredi cromosomici da due o più specie ancestrali. Attualmente sembra invece che quasi tutte le specie di piante poliploidi naturali siano allopoliploidi, cioè di origine ibrida. Cosi la graminacea Triticum aestivum è un esaploide, i cui sei assetti cromosomici sono derivati due dal diploide Triticum monococcum, due da Aegilops longissima (o una specie strettamente correlata) e due da Aegilops squarrosa (v. Peacock e altri, 1981).
La speciazione da poliploidia è dunque un meccanismo importante nel regno vegetale. Nella maggior parte dei casi questo processo inizia con l'ibridazione tra due specie e produce un ibrido parzialmente sterile. Se le specie parentali sono diploidi potremmo designare i loro assetti cromosomici con AA e BB; gli ibridi saranno dunque AB e la loro sterilità sarà dovuta all'incapacità dei cromosomi di un corredo di appaiarsi regolarmente con quelli dell'altro nella meiosi. Se però avviene un raddoppiamento dei cromosomi (dovuto a una mitosi inefficace o solo non completata) avremo una cellula tetraploide con un corredo AABB. Se si sviluppa un ramo o una parte di una pianta con tale corredo cromosomico, essa sarà sessualmente fertile perché alla meiosi sarà possibile l'appaiamento dei due assetti cromosomici uguali tra di loro.
Circa la metà delle piante superiori sono probabilmente poliploidi (v. Grant, 1971): la frequenza è particolarmente elevata tra le Pteridofite, che infatti sono poliploidi per circa il 95% delle specie, e fra alcune famiglie di Angiosperme come le Graminacee, che sono poliploidi in circa il 70% dei casi. Fra le Conifere, invece, solamente l'1,5% delle specie è poliploide.
Le specie animali poliploidi sono poche; la poliploidia, infatti, ha svolto un ruolo molto meno importante nella speciazione animale. La ragione principale di questo fenomeno va ricercata senza dubbio nel fatto che la maggioranza delle specie animali è a sessi separati e la fecondazione incrociata è abitualmente obbligatoria perfino in gruppi di animali ermafroditi come i molluschi polmonati e i lombrichi. Così se in qualche modo viene prodotto un individuo tetraploide, potrà incrociarsi solo con un individuo diploide, nel qual caso la progenie sarà rappresentata da triploidi sterili. Fra le piante, invece, una certa possibilità di autofecondazione è quasi sempre presente. Le specie animali che si riproducono per partenogenesi rappresentano il quasi unico esempio di specie poliploidi; vi sono comunque rarissimi casi di specie sicuramente poliploidi fra le rane e si pensa in generale che lo stato poliploide si sia verificato durante l'evoluzione di alcune famiglie di Pesci.
Sembra probabile che, oltre alla poliploidia, tutti i meccanismi di speciazione presenti tra gli animali siano allo stesso modo presenti anche nel regno vegetale. La speciazione cromosomica è ben documentata fra alcune piante diploidi con numero cromosomico piccolo (Clarkia, Haplopappus, Brachycome), in cui sembra che sia di natura invasiva. Alcuni autori l'hanno definita speciazione quantica, ma questo termine è stato usato in senso più generale per includere tutti i tipi di speciazione in cui una specie incipiente si origina da una popolazione aberrante, periferica e semi-isolata, di una specie ancestrale.
Una caratteristica per la quale le piante sembrano differire dagli animali è la maggior frequenza di ibridazione tra specie diverse, con conseguente trasferimento di materiale genetico dal pool di una specie a quello di un'altra. Questo fenomeno, che è stato denominato ‛ibridazione introgressiva', sembra dovuto al fatto che tra le specie vegetali non esiste niente di paragonabile all'isolamento etologico. Fra gli animali, invece, l'isolamento etologico tra specie strettamente correlate è quasi sempre presente. C'è comunque una controversia di vecchia data su quanto questo fenomeno sia una conseguenza automatica della divergenza evolutiva, o sia piuttosto favorito dalla selezione naturale nelle aree in cui due specie si sovrappongono geograficamente (il cosiddetto effetto Wallace, dal nome del naturalista A.R. Wallace).
8. Il problema della rettangolarità nella speciazione
Negli ultimi anni un certo numero di paleontologi ha prospettato un'ipotesi secondo cui la maggior parte dei cambiamenti morfologici avvenuti durante l'evoluzione sono stati associati a eventi di speciazione. Questo fenomeno è stato indicato come ‛modello di interpunzione' dell'evoluzione (v. Eldredge e Gould, 1972; v. Stanley, 1979)0 come ‛rettangolarità'. Secondo il punto di vista opposto e più tradizionale, che è generalmente chiamato ‛gradualismo filetico' o ‛modello gradualistico', i cambiamenti morfologici avvengono con una frequenza più o meno costante durante tutto il periodo evolutivo, senza una particolare accelerazione nel periodo della speciazione. Il modello della rettangolarità era basato sulla conoscenza, acquisita da documenti paleontologici, della durata della morfospecie. Per i Mammiferi, durante il periodo cenozoico, questa durata era di circa 2 milioni di anni; vale a dire, l'entità della differenziazione morfologica degli animali, la cui evoluzione filetica è di tipo continuo, è all'incirca equivalente a quella riscontrabile fra due specie viventi strettamente correlate appartenenti allo stesso gruppo. Nonostante che la durata della morfospecie sia relativamente lunga, alcuni ordini di Mammiferi, come i pipistrelli e le balene (dei quali, grazie ai reperti fossili, è attualmente nota la storia), si separarono dai primitivi progenitori non specializzati, probabilmente simili agli attuali insettivori, in circa 12 milioni di anni. Come dice Stanley: ‟In questo intervallo di tempo [...] possono essersi succedute solo cinque o dieci specie di normale longevità che hanno prodotto questa enorme quantità di mutamenti evolutivi". Poiché non vi è nessuna prova di una generale accelerazione dei cambiamenti morfologici durante l'intera durata di queste discendenze, Stanley deduce che i mutamenti morfologici devono essere stati notevolmente accelerati durante ogni evento di speciazione.
A sostegno di tale punto di vista Stanley (v., 1979) ha presentato notevoli prove tratte da studi fatti sulla base di testimonianze fossili sia di Vertebrati che di Invertebrati. Comunque, la ‛rettangolarità' è quasi sicuramente una caratteristica di speciazione non universale, altrimenti non avremmo un numero così grande di specie correlate, come discusso in precedenza, difficilmente differenziabili dal punto di vista morfologico. Sembra che i sostenitori del concetto della rettangolarità basino le loro ipotesi soprattutto sul modello di Ernst Mayr (v., 1954) secondo il quale, in alcuni tipi di speciazione allopatrica, una specie incipiente o potenziale andrebbe incontro a una ‛rivoluzione genetica' (cioè alla sostituzione di molti geni da parte di nuovi alleli ottenuti per mutazione).
Sembra comunque più probabile che proprio nella speciazione allopatrica la rettangolarità si verifichi difficilmente. Le popolazioni allopatriche che stanno subendo un processo di speciazione in genere si troveranno in habitat simili e avranno dei modi di vita analoghi a quelli della popolazione ancestrale; di conseguenza ci sarà una selezione abbastanza piccola per modificazioni morfologiche notevoli. D'altra parte, la speciazione simpatrica sembra implicare nella maggior parte dei casi un adattamento a un nuovo habitat, a una nuova fonte di cibo o a un nuovo tipo di vita (o a tutte le cose contemporaneamente). È quindi verosimile che la selezione per nuovi caratteri morfologici sia molto forte. Si sono avuti alcuni casi di ‛speciazione esplosiva' nei pesci di acqua dolce, in cui specie notevolmente diverse sembrano essersi evolute simpatricamente in poche migliaia di anni: esse possono essere considerate, con ogni probabilità, esempi di forte rettangolarità, dovuta, nella maggior parte dei casi, allo sviluppo di nuove specializzazioni trofiche.
Chiaramente la natura delle specie e dei processi evolutivi mediante i quali esse si originano non sarà interamente chiarita finché non avremo un quadro molto più completo dell'architettura molecolare del materiale genetico. La tanto decantata modem synthesis di J. Huxley (v., 1942) della teoria evolutiva è attualmente considerata come un inizio piuttosto che come un punto di arrivo di una completa comprensione del processo evolutivo. In ultima analisi, il concetto di specie e di speciazione occuperà un posto centrale nella teoria evolutiva a prescindere dalla possibilità o meno che la rettangolarità dimostri di essere un reale e importante aspetto dell'evoluzione.
Bibliografia
Ayala, F. J., Valentine, J. W., Evolving: the theory and process of organic evolution, Menlo Park 1979.
Bateson, W., Evolutionary faith and modern doubts, in ‟Science", 1922, LV, pp. 55-61.
Bocquet, C., Recherches sur le polymorphisme naturel de Jaera marina (Fahr.) (Isopoted Asellotes). Essai de systématique évolutive, in ‟Archives de zoologie expérimentale et générale", 1953, XC, pp. 187-450.
Britton, J., Thaler, L., Evidence for the presence of two sympatric species of mice (genus Mus L.) in southern France based on biochemical genetics, in ‟Biochemical genetics", 1978, XVI, pp. 213-225.
Britton-Davidian, J., Bonhomme, F., Croset, H., Capanna, E., Thaler, L., Variabilité génétique chez les populations de souris (genre Mus L.) à nombre chromosomique réduit, in ‟Comptes rendus hebdomadaires des séances de l'Académie des Sciences" (Série D), 1980, CCXC, pp. 195-198.
Bullini, L., Nascetti, G., Ciafrè, S., Rumore, F., Biocca, E., Ricerche cariologiche ed elettroforetiche su Parascaris univalens e Parascaris equorum, in ‟Rendiconti dell'Accademia Nazionale dei Lincei. Classe di scienze fisiche, matematiche e naturali", 1978, LXV, pp. 151-156.
Bush, G. L., Modes of animal speciation, in ‟Annual review of ecological systematics", 1975, VI, pp. 339-364.
Bush, G. L., Sympatric speciation in phytophagous parasitic insects, in Evolutionary strategies of parasitic insects (a cura di P. W.Price), London 1975, pp. 187-206.
Capanna, E., Civitelli, M. V., Cristaldi, M., Chromosomal rearrangement, reproductive isolation and speciation in Mammals: the case of Mus musculus, in ‟Bollettino di zoologia", 1977, XLIV, pp. 213-246.
Carson, H. L., Chromosome tracers of the origin of species, in ‟Science", 1970, CLXVIII, pp. 1414-1418.
Carson, H. L., Hardy, D. E., Spieth, H. T., Stone, W. S., The evolutionary biology of the Hawaiian Drosophilidae, in Essays in evolution and genetics in honor of Theodosius Dobzhansky (a cura di M. K. Hecht e W. C. Steare), New York 1970, pp. 437-543.
Carson, H. L., Okada, T., The ecology and evolution of some flowerbreeding Drosophilidae of New Guinea, in Abstracts of the XVI international congress of entomology, Kyoto 1980, p. 7.
Coluzzi, M., Sabatini, A., Petrarca, V., Di Deco, M. A., Chromosomal differentiation and adaptation to human environments in the Anopheles gambiae complex, in ‟Transactions of the Royal Society of Tropical Medicine and Hygiene", 1979, LXXIII, pp. 483-497.
Craddock, E. M., Chromosome number variation in a stick insect, Didymuria violescens (Leach), in ‟Science", 1970, CLXVII, pp. 1380-1382.
Craddock, E. M., Intraspecific karyotypic differentiation in the Australian phasmatid Didymuria violescens (Leach). I. The chromosomal races and their structural and evolutionary relationships, in ‟Chromosoma", 1975, LIII, pp. 1-24.
Daly, J. C., Wilkinson, P., Shaw, D. D., Geographic distribution of allozyme variation within and between the four chromosomal taxa of the grasshopper Caledia captiva and its relationship to population subdivision and reproductive isolation, in ‟Evolution", 1981, XXXV.
Dobzhansky, Th., Genetics and the origin of species, New York 1937.
Eldredge, N., Gould, S. J., Punctuated equilibria: an alternative to phyletic gradualism, in Models in paleobiology (a cura di T. J. M. Schopf), San Francisco 1972, pp. 82-115.
Fisher, R. A., The genetical theory of natural selection, Oxford 1930.
Grant, V., Plant speciation, New York 1971.
Highton, R., Larson, A., The genetic relationships of the salamanders of the genus Plethodon, in ‟Systematic zoology", 1979, XXVIII, pp. 579-599.
Huxley, J. S., Evolution: the modern synthesis, London 1942.
Key, K. H. L., Speciation in the Australian morabine grasshoppers: taxonomy and ecology, in Genetic mechanisms of speciation in Insects (a cura di M. J. D. White), Sydney 1974, pp. 43-56.
Lécher, P., Cytogénétique de l'hybridization expérimentale et naturelle chez l'isopode Jaera (albifrons) Syei Bocquet, in ‟Archives de zoologie expérimentale et générale", 1967, CVIII, pp. 633-698.
Lécher, P., Polymorphisme chromosomique dans les populations baltes et scandinaves de l'isopode Jaera (albifrons) Syei Bocquet, in ‟Archives de zoologie expérimentale et générale", 1968, CIX, pp. 211-227.
Mancino, G., Ragghianti, M., Bucci-Innocenti, S., Cytotaxonomy and cytogenetics in European newt species, in The reproductive biology of Amphibians (a cura di D. H. Taylor e S. I. Guttman), New York 1977, pp. 411-447.
Mayr, E., Systematics and the origin of species, New York 1942.
Mayr, E., Change of genetic environment and evolution, in Evolution as a process (a cura di J. S. Huxley, A. C. Hardy ed E. B. Ford), London 1954, pp. 157-180.
Mayr, E., Animal species and evolution, Cambridge, Mass., 1963 (tr. it.: L'evoluzione delle specie animali, 2 voll., Torino 1970).
Mayr, E., Populations, species and evolution, Cambridge, Mass., 1970.
Nei, M., Genetic distance between populations, in ‟American naturalist", 1972, CVI, pp. 283-292.
Nei, M., Molecular population genetics and evolution, Amsterdam 1975.
Peacock, W. J., Gerlach, W. L., Dennis, E. S., Molecular aspects of wheat evolution: repeated DNA sequences, in Wheat science. Today and tomorrow (a cura di L. T. Evans e W. J. Peacock), London 1981, pp. 41-60.
Reig, O., Aquilera, M., Barros, M. A., Useche, M., Chromosomal speciation in a rassenkreis of Venezuelan spiny rats (genus Proechimys, Rodentia, Echimyidae), in ‟Genetica", 1980, LII-LIII, pp. 291-312.
Sarich, V. M., Rates, sample sizes and the neutrality hypothesis, in ‟Nature", 1977, CCLXIII, pp. 24-28.
Stanley, S. M., Macroevolution: pattern and process, San Francisco 1979.
Wasserman, M., Cytology and phylogeny of Drosophila, in ‟American naturalist", 1963, XCVII, pp. 333-352.
White, M. J. D., Models of speciation, in ‟Science", 1968, CLIX, pp. 1065-1070.
White, M. J. D., Models of speciation, San Francisco 1978.
Zimmerman, E. G., Kilpatrick, C. W., Hart, B. J., The genetics of speciation in the rodent genus Peromyscus, in ‟Evolution", 1978, XXXII, pp. 565-579.