
specializzazione emisferica
specializzazione emisferica
Nell’uomo l’emisfero di sinistra è specializzato nelle funzioni del linguaggio, nel controllo della mano dominante destra e nell’organizzazione della motilità volontaria. L’emisfero destro prevale invece nella cognizione dello spazio e nella regolazione dell’emotività. Lo stile cognitivo dell’emisfero sinistro è sostanzialmente verbale, analitico e locale, mentre quello dell’emisfero destro è spaziale, sintetico e globale. Esistono asimmetrie comportamentali e specializzazioni emisferiche cerebrali in molte specie animali, con una tendenza dell’emisfero sinistro a controllare i comportamenti esplorativi e alimentari e dell’emisfero destro a controllare i comportamenti a carattere emotivo. Esistono modelli genetici ed evolutivi per la spiegazione dei fenomeni di lateralità e di specializzazione emisferica, ma nessuno di essi ha trovato finora una conferma definitiva. [➔ cervello, evoluzione del; corteccia cerebrale; funzioni cerebrali superiori; imaging cerebrale, diagnosi e ricerca; linguaggio] La s. e. denota un’asimmetria funzionale tra gli emisferi del cervello umano. Come il sistema nervoso centrale dei vertebrati, il cervello umano presenta una simmetria strutturale bilaterale per la quale la metà destra è all’incirca l’immagine speculare di quella sinistra e viceversa. A questa simmetria strutturale non corrisponde una simmetria funzionale poiché ciascun emisfero cerebrale dell’uomo possiede funzioni proprie, almeno in parte differenziate da quelle dell’altro emisfero. Fin dal 19° sec., studi clinici e anatomopatologici avevano dimostrato disturbi specifici del linguaggio parlato e scritto (➔ afasia) a seguito di lesioni di determinate regioni corticali dell’emisfero sinistro, ma non del destro, quali l’area di Broca nel lobo frontale e l’area di Wernicke nel lobo temporale. A sostegno di una sua superiorità funzionale, sta anche il fatto che l’emisfero sinistro presiede alle funzioni di senso e di moto della mano destra, che per una cospicua caratteristica della specie umana è utilizzata di preferenza nelle varie attività manuali della vita quotidiana. Inoltre, lesioni cerebrali specifiche a sinistra, ma non a destra, possono causare l’incapacità di eseguire azioni e gesti su comando verbale o imitazione (➔ aprassia), pur in assenza di paralisi, a indicare il possesso del controllo superiore della motilità intenzionale da parte dell’emisfero sinistro. Tutti questi argomenti, a favore di una dominanza funzionale assoluta dell’emisfero sinistro su quello destro, per molto tempo hanno fatto sì che a quest’ultimo venisse attribuito un ruolo funzionale minore e subordinato.
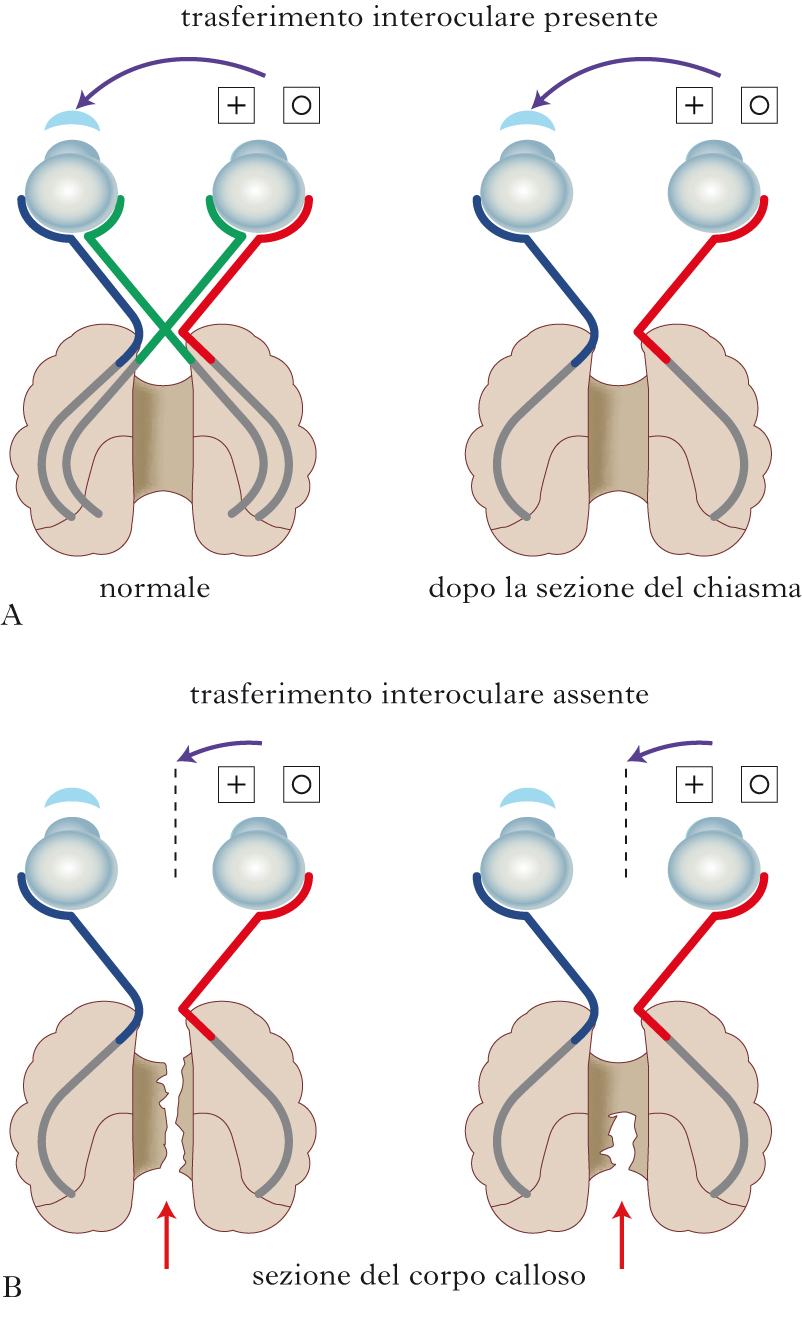
Specializzazione contro dominanza assoluta
Nella seconda metà del 20° sec. la dottrina della dominanza assoluta dell’emisfero sinistro è stata riveduta e corretta da importanti osservazioni sperimentali: l’analisi accurata di disturbi indipendenti dal linguaggio, causati da lesioni specifiche dell’emisfero destro; lo studio delle prestazioni cognitive controllate da ciascun emisfero in pazienti con sezione chirurgica delle connessioni interemisferiche; la misura, in soggetti normali, di piccole differenze di accuratezza e velocità nell’elaborazione di informazioni limitate sperimentalmente all’emisfero destro o sinistro. A questi reperti si sono aggiunti più recentemente quelli forniti dalle tecniche non invasive delle neuroimmagini (➔ imaging cerebrale, diagnosi e ricerca), che consentono di determinare la sede e le dimensioni di lesioni emisferiche in vivo, nonché di correlare in soggetti sani le attivazioni regionali che in ciascun emisfero si associano selettivamente con lo svolgimento di compiti comportamentali e mentali. La separazione chirurgica degli emisferi, tramite la sezione del corpo calloso, eseguita al fine di controllare forme di epilessia refrattarie ai farmaci, ha offerto l’opportunità di distinguere le varie specializzazioni funzionali degli emisferi di destra e di sinistra dello stesso cervello. Quando il corpo calloso è tagliato, le informazioni elaborate da un solo emisfero generano esperienze coscienti limitate a quell’emisfero, che quindi non vengono a conoscenza dell’altro. In accordo con gli effetti di lesioni uniemisferiche circoscritte, nonché con i risultati ottenuti su soggetti con cervello intatto, gli studi sui pazienti con sconnessione interemisferica hanno dimostrato che l’emisfero destro, pur privo di linguaggio, prevale di poco, ma sistematicamente, sul sinistro nel riconoscimento dei volti e delle espressioni facciali, nell’inserimento appropriato di un frammento in un disegno, nella discriminazione e nella memorizzazione di forme prive di senso, nella distinzione di note musicali, nella capacità di ricostruire un tutto da componenti separate, nella rotazione mentale di forme bi- e tridimensionali, nell’intonazione emotiva del linguaggio parlato. I due emisferi hanno anche ruoli diversi nelle funzioni della memoria: la corteccia prefrontale dell’emisfero sinistro prevale nella memorizzazione di accadimenti personali, e la corteccia prefrontale dell’emisfero destro prevale nella rievocazione di tali accadimenti. Non esiste pertanto una dominanza funzionale assoluta di un emisfero sull’altro, bensì una s. e. funzionale e una divisione del lavoro fra due parti di un organo in possesso di competenze parzialmente diverse e complementari, che normalmente contribuiscono ai processi psicologici e al controllo del comportamento unitamente, ma in modi differenziati. Nel tentativo un poco semplicistico di caratterizzare le modalità operative dei due emisferi si usano aggettivi come verbale, analitico e locale per l’emisfero sinistro, e spaziale, sintetico e globale per l’emisfero destro. Altri tentativi più estremi di attribuire ai due emisferi caratteristiche personali come la razionalità e il conformismo all’emisfero sinistro e l’intuitività e la creatività all’emisfero destro sono esagerazioni caricaturali assolutamente da evitare.
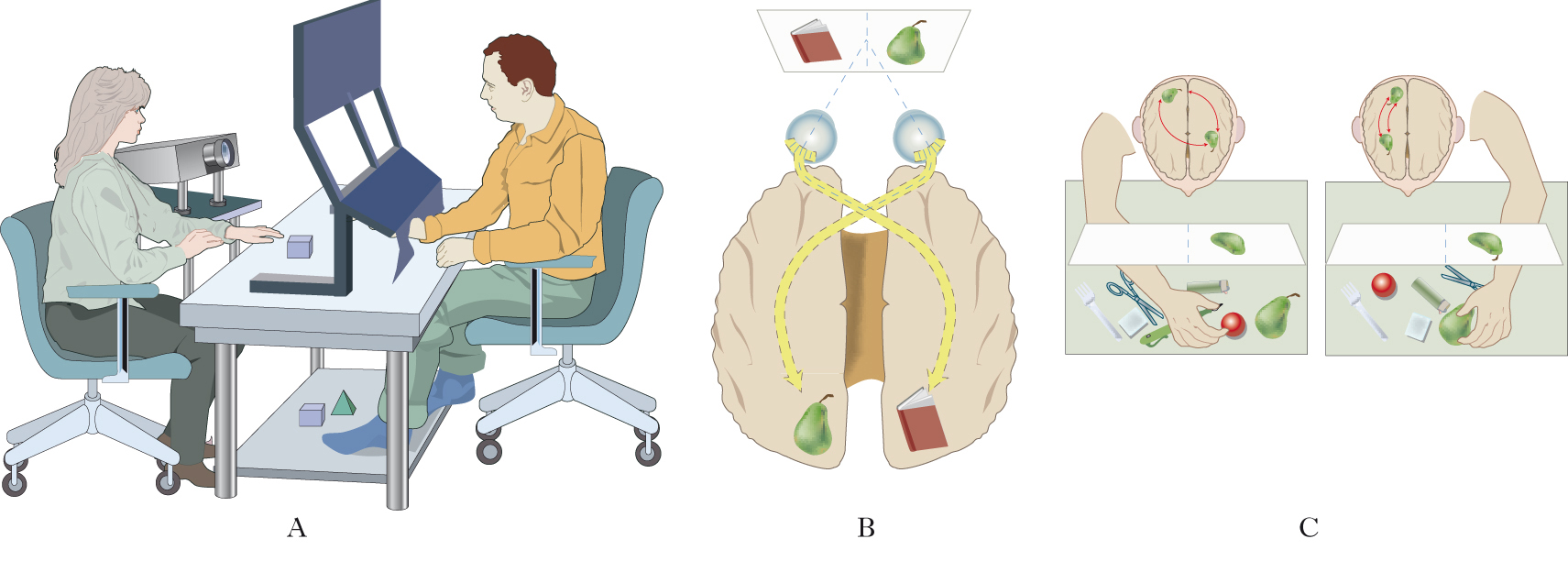
Dominanza manuale e lateralizzazione dei centri del linguaggio
In tutte le popolazioni umane oggi esistenti ca. 8 o 9 individui su 10 utilizzano di preferenza la mano destra in tutte o quasi tutte le normali attività manuali, contro una minoranza di individui che preferiscono invece la mano sinistra o sono ambidestri. Ci si riferisce qui alla scelta manuale spontanea, non contaminata da fattori culturali come l’imposizione antica della mano destra per la scrittura. Da ricerche paleoantropologiche si sa che il rapporto di 8÷9 a 1 fra destrimani e mancini è rimasto stabile fin dalla preistoria dell’uomo. Per localizzare i centri del linguaggio in cervelli di destrimani e mancini si sono utilizzate le alterazioni della capacità di parlare quando l’uno o l’altro emisfero era stato inattivato con farmaci o stimolazioni elettriche per ragioni terapeutiche. In circa il 95% dei destrimani e nel 70% dei mancini l’emissione verbale è soppressa dalla sola inattivazione dell’emisfero sinistro; l’opposto avviene nel rimanente 5% dei destrimani e nel 15% dei mancini, nei quali è l’inattivazione dell’emisfero destro a interferire con il linguaggio. Nel rimanente 15% dei mancini il linguaggio non viene soppresso dall’inattivazione dell’emisfero sinistro né del destro, a dimostrazione di una presumibile bilateralità dei centri del linguaggio. Recenti esperimenti (2009), con tecniche non invasive su soggetti normali, pur confermando in buona parte questi risultati, suggeriscono che le percentuali di mancini con linguaggio a sinistra e a destra sarebbero pari rispettivamente all’80 e al 20%, e che la rappresentazione bilaterale del linguaggio probabilmente sarebbe presente solo in pochi soggetti autenticamente ambidestri o con lesioni neurologiche. Complessivamente il linguaggio risulterebbe localizzato a sinistra in 90 individui su 100, ma di questi 83 sarebbero destrimani e 7 mancini o ambidestri. Quindi l’emisfero sinistro non è la sede esclusiva del linguaggio in tutti gli individui, né la dominanza manuale, imperfettamente correlata con il lato dominante per il linguaggio, può fornire di per sé indicazioni certe sull’emisfero parlante nel singolo individuo. L’associazione in uno stesso emisfero dei centri del linguaggio e del controllo della mano dominante potrebbe essere avvenuta nell’evoluzione umana durante uno stadio di transizione fra la comunicazione gestuale, eseguita prevalentemente dalla mano dominante, e la comunicazione vocale.
Genetica e maturazione della lateralizzazione manuale e cerebrale
La lateralità manuale è almeno in parte sotto controllo genetico, che peraltro è indipendente dal controllo genetico della posizione asimmetrica di visceri impari come il cuore e il fegato. Nella condizione chiamata situs inversus, con cuore a destra e fegato a sinistra, la lateralità manuale e quella cerebrale non risultano alterate. Uno dei modelli genetici più noti postula l’esistenza di un polimorfismo genetico bilanciato, basato su un allele RS+ favorente la lateralizzazione manuale a destra e la lateralizzazione del linguaggio a sinistra. Gli omozigoti RS++ sarebbero fenotipicamente destrimani con il linguaggio localizzato a sinistra, al pari del 75% degli eterozigoti RS+–, mentre nei restanti eterozigoti e negli omozigoti RS– lateralità manuale ed emisferica per il linguaggio sarebbero distribuite sostanzialmente a caso e indipendentemente l’una dall’altra, per azione di fattori epigenetici pre- e postnatali, dalla posizione nell’utero all’azione di ormoni, alle influenze sociali e culturali. Ancorché ipotetico, e privo di riscontri in termini di ubicazione cromosomica del presunto locus genetico, il modello può spiegare vari dati demografici, inclusa la discordanza per la preferenza manuale che si riscontra in qualche coppia di gemelli monozigotici. La preferenza di una mano sull’altra nella specie umana è molto precoce, tanto che si manifesta già prima della nascita. Il feto comincia a succhiarsi un pollice già dalla 15a settimana di vita intrauterina, e i feti che succhiano il pollice destro sono in netta maggioranza rispetto a quelli che succhiano il sinistro. Quanto agli emisferi, il destro matura prima del sinistro, e la ritardata maturazione dell’emisfero sinistro può spiegare la sua maggiore vulnerabilità agli agenti patologici durante la prima infanzia. Uno dei fattori ritardanti la maturazione dell’emisfero sinistro è l’ormone maschile testosterone. Studi con varie tecniche dimostrano che l’emisfero destro prevale sul sinistro nella discriminazione di timbri musicali a partire da due mesi di età, e nella discriminazione di facce dal quarto mese. L’esposizione anche breve al linguaggio nativo produce un’attivazione specifica in sede temporale sinistra a partire dai due anni di età, probabilmente in relazione allo sviluppo della comprensione verbale.
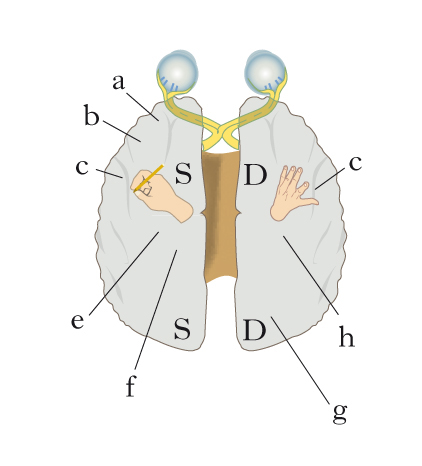
Asimmetrie strutturali fra gli emisferi
Pur nella loro sostanziale simmetria strutturale, gli emisferi cerebrali dell’uomo presentano alcune differenze relativamente modeste fra destra e sinistra, la cui correlazione con le asimmetrie e specializzazioni funzionali di lato è ancora in buona parte sconosciuta. Fra le asimmetrie macroscopiche, vi è un maggior sviluppo relativo della parte anteriore dell’emisfero destro e della parte posteriore dell’emisfero sinistro, mentre la scissura silviana di sinistra è più lunga e rettilinea di quella di destra. La correlazione potenziale più studiata fra asimmetrie cerebrali morfologiche e specializzazione funzionale riguarda proprio la scissura silviana, il cui labbro inferiore contiene il planum temporale che già in antichi studi autoptici era risultato più esteso nell’emisfero sinistro che in quello destro nella maggioranza dei casi, in una percentuale oggi valutabile almeno pari all’80%. Il grado di asimmetria a favore del planum di sinistra è massimo nei destrimani, intermedio nei mancini non familiari (cioè senza nessun parente di primo grado mancino), minimo o nullo nei mancini familiari. Peraltro il significato di questa correlazione con la dominanza manuale non può che essere indiretto, poiché la corteccia del planum temporale, corrispondente in parte all’area di Wernicke, non ha funzioni motorie bensì uditive e linguistiche. Studi recenti (2009) indicano che le asimmetrie morfologiche importanti per quelle funzionali vadano ricercate più sul piano dell’organizzazione istologica e citoarchitettonica di aree come quelle di Wernicke e di Broca. Tali aree presentano infatti caratteri microstrutturali, come la presenza di grandi neuroni, la ricchezza degli alberi dendritici, la spaziatura dei moduli colonnari e la connettività intra- e interareale che distinguono l’emisfero sinistro da quello destro. Molte, se non tutte, le differenze tra emisfero destro e sinistro osservate in adulti sono presenti anche in feti e infanti, ad attestare una loro origine almeno in parte genetica.
Asimmetrie comportamentali e cerebrali in animali non umani
I fenomeni di lateralità comportamentale e le differenze funzionali fra gli emisferi cerebrali sono stati a lungo ritenuti appannaggio della sola specie umana, in relazione a capacità uniche come il linguaggio, la fabbricazione di utensili e gli sviluppi intellettuali e tecnologici che ne derivano. Da alcuni anni questa visione è cambiata radicalmente poiché sono state descritte e studiate in molti ordini e specie di vertebrati lateralizzazioni comportamentali e cerebrali individuali e di popolazione, per le quali in una data funzione la maggioranza degli individui (dal 65 al 95%) di una data specie manifesta un’asimmetria nella stessa direzione. Sono state accertate lateralizzazioni di popolazione in pesci, anfibi, uccelli e mammiferi in prestazioni come il riconoscimento di prede, predatori e conspecifici, il corteggiamento e l’accoppiamento, i comportamenti di fuga e di aggressione. Vi è una tendenziale specializzazione del lato sinistro del cervello per comportamenti di approccio, alimentari ed esplorativi dell’ambiente, e del lato destro per comportamenti di evitamento o aggressione, e in genere per risposte emotive. A somiglianza con la specie umana, l’emisfero sinistro di varie specie di mammiferi non umani sembra essere specializzato per il riconoscimento di segnali vocali di membri della stessa specie. Recentemente (2009) è stata confermata una preferenza manuale destra nel primate evolutivamente più vicino all’uomo, lo scimpanzé. Due scimpanzé su tre mostrano una preferenza stabile della mano destra sulla sinistra in attività come estrarre cibo da un tubo, fare gesti e lanciare oggetti. Poiché alcuni reperti fossili indicano una preferenza per la mano destra in ominidi vissuti 2 milioni di anni fa, non si esclude la possibilità che tale preferenza fosse già presente in un antenato comune all’uomo e allo scimpanzé. Di notevole interesse, quali possibili omologie con la specie umana, sono le asimmetrie di lato rilevate con la risonanza magnetica funzionale in aree corticali dello scimpanzé corrispondenti alle aree per il linguaggio nell’uomo, come il giro frontale inferiore e il planum temporale, che sono risultate significativamente più estese a sinistra che a destra. È possibile che queste aree corticali dello scimpanzé servano alla comunicazione gestuale.
Le ragioni delle asimmetrie di lato e della specializzazione funzionale emisferica
Elaborazioni complesse così diverse come il linguaggio e la cognizione dello spazio richiedono computazioni specifiche eseguite da circuiti neuronali dedicati, ed è verosimile che vi sia stato un vantaggio evolutivo nel dividere tali circuiti in emisferi diversi per evitare sovrapposizioni, duplicazioni e reciproche interferenze. Una mancanza di lateralizzazione (ambidestrismo e bilateralità della rappresentazione cerebrale del linguaggio) sembra essere associata a una riduzione significativa dell’abilità scolastica, e la mancata lateralizzazione emisferica della componente fonologica del linguaggio sembra essere associata con la schizofrenia. D’altra parte anche la lateralizzazione estrema può presentare costi: l’eccellenza nelle funzioni del linguaggio, tipica dei destrimani omozigoti per l’allele RS+, comporta una ridotta efficienza nelle abilità spaziali e sportive, e i mancini sono più suscettibili dei destrimani ai disturbi della lettura. Molte lateralizzazioni di popolazione, come la preferenza manuale destra nell’uomo, sono caratterizzate dall’ostinata persistenza di una minoranza che va contro la direzione favorita dalla maggioranza. Questa minoranza potrebbe avere vantaggi adattativi (per es. nella lotta e in attività sportive individuali, come il tennis), rimanendo appunto minoranza, per la capacità di mettere in difficoltà destrimani abituati a scontrarsi con altri destrimani. Similmente è stato proposto che per i pesci che vivono in banchi vi sia una maggiore probabilità di sopravvivenza della specie, se nell’incontro con un predatore una maggioranza fugge costantemente in una direzione e una minoranza fugge costantemente nella direzione opposta. Giovanni Berlucchi