
plasticita neurale
plasticità neurale
Un concetto fondamentale della neurobiologia moderna è che le connessioni del sistema nervoso possono essere modificate dall’esperienza, sia in termini funzionali (variazione delle quantità di neurotrasmettitore rilasciato), sia in termini di struttura (espansione o retrazione delle connessioni). Tali processi sono noti come fenomeni di plasticità neurale e rappresentano una proprietà caratteristica, ma non esclusiva, della corteccia cerebrale. Il nostro cervello è continuamente soggetto al flusso della stimolazione ambientale, che influisce sulle nostre capacità di percepire gli stimoli, compiere movimenti, pensare, apprendere, ricordare e pianificare strategie comportamentali. Quando diciamo «Ho cambiato idea» asseriamo senza saperlo che qualcosa è cambiato nella funzione o nella struttura del nostro cervello. Questo perché le idee sono, per così dire, ‘stampate’ nei circuiti nervosi e non si può cambiare idea se non cambiando il ‘testo’ cerebrale che la descrive. La plasticità è una caratteristica peculiare del sistema nervoso in sviluppo; con il passaggio all’età adulta si verifica una notevole riduzione delle potenzialità plastiche dei circuiti nervosi, anche se una serie di studi ha evidenziato che un’adeguata stimolazione ambientale è in grado di indurre fenomeni di plasticità anche nel cervello adulto. [➔ ambiente e cervello; apprendimento; depressione a lungo termine; neurotrofine; periodi critici; potenziamento a lungo termine; visione] «Immagina un uomo adulto e cieco dalla nascita e che questi abbia imparato a distinguere un cubo da una sfera […] mediante il tatto. Supponi poi che […] il cieco recuperi la vista. La questione è se, servendosi della sola vista e prima di toccarli, egli saprebbe distinguere la sfera dal cubo» (John Locke, An essay concerning human understanding, 1690). Questa riflessione, nota come il problema di Molyneux, rappresenta una delle prime testimonianze di un dibattito che ha attraversato la storia della filosofia della scienza e che si è focalizzato sul contributo del patrimonio genetico e dell’esperienza nello sviluppo del sistema nervoso e del comportamento. Oggi sappiamo che i geni guidano le prime fasi dello sviluppo cerebrale e la formazione iniziale delle connessioni neurali (➔ cervello, sviluppo del), mentre le interazioni con l’ambiente sono fondamentali per completare in maniera appropriata e individuo-specifica la maturazione dei circuiti deputati al controllo della maggior parte delle funzioni cerebrali. Più specificamente, i processi della p. n. sono alla base di un fenomeno di selezione che prevede il rafforzamento di alcune connessioni neurali e l’eliminazione di altre, determinando la cablatura definitiva dei circuiti del sistema nervoso. La finestra temporale in cui il cervello è molto ricettivo relativamente alle informazioni provenienti dal mondo esterno e in cui l’esperienza gioca un ruolo essenziale nel piano di sviluppo è nota come periodo critico (➔). Si consideri, però, che la maturazione di funzioni cerebrali differenti avviene in tempi diversi: non esiste, quindi, un unico periodo critico nello sviluppo del cervello, bensì un periodo critico diverso (in termini di momento di inizio e di durata) per ogni funzione. I cospicui livelli di p. n. che sono funzionali allo sviluppo dei circuiti nervosi si riflettono in considerevoli capacità di apprendimento e memoria: è ben noto, infatti, che durante l’infanzia e l’adolescenza si ha una facilità ad apprendere e ricordare, per es., una serie di nozioni o avvenimenti, una sequenza di movimenti o una seconda lingua, che non ha eguali durante la vita adulta. Alterazioni dell’input ambientale durante queste fasi, tuttavia, possono determinare l’instaurarsi di processi di p. n. ‘maladattativa’, che sfociano in modificazioni permanenti dell’organizzazione dei circuiti neurali e nella conseguente insorgenza di condizioni patologiche. A titolo esemplificativo, consideriamo quanto accade nei bambini affetti da cataratta congenita bilaterale (ovvero a carico di entrambi gli occhi): la mancanza di un’adeguata stimolazione visiva, dovuta all’opacizzazione del cristallino, impedisce la maturazione di diverse proprietà del sistema visivo e se la cataratta non viene rimossa tempestivamente (ovvero entro le prime settimane di vita) causa la compromissione irreversibile della percezione visiva. Altrettanto invalidante è la patologia che si instaura a causa di un’alterazione dell’esperienza sensoriale a carico di un solo occhio (per es., dovuta a difetti di rifrazione o cataratta), nota come ambliopia (➔).
Modello del sistema visivo
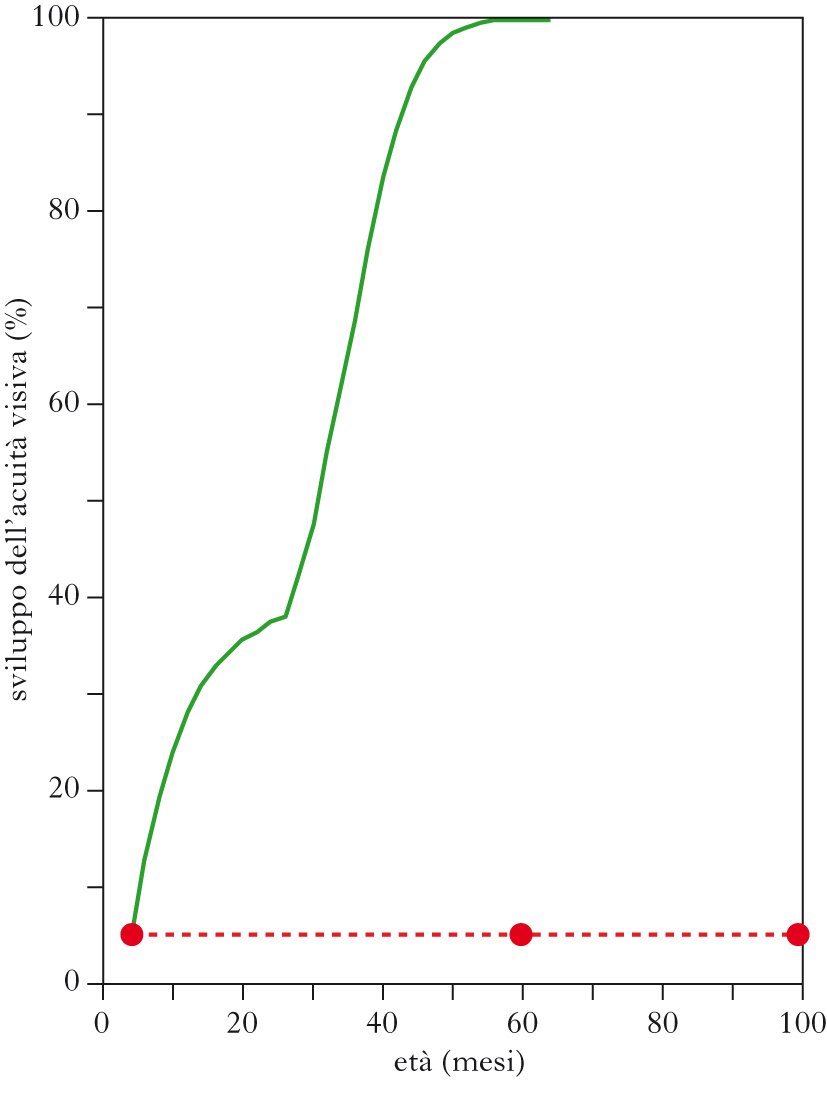
Il sistema visivo è storicamente il terreno d’elezione per l’indagine sui meccanismi alla base dei fenomeni di p. n., dal momento che l’esperienza visiva può essere facilmente manipolata e le conseguenze di tali manipolazioni possono essere misurate a livello fisiologico, anatomico e molecolare (➔ visione). In particolare, è stato ampiamente studiato, come paradigma sperimentale di p. n. nei modelli animali, il protocollo della deprivazione monoculare, che consiste nell’occlusione di un occhio tramite sutura delle palpebre con conseguente sbilanciamento nella stimolazione sensoriale che afferisce ai due occhi. Gli esperimenti classici di David H. Hubel e Torsten Wiesel hanno dimostrato che la deprivazione monoculare determina un indebolimento irreversibile delle facoltà visive dell’occhio chiuso (ambliopia) e un drammatico cambiamento dell’organizzazione funzionale della corteccia visiva primaria. L’occhio deprivato, infatti, perde gran parte della sua capacità di attivare la corteccia visiva e questo si riflette in una riduzione del numero di neuroni che rispondono preferenzialmente alla stimolazione di quell’occhio o parimenti a entrambi gli occhi (neuroni binoculari), associata a un aumento delle cellule attivate dall’occhio rimasto aperto. Questo fenomeno prende il nome di plasticità di dominanza oculare ed è stato riscontrato in tutti i mammiferi in cui sono stati indagati gli effetti della deprivazione monoculare. Come correlato anatomico, si osservano un’alterazione delle connessioni orizzontali della corteccia visiva primaria seguita da una contrazione delle regioni a cui afferiscono i terminali assonici del nucleo genicolato laterale che trasmettono gli stimoli provenienti dall’occhio chiuso (dovuta ad atrofia di questi ultimi) e l’espansione delle aree di competenza delle fibre che veicolano le informazioni dall’occhio aperto. L’efficacia della deprivazione monoculare è ristretta a un periodo critico, che è di lunghezza diversa a seconda della specie studiata, ma che invariabilmente si colloca in una finestra dello sviluppo del sistema visivo. Se si sottopone a deprivazione monoculare un individuo dopo la fine del periodo critico o in età adulta, non si osserva nessuna perdita funzionale dell’occhio deprivato né modificazioni dell’organizzazione della corteccia visiva.
Meccanismi d’azione della plasticità
Gli effetti della deprivazione sensoriale si possono spiegare in base ai termini del principio di Hebb: questo modello di modificabilità delle sinapsi prevede che il rafforzamento delle connessioni sia dovuto alla coincidenza temporale tra le attività di scarica del neurone presinaptico e di quello postsinaptico. Le fibre provenienti dall’occhio aperto hanno un’attività ricca, in quanto modulata dall’esperienza visiva, che depolarizza efficacemente i neuroni corticali, mentre le fibre guidate dall’occhio deprivato hanno un’attività molto ridotta (essenzialmente costituita dall’attività spontanea della retina e del talamo), che risulta scorrelata da quella dell’altro gruppo e non è sufficiente a oltrepassare la soglia di attivazione della corteccia. Questo fa sì che un’appropriata coincidenza temporale tra l’attività afferente e quella corticale abbia luogo solo a livello delle sinapsi che veicolano l’informazione in ingresso dall’occhio aperto, determinando il conseguente potenziamento di queste ultime a scapito di quelle dell’occhio deprivato. È stato suggerito il coinvolgimento nella plasticità corticale dei fenomeni di plasticità sinaptica che vengono definiti potenziamento a lungo termine (➔) e depressione a lungo termine (➔); questo punto, tuttavia, è ancora molto dibattuto. Date le sue peculiari caratteristiche, il recettore per il glutammato di tipo NMDA (N-metil-D-aspartato) gioca un ruolo chiave per rilevare la correlazione tra attività presinaptica e attività postsinaptica: il blocco farmacologico di questi recettori, infatti, impedisce l’instaurarsi degli effetti della deprivazione monoculare. Tuttavia, anche la trasmissione sinaptica GABAergica è determinante per l’induzione dei fenomeni di p. n. attraverso la modulazione del pattern di attività dei circuiti corticali: è stato dimostrato che la maturazione dei circuiti inibitori durante lo sviluppo della corteccia visiva è cruciale per l’apertura del periodo critico di plasticità. Alcuni esperimenti, inoltre, hanno suggerito che i sistemi di neuromodulazione (➔) a proiezione diffusa del tronco dell’encefalo, in partic. quelli dell’acetilcolina, della noradrenalina e della serotonina, esercitino un ruolo di facilitazione sulla plasticità corticale. Una volta innescati dall’attività elettrica, i processi di p. n. si attuano tramite l’attivazione di una serie di proteine intracellulari, tra cui le proteine chinasi PKA (proteina chinasi A), ERK (proteina chinasi regolata da segnali extracellulari) e CAMKII (proteina chinasi calcio-calmodulina dipendente II), che possono regolare localmente l’inserzione di nuovi recettori nella membrana dei neuroni postsinaptici o determinarne la rimozione, causando rispettivamente un rapido aumento o una riduzione della risposta sinaptica. L’attivazione della trascrizione genica e la sintesi di nuove proteine sono necessarie per rendere permanenti le modificazioni dell’efficacia sinaptica e per realizzare i cambiamenti morfologici alla base dei fenomeni di plasticità. Questa riorganizzazione anatomica è accompagnata dall’induzione dell’attività proteolitica di alcuni enzimi, finalizzata a degradare alcune componenti della matrice extracellulare, che, esercitando la loro funzione di supporto e ancoraggio dei neuroni, fungono da fattori inibitori per la dinamicità sinaptica.
Ruolo delle neurotrofine. Una questione cruciale emersa da numerosi esperimenti è che la deprivazione monoculare gioca su un meccanismo di competizione tra i due occhi. La sutura binoculare delle palpebre, infatti, non determina drammatiche modificazioni dell’organizzazione funzionale e anatomica della corteccia visiva. È stato suggerito che le fibre provenienti dai due occhi competano per un fattore neurotrofico, prodotto dai neuroni corticali. Questa ipotesi neurotrofica (➔ cervello, sviluppo del) prevede che le neurotrofine siano sintetizzate e rilasciate dai tessuti bersaglio in maniera attività-dipendente, siano catturate dai terminali nervosi più attivi e trasportate per via retrograda lungo gli assoni fino al corpo cellulare, dove esplicano il loro effetto in termini di sopravvivenza e rafforzamento delle connessioni sinaptiche a livello sia funzionale sia morfologico. Il trasporto dei fattori neurotrofici, tuttavia, si verifica non solo in modo retrogrado, ma anche in senso anterogrado (dal corpo cellulare ad assoni e dendriti) ed è stato recentemente suggerito un ruolo per un’azione anterograda del BDNF nella plasticità della corteccia visiva indotta dalla deprivazione monoculare. Altri esperimenti hanno dimostrato che le neurotrofine hanno un’azione ancora più complessa nella regolazione dei processi di p. n. con effetti differenti su specifiche popolazioni cellulari: è stato ampiamente documentato, per es., che il BDNF è coinvolto nella maturazione dei neuroni inibitori e che l’NGF promuove la trasmissione colinergica. Declino della plasticità
La modificabilità delle connessioni corticali in risposta all’esperienza si riduce progressivamente e, dopo la fine del periodo critico, è molto limitata, se non del tutto assente. Sono stati condotti numerosi studi per identificare i meccanismi molecolari responsabili del declino della p. n. che accompagna il completamento dello sviluppo del sistema nervoso. Il primo fattore che regola la chiusura dei periodi critici è l’esperienza stessa. La mancanza di una stimolazione ambientale appropriata, infatti, prolunga la durata dei periodi critici; ancora una volta è paradigmatico l’esempio del sistema visivo: è stato dimostrato che la corteccia di animali allevati al buio dalla nascita permane in uno stato di immaturità fino all’età adulta e la deprivazione monoculare rimane efficace nel modificare le proprietà di dominanza oculare dei neuroni corticali molto più a lungo di quanto accade negli animali vissuti in normali condizioni di luce. Questo suggerisce che l’esperienza regola l’espressione di alcuni fattori molecolari, la cui maturazione è responsabile della chiusura del periodo critico. I fattori neurotrofici sono stati il primo gruppo di molecole per cui è stata stabilita una relazione causale tra la loro azione e la durata del periodo critico di plasticità della corteccia visiva: è stato osservato, infatti, che l’allevamento al buio riduce l’espressione corticale di tali fattori e che il blocco della loro azione, indotto dalla somministrazione di anticorpi specifici, ritarda la chiusura del periodo critico, mentre l’incremento dell’espressione di BDNF in animali transgenici la accelera. Analogamente, l’allevamento al buio ritarda la maturazione dei circuiti inibitori della corteccia visiva, mentre un’anticipazione dello sviluppo della trasmissione GABAergica determina una precoce chiusura del periodo critico della plasticità di dominanza oculare. Il ruolo centrale esercitato dal tono inibitorio nella regolazione del decorso del periodo critico è stato recentemente (2010) consolidato dall’osservazione che la riduzione farmacologica del bilancio inibizione/eccitazione nella corteccia visiva di animali adulti ripristina completamente i fenomeni di plasticità indotti dalla deprivazione monoculare. Infine, è stato documentato che la maturazione della matrice extracellulare e la mielinizzazione determinano la stabilizzazione delle connessioni neurali, sopprimendo la p. n. nel cervello maturo.
Plasticità crossmodale
Una questione che ha sempre sollevato molto interesse, anche in ambito non strettamente scientifico, è se soggetti ciechi dalla nascita, o diventati ciechi nei primi anni di vita, sviluppino capacità superiori a quelle degli individui con normali abilità visive nelle modalità sensoriali.
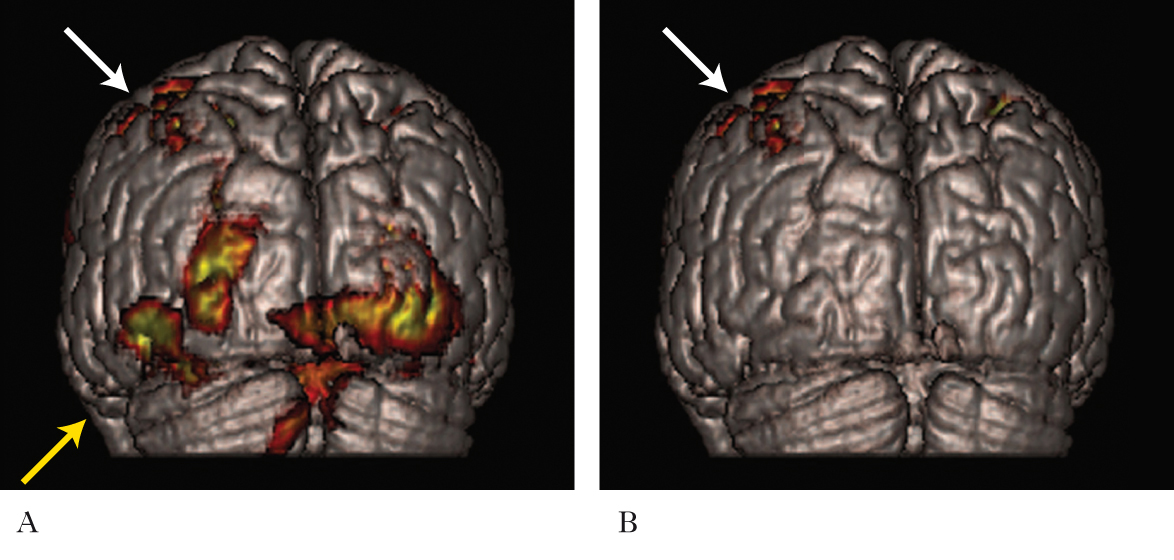
Le numerose evidenze aneddotiche a favore di questa ipotesi hanno aperto un nuovo campo nello studio della p. n. ed è stato dimostrato come la riorganizzazione delle connessioni del sistema nervoso, che si verifica in seguito alla deprivazione di un canale sensoriale, non interessi solo le aree cerebrali della modalità deprivata, ma si estenda anche ad altri sistemi sensoriali. È stato documentato, per es., che i soggetti ciechi hanno capacità tattili e acustiche superiori rispetto ai normovedenti e che l’esecuzione di compiti tattili raffinati, come la lettura Braille, o prove di localizzazione di una sorgente sonora, inducono l’attivazione della corteccia visiva di entrambi gli emisferi, in aggiunta, rispettivamente, alla corteccia somatosensoriale e a quella acustica. È interessante sottolineare, però, che informazioni sensoriali di natura diversa attivano aree differenti della corteccia visiva. Questa attivazione, non riscontrabile nella corteccia degli individui vedenti, è il segno della plasticità crossmodale e ha un preciso ruolo funzionale nell’elaborazione degli stimoli non visivi, che si riflette nelle migliori prestazioni percettive. È stato osservato, infatti, che alterando l’attività della corteccia visiva tramite una stimolazione magnetica transcranica (➔ stimolazione cerebrale elettrica e magnetica) durante un compito di lettura Braille in soggetti ciechi, essi commettono numerosi errori. La plasticità crossmodale non è limitata ai casi di deprivazione della modalità visiva, ma è presente anche nei soggetti con sordità congenita o diventati sordi nei primi anni di vita, che mostrano migliori abilità tattili e di visione periferica, correlate all’attivazione della corteccia uditiva, rispetto ai soggetti normoudenti. Il modello utilizzato per spiegare il fenomeno della plasticità crossmodale prevede gli stessi meccanismi hebbiani di consolidamento e indebolimento delle sinapsi visti in precedenza. Non è ancora del tutto chiaro, tuttavia, a quale livello agiscano questi processi. Una possibilità è che ingressi sensoriali diversi competano in maniera attività-dipendente per il controllo dei neuroni corticali delle aree associative polimodali: le afferenze più attive, ovvero quelle provenienti dalle modalità sensoriali intatte, rafforzerebbero le loro connessioni a scapito dell’ingresso deprivato, meno attivo, e la conseguente modifica delle proprietà dei neuroni polimodali sarebbe responsabile dell’attivazione della corteccia sensoriale deprivata in risposta a stimoli non specifici. Una seconda ipotesi, che non esclude la prima, prevede la presenza di connessioni ridondanti tra le diverse regioni del sistema nervoso: in caso di deprivazione del canale dominante di una corteccia sensoriale, le afferenze che veicolano le informazioni degli altri ingressi si potrebbero espandere, determinando il cambiamento delle proprietà di risposta dei neuroni corticali. Il risultato di questa competizione intermodale è un rimodellamento delle connessioni neurali, che ha l’effetto di compensare la deprivazione di un canale sensoriale con migliori prestazioni nei compiti che coinvolgono altre modalità. Laura Baroncelli, Lamberto Maffei
Plasticità nell’adulto
Nel cervello adulto la modificabilità delle connessioni tra cellule nervose in risposta all’esperienza si riduce drasticamente. Tuttavia, la plasticità è una proprietà del sistema nervoso che ci accompagna durante tutta la vita. In primo luogo, essa è alla base dei fenomeni di apprendimento (➔) e memoria (➔): la possibilità di immagazzinare nuove informazioni è correlata all’elevata dinamicità che caratterizza i neuroni delle aree deputate all’elaborazione di questi processi, prima fra tutte quella dell’ippocampo. L’incredibile flessibilità di questa struttura è sottolineata dal fatto che le modificazioni anatomiche indotte dall’esperienza possono tradursi in una variazione delle sue dimensioni. Paradigmatico è l’esempio dei tassisti di Londra, che devono acquisire una dettagliata conoscenza delle strade per esercitare la loro professione: uno studio del 2000 ha mostrato che questi soggetti presentano un ippocampo molto più sviluppato della media. È noto che anche la corteccia cerebrale può andare incontro a riorganizzazioni funzionali a seguito di modificazioni prolungate dell’esperienza ambientale: numerosi studi hanno dimostrato che la topografia corticale può mantenere un certo grado di dinamicità, regolato dai livelli di attività periferica.
Plasticità corticale e apprendimento
Il miglioramento delle capacità di discriminazione sensoriale attraverso l’esercizio è un processo detto apprendimento percettivo. Un esempio classico è la raffinata attitudine a identificare toni musicali differenti, propria dei musicisti, o l’abilità di riconoscere i volti diversi di persone della propria etnia. Negli studi effettuati per chiarire i meccanismi coinvolti nel miglioramento delle capacità percettive il modello più utilizzato prevede il rimodellamento funzionale dei territori corticali. È stato osservato che scimmie adulte, addestrate a usare solo le falangi distali di alcune dita per eseguire compiti di discriminazione tattile, mostrano un’apprezzabile espansione della rappresentazione corticale delle parti che ricevono una maggiore stimolazione tattile e propriocettiva. Tale fenomeno presuppone una ridondanza delle afferenze periferiche della corteccia somatosensoriale: il consolidamento hebbiano delle sinapsi che veicolano l’informazione proveniente dalla porzione di superficie corporea impiegata per l’apprendimento percettivo (concomitante all’indebolimento delle connessioni formate dalle fibre derivanti dalle parti adiacenti), determina il reclutamento di un maggior numero di neuroni in seguito alla stimolazione della superficie allenata e il conseguente ampliamento del suo territorio corticale a scapito della rappresentazione delle zone del corpo vicine. Un meccanismo simile si osserva nel sistema uditivo: animali addestrati a distinguere piccole differenze nella frequenza di singoli toni mostrano un miglioramento delle capacità di discriminazione acustica, accompagnato dall’allargamento delle porzioni di corteccia uditiva che rispondono preferenzialmente alle frequenze utilizzate nel compito di apprendimento. È stato dimostrato nell’uomo (2004) che imparare a fare il giocoliere induce modificazioni strutturali nella porzione di corteccia visiva devoluta alla percezione del movimento. Una relazione tra la ripetizione di un esercizio e l’espansione di un territorio corticale è stata individuata anche per la corteccia motoria. È stato evidenziato che la rappresentazione corticale delle dita della mano sinistra è più sviluppata nei suonatori di strumenti a corda, come violinisti e chitarristi, rispetto a soggetti di controllo. Sebbene il grado di riorganizzazione della mappa motoria sia funzione dell’età di inizio della pratica musicale, questi fenomeni di plasticità sono presenti anche nel cervello adulto. A conferma di ciò, la risonanza magnetica funzionale di un gruppo di persone adulte, a cui si richiedeva di eseguire ripetutamente una rapida sequenza di movimenti delle dita, ha mostrato che il compito motorio in questione evocava l’attivazione di un’area della corteccia motoria più grande rispetto a una sequenza di controllo.
Plasticità indotta da lesione
Un robusto rimodellamento della corteccia cerebrale di individui adulti si verifica anche in seguito a una riduzione della stimolazione periferica, dovuta a lesioni o amputazioni. I primi dati in merito sono stati ottenuti nel sistema somatosensoriale della scimmia: la lesione di un nervo periferico determina la comparsa di una zona corticale silente, dove le cellule non ricevono più segnali dai recettori; successivamente questi neuroni iniziano a rispondere alla stimolazione della superficie corporea adiacente a quella deprivata e si determina una riorganizzazione della mappa somatosensoriale (➔ tatto). Un fenomeno simile si manifesta nei pazienti che sperimentano la sindrome dell’arto fantasma (➔). Analogamente, nel sistema visivo una piccola lesione della retina (scotoma) fa sì che l’area corticale devoluta alla percezione della porzione di campo visivo interessata, non ricevendo più informazioni sensoriali, sia attivata dai recettori retinici situati vicino alla zona lesionata. Anche la corteccia motoria (➔) è condizionata dalla stimolazione proveniente dalla periferia: studi sul ratto hanno dimostrato che l’attivazione dell’area motoria deputata al controllo delle vibrisse, dopo avere lesionato le fibre che le innervano, provoca il movimento dell’arto anteriore. Tali modificazioni plastiche avvengono tramite lo smascheramento di connessioni (intracorticali o sottocorticali) già esistenti, che vengono disinibite dall’indebolimento di quelle private della stimolazione sensoriale. Un altro meccanismo, che non esclude il precedente, è la manifestazione di fenomeni di sprouting, ossia l’emissione di collaterali assoniche a carico delle afferenze che non hanno subito la lesione e la formazione di nuove sinapsi tra queste ultime e le aree corticali temporaneamente silenziate dalla lesione.