
Radicali liberi
Radicali liberi
Il significato di r. l. in biologia è uno dei concetti chiave per la comprensione non solo dei danni ossidativi prodotti dai radicali stessi, ma di tutte le funzioni degli organismi che utilizzano l'ossigeno molecolare per la produzione di energia (aerobiosi). Mentre il danno ossidativo da r. l. implica modifiche chimiche irreversibili del bersaglio molecolare colpito e indiscriminate rispetto al tipo di molecola e di gruppo chimico modificati, gli effetti di natura fisiologica si esplicano attraverso alterazioni chimiche reversibili e selettive e, pertanto, potenzialmente soggette a regolazione metabolica.
Nuovi radicali liberi con nuovi effetti biologici
In una prima serie di ricerche erano considerati r. l. di importanza biologica solamente i prodotti intermedi della riduzione dell'ossigeno nei vari distretti della cellula (superossido, perossido di idrogeno e radicale ossidrile) e i perossidi derivanti dall'addizione dell'ossigeno ai radicali degli acidi grassi delle membrane. Queste specie sono correntemente indicate con l'acronimo ROS (Reactive Oxygen Species, specie reattive dell'ossigeno). A esse si è poi aggiunta una nuova famiglia di specie radicaliche caratterizzate dalla presenza, oltre che dell'ossigeno, di un atomo d'azoto, e pertanto indicate come RNS (Reactive Nitrogen Species, specie reattive dell'azoto). Il capostipite di questi radicali è l'ossido d'azoto o nitrico (NO·), che nei sistemi biologici si genera per ossidazione di uno degli azoti del gruppo guanidinico dell'aminoacido arginina, catalizzata dall'enzima sintetasi dell'ossido nitrico o NO-sintasi (NOS). ROS e RNS interagiscono fra loro in vario modo: per es., la NOS è attivata da dosi fisiologiche di perossido d'idrogeno e l'interazione diretta di superossido e NO· è all'origine di una specie generatrice di danno ossidativo, il perossinitrito. Le ricerche hanno però mostrato che la maggior parte degli effetti dei RNS si esplica a livello della regolazione fisiologica delle funzioni dell'organismo. Nel 1998 il premio Nobel per la fisiologia o la medicina è stato assegnato agli scienziati statunitensi R.F. Furchgott, L.J. Ignarro e F. Murad per le loro ricerche sul ruolo dell'ossido nitrico come mediatore di segnali fisiologici nel sistema cardiovascolare, in particolare quelli che regolano la vasodilatazione per effetto del rilassamento della muscolatura liscia. Queste proprietà dell'ossido di azoto sono alla base dell'attività vasodilatante della nitroglicerina, scoperta dall'italiano A. Sobrero nel 1847, insieme alla sua azione esplosiva che sarà sfruttata pochi anni dopo da A. Nobel per la preparazione della dinamite. Le conseguenze degli studi sulle proprietà biologiche di questo r. l. è stata la scoperta dei meccanismi molecolari dell'erezione del pene e la successiva diffusione di farmaci oggi molto popolari per la cura dell'impotenza maschile. Un'altra specie radicalica della quale non si conoscevano finora effetti biologici è il radicale carbonato, CO3·−−. Si è visto che esso è molto importante nel meccanismo dell'attività aggiuntiva dell'enzima superossido dismutasi a rame zinco (SOD1) che si esplica in alcune situazioni patologiche. Questo enzima è la difesa principale della cellula verso i ROS perché elimina il radicale primario dell'ossigeno, l'O2·− o anione superossido, mediante una reazione di dismutazione che lo trasforma in perossido d'idrogeno, H2O2. Questo prodotto di reazione, se non decomposto dagli altri enzimi antiossidanti catalasi o glutatione perossidasi genera radicali ossidrile al sito attivo dell'enzima, inattivandolo. In presenza di bicarbonato, si formano radicali carbonato, e questa reazione risparmia il centro attivo dell'enzima ma produce danno ossidativo su altri bersagli biologici.
Queste reazioni configurano un'attività proossidante di un enzima antiossidante, e sono rilevanti, per es., quando esistono delle varianti genetiche della SOD1 che, a parità di attività superossido dismutasica, hanno una conformazione del sito attivo che favorisce la reazione con il perossido di idrogeno (v. oltre: Malattie neurodegenerative).
Nuove funzioni biologiche per i ROS
Il caso dell'ossido nitrico con la sua duplicità di effetti, danno ossidativo e regolazione fisiologica, ha stimolato nuovi studi che hanno dimostrato che anche i ROS possono intervenire come mediatori fisiologici di molti processi funzionali.
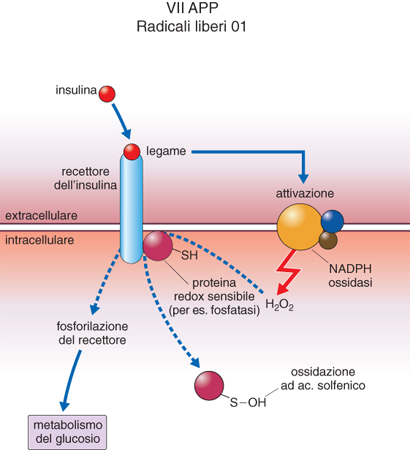
Ruolo dei radicali liberi nella transduzione di segnali biologici. - I cosiddetti recettori di membrana trasferiscono all'interno della cellula il segnale chimico che arriva dall'esterno veicolato da ormoni, fattori di crescita e altre molecole bioattive, quali farmaci e componenti alimentari, che non sono in grado di attraversare la membrana cellulare (transduzione del segnale). Un caso tipico riguarda l'insulina, l'ormone essenziale al metabolismo cellulare del glucosio (fig. 1). Il legame dell'insulina al suo recettore produce l'attivazione di un enzima di membrana (NADPH ossidasi) simile a quello che si attiva sulla membrana dei fagociti nella risposta infiammatoria, capace di produrre perossido di idrogeno dall'ossidazione di un coenzima ridotto (NADPH), che si genera esclusivamente per ossidazione diretta del glucosio e quindi fa da rivelatore della sua concentrazione nei tessuti. È questo flusso di perossido che ossida reversibilmente specifiche cisteine di proteine incorporate nella parte intracellulare del recettore. In questo stato di blanda ossidazione il recettore aumenta la sua affinità per i gruppi fosforici, e nella sua forma fosforilata può attivare enzimi della cellula cruciali per il metabolismo del glucosio che non avrebbe 'incontrato e riconosciuto' nella sua forma priva di fosforo. Questo tandem ROS-fosforilazione, mediato da ossidazione blanda delle cisteine di proteine specializzate, opera in molti altri eventi di attivazione recettoriale ed è un caso particolare del fenomeno più generale della regolazione delle chinasi da parte dei ROS. Le chinasi sono gli enzimi che catalizzano il trasporto di gruppi fosforici dal donatore universale di fosforo adenosintrifosfato (ATP) a determinate proteine, modulandone l'attività positivamente o negativamente. Molte chinasi sono preposte alla regolazione del ciclo cellulare, per es., le chinasi regolate dai mitogeni o MAPK (Mytogen Activated Protein Kinase). Ora sappiamo che intervalli molto stretti nel flusso di ROS e RNS possono indirizzare il ciclo cellulare verso la sua attivazione o il suo arresto attraverso la modificazione ossidativa di cisteine delle proteine che si interpongono tra la fonte primaria di radicali e le MAPK.
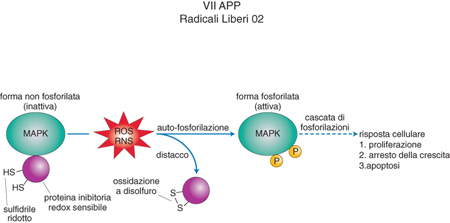
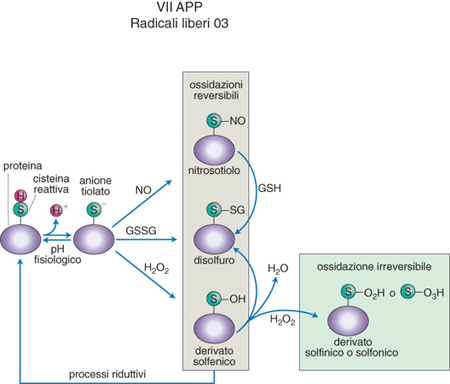
Queste reazioni sono quindi molto importanti nel determinare l'accelerazione della riproduzione cellulare (proliferazione), come nello sviluppo degli organismi e nella carcinogenesi, o per converso il suo rallentamento, come nell'invecchiamento e nella apoptosi o morte cellulare programmata (fig. 2). Anche altre chinasi sono soggette a regolazione ossidoriduttiva, che può avvenire non solo per ossidazione di loro cisteine specifiche da parte dei ROS, ma anche, al contrario, per effetto del legame di molecole antiradicaliche come le vitamine A ed E, che impediscono queste ossidazioni. Se lo stimolo ossidativo proveniente dall'ambiente si limitasse alla formazione di ROS e RNS, sarebbe inefficace come regolatore dei processi fisiologici degli organismi, perché queste specie hanno nella cellula una vita molto breve a causa della loro instabilità e della loro reattività indiscriminata con le molecole circostanti. Invece, il potenziale ossidativo dei ROS, quando prodotti in concentrazioni fisiologiche, è fissato su molecole che nello stato ossidato sono relativamente stabili e ripristinano il loro stato ridotto solo mediante reazioni dotate di alta specificità. In questi processi il perossido di idrogeno e l'ossido nitrico sono più adatti dei loro derivati radicale ossidrile e perossinitrito, responsabili di reazioni ossidanti più potenti, indiscriminate e irreversibili. Il sistema chimico che l'evoluzione molecolare ha selezionato come bersaglio ideale di queste ossidazioni blande è stato già accennato nel caso delle chinasi ed è costituito da composti contenenti, nel loro stato ridotto, il gruppo sulfidrilico o tiolico (-SH) dell'aminoacido cisteina (fig. 3). Per azione del perossido di idrogeno esso diventa gruppo solfenico (-SOH) o disolfuro (-S-S) mentre per effetto dell'ossido nitrico si trasforma in gruppo nitrosotiolico (SNO). Queste reazioni avvengono o come modificazioni postgenetiche di cisteine di proteine specificamente deputate al trasporto del segnale innescato dai r. l. o, in modo più ubiquitario e quantitativamente più rilevante, al livello della cisteina del tripeptide glutatione (acido glutammico, cisteina e glicina, siglato come GSH nello stato tiolico e GSSG nello stato più comune di blanda ossidazione, quello disolfurico). In virtù della sua abbondanza il glutatione è il vero tampone delle reazioni radicaliche nella cellula perché i r. l. in eccesso reagiscono con il GSH trasformandolo nella sua forma radicalica, e due radicali di GSH reagiscono poi generando GSSG. Quest'ultimo può propagare il segnale radicalico mediante una successione di reazioni tiolo-disolfuro a livello di proteine selezionate all'interno di un determinato processo regolatorio, come, per es., nel caso della transduzione del segnale e nell'attivazione genica.
Radicali liberi e geni. - La difesa degli organismi contro il danno ossidativo è basata essenzialmente sull'espressione, costitutiva o induttiva, di geni che regolano la sintesi di molecole antiradicaliche o mediano l'azione fisiologica dei radicali stessi. Questo equivale a dire che le reazioni radicaliche intervengono sia a valle, sulle proteine terminatrici del segnale, sia a monte, sui dispositivi molecolari dell'inizio del segnale a livello genetico. Questi dispositivi sono anch'essi proteine, denominate fattori di trascrizione, perché, legandosi a zone specifiche del DNA dei geni (promotori) innescano la sintesi del RNA complementare (trascrizione) che detterà la sintesi della proteina corrispondente (traduzione). Nei Procarioti i r. l. hanno effetti diretti sul gene perché i fattori di trascrizione contengono essi stessi un elemento ROS-sensibile, come gruppi di cisteine ridotte o centri ferro-zolfo, costituiti da un atomo di ferro al centro di un complesso di zolfi sulfidrilici. Nei batteri è stato individuato un sistema di risposta allo stress ossidativo che comprende una proteina regolatoria, OxyR, la quale, quando le sue cisteine sono ossidate da bassi livelli di ROS ad acido sulfenico e a disolfuro, favorisce l'espressione dei geni di numerose molecole antiradicaliche. Negli Eucarioti l'espressione genica è più spesso controllata dai ROS in maniera indiretta e complessa, a causa dei diversi compartimenti cellulari che concorrono alle fasi successive di induzione e di esecuzione del processo di attivazione dei geni, e che possono avere omeostasi ossidoriduttive differenti. Per es., molti fattori regolatori della trascrizione dei geni sono attivati nel citoplasma mediante fosforilazione da parte di chinasi a loro volta attivate dai ROS, ma l'evento che termina il processo, cioè il legame del fattore al DNA per iniziare la trascrizione richiede, al contrario che nel caso esaminato sopra per i Procarioti, condizioni riducenti, assicurate nel nucleo da alte concentrazioni di GSH e di una proteina ricca di tioli, la tioredossina. La necessità di regolare in modo accurato il flusso di r. l. per garantire questa doppia esigenza di condizioni proossidanti e antiossidanti è alla base dell'esposizione degli organismi superiori a rischi di squilibrio ossidoriduttivo che si traducono nelle varie situazioni patologiche collegate ai radicali liberi.
Dalla patologia da radicali liberi a una medicina molecolare della regolazione ossidoriduttiva
Le conoscenze acquisite sul ruolo dei radicali in tutti gli stadi di generazione e trasferimento dei segnali biologici di regolazione, ha permesso di rivisitare la patologia da radicale libero alla ricerca del coinvolgimento radicalico nei meccanismi patogenetici più intimi al di là della mera constatazione di un danno terminale ipoteticamente di natura ossidativa. Questo ha anche aperto nuovi orizzonti alle prospettive terapeutiche. Tutta la medicina, nei suoi aspetti molecolari, è stata coinvolta in questa transizione, ma alcuni campi hanno assistito a progressi più significativi.
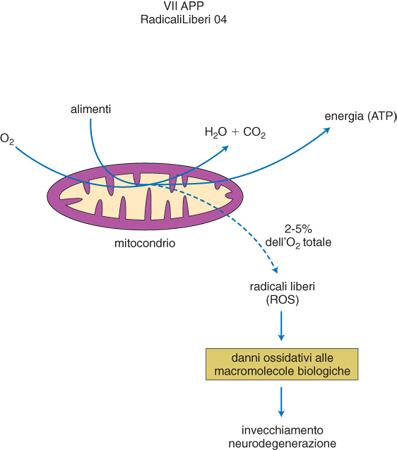
Invecchiamento. - L'invecchiamento da r. l., cioè l'ipotesi semplicistica che la senescenza è data dall'accumulo di danni cellulari causati dai ROS prodotti durante il metabolismo dell'ossigeno, è quasi un luogo comune da quando nel 1956 D. Hartman la propose per primo, indicandola, nel contesto delle conoscenze del tempo, come "una teoria basata sulla chimica dei r. l. e delle radiazioni". Ora, però, disponiamo di dati meno descrittivi e fenomenologici, basati sulla genetica. Sono stati ottenuti animali o cellule transgenici nei quali l'alterazione dell'espressione genica di un enzima antiossidante modifica la lunghezza della vita del sistema biologico trasformato. Per quanto riguarda il meccanismo alla base di questi effetti, i dati si sono focalizzati sul mitocondrio (fig. 4) come sede principale della connessione fra r. l. e invecchiamento, non solo come sede preponderante della produzione di ROS nella cellula ma anche come bersaglio di scelta da parte dei r. l. nei processi di senescenza. Si è potuto vedere che la morte cellulare programmata o apoptosi, processo molto importante nella senescenza dei tessuti, è in gran parte connessa con la liberazione nel citoplasma di fattori mitocondriali, a sua volta resa possibile dalla perdita d'integrità della membrana dell'organello per azione dei radicali liberi. I ROS possono essere prodotti in eccesso dal mitocondrio stesso non solo in relazione a situazioni patologiche ma anche in seguito all'azione di molecole che segnalano la scadenza, programmata geneticamente, del tempo assegnato alla vita di un determinato sistema biologico. Un locus genico implicato in segnali di questo tipo è stato individuato mediante mutazioni mirate del gene shc: la soppressione della sintesi di un suo prodotto proteico, denominato p66 dal suo peso molecolare, ha conferito ai topi trattati minore incidenza di malattie collegate all'invecchiamento, come, per es., l'aterosclerosi, e un significativo prolungamento di vita. È stato quindi scoperto che p66 è un enzima ossidoriduttivo che, una volta attivato da segnali proapoptotici di tipo ossidativo che modulano il suo grado di fosforilazione, è in grado di deviare elettroni dal loro flusso normale nella membrana mitocondriale verso la produzione di perossido d'idrogeno, con conseguente stress ossidativo, senescenza e morte del sistema interessato.
Malattie neurodegenerative. - La malattia di Alzheimer e la malattia di Parkinson insorgono in tarda età in zone specifiche del cervello che, ai rilievi autoptici, mostrano segni di danno ossidativo e appaiono come casi locali di invecchiamento in un tessuto, come quello nervoso, particolarmente suscettibile al danno ossidativo per l'intenso metabolismo aerobio e la relativa carenza di difese antiossidanti. Nell'Alzheimer, il legame improprio di alcuni ioni metallici a una determinata proteina neuronale potrebbe dar luogo a reazioni monovalenti con l'ossigeno (autossidazione) che innescano la catena radicalica dei ROS. Nel Parkinson l'autossidazione riguarda i metaboliti della dopamina, neurotrasmettitore che si libera in alte concentrazioni proprio nelle aree cerebrali interessate dalla malattia. Ma è nello studio della sclerosi laterale amiotrofica (SLA) che la medicina molecolare dei fenomeni ossidoriduttivi ha fornito gli spunti più interessanti e ancora una volta questi progressi sono avvenuti per il contributo della genetica. La SLA è una paralisi progressiva che insorge in genere in età matura e che porta inesorabilmente alla morte in pochi anni per degenerazione specifica delle cellule nervose che mandano gli impulsi per la contrazione muscolare (neuroni motori o motoneuroni). Si è visto che in una frazione molto ridotta di casi di SLA, con incidenza familiare ben definita, sono presenti mutazioni del gene della SOD1, che alterano la funzione dell'enzima prodotto, o nel senso di diminuirla o nel senso di affiancarla a una nuova attività che, invece di essere antiossidante, è di tipo perossidasico e perciò proossidante. In ogni caso si ha un aumento di ROS che porta alla morte dei motoneuroni. I casi, molto più numerosi, di SLA privi di questa mutazione (forma sporadica), sono indistinguibili dalle forme familiari per sintomi, decorso e reperti autoptici. Si può ipotizzare che nei casi sporadici intervenga un altro fattore che comunque aumenti il flusso dei ROS nei motoneuroni. La comparsa di questa malattia, tipica dell'età intorno ai 50 anni, in giovani che esercitano attività sportive di tipo agonistico spesso accompagnate da abuso di farmaci, è in linea con questa ipotesi. In Italia, un numero significativo di casi è stato diagnosticato in calciatori a fine carriera.
Malattie neoplastiche. - Negli stati trattati precedentemente la bilancia ossidoriduttiva è spostata verso condizioni proossidanti che favoriscono la degenerazione e la morte cellulari. La crescita di un tumore è invece marcata da una tendenza incontrollata alla proliferazione di determinate cellule. In realtà nel cancro i r. l. hanno un ruolo differente a seconda che consideriamo le diverse fasi del processo tumorale, la carcinogenesi (fase di trasformazione), lo sviluppo del tumore (fase di progressione) o la terapia. Mentre nella prima fase domina il ruolo mutageno dei ROS (è assodato il rapporto fra cancro e presenza nell'ambiente di condizioni note per la loro capacità di produrre radicali, come l'inquinamento chimico o quello radioattivo), per lo sviluppo del cancro sono determinanti condizioni locali che favoriscono la proliferazione cellulare, cioè, in genere, uno stato ridotto degli antiossidanti tessutali. Al momento della terapia si cerca invece di produrre flussi radicalici imponenti a livello della neoplasia, con trattamenti radio- oppure chemioterapici, sfruttando la più alta sensibilità delle cellule tumorali ai r. l. per il maggiore ritmo riproduttivo e la minore presenza di antiossidanti tipica delle cellule poco differenziate.
Alimentazione ed esercizio fisico. - Gran parte delle calorie introdotte con gli alimenti costituiscono il potenziale di riduzione dell'ossigeno nel mitocondrio, al fine di produrre energia sotto forma di ATP. Il 2-5% dell'ossigeno consumato in questo processo dà origine in condizioni fisiologiche a superossido e agli altri ROS (fig. 4). Quindi, quanto maggiore è il contenuto calorico di una dieta, tanto più elevato è il flusso di ROS dai mitocondri. Poiché si è visto (v. sopra) che la produzione mitocondriale di ROS è correlata ai processi di invecchiamento, ci si è chiesto se la riduzione di calorie assunte durante la vita di un individuo (restrizione calorica) possa portare a un allungamento della vita. Le ricerche su animali da esperimento hanno dimostrato che questa relazione esiste ed è associata a ridotto danno ossidativo degli enzimi mitocondriali coinvolti nella riduzione dell'ossigeno da parte dei derivati metabolici di componenti alimentari ricchi di energia. Oltre a questa rilevanza dei ROS nella quantità globale di apporto alimentare, sono oggetto di studi molto intensi le specifiche attività pro- o anti ROS dei vari componenti della dieta, sia naturali sia aggiunti (additivi e integratori alimentari). Molte vitamine (soprattutto A, C ed E) neutralizzano direttamente i ROS, mentre alcuni ioni minerali (rame e ferro in particolare) sono coenzimi essenziali di enzimi che neutralizzano i ROS come la superossido dismutasi e la catalasi. Oltre che di questi nutrienti indispensabili i vegetali sono ricchi di sostanze che, pur non essendo necessarie al nostro metabolismo hanno ugualmente un'azione positiva nella prevenzione delle malattie collegate ai r. l. proprio per la loro elevata capacità antiossidante: i polifenoli, composti che conferiscono ai vegetali colori molto intensi e sono molto abbondanti soprattutto in bevande come il tè e il vino rosso. La necessità di preservare le proprietà antiossidanti degli alimenti nelle condizioni di distribuzione dilatata nel tempo e nello spazio tipiche delle società moderne ha fatto diffondere l'uso di additivi chimici ad azione antiradicalica. È inoltre sempre più diffusa l'abitudine di surrogare la capacità antiossidante di una alimentazione naturale variata e bilanciata con l'assunzione di vitamine e minerali in compresse o in estratti concentrati. Si deve però tener presente che un antiossidante è tale in quanto dotato di capacità riducenti e quindi la sua assunzione al di fuori delle sinergie e delle compensazioni presenti in un alimento o in una dieta può portare a interferenze con le azioni fisiologiche dei r. l. o a effetti proossidanti solo apparentemente paradossali, perché un riducente in concentrazione eccessiva tende ad autossidarsi in presenza di ossigeno, innescando a sua volta produzione di ROS.
L'esercizio fisico è collegato all'alimentazione in quanto consuma nel muscolo l'ATP derivato dalle calorie alimentari. In questo senso ha un'azione protettiva analoga alla restrizione calorica. Sforzi eccessivi portano però ad aumentata produzione di ROS sia per l'aumento del consumo di ossigeno sia per i ripetuti cicli di ipossia/riossigenazione che si hanno soprattutto negli sport anaerobi e che riproducono una situazione, a livello di tutto l'organismo, paragonabile a quella che si crea localmente nell'infarto cardiaco. Queste complicazioni sono prevenute dall'allenamento che ha, fra i suoi molti effetti, quello di indurre l'espressione dei geni degli enzimi antiossidanti. Un particolare tipo di esercizio fisico è quello che si effettua in alta montagna, dove la pressione parziale di ossigeno diminuisce fino ad arrivare, sulla vetta del monte Everest, a un terzo di quella presente a bassa quota. È stato dimostrato che nelle condizioni più spinte il consumo di ossigeno produce per il 50% r. l. (contro il 2-5% fisiologico), a causa della drastica riduzione e inefficienza del metabolismo dei mitocondri e dell'alternanza di fasi ipossiche durante lo sforzo e di riossigenazione durante il riposo. Gli adattamenti molecolari delle popolazioni acclimatate all'altitudine comprendono un maggiore livello di espressione genica degli enzimi antiossidanti, e questo si riscontra anche negli individui viventi a quote inferiori dopo un congruo periodo di allenamento in altura.
bibliografia
W. Droege, Free radicals in the physiological control of cell functions, in Physiological review, 2002, 82, 1, pp. 47-95.
G. Filomeni, G. Rotilio, M.R. Ciriolo, Cell signalling and the glutathione redox system, in Biochemical pharmacology, 2002, 64, 5-6, pp. 1057-64.
G. Filomeni, G. Rotilio, M.R. Ciriolo, Disulfide relays and phosphorylative cascades: partners in redox-mediated signaling pathways, in Cell death and differentiation, 2005, 12, pp. 1555-63.