
limbico, sistema
limbico, sistema
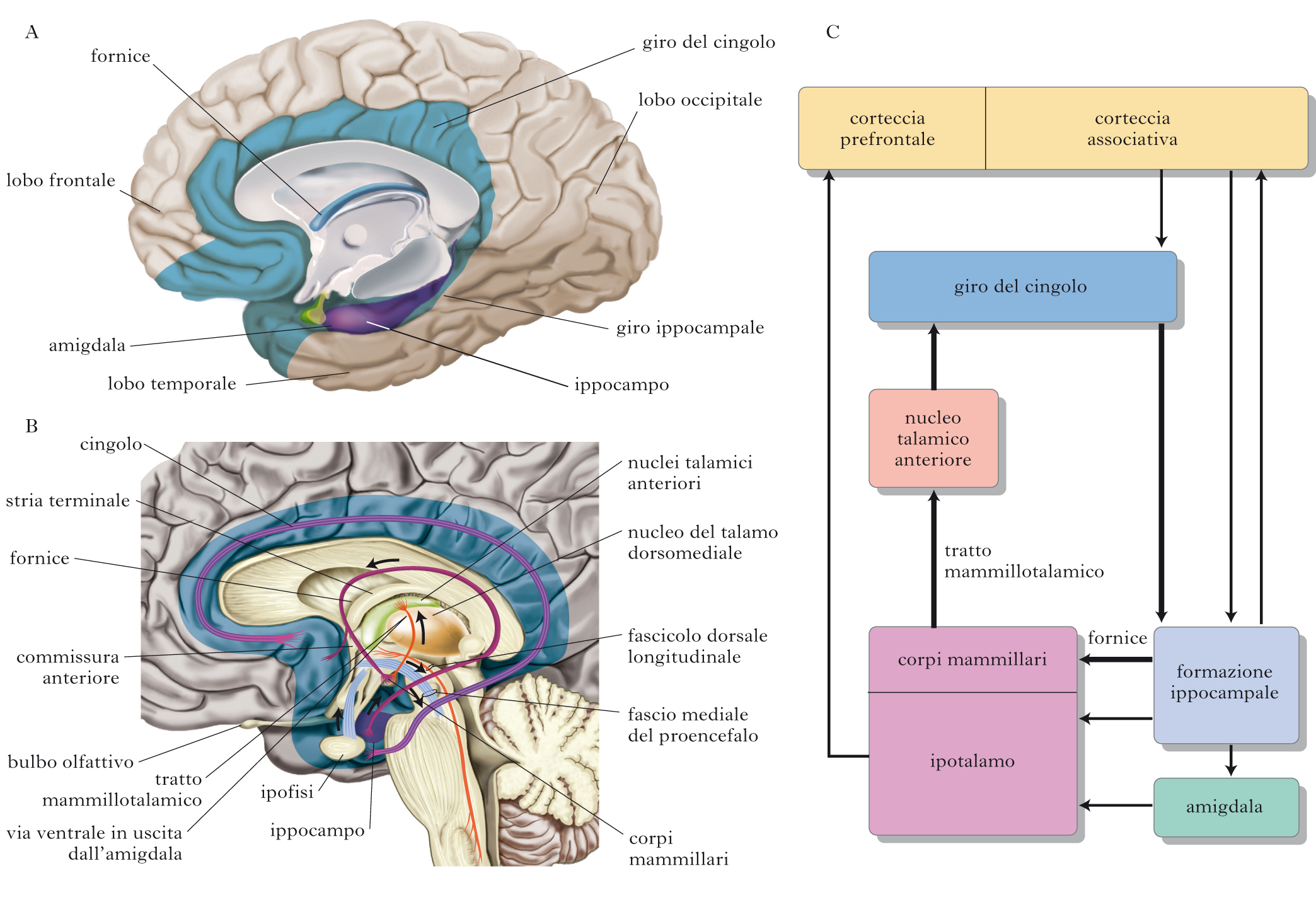
Insieme di formazioni cerebrali del telencefalo e del diencefalo implicate nel controllo delle emozioni e delle reazioni comportamentali legate alla sopravvivenza della specie (paura, attacco e difesa, attività sessuale) e della memoria. Nel 1937 James Papez, basandosi sui risultati di una serie di esperimenti che suggerivano un ruolo determinante dell’ipotalamo (➔) nell’espressione delle emozioni, suggerì che il sistema l. potesse essere il substrato anatomico delle emozioni (➔). Il concetto di sistema l. deriva dal lobo limbico, una regione identificata da Paul Broca, composta da aree filogeneticamente primitive, posizionate a formare un anello intorno al tronco encefalico. Il lobo limbico comprende il giro paraippocampale, il giro del cingolo e il giro subcallosale e la sottostante corteccia della formazione ippocampale (morfologicamente più semplice rispetto alle cortecce superiori). Le aree del sistema l. formano circuiti complessi che insieme giocano un ruolo essenziale in alcune funzioni cognitive superiori, come l’apprendimento, la memoria e le emozioni. Papez propose che, dal momento che le emozioni raggiungono il livello del pensiero consapevole e i pensieri possono influenzare le emozioni, debba esserci una comunicazione reciproca tra i centri corticali e l’ipotalamo. Egli suggerì quindi che la corteccia influenza l’ipotalamo tramite la connessione diretta con il giro del cingolo e la connessione tra il giro del cingolo e la formazione ippocampale. In linea con tale ipotesi, la formazione ippocampale avrebbe la possibilità di processare le informazioni in arrivo dal giro del cingolo e inviarle direttamente ai corpi mammillari dell’ipotalamo (tramite il fornice). L’ipotalamo, a sua volta, convoglierebbe le informazioni al giro del cingolo tramite un percorso che parte dai corpi mammillari, arriva ai nuclei talamici anteriori e, da questi, conduce al giro del cingolo. Il concetto di sistema l. è stato successivamente ampliato da Paul MacLean per includere altre strutture funzionalmente e anatomicamente collegate a quelle descritte da Papez. MacLean incluse nel sistema l. anche parti dell’ipotalamo, l’area del setto, il nucleo accumbens (parte dello striato), aree corticali come la corteccia orbitofrontale e l’amigdala. Numerosi studi anatomici hanno dimostrato l’effettiva esistenza di circuiti di connessione tra aree corticali superiori, formazione ippocampale e amigdala.
Studi sull’ipotalamo
Stephen W. Ranson e Walter R. Hess idearono una serie di esperimenti che, tramite l’impianto di elettrodi permanenti, permisero di osservare gli effetti della stimolazione cerebrale in animali svegli e liberi di muoversi. La stimolazione di parti differenti dell’ipotalamo produce una costellazione di risposte che dà origine a comportamenti organizzati. Per es., la stimolazione elettrica dell’ipotalamo laterale evoca, nel gatto, risposte autonomiche e somatiche tipiche della rabbia, quali aumento della pressione sanguigna, costrizione pupillare, inarcamento del dorso e sollevamento della coda. Tali osservazioni fornirono le basi per giungere all’importante conclusione che l’ipotalamo, diversamente da quanto ritenuto in passato, non è unicamente un nucleo motorio, ma piuttosto un centro dove input di natura diversa si integrano per generare risposte somatiche e autonomiche complesse, organizzate e coerenti. Dato che molte risposte prodotte dall’attivazione dell’ipotalamo coincidevano con risposte osservabili durante esperienze emotive tipiche, Hess suggerì che l’ipotalamo avesse un ruolo specifico nel coordinare l’espressione dei comportamenti associati agli stati emotivi. Indagini successive hanno effettivamente dimostrato che aree distinte dell’ipotalamo regolano stati emotivi specifici: mentre la stimolazione dell’ipotalamo laterale evoca una risposta di rabbia, lesioni alla stessa area inducono stati di calma; al contrario, lesioni dell’ipotalamo mediale provocano una maggiore suscettibilità agli stati eccitatori e rendono l’animale maggiormente propenso a manifestare risposte aggressive; livelli di irritabilità paragonabili sono osservabili anche a seguito di decorticazione. Pertanto, l’azione reciproca tra l’attività dell’ipotalamo e l’attività dei centri superiori dà origine alle esperienze emotive che descriviamo come paura, rabbia, piacere e soddisfazione. L’idea è supportata dal comportamento di pazienti con rimozione di parti del sistema l. (zone della corteccia frontale o del giro del cingolo). Tali pazienti, nonostante percepiscano il dolore ed esibiscano risposte autonomiche appropriate, non sperimentano più la profonda esperienza emotiva di afflizione associata al dolore percepito.
Sistema limbico e controllo delle emozioni
L’attuale modello interpretativo delle basi neuronali delle emozioni è principalmente basato sulle idee proposte da Papez, che descrisse correttamente il ruolo rilevante della corteccia cingolata e del giro paraippocampale nella percezione delle emozioni. Non è invece risultata valida l’ipotesi di Papez secondo cui l’ippocampo sarebbe il coordinatore dell’attività dell’ipotalamo, ruolo oggi attribuito all’amigdala (➔). Attualmente, anche il ruolo dei corpi mammillari e dei nuclei talamici anteriori nelle emozioni è stato molto ridimensionato; tali strutture sono piuttosto legate ai processi di immagazzinamento mnestico. Secondo il modello, l’ipotalamo integra le risposte motorie ed endocrine generando il comportamento emotivo appropriato, mentre il telencefalo filtra il significato degli stimoli ambientali sopprimendo le risposte emotive a quelli privi di senso. Tramite il controllo operato dalle cortecce superiori che mediano il passaggio degli stimoli esterni all’ipotalamo, le risposte autonomiche e somatiche concomitanti alle emozioni prodotte dagli stessi centri ipotalamici risultano appropriate alle condizioni esterne. La corteccia è anche necessaria per l’esperienza conscia delle emozioni, mentre l’amigdala ha un ruolo chiave nel controllo della comunicazione tra le aree che organizzano l’espressione somatica delle emozioni (ipotalamo e nuclei del tronco encefalico) e le aree che realizzano l’esperienza emotiva conscia.