
RNA
RNA
Introduzione
Fino agli anni Settanta del 20° sec. le funzioni biologiche note dell'acido ribonucleico o RNA (Ribonucleic Acid) erano ristrette alla funzione codificante (RNA messagero o mRNA) e alla sintesi proteica (RNA ribosomale o rRNA, RNA transfer o tRNA) e l'idea prevalente era che il RNA svolgesse un ruolo minimo nella regolazione dell'espressione dei geni. Tuttavia, nelle decadi successive agli anni Settanta è diventato evidente che il RNA è coinvolto in tutti gli aspetti che riguardano il mantenimento, il trasferimento, il processamento e la regolazione dell'informazione genetica.
È stato dimostrato che nella cellula esiste una varietà incredibile di molecole di RNA e che il RNA può svolgere funzioni enzimatiche, dirigere la rimozione degli introni dai trascritti nucleari precursori del mRNA, dirigere il processamento del RNA precursore dei RNA ribosomali, funzionare come guida per gli enzimi che modificano i precursori del rRNA ribosomale o altri piccoli RNA e controllare l'espressione genica sia a livello trascrizionale sia a livello post-trascrizionale. Tutte queste funzioni sono possibili perché il RNA è una biomolecola altamente reattiva con proprietà chimiche uniche, può formare strutture secondarie e terziarie estremamente complesse e riconoscere in maniera specifica sequenze complementari presenti sia in DNA sia in RNA.
Piccoli RNA
Negli organismi eucarioti il RNA comprende una vasta gamma di molecole che in generale vengono classificate a seconda della loro funzione, unità di sedimentazione oppure dimensioni, e localizzazione nella cellula. Dal punto di vista storico, l'espressione piccoli RNA fu originariamente introdotta per indicare molecole di RNA non codificanti di lunghezza compresa fra 50 e 300 nucleotidi. Questa classe di RNA comprende i piccoli RNA nucleari, i piccoli RNA nucleolari, i piccoli RNA citoplasmatici, i piccoli RNA interferenti e i piccoli RNA microscopici.
Piccoli RNA nucleari
Sono fattori essenziali per il processamento dei precursori primari degli mRNA.
Piccoli RNA nucleolari
Funzionano nel processo di taglio del precursore del rRNA o come guida nei processi di modificazione post-trascrizionale di certi nucleotidi dei rRNA o di altri piccoli RNA.
Piccoli RNA citoplasmatici
Comprendono il tRNA, che funziona nella sintesi proteica (RNA 5S e RNA 7S).
Piccoli RNA interferenti e piccoli RNA microscopici
Alla fine degli anni Novanta la scoperta dell'interferenza genetica mediata da RNA a doppio filamento (dsRNA) ha portato all'identificazione di nuove classi di piccoli RNA con una lunghezza compresa fra i 20 e i 30 nucleotidi, i più corti RNA conosciuti negli organismi eucarioti. Questi piccoli RNA sono noti come piccoli RNA interferenti o siRNAs (small interfering RNAs) e piccoli RNA microscopici o miRNAs (microRNAs). In generale questi piccoli RNA riconoscono per complementarità sequenze omologhe presenti in mRNA (o in altri RNA) e servono come guida per indurre la degradazione del RNA bersaglio o per bloccare il processo di sintesi proteica. La scoperta della interferenza genica mediata da questi piccoli RNA ha generato una completa rivoluzione della concezione di come l'espressione dei geni è regolata negli organismi eucarioti. Si calcola che nell'uomo più di un terzo dei geni vengano regolati attraverso i miRNAs e, con buona probabilità, il numero dei geni regolati è ancora più alto. È chiaro che questi RNA hanno sufficiente specificità per funzionare con accuratezza. Non è esagerato affermare che non c'è campo della biologia che non sia stato profondamente cambiato dalla scoperta dei siRNAs e dei miRNAs.
La scoperta della interferenza genica o silenziamento genico mediati da piccoli RNA
Come succede spesso per scoperte scientifiche rivoluzionarie, il concetto e la dimostrazione dell'interferenza genica mediata da piccoli RNA non sono risultati da un unico esperimento ma si sono sviluppati attraverso molteplici osservazioni riportate dagli anni Novanta in poi da parecchi laboratori che utilizzavano organismi completamente diversi. Nel 1990 due gruppi avevano cercato di produrre piante di petunia con fiori di colore viola più scuro del normale e, come strategia, ambedue i gruppi avevano scelto di introdurre copie transgeniche di un enzima il quale determina la formazione del pigmento. L'idea era molto semplice: più geni per l'enzima avrebbero prodotto più enzima ed era logico pensare che la quantità di pigmento sarebbe aumentata risultando in fiori viola scuro. Al contrario, la maggior parte dei fiori che si svilupparono dalle piante transgeniche avevano petali bianchi, quindi senza pigmento. Questa osservazione e altri esperimenti dimostrarono che nella petunia, in presenza di copie transgeniche, la sintesi dell'enzima responsabile per la formazione del pigmento era stata seriamente repressa invece di essere aumentata. Questo fenomeno fu battezzato cosuppression, per indicare che era stata simultaneamente repressa sia l'espressione del gene endogeno sia quella delle copie transgeniche. Risultati simili furono descritti nel fungo Neurospora e il fenomeno fu battezzato quelling. Durante gli anni che seguirono la maggior parte dei tentativi di esprimere geni soprannumerari nelle piante fallirono per il fenomeno di cosuppression. Inoltre, alcuni esperimenti chiave dimostrarono che la cosuppression si verificava solamente quando tanto la sequenza codificante quanto la sequenza non codificante di un gene venivano espresse contemporaneamente. A questo punto cominciò a prendere forma l'idea che la cosuppression dipendesse dalla formazione di RNA a doppio filamento, ma come funzionassero questi dsRNA non era ancora chiaro.
Mentre questi primi esperimenti sulle piante procedevano, molti ricercatori cercavano di reprimere l'espressione genica utilizzando la cosiddetta strategia antisense. Anche questa era un'idea semplice: si pensava che l'espressione di sequenze antisense avrebbe ridotto l'espressione di geni specifici. Questi esperimenti antisense a volte funzionavano, ma nella maggior parte dei casi fallivano. A queste osservazioni si aggiunse, nel 1995, la pubblicazione di un lavoro che riportava come l'espressione genica nel nematode Caenorhabditis elegans potesse essere repressa mediante iniezione di molecole sintetiche di RNA o di tipo sense o antisense.
Dobbiamo ai laboratori di A. Fire e C.C. Mello, ai quali è stato assegnato il premio Nobel per la fisiologia o la medicina nel 2006, l'osservazione che tutte le preparazioni di molecole di RNA a singolo filamento (che si usano comunemente in laboratorio) sono contaminate da basse concentrazioni di dsRNA e la fondamentale scoperta che sono le molecole di dsRNA a iniziare il processo di silenziamento genico. Il fenomeno fu battezzato RNA interference o RNAi. In seguito al lavoro di Fire e Mello il silenziamento dell'espressione genica mediata da dsRNA fu scoperto e utilizzato sperimentalmente in un ampio spettro di organismi eucarioti, dai protozoi all'uomo. L'evidenza che piccoli RNA interferenti, i siRNAs, erano correlati con il fenomeno di cosuppression fu documentata per la prima volta da D. Baulcombe e colleghi nel 1999 nelle piante e, poco tempo dopo, il laboratorio di T. Tuschl dimostrò per la prima volta che siRNAs sintetici erano capaci di indurre RNAi in cellule di mammifero. Questa scoperta è stata fondamentale dal momento che nei mammiferi il dsRNA di lunghezza superiore a 30 nucleotidi provoca una generale distruzione di tutti gli mRNA e le cellule vanno incontro successivamente ad apoptosi. Tutti gli organismi nei quali, come nei mammiferi, i metodi di genetica classica sono di difficile applicazione, hanno tratto benefici enormi dalla scoperta dell'RNAi e dall'inizio del 2000 lo studio della funzione di geni si è sviluppato in maniera esponenziale.
La scoperta degli RNA microscopici
La scoperta dei miRNAs avvenne indipendentemente dai siRNAs e iniziò nel 1993 con l'identificazione di un piccolo RNA di 22 nucleotidi, necessario per il corretto sviluppo temporale delle larve dell'organismo modello Caenorhabditis elegans. Nel genoma umano si calcola che possano essere codificati circa 1000 miRNAs, ma il loro numero è molto probabilmente destinato ad aumentare.
È l'origine e non la funzione a distinguere miRNAs e siRNAs. È importante notare che i miRNAs sono stati identificati solo in piante e animali e nei loro virus, ma non in organismi unicellulari. I siRNAs sono derivati da RNA a doppio filamento (dsRNA) che possono essere lunghi centinaia o forse migliaia di basi. Nelle cellule di piante, nematodi e anche organismi unicellulari, queste lunghe molecole di dsRNA si possono formare come conseguenza della trascrizione di ambedue le eliche del DNA di famiglie di sequenze ripetute. Una volta sintetizzati, questi RNA formerebbero dsRNA attraverso l'appaiamento di regioni di estensiva complementarità. Alternativamente, nei nematodi, nelle piante e nei funghi il dsRNA può essere formato mediante copiatura di RNA a singolo filamento da parte di una specifica RNA polimerasi. Nell'accezione odierna i siRNAs sono perfettamente o estensivamente complementari a sequenze dei trascritti bersaglio e ne guidano la specifica degradazione.
MiRNA. I miRNAs derivano da lunghi transcritti primari (primary-miRNA o pri-miRNA), sintetizzati dalla RNA polimerasi ii, contenenti numerose regioni con strutture secondarie (hairpins, letteralmente forcine per capelli) di circa 70 nucleotidi. Nella Drosophila questi hairpins sono liberati dal pri-miRNA da una endonucleasi nucleare e i prodotti di questa digestione sono chiamati pre-miRNAs. I pre-miRNAs vengono esportati dal nucleo nel citoplasma dove vanno incontro a un secondo step di maturazione da parte dell'enzima Dicer. Quest'ultimo taglia da ambedue le parti della doppia elica dell'hairpin producendo miRNA maturo. I miRNAs possono essere parzialmente o completamente complementari a sequenze codificanti o non codificanti dei mRNA bersaglio. Negli organismi vegetali tutti i miRNAs conosciuti sono del tutto complementari ai loro rispettivi RNA bersagli. È importante notare che alcuni mRNA hanno la potenzialità di legare più di un tipo di miRNA e questa possibilità ha stimolato l'idea che la regolazione mediata dai miRNAs possa avvenire in maniera combinatoria.
SiRNA. I siRNAs possono inibire l'espressione genica a livello post-trascrizionale, come accennato sopra, e anche a livello della trascrizione. Una delle importanti funzioni dell'RNAi è prevenire la trascrizione di questi elementi mobili mediante formazione di eterocromatina o indurre la degradazione dei corrispondenti trascritti attraverso i siRNAs. Negli organismi vegetali, l'RNAi funziona anche come sistema di protezione contro infezioni virali, specificamente infezioni da virus con genoma di RNA a singolo filamento.
La biochimica del silenziamento genico mediato da piccoli RNA
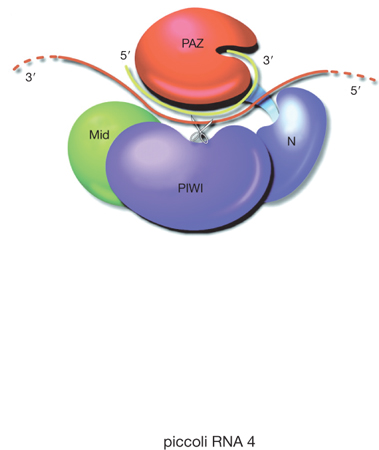
Il meccanismo più chiaro dal punto di vista biochimico è il processo di degradazione di mRNA mediato dai siRNAs (o dai miRNAs perfettamente complementari al RNA bersaglio). Questo processo è stato ricostruito in vitro usando estratti di cellule in coltura o di embrioni di Drosophila. Una volta che i siRNAs sono stati processati da Dicer, si pensa che vengano dissociati nei due filamenti da una attività elicasica e che uno dei due filamenti, il filamento guida, si associ con la proteina Argonaute2 (Ago2). I membri della famiglia delle proteine Argonaute contengono due domini caratteristici: il dominio PAZ e il domino Piwi, oltre ai domini Mid e N-terminale (v. figura). Negli organismi superiori sono presenti molteplici proteine Ago. L'evidenza sperimentale accumulata finora suggerisce che le diverse proteine Ago hanno funzioni specializzate che possono dipendere dal tipo di cellula in cui sono espresse e dal periodo di sviluppo dell'organismo.
La proteina Ago2 dell'uomo e della Drosophila insieme ad altre proteine forma un complesso multiproteico chiamato RISC (RNA-Induced Silencing Complex) che possiede attività endonucleasica ed è capace di degradare in maniera specifica un RNA bersaglio contenente sequenze complementari alla sequenza guida del siRNA.
Le ribonucleoproteine Ago svolgono funzioni essenziali nel processo di silenziamento attraverso siRNAs e miRNAs. Sia i siRNAs sia i miRNAs cominciano la loro esistenza come molecole a doppio filamento, ma esercitano la loro funzione come molecole a singolo filamento. La trasformazione da doppio a singolo filamento avviene durante il trasferimento di queste molecole a proteine della famiglia Argonaute, con l'aiuto di altre proteine che contengono domini di riconoscimento per dsRNA.
bibliografia
A.R. van der Krol, L.A. Mur, M. Beld et al., Flavonoid genes in petunia: additino of a limited number of gene copies may lead to a suppression of gene expression, in The plant cell, 1990, 291, 2.
C. Napoli, C. Lemieux, R.A. Jorgesen, Introduction of a chimeric chalcone synthase gene into petunia results in reversibile co-suppression of homologus genes in trans, in The plant cell, 1990, 2, pp. 279-89.
R.C. Lee, R.L. Feinbaum, V. Ambros, The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarità to lin-14, in Cell, 1993, 75, pp. 843-54.
S. Guo, K.J. Kemphues, Par-1, a gene required for establishing polarità in C. elegans embryos, encodes a putative Ser/Thr kinase taht is asymmetrically distributed, in Cell, 1995, 81, pp. 611-20.
A. Fire, M.K. Montgomery, S.A. Kostas et al., Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans, in Nature, 1998, 391, pp. 806-11.
A.J. Hamilton, D.C. Baulcombe, A species of small antisense RNA in post-transcriptional gene silencing in plants, in Science, 1999, 286, pp. 950-52.
S.M. Hammond, E. Bernstein, D. Beach et al., An RNA-directed nucleare mediates post-transcriptional gene silencing in Drosophila cells, in Nature, 2000, 404, pp. 293-96.
S.M. Elbashir, M.J. Harborth, W. Lendeckel et al., Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells, in Nature, 2001, 411, pp. 494-98.
J. Liu, M.A. Carmell, F.V. Rivas et al., Argonaute2 is the catalytic engine of mammalian RNAi, in Science, 2004, 305, pp. 1437-41.