
Orientamento degli animali
Orientamento degli animali
di Leo Pardi
Orientamento degli animali
sommario: 1 Che cosa è l'orientamento. 2. Un cenno sulla storia del problema. 3. Strategie per trovare una meta. a) Ricerca a caso e ricerca sistematica (orientamento mediato a meta indefinita). b) Orientamento mediato a meta definita. 4. Conclusione. □ Bibliografia.
1. Che cosa è l'orientamento
In una bella pagina autobiografica uno scrittore italiano narra come il padre suo, pastore, gli insegnasse quando era bambino a non perdersi mai nella solitudine della campagna: ‟Mentre avanzavamo, lui si industriava a fissarmi nella mente i punti caratteristici del terreno, le querce che spiccavano in modo particolare, per la loro forma, per la loro grandezza, curvature varie e per i loro difetti: buchi e gibbosità. Oppure i macigni e i cespugli. Con tutta la sua esperienza mi aiutava a vederli nel loro insieme [...] a farmi un quadro della loro disposizione [...]". (G. Ledda, Padre padrone. L'educazione di un pastore, Milano 1975, p. 15).
Questo - costruito sulla base di un reticolato di punti di riferimento via via arricchito dall'apprendimento - è certo un processo d'orientamento: lo è, anzi, nel senso più comune, anche se l'uomo d'oggi vi ricorre più spesso nell'ambiente urbano. Ma è pure orientamento, stavolta proprio nel senso etimologico, quello di colui che - perduto nel deserto - decidesse di andare diritto il più a lungo possibile, lasciando alle spalle l'oriente. E di orientamento, ancora, si tratterebbe se questo stesso individuo - determinata la propria posizione con il cronometro e il sestante - assumesse e mantenesse una rotta prefissata con l'aiuto di una bussola. L'elenco non finisce qui, poiché azioni di orientamento sono ancora, per esempio, navigare verso un faro, dirigersi verso una persona, una casa, un albero, allontanarsi da un fuoco. Non solo, ma coinvolgono orientamento ‛azioni parziali' come allungare il braccio per afferrare un oggetto o per fare ginnastica, riassestare e mantenere l'equilibrio del corpo, assumere una certa posizione nel riposo, porsi il cappello in capo, disporre i mattoni per costruire un muro o, infine, scrivere queste righe: un elenco già lungo e, certo, non completo.
Gli animali non sono da meno, per la varietà dei fenomeni. Non solo l'orientamento del corpo e delle appendici interviene in cicli funzionali strettamente confrontabili con quelli ricordati per l'uomo (mantenimento dell'equilibrio o della posizione preferita, ricerca del nutrimento e del compagno, ritrovamento e costruzione del rifugio e del nido, ricerca del territorio o dell'area comunque più favorevole, allontanamento da sorgenti di stress), ma si manifesta sovente attraverso rilevamenti sensoriali e meccanismi di cui non sempre si trova l'equivalente nella nostra specie.
Potremmo definire questo insieme apparentemente molto eterogeneo di fenomeni, l'orientamento nello spazio, come ‟la capacità degli organismi di assumere e controllare attivamente posizioni e movimenti totali o parziali, in rapporto a informazioni (esterne o interne) di natura spaziale" (v. Schöne, 1975). Così inteso, l'orientamento è caratteristica veramente universale degli organismi, poiché vi sono compresi a buon diritto non solo quasi tutti i movimenti (ai vari livelli) dell'individuo intero o delle sue parti (sino alle risposte orientate delle singole cellule e, persino, di certi organuli intracellulari, come, ad esempio, quelle dei cloroplasti rispetto alla luce nelle cellule vegetali), ma anche molti movimenti connessi con l'accrescimento e con lo sviluppo, come i movimenti morfogenetici.
E ciò vale per tutti gli organismi, vegetali o animali, fissi o mobili che siano. Qui dobbiamo tuttavia limitarci a considerare i movimenti che fanno parte del comportamento, la loro organizzazione spaziale e, soprattutto, alcuni aspetti del loro valore adattativo.
2. Un cenno sulla storia del problema
Poiché quasi tutto il comportamento motorio è orientato, il problema è centrale in etologia e in fisiologia comparata. Ed è un problema del nostro secolo, se si prescinde dalle ricerche anticipatrici di L. Spallanzani sull'orientamento dei Chirotteri nell'evitare gli ostacoli (1793). La teoria dei ‛tropismi' di J. Loeb, il primo tentativo di interpretazione fisiologica unitaria dei fenomeni di orientamento, nasce nell'ultimo decennio del secolo scorso e si afferma negli anni che precedono la prima guerra mondiale. Per Loeb ‟i movimenti causati dalla luce o da altri fattori sembrano al profano l'espressione di una volontà e di una finalità presenti nell'animale, mentre, in realtà, l'animale è forzato ad andare dove le gambe lo portano: poiché la condotta degli animali consiste di movimenti obbligati" (v. Loeb, 1918, p. 1). La spiegazione loebiana del fototropismo positivo di un animale, che si orienta (e procede) verso una sorgente luminosa, è schematicamente questa: eguale stimolazione sui fotorecettori simmetrici produce eccitazioni identiche nel sistema nervoso centrale e, quindi, eguale ‛tensione' nei muscoli simmetrici, a differenza di una stimolazione ineguale. Nel primo caso l'animale si orienta (e si muove rettilineamente) verso la luce, nel secondo è forzato prima a ruotare fino a ristabilire l'eguaglianza di stimolazioni. Insomma, animal non agit, agitur. Pur contrastati da molti (v., per es., Mast, 1911), e con varie limitazioni, attenuazioni e correzioni, questi concetti dominano a lungo nello studio dell'orientamento nel periodo fra le due guerre.
A. Kühn (v., 1919) basa il suo sistema sulle concezioni loebiane distinguendo fra ‛tropismi' e ‛tassie'. Egli riserva il termine di ‛tropismi' (in armonia con l'originale accezione botanica) alle disposizioni orientate che gli animali sessili assumono, per effetto di accrescimenti differenziali (o, forse, di variazioni di turgore) di fronte alla luce o nel campo gravitazionale: fenomeni, questi, che non sarebbero da considerarsi comportamentali, nel senso corrente del termine. Sono invece - a pieno diritto - comportamento le ‛tassie', modalità elementari di orientamento degli animali mobili rispetto agli stimoli, che sono pur sempre assimilabili per Kühn a ‛movimenti riflessi' dell'animale intero. Una letteratura estesissima, basata su analisi accurate del comportamento orientato degli animali più diversi, dagli Unicellulari ai Vertebrati e di fronte alle più varie stimolazioni (fototassie, chemotassie, geotassie, galvanotassie, anemotassie, reotassie, ecc.) fiorisce con Kühn sulla base prevalente di esperimenti di laboratorio, conducendo a risultati assai importanti. Si riconosce, soprattutto, che il meccanismo descritto da Loeb (‛tropotassia' secondo Kùhn) è, se mai, uno solo dei molti possibili fra le reazioni riflesso-simili dell'animale intero agli stimoli esterni. Sono dovuti a Kühn i nuovi concetti di ‛telotassia' e di ‛menotassia'. G. S. Fraenkel e D. L. Gunn (v., 1940) riassumono e discutono egregiamente questa prima fase delle ricerche in un'opera ormai classica proponendo un sistema kühniano modificato dei meccanismi elementari, che esclude ancora i tropismi in senso stretto, ma fa maggior posto - accanto alle tassie - alla nuova categoria delle cinesi. Poiché questo sistema, nonostante critiche del resto assai fondate (v. Schöne, 1975), non ha solo valore storico ma è tuttora largamente in uso (almeno per esigenze di descrizione) e poiché a esso ci riferiremo necessariamente più volte, riteniamo opportuno riportarlo per esteso in forma tabellare (v. tabella), aggiungendo brevi definizioni.

Ma, frattanto, nell'analisi scientifica del comportamento in generale e di quello istintivo in particolare, sta accadendo qualcosa di simile a una rivoluzione. Negli anni fra le due guerre, difatti, per una felice confluenza fra gli indirizzi etologico e fisiologico, si affermano nuove dottrine che contrastano radicalmente sia con le posizioni vitalistiche, sia con quelle, strettamente riduzionistiche, che interpretano ogni condotta animale unicamente in termini di stimolo e di risposta e concepiscono il sistema nervoso centrale quasi solo come semplice commutatore fra le informazioni esterne e i comandi motori. La ‛spontaneità' del comportamento, intesa come mutamento dello stato centrale non accompagnato da variazioni della situazione stimolante esteriore, e i suoi correlati fisiologici cominciano a essere oggetto di analisi sperimentale e le teorie dell'azione istintiva come riflesso o catena di riflessi rivelano sempre di più la loro inadeguatezza.
Singolarmente, questi concetti si fanno strada un poco più tardi nell'analisi dell'orientamento. Ed è solo negli anni cinquanta che, soprattutto per opera di E. von Holst, si perviene al concetto - e lo si dimostra sperimentalmente anche per animali ‛inferiori' - che i centri determinano, momento per momento, in dipendenza o meno da variazioni ambientali, la posizione che l'animale dovrà assumere rispetto alle grandezze di riferimento, cioè la ‛direzionenorma' e, insieme, il suo verso (Sollrichtung). Poiché anche con una costellazione di stimoli invariata l'animale risponde in modo differente a seconda della ‛disposizione centrale' (Stimmung), il processo di orientamento non si esaurisce in termini di stimolo-reazione, o - meglio - è reazione solo in quanto ogni deviazione registrata dalla direzione-norma centralmente stabilita viene ‛automaticamente' annullata.
È ancora a von Holst (e Mittelstaedt) che si deve, nel 1950, con l'introduzione del ‛principio di reafferenza', il primo modello concreto circa l'intervento regolatore del sistema nervoso centrale, nel meccanismo che connette l'afferenza sensoriale all'efferenza motoria. R. W. Sperry, ancora nel 1950, propone un'ipotesi analoga. Da allora in poi si preferirà quindi parlare di ‛azione di orientamento' piuttosto che di ‛reazione', e assumerà uno sviluppo sempre più rilevante, soprattutto per opera di Mittelstaedt (1958), di Hassenstein (1960), di Schöne (1973) e di altri, l'interpretazione dei processi relativi nel linguaggio della cibernetica.
Ma negli anni della seconda guerra mondiale e nell'im- mediato secondo dopoguerra vi è un altro sensibile muta- mento, non meno significativo del precedente. A un periodo caratterizzato (in prevalenza, beninteso) dallo studio di laboratorio su reazioni elementari isolate (cinesi e tassie) e in condizioni talvolta estremamente innaturali (classico, ad esempio, il saggio per vedere cosa fanno i più diversi animali fra due sorgenti di luce artificiale) subentra un maggiore interesse per i metodi di orientamento, multipli, complessi e complementari, che l'animale effettivamente adopera in natura entro i normali cicli funzionali, quando ricerca le ‛mete' necessarie alla vita ed evita le sorgenti di stress. È il problema del ‛significato adattativo', dell'ecologia dell'orientamento, che giunge in primo piano e la stessa conoscenza della fisiologia dei meccanismi se ne avvantaggia grandemente. Sono di questo periodo, che si prolunga fino ai giorni nostri, le scoperte senza dubbio più significative sull'effettiva ‛funzione naturale' delle risposte orientate: la ecolocalizzazione ultrasonica dei Chirotteri (Griffin e Galambos, 1942; Dijkgraf, 1943; Möhres, 1943), l'orientamento solare nelle api (von Frisch, 1950) e negli Uccelli (Kramer, 1950), l'orientamento solare e lunare degli animali ripari e intertidali (Pardi e Papi, 1952), l'orientamento stellare negli Uccelli (Sauer, 1955), l'elettrolocalizzazione nei Pesci (Lissmann, 1958), l'orientamento magnetico in molti gruppi (Schneider, 1957, negli Insetti; Barnwell e Brown, 1964, nei Molluschi; Merkel e Wiltschko, 1965, negli Uccelli), lo homing chemoreotattico nella migrazione anadroma dei salmoni (Hasler, 1956) e il complesso sistema ‛navigazionale' olfattivo del colombo viaggiatore (Papi e altri, 1971-1975). Vi sono spesso risultati paralleli non meno importanti di quello principale in parecchie di queste indagini, poiché non si tratta tanto di sapere ‛che cosa fa' l'animale di fronte a uno stimolo noto e isolato, ma proprio di scoprire come si orienta in natura, come trova la meta nel suo ambiente; l'esclusione dei riferimenti sensoriali possibili, uno per uno o, meglio ancora, di più d'uno simultaneamente, non conduce lo studioso a chiedere spiegazioni allo ‛stregone' (come ebbe a dire scherzosamente uno zoologo quasi disperato di fronte al ‛mistero' dello homing di certi Anfibi), ma porta a scoprire nuove modalità sensoriali (percezione della luce polarizzata o dell'ultravioletto, di ultrasuoni o di infrasuoni, sensibilità al campo magnetico terrestre o a campi elettrici) o a rivelare nuove interazioni o prestazioni di quelle note, assolutamente inattese, come nello homing chemoreotattico dei salmoni o nella navigazione olfattiva dei colombi.
Nello studio dell'orientamento in natura, anche i metodi di esperimento e di rilevamento si evolvono: a quelli, classici, della cattura, marcatura e ricattura, si aggiunge - semplicissimo ma oltremodo ricco di risultati fino a oggi - il metodo della determinazione dei ‛punti di sparizione' degli animali al momento del rilascio, in condizioni naturali o sperimentali variate di cui parleremo in seguito. Si perfeziona l'uso di tecniche particolari adatte a seguire il percorso degli animali (radio-tracking, anche mediante satelliti, radar-tracking, inseguimento con elicotteri o piccoli aerei). Per piccole specie si diffonde lo studio - inaugurato da Kramer (1950) per gli Uccelli e da Pardi e Papi (1952) per gli Artropodi - delle distribuzioni assunte da individui singoli o in gruppo in ambienti circolari confinati, in condizioni tali da sottrarre o modificare agevolmente le informazioni esterne che possono pervenire agli animali.
Le distribuzioni circolari ottenute con questi metodi divengono - proprio per la sollecitazione da parte degli studiosi dell'orientamento - oggetto di più attenta considerazione da parte dei biomatematici (v. soprattutto Batschelet, 1965), che suggeriscono sempre nuovi test atti a giudicarne attendibilità e differenze statistiche.
3. Strategie per trovare una meta
Il titolo di questo capitolo indica la natura dei problemi a cui, come si è detto, intendiamo limitarci. Un'escursione anche sommaria attraverso i vari e complessi temi dell'orientamento richiederebbe uno spazio enorme. Qui vogliamo considerare, perciò, soltanto gli aspetti ecologici principali del cosiddetto orientamento ‛mediato'.
Se definiamo ‛meta positiva' (goal o target) ogni oggetto, luogo, area che permetta lo svolgimento di qualche funzione essenziale per il mantenimento dell'individuo e la propagazione della specie, e ‛meta negativa' ogni sorgente localizzata di stress, l'orientamento caratterizzato dalla presenza di una meta, quasi sempre extracorporea (‛orientamento secondario'), si separa assai bene dalle forme dell'orientamento ‛primario' o posturale, di cui è tipico esempio - fra i molti - il mantenimento della normale posizione di equilibrio.
Ora, una meta positiva - e a una meta positiva ed extracorporea ci riferiremo d'ora in poi, parlando semplicemente di ‛meta' salvo esplicito riferimento - può fornire essa medesima gli stimoli che dirigono l'animale (come nel caso del predatore che insegue la preda). L'orientamento secondario dicesi allora ‛immediato'. Ma in moltissimi altri casi la meta o non emette alcuno stimolo ‛orientante' o può emetterlo solo al di fuori dell'ambito sensoriale, oppure, infine, emette soltanto stimoli ‛motivanti', capaci cioè di indurre un comportamento di ricerca della meta, ma non di orientarlo verso di essa. Tutti i meccanismi che, in tali condizioni, aumentano la probabilità di raggiungere la meta, possono indicarsi come orientamento secondario ‛mediato'. Quasi sempre il raggiungimento finale è poi dovuto a una seconda fase di orientamento ‛immediato', allorché la meta viene a cadere nell'ambito sensoriale, ma ciò non è sempre necessario: taluni animali finiscono per raggiungere un'area (meta) favorevole e riconoscerla senza esser mai guidati da stimoli da essa prodotti.
La natura e la complessità dei meccanismi dell'orientamento mediato (come pure di quello immediato, del resto) dipendono innanzitutto dagli strumenti sensoriali e dall'organizzazione neurale della specie. Ma poiché qui vogliamo soprattutto cogliere - secondo quell'indirizzo a cui desideriamo attenerci - il valore adattativo delle forme di orientamento e i modi della diversa interazione dell'organismo con l'ambiente (ecologia dell'orientamento), sarà opportuno fare attenzione soprattutto alla ‛distribuzione spaziale' delle mete e alle informazioni che l'organismo ne riceve.
a) Ricerca a caso e ricerca sistematica (orientamento mediato a meta indefinita)
Gli animali assumono sovente direzioni che non sembrano avere alcuna relazione determinata con una meta. In condizioni interne opportune l'animale, spesso (ma non sempre) motivato - ma non orientato - da variazioni d'intensità dei fattori ambientali, ‛ricerca' o ‛esplora', con effetti che possono portare al raggiungimento di condizioni favorevoli o di oggetti di importanza vitale: una ‛meta' nel nostro senso, che non è definita quando il processo s'inizia.
La ricerca può essere apparentemente casuale, combinando variazioni di velocità lineare della locomozione e/o frequenza dei periodi di moto (‛ortocinesi'). Se è la variazione d'intensità di uno stimolo esterno a scatenare il comportamento, a seconda della natura dello stimolo si parla di igro-, foto-, chemiocinesi ecc. Con la sola igroortocinesi taluni Crostacei Isopodi terrestri fortemente igrofili riescono nella maniera più semplice ad aggregarsi nelle aree di umidità preferita, decelerando il moto fino a fermarsi a mano a mano che le condizioni esterne diventano più favorevoli e accelerandolo in caso contrario (Gunn, 1937). Con meccanismi chemotermo-clinocinetici (mutando direzione a caso in condizioni non ottimali) anche Batteri e Protozoi riescono ad aggregarsi nelle zone più adatte di un gradiente chimico o termico. Filogeneticamente, quindi, si tratta di meccanismi fra i più elementari e primitivi dell'orientamento, anche dal punto di vista dell'organizzazione sensoriale, poiché essi presuppongono unicamente - quando sono esternamente motivati - una capacità di percezione delle variazioni di intensità degli stimoli, che può essere attribuita anche a sensibilità generalizzata e non essere necessariamente mediata da speciali recettori.
Di gran lunga più frequenti sono i casi in cui l'esplorazione diventa più organizzata e sistematica e ciò specialmente quando l'animale è motivato alla ricerca di mete spazialmente concentrate (oggetti, aree più ristrette rispetto all'ambito d'azione dell'individuo). L'esplorazione può investire allora successivamente, ad esempio, aree limitate sempre diverse, con movimenti formalmente determinati, talvolta specie-specifici. Jander (v., 1975) ne distingue tipi diversi: ranging (una mosca che esplora un tavolo percorrendo tratti rettilinei e curve secche improvvise), esplorazioni pendolari, vari tipi di spostamenti sistematici durante la pastura o, infine, semplicemente ‛tirar diritto' o transecting. Andare diritto, se le mete sono distribuite a caso e/o se si tratta di uscire il più economicamente possibile da un'area di stress, può essere un ottimo espediente che useremmo noi stessi. Mentre però le forme già nominate di movimento esplorativo possono essere programmate anche solo su informazioni interne, andare diritto o quasi è difficile senza utilizzare riferimenti esterni, diversi a seconda della specie, che tuttavia - ripetiamo - non hanno relazione alcuna, fissa e determinata, con una meta: affidandovisi, l'animale aumenta semplicemente la probabilità di trovarla con un minimo dispendio di energia. Eccone alcuni esempi.

Durante certe emigrazioni massicce conseguenti a moltiplicazione eccessiva, nella ricerca di nuove aree da colonizzare, taluni insetti si valgono per mantenere una rotta rettilinea (che non conduce necessariamente alla meta) di elementi del paesaggio e/o di correnti aeree. Il lepidottero Ascia monuste L., studiato da Nielsen (1961) in Florida, sciama a migliaia sulla costa dai mangrovieti delle lagune interne e segue verso sud o nord, indifferentemente, la linea di riva o le strade litorali, persino le linee telegrafiche, un comportamento che - a differenza di una ricerca disordinata - aumenta le probabilità di pervenire in aree meno affollate. Forse hanno lo stesso significato alcune emigrazioni delle libellule lungo le spiagge, in cui - spesso ma non sempre - anche il vento è fattore orientante.
Su scala minore ha lo stesso significato di ricerca sistematica in base a riferimenti esterni il comportamento delle formiche che cercano il nutrimento seguendo fedelmente il contorno delle superfici da esplorare (v. Jander, 1975).
Ma in taluni biotopi monotoni (sabbie litorali o desertiche, ad esempio) possono mancare del tutto riferimenti topografici prossimi o remoti; alcuni Artropodi (Insetti soprattutto) si dimostrano allora capaci di mantenere rotte rettilinee in direzioni qualsiasi, prendendo - rispetto all'azimut solare, forse anche lunare - un angolo fisso per un certo tempo: dal punto di vista ecologico è un transecting solare (lunare), secondo il linguaggio kühniano una ‛fotomenotassia' (v. tabella). Che il sole sia il fattore orientante si può provare schermandolo e proiettandone l'immagine riflessa dall'azimut opposto (esperimento di Santschi, 1911): l'animale fa dietrofront e prende, rispetto allo specchio, lo stesso angolo che faceva col sole.
È subito evidente però che una rotta indefinitamente rettilinea non può essere mantenuta con questo mezzo: variando la posizione del sole, se l'angolo rimane immutato, cambia la direzione dell'animale. Ciò non ha grande importanza per brevi percorsi e, d'altronde, questi animali mutano spontaneamente e senza regola alcuna la loro disposizione rispetto al sole, il che può - entro certi limiti - essere pure vantaggioso. Ma vi sono specie che sanno mantenere una direzione costante variando regolarmente il proprio angolo di orientamento col sole: la fotomenotassia, da ‛non cronometrica', diventa ‛cronometrica'. Ora, in talune specie, questa più sofisticata capacità (che vedremo pienamente utilizzata nei meccanismi di orientamento con meta definita) non sembra avere alcun risultato immediatamente comprensibile, poiché la direzione costante assunta non ha relazione alcuna con una meta: è il caso della non-sense orientation solare verso NO di certi Anatidi studiata da Matthews (1961) o verso direzioni diverse da popolazione a popolazione nel caso del coleottero Geotrupes sylvaticus F. (Birukow, 1953). Si tratta di una forma d'esplorazione interpretabile come un transecting? Non si può affermarlo con sicurezza.
Vi è per contro, molto più probabilmente, un transecting magnetico. In quest'ultimo decennio si sono moltiplicate prove inoppugnabili che gli animali sono capaci di rilevare le caratteristiche direzionali del campo magnetico terrestre e le sue variazioni stagionali e giornaliere (Martin e Lindauer, 1973). È possibile che uno degli usi della ‛bussola magnetica', oltre a quelli che ha in certe forme di orientamento posturale (cioè primario) e nell'orientamento mediato con meta definita, sia appunto il semplice mantenimento di rotte rettilinee qualsiasi, atte a far sortire più o meno rapidamente da condizioni di stress: tale significato potrebbero avere le direzioni preferenziali più volte accertate, in assenza di altri riferimenti, in Molluschi, in Planarie e persino in Coleotteri Carabidi (Mletzko, 1969).
b) Orientamento mediato a meta definita
Molti animali (oltre all'uomo) sanno risolvere egregiamente il problema di trovare la direzione di una meta determinata, che sia al di fuori dell'ambito sensoriale. E con due ordini di meccanismi radicalmente diversi. Talvolta l'orientamento viene assunto soltanto per informazioni interne (orientamento ‛idiotetico') senza riferimenti esterni, ma nella maggioranza dei casi, ovviamente interagenti con informazioni endogene di varia natura, i riferimenti esterni sono indispensabili: l'orientamento si dice allora ‛allotetico'.
1. Orientamento senza alcuno stimolo orientante esterno. - Anche in opere recenti si legge che i movimenti orientati sono sempre legati alla presenza di stimoli orientanti. Se per stimolo s'intende una qualsiasi informazione di origine esterna, mediata dagli organi di senso, l'asserzione è erronea. Gli esempi di sequenze motorie perfettamente orientate anche verso mete extracorporee definite, che non emettono alcun segnale e senza alcun altro riferimento esterno, sono numerosissimi. Per la nostra specie un esempio molto citato è quello del pianista che trova i tasti giusti anche senza controllo visivo.
Per gli animali gli esempi meglio analizzati ce li offrono i ragni. Agelena labyrinthica Clerk, che costruisce una tela con una tana imbutiforme eccentrica, è capace di ritornarvi rettilineamente da qualunque punto della tela anche se ha raggiunto questo punto con un percorso molto circonvoluto: di ciò è capace anche in assenza assoluta di ogni possibile stimolo esterno (luce, gravità, stimoli olfattivi, gradiente di elasticità della tela) (Görner, 1958 e 1973).
Cupiennus salei Keys, un ragno che non fa tele, scacciato da un punto in cui ha trovato una preda, non solo vi torna poco dopo ben orientato anche in assenza di stimoli visivi (accecato), ma cerca attivamente, nel punto dov'era, la preda che lo sperimentatore ha frattanto spostato appena più in là (Barth e Seyfahrt, 1971). In questo caso, dunque, l'animale controlla in qualche modo direzione e distanza della meta: l'orientamento è ‛vettoriale'.
L'interpretazione corrente di questi e altri simili comportamenti è che l'animale, durante il percorso (purché sia stato ‛attivamente' compiuto), registra la grandezza delle successive rotazioni e traslazioni, cosicché, in definitiva, gli è precisamente nota la propria posizione rispetto al punto di partenza. L'interpretazione cibernetica del procedimento, dovuta a H. e L. Mittelstaedt (1972), ammette l'interazione fra due accumulatori d'informazioni, uno per le deviazioni, l'altro per i percorsi compiuti verso ciascun azimut. La capacità del ritorno orientato viene fortemente compromessa dall'ablazione di certi propriocettori localizzati in corrispondenza delle articolazioni delle zampe (organi liriformi) (Seyfahrt e Barth, 1972).
2. Orientamento con l'uso dei riferimenti di guida. - In ordine di localizzazione sempre più limitata si distinguono tre categorie di riferimenti (nell'orientamento mediato, si intende): a) riferimenti spazialmente continui, di validità universale, specie-specifica, come il campo gravitazionale, il campo magnetico terrestre, altri eventuali gradienti geofisici orizzontali o verticali e i riferimenti astronomici o celesti (sole, luna, stelle, luce polarizzata del cielo azzurro); b) riferimenti discontinui, di validità universale, specie-specifica. Ad esempio: 1) campi di stimolazione (come le correnti aeree, le correnti marine e fluviali, il moto ondoso) o 2) elementi salienti del paesaggio visuale; c) riferimenti discontinui, a validità localizzata, individual-specifica: ad esempio riferimenti visuali, olfattivi, tattili dell'ambito direttamente (o indirettamente) esplorato dall'individuo, le cui caratteristiche particolari di struttura e di posizione gli sono singolarmente note.
Per i riferimenti della classe a), ma spesso anche per quelli del gruppo b, 1) (v. Schöne, 1975), si parla di orientamento ‛alla bussola': (astronomico per i riferimenti celesti, cr0nometrico quando comporta variazione regolare nel tempo dell'angolo di orientamento). Per i riferimenti b, 2) ma soprattutto per quelli della categoria c) si usa il termine nautico di ‛pilotaggio'. La navigazione, di cui si dirà in seguito, non può definirsi solo per la classe di riferimenti usati.
Per raggiungere una meta definita al di fuori della portata degli organi di senso mediante l'uno o l'altro dei riferimenti indiretti ora specificati, l'animale deve aver acquisito in qualche modo informazioni circa la relazione spaziale fra quel riferimento e la posizione della meta. Il problema comporta soluzioni diverse, come si vedrà dagli esempi che seguono.
A. I sistemi unidirezionali. Sono fra i più noti e realizzano soluzioni di problemi relativamente più semplici. La meta o le mete hanno una posizione fissa nello spazio per una o più specie o, almeno, per una data popolazione. I movimenti verso la (o via dalla) meta si compiono o assumendo un angolo determinato rispetto a un riferimento di direzione determinata o, eventualmente, se il riferimento varia di posizione nelle 24 ore come il sole, variando l'angolo di orientamento secondo un ciclo praticamente uguale nel corso della vita e identico per tutti gli individui di una data popolazione.
Le forme tipiche di orientamento unidirezionale sono l'orientamento ‛stratale' (ad esempio, le migrazioni verticali del plancton) e l'orientamento ‛zonale' degli animali ripari e intertidali.
Forme speciali di orientamento unidirezionale si hanno quando il sistema di riferimento è di per sé distorto nello spazio secondo configurazioni costanti (come la corrente di un fiume) o variabili (come le correnti aeree o marine). Vi sono animali che, nel raggiungimento delle loro mete, mantengono una determinata posizione rispetto a tali riferimenti (generalmente reotassia, anemotassia positiva o negativa), seguendo perciò le distorsioni del sistema; ciò si verifica, ad esempio, nelle migrazioni fluviali di vari pesci.
Alla stessa categoria speciale di orientamenti unidirezionali è riferibile anche l'orientamento basato sull'utilizzazione di ‛piste' o tracciati odorosi, comune nei Formicidi, nelle termiti e anche in talune api sociali (Melipone), con l'importante differenza che il sistema di riferimento (chimico) è spesso autoprodotto dalla stessa specie che se ne vale e che non è di solito continuo, ma segmentale.
Quando, valendosi di sistemi unidirezionali, gli animali, oltre a informazioni sulle direzioni, mostrano di possedere in partenza anche informazioni sulla distanza della meta, l'orientamento unidirezionale è, per di più, ‛vettoriale' (univettoriale).
I. L'orientamento stratale. Un ecosistema sufficientemente sviluppato in senso verticale si differenzia di regola in sottosistemi (o ‛strati') ecologicamente diversi, praticamente illimitati secondo due dimensioni. Ciò avviene, ad esempio, in una foresta coi suoi piani diversi (lettiera di foglie, piano erbaceo, arbustivo, chioma arborea) o nel suolo, in un lago, nel mare. Il mantenimento di certi livelli, per talune specie, o la capacità di migrazione (periodica o aperiodica) fra di essi, per altre, sono essenziali. Come nella distribuzione zonale di cui diremo più oltre, anche in quella stratale le variazioni - entro breve spazio - delle condizioni di ambiente possono essere molto intense, anche se più o meno graduali.
Molto studiate sono le migrazioni unidirezionali a periodicità giornaliera di specie marine o limniche di organismi planctonici (Crostacei soprattutto: Cladoceri, Copepodi, Misidacei, ecc.) che talvolta si estendono, a seconda delle specie e degli ambienti, per varie centinaia di metri fra acque più superficiali durante la notte (generalmente) e acque profonde durante il giorno.
Queste migrazioni, che rappresentano la più importante dislocazione periodica di biomassa del nostro pianeta, hanno sicuramente significato trofico e, fors'anche, protettivo. La risalita notturna (generalmente) porta gli animali negli strati superficiali eufotici ricchi di fitoplancton, mentre la discesa in strati profondi durante il giorno può proteggere dagli effetti dannosi della luce solare e dalla più alta temperatura superficiale o anche - come è stato suggerito - da una predazione eccessiva. Il decorso delle migrazioni è governato - in alcune specie almeno - da un ritmo circadiano.
La direzione della migrazione è una sola, ovunque e per tutte le specie. I riferimenti sono a direzionalità costante, come la luce dall'alto, o anche - per di più - costantemente presenti, come quello gravitazionale: una situazione certo più uniforme e più semplice di quella che troveremo nell'orientamento zonale. Tuttavia l'intero meccanismo è molto complesso e, sotto certi aspetti, ancora ricco di problemi irrisolti. Le variazioni dell'intensità luminosa sono, senza dubbio alcuno, il fattore più importante che regola inizio, termine e verso della migrazione. Anche la pressione ha importanza. La luce è comunque, innanzi tutto, il fattore motivante principale e al tempo stesso quello che determina la ‛direzione-norma'. I planctonti seguono con precisione gli spostamenti verticali di una zona preferenziale specifica di intensità luminosa: durante un'eclissi solare risalgono, come al tramonto, per discendere nuovamente alla sua fine. Nella luce continua dell'estate artica il ritmo tipico è assente.
I fattori guida, cioè gli stimoli orientanti, sono - ancora la luce dall'alto e, inoltre, la gravità. La locomozione è di tipo tattico (v. tabella): fototassia positiva e negativa, e geotassia negativa e positiva, rispettivamente, nelle due fasi di ascesa e discesa. Effetti fotoclinocinetici (v. tabella) non sembrano dimostrati e appaiono improbabili, considerando le velocità con cui organismi anche minuti coprono notevoli distanze (Creutzberg, 1978). Le precise modalità di intervento e di interazione della foto- e geotassia sono assai varie e, per molte forme, ancora oggetto di discussione.
II. L'orientamento zonale degli animali ripari e intertidali. Nelle zone di contatto fra ecosistemi diversi, ad esempio sulle rive marine, lacustri o fluviali, si differenziano sempre una o più zone con caratteri di passaggio e soggette, di regola, a variazioni di livello. Più o meno estese secondo la direzione perpendicolare al confine, tali zone possono considerarsi praticamente illimitate nell'altro senso. Molte specie mobili, legate a una zona particolare e incapaci di vivere a lungo al di fuori di essa, sono dotate di precisi meccanismi per mantenervisi o ritornarvi con un percorso di minimo dispendio, cioè praticamente perpendicolare ai confini della zona: esse tendono a seguire, perciò, ogni variazione di livello della loro zona di elezione, periodica o aperiodica che sia. Questo tipo di orientamento si dice ‛zonale'.
I meccanismi di orientamento unidirezionale zonale possono essere assai diversi e comunque, di regola, la stessa specie ne ha a disposizione più d'uno. I più semplici, poiché utilizzano riferimenti che hanno una relazione fissa con la direzione da assumere (visione diretta di caratteristiche del paesaggio, pendenza del substrato, ecc.), non sono cronometrici, cioè non implicano il senso del tempo. Altri, più complessi, si valgono di riferimenti astronomici o celesti (sole, luna, luce polarizzata del cielo) e sono ‛cronometrici'.
Il più noto dei sistemi di quest'ultimo tipo è quello solare di certi Anfipodi del sopralitorale, studiato da Pardi (1952-1976), da Papi (1952-1966), da Ercolini (1963-1976) e da vari altri autori (recentemente da Hartwick, 1976). Questi piccoli crostacei (il loro nome volgare è ‛pulci di mare', Sandflohe o Sandhoppers, perchè capaci di salto) sono diffusi in gran parte del mondo con varie specie assai simili (dei generi Talitrus, Orchestia, Talorchestia, ecc.) che vivono di norma nella sabbia umida. Detritivori e attivi in superficie soprattutto al crepuscolo e di notte, ma anche in pieno giorno (in certe condizioni), possono sia allontanarsi dalla loro zona di elezione per migrazioni attive con ritmo diurnale e/o tidale alla ricerca di nutrimento, sia esserne rimossi per effetti accidentali (moto ondoso, vento, fuga da predatori). Essi evitano però sia la sommersione come il soggiorno prolungato nella sabbia asciutta, con un meccanismo assai preciso di ritorno, tanto dal mare come dall'entroterra. La sommersione in acqua di mare motiva l'animale alla fuga verso terra, il disseccamento sulla sabbia asciutta a quella verso il mare: la direzione viene assunta valendosi del sole o di altri riferimenti (stimoli orientanti). Si noti che gli animali non tendono affatto a ritornare a quel punto particolare della zona da cui si sono allontanati, ma alla zona ecologicamente equivalente per tutta la sua estensione. In acqua dolce la fuga è verso il mare (Terracini-De Benedetti, 1963): diluendo progressivamente l'acqua di mare si ottiene inversione di fuga per concentrazioni minori di 3,5‰ (Scapini).
Per semplicità, si considera qui solo il ritorno orientato verso il mare, che si può studiare statisticamente con un espediente assai semplice. L'orientamento - pressoché costante alle varie ore del giorno - permane immutato se si esclude la visione del paesaggio e del mare e pressoché immutato se l'esperimento viene compiuto a distanza dal mare o su rive con il mare dalla parte opposta o diversa.
L'orientamento è dunque essenzialmente indipendente da fattori orientanti locali (dovuti alla vicinanza del mare). Con un espediente pure molto semplice si può dimostrare che esso si basa sulla visione del sole. Poiché il sole è un punto di riferimento mobile, gli animali - come von Frisch e Kramer avevano già dimostrato rispettivamente per le api e gli Uccelli - debbono essere in grado di compensare la variazione di azimut solare, variando regolarmente nel corso del giorno il proprio angolo di orientamento con il sole, cioè la ‛direzione-norma' da assumere rispetto al sole: ad esempio, per una popolazione di una riva rispetto alla quale il mare sia a ovest, l'angolo (prescindendo da variazioni stagionali) varia da circa +180° al mattino, col sole a tergo, fino a circa 0° (sole di fronte) al tramonto passando per +90° circa a mezzogiorno (sole sulla sinistra).
Questa capacità è basata su un orologio interno, un ritmo circadiano sincronizzato con la rotazione terrestre dalla alternanza luce-buio: ciò si può dimostrare per Talitrus e altre forme dotate di simile meccanismo per mezzo di esperimenti di vario tipo, che comportano sfasamento del foto- periodo (Pardi e Grassi, 1955; esperimenti simili sono stati eseguiti per l'orientamento solare degli Uccelli), oppure rapido trasferimento ad altra longitudine (Papi, 1955; Pardi, 1955, per Tylos, un isopodo), o ancora arresto o accelerazione temporanea dell'‛orologio' mediante refrigerazione o esposizione ad alte temperature (v. Pardi, 1957).
Popolazioni di rive diverse hanno orientamento (‛direzioni di fuga') diverso e quindi cicli differenti di variazione dell'angolo, adeguati alla disposizione della propria riva. Ma il meccanismo dimostra altresì adattamenti precisi a condizioni geografiche diverse: in zone intertropicali, dove l'azimut solare si muove in senso orario e culmina a sud per una certa parte dell'anno, mentre durante l'altra parte si sposta in senso antiorario e culmina a nord, vi sono popolazioni di Anfipodi che sanno compensare altrettanto bene il moto orario e quello antiorario (Pardi e Ercolini, 1965). Non sappiamo ancora, se non per ipotesi, come avvenga il passaggio dall'una all'altra modalità di compensazione, ma lo studio dell'orientamento di specie intertropicali fornisce una prova importante dell'orientamento solare: quando la distanza del sole dallo zenit è inferiore a un certo limite (5-6°) gli animali non sono più in grado di ritrovare la direzione giusta e, nel loro habitat, tendono a insabbiarsi rapidamente (Ercolini, 1964). Lindauer (1957) aveva già osservato fenomeni confrontabili con questi nell'orientamento solare delle api.
Non v'è economia nei processi evolutivi quando il risultato è di importanza vitale e non sorprende che gli Anfipodi abbiano diversi altri mezzi a disposizione per assicurarsi il ritorno nel caso in cui non possano servirsi del sole; di giorno, se si trovano in ombra, o nelle ore del crepuscolo - ad esempio - gli animali possono orientarsi, come le api e molti altri Artropodi, con il pattern di luce polarizzata del cielo, che ha un moto apparente solidale con quello solare. Di notte è dimostrato (Papi e Pardi, 1953; Enright, 1961) che possono orientarsi con la luna, secondo un meccanismo di regolazione dell'angolo indipendente da quello solare e certo più complesso (orientamento lunare cronometrico). In una località equatoriale il disorientamento caratteristico nel momento in cui l'astro passa allo zenit si verifica prevedibilmente due volte per ogni ciclo sinodico (Pardi ed Ercolini, 1965).
Altri meccanismi che talitri e specie affini possono usare per ritrovare la loro zona di elezione non sono cronometrici: di giorno e di notte, a cielo coperto, la visione diretta del paesaggio (profilo della duna, della vegetazione, della scogliera, ecc.) ha un peso e così pure, in talune condizioni, la direzione del vento. Anche la pendenza del substrato ha importanza, poiché gli animali tendono a salire se la sabbia è bagnata, a scendere se è asciutta (Craig, 1973). Ercolini e Scapini (1974) hanno dimostrato come questa inversione della geotassia cooperi e/o interferisca con l'orientamento solare. Vi è pure - come fattore coadiuvante secondario - un ‛fattore locale' presumibilmente dovuto a eterogeneità visuali dipendenti dalla vicinanza del mare : animali tirrenici e adriatici, trattati nello stesso modo e saggiati contemporaneamente in esclusione di paesaggio sul Tirreno e sull'Adriatico, risultano statisticamente meglio orientati (sia per direzione che per concentrazione) sul ‛loro mare' che sul mare ‛opposto' (Pardi e coll., 1957).
Lo stesso meccanismo di orientamento zonale solare è stato dimostrato, dopo le ricerche iniziali su Talitrus, in molte altre specie terrestri o semiterrestri di Artropodi (Decapodi: Schöne, 1963; Altevogt, 1965; v. Herrnkind, 1972; Isopodi: Pardi, 1954; Araneidi: Papi, 1955; Insetti: Papi, 1955; Pardi, 1955; Ercolini, 1962) di rive marine, lacustri o fluviali. Per varie di queste forme è accertato anche l'orientamento lunare (nell'isopodo Tylos e nel ragno Arctosa) e sulla base della luce polarizzata. Più recentemente si è stabilito che un orientamento zonale solare esiste anche in specie francamente marine o limniche di acque sub-, circa- o infralitorali: Isopodi, Anfipodi, taluni Pesci e Anfibi. Alcuni Scaridi delle scogliere coralline si valgono probabilmente del meccanismo solare nei loro regolari spostamenti da acque litorali basse ad acque più profonde (Winn e coll., 1964), girini di rospi o di rane evitano nello stesso modo lo spiaggiamento (Ferguson e Lanreth, 1966).
Il problema delle componenti innate e acquisite del meccanismo solare non sempre è stato affrontato e risolto. Anche in tal caso gli Anfipodi (Pardi e coll., 1957-1960) e i ragni (Papi e Tongiorgi, 1963) sono a questo riguardo tra le forme più note. La capacità di utilizzare il senso interno del tempo nella compensazione del moto solare apparente è sicuramente innata in molti gruppi, ma negli Anfipodi sono geneticamente determinati entro larghi limiti anche il ciclo particolare di variazione dell'angolo adatto alla direzione della riva o, in una parola, la direzione utile di fuga, almeno per popolazioni di coste con disposizione stabile nel tempo e nello spazio: piccoli nati in laboratorio, che non hanno mai visto il sole, hanno direzioni di fuga statisticamente corrispondenti, anche se meno precise, a quelle della popolazione di origine (Pardi, 1960). Nei ragni del genere Arctosa, per contro, i giovani di tutte le popolazioni di rive differentemente disposte hanno direzione di fuga verso nord, che può essere eventualmente vantaggiosa come transecting unidirezionale (v. sopra, cap. 3, § a). Per apprendimento individuale, agevolato dalla pendenza del substrato e da altri fattori, alla direzione di fuga innata verso nord si sovrappone quella particolare, adatta alla riva su cui si trovano. In condizioni opportune gli animali possono apprendere nuove direzioni di fuga (Papi e Tongiorgi, 1963). Il coleottero stafilinide Paederus delle rive lacustri o fluviali apprende nuovamente la direzione utile (via dall'acqua) quando fuoriesce dall'inattività invernale (Ercolini e Badino, 1961).
La molteplicità dei meccanismi di orientamento a disposizione di molti organismi ripari e intertidali pone il problema della loro importanza funzionale relativa nelle condizioni naturali. Non sorprende che in gruppi diversi - a seconda dell'organizzazione e delle differenze eco-etologiche - si abbiano condizioni assai diverse. Nei granchi del genere Uca che hanno, per la complessità strutturale degli occhi e per le loro stesse dimensioni e postura, una possibilità di maggior controllo dell'ambiente visuale prossimo e remoto, la diretta visione della costa e un orientamento (telotattico e scototattico) verso oggetti salienti della riva hanno grande importanza nei movimenti orientati sulla spiaggia, forse ancor più dell'orientamento solare (v. Herrnkind, 1972). Il paguro Coenobita rugosus Milne Edwards usa probabilmente l'orientamento solare per ritornare al mare quando si trova nell'interno a una certa distanza dalla spiaggia, mentre le sue consuete migrazioni ritmiche secondo l'asse mare-terra sono guidate dalla visione diretta del paesaggio (duna, mare, ‛scototassia' verso oggetti salienti della spiaggia alta) e, in certe condizioni, da una anemotassia positiva (Vannini, 1975). Un orientamento solare cronometrico non è mai stato provato con sicurezza nei Molluschi, benché fotomenotassia e sensibilità alla luce polarizzata siano note per questo tipo, senza che se ne sappia la precisa utilizzazione in natura. Ricordiamo ancora i casi in cui, con il controllo attivo dei cicli di emersione dal fondo sabbioso, certi bivalvi (ad es. Donacidi) riescono a sfruttare come mezzo di trasporto (non come riferimento orientante!) le correnti di marea e il moto ondoso, compiendo migrazioni periodiche tidali di notevole estensione. Fenomeni comparabili - per ciò che riguarda lo sfruttamento di correnti di marea - sono noti anche in Crostacei propriamente intertidali come l'anfipodo Synchelidium, in cui il controllo circadiano delle reazioni alla pressione e alla luce permette il tempestivo affioramento dal fondo sabbioso e quindi il trasporto verso terra a marea ascendente e in senso opposto a marea calante (Enright, 1963). Nei Molluschi intertidali (Bivalvi, Gasteropodi) di substrati rocciosi a forte pendenza prevalgono risposte geotattiche nel mantenimento di zona. Un orientamento scototattico verso la silhouette scura della costa, soprattutto in caso di distacco dal substrato e allontanamento dalla costa per effetto del moto ondoso, è dimostrato per alcune specie (Littorina punctata L. e Nerita textilis Dill.; Chelazzi e Vannini, 1976), mentre in Littorina littorea L. il recupero di zona sembra fondato sulla capacità di utilizzare come riferimento direzionale la direzione di provenienza delle onde (Gewdrow, 1977).
Il sistema di guida visuale delle piccole tartarughe (Chelonia mydas L.), che debbono compiere una corsa lunga e pericolosa per raggiungere rapidamente il mare dal punto della spiaggia in cui si sono schiuse, è ancora diverso (Mrosovsky, 1967). I piccoli, che generalmente schiudono su spiagge in cui v'è netta differenza d'intensità tra il flusso luminoso proveniente dall'orizzonte aperto marino e quello di terra incombente e ricco di vegetazione, si orientano tropotatticamente in modo da ricavare una stimolazione luminosa egualmente intensa nei due occhi: ciò li conduce nella direzione giusta, anche se, inizialmente, non vedono il mare. Tale risposta è assolutamente innata. Benché altri cheloni (e forse gli stessi adulti di Chelonia mydas L.) dispongano del meccanismo solare cronometrico, nei piccoli sarebbe impossibile una sua utilizzazione, se non altro perché le madri possono deporre su spiagge comunque orientate.
Meccanismi dunque molto diversi per gruppi e situazioni diverse, come sottolinea Enright (1978); tuttavia, se il riferimento solare si è indipendentemente affermato in un così alto numero di forme litorali, dagli Invertebrati ai Vertebrati (marini o d'acqua dolce), ciò dev'essere dovuto evidentemente alla sua disponibilità per forme di habitat diverse e al suo alto grado di affidabilità, in una parola al forte vantaggio selettivo che offre. Per le specie sopralitorali (Anfipodi, Isopodi, ragni) il riferimento solare o celeste (luce polarizzata) assai raramente fa difetto. Non altrettanto sicuri sono gli altri sistemi: non la visione del paesaggio, che può essere cancellata o confusa da ostacoli grossi e piccoli, non la pendenza localmente variabile, non - a maggior ragione - la direzione del vento o delle correnti, o del moto ondoso. Che talune specie di questi gruppi ricorrano pure - durante la notte - a un orientamento astronomico ancora più complesso, come quello lunare, rafforza nella precedente opinione. Ciò non esclude che, in talune popolazioni e in circostanze speciali, altri meccanismi (ad esempio, la visione della costa saliente, la stessa pendenza del substrato) possano, in una situazione di conflitto, prevalere sul meccanismo solare (Hartwick, 1976). Nelle api domestiche troveremo fatti paragonabili a questi.
III. La migrazione anadroma dei salmoni. La storia naturale dei salmoni, di quelli del Pacifico in particolare, è assai ben nota (Hasler, 1954-1978). I salmoni (genere Oncorhynchus) si riproducono in autunno e inverno in acque interne fluviali o lacustri ben ossigenate e i giovani di una certa dimensione migrano poi a favor di corrente verso l'oceano. Qui passano un lungo periodo (da 2 a 3 anni) di vita pelagica (i salmoni del Pacifico soprattutto nel Golfo d'Alasca) fino a completare l'accrescimento.
Inizia allora una migrazione a rovescio di qualche migliaio di chilometri che riporta i salmoni alle foci del loro sistema di origine. Se la migrazione oceanica sia completa- mente o solo in parte attiva e quali siano i fattori che la guidano, è tuttora un gran mistero, benché alcuni autori - su basi indirette - sostengano l'intervento di un orienta- mento o di una navigazione solare. Si noti che l'orienta- mento cronometrico solare nei Pesci è stato supposto per taluni spostamenti in natura e dimostrato in esperimenti in cui si riesce ad addestrare certe specie a trovare un rifugio basandosi unicamente sulla visione del sole (Braemer e Schwassman, 1963 e altri). Una sensibilità alla luce polarizzata è stata dimostrata almeno per alcuni Teleostei (Groot, 1965; Waterman, 1972).
Vi sono comunque ottime prove che un buon numero di adulti in qualche modo ritrova e risale proprio il sistema fluviale nel cui alto bacino era nato qualche anno prima. Questa parte della vicenda è oggi assai più chiara, perché appare ormai dimostrato (Hasler, 1956) che, risalendo il loro sistema fluviale, gli animali riconoscono olfattivamente le qualità chimiche ‛uniche' delle acque provenienti dal loro bacino di origine e scelgono ogni volta il ramo fluviale ‛giusto'. Soggetti marcati e riportati a valle di una certa biforcazione non ritrovano la via e si disperdono a caso (a differenza dei controlli) se hanno le cavità nasali otturate. Esperimenti di condizionamento dimostrano d'altra parte che i salmoni sono dotati di olfatto sviluppatissimo e che possono discriminare egregiamente acque di fiumi diversi, oltreché quelle del proprio, nonché sostanze chimiche in diluizioni estreme. L'acqua del fiume originario determina risposte elettriche, a livello del bulbo olfattivo, molto più intense rispetto a quelle provocate da acque di varia origine.
Si suppone fondatamente che questa straordinaria capacità di ritorno a un'area circoscritta determinata (homing) sia dovuta a un imprinting, ovverosia un apprendimento irreversibile in una fase sensibile più o meno precoce della vita, di carattere olfattivo rispetto alle qualità chimiche delle acque del bacino originario. Ciò è confermato anche da alcuni esperimenti recenti (Hasler e Schuz, 1977) in cui salmoni giovani (marcati) furono esposti per un mese ad acque con morfolina o alcool feniletilico e quindi rilasciati. Durante la successiva migrazione anadroma la maggioranza dei soggetti già esposti a morfolina fu catturata in un ramo fluviale in cui veniva versata tale sostanza, e analogamente per i pesci esposti all'alcool feniletilico. Un gran numero di controlli vennero catturati per contro in località diverse.
L'orientamento dei salmoni nella migrazione anadroma deve considerarsi ‛mediato', secondo la nostra classificazione, poiché lo stimolo chimico, come in molti altri casi di cosiddetta chemotassia (v. Creutzberg, 1975), non è ‛orientante', ma solo ‛motivante': l'animale, una volta motivato, si orienta controcorrente (reotassia positiva), una forma speciale di orientamento unidirezionale.
Che l'animale possegga anche in questo caso, come in tutte (forse) le migrazioni aviarie, informazioni sulla distanza da percorrere è presumibile (ma non dimostrato), soprattutto sulla base di esperimenti riguardanti la migrazione di discesa, per cui si fa appello di nuovo all'intervento di una bussola solare (Johnson e Groot, 1963).
B. I sistemi pluridirezionali. Nei sistemi di questo tipo il raggiungimento mediato delle mete comporta sempre, anche per l'individuo singolo, una molteplicità di disposizioni possibili (angoli di orientamento) rispetto a riferimenti fissi e/o una molteplicità di cicli di variazione angolare rispetto a quelli regolarmente variabili come il sole. Considereremo a questo riguardo l'orientamento delle api, l'orientamento migratorio degli Uccelli, lo homing e la navigazione.
I. L'orientamento nelle api. Le ricerche magistrali di Karl von Frisch (uno dei più grandi biologi viventi) e della sua scuola hanno scoperto e chiarito i complessi sistemi di orientamento e di comunicazione in Apis mellifica L. e nelle specie congeneri. Benché siano state ampiamente divulgate, ne riassumiamo gli aspetti essenziali.
Mete ecologiche delle api sono le sorgenti di nutrimento e di acqua, l'alveare stesso - nel viaggio di ritorno dall'ambito di volo (alcuni chilometri di raggio) - e il nuovo posto di nidificazione al momento della sciamatura.
Una concentrazione imponente di individui qual è quella dell'alveare (da 20.000 a 80.000 adulti e varie migliaia di larve in accrescimento) impone un sistema rapido e tempestivo di sfruttamento e di afflusso delle risorse trofiche disperse su una larga superficie: tale sistema si basa sul reclutamento di altre numerose operaie foraggiatrici (‛novizie') non appena un'operaia esploratrice abbia scoperto una fonte trofica degna di attenzione.
La tecnica di studio è relativamente semplice: l'alveare è costruito in modo da poter vedere ciò che fanno le api sui favi. Gli individui in osservazione sono numerati con un codice speciale di macchie colorate. Per addestrare le operaie a raccogliere in un punto determinato (un tavolinetto a un piede con un vetrino da orologio contenente soluzioni zuccherine di concentrazione controllata) è sufficiente attrarre su un supporto con una goccia di miele qualche ape operaia e spostarsi gradatamente fino al punto voluto. Se anche un'ape soltanto ha seguito lo spostamento, di lì a poco - nello stesso luogo - ne arriveranno a decine.
Con questo metodo si dimostra innanzitutto che le api hanno un senso del tempo assai preciso, basato su un ritmo circadiano: fornendo cibo ad api marcate a un'ora determinata, le vedremo ricomparire in quel luogo nei giorni successivi, con grande puntualità. Il significato biologico è evidente: la produzione di nettare ha orari specifici e determinati.
Tre sono i metodi che le api hanno a disposizione per localizzare e ritrovare la meta: orientamento su caratteri salienti del paesaggio (‛pilotaggio visuale'), orientamento ‛olfattivo' e orientamento basato sulla ‛bussola astronomica' (sole e luce polarizzata del cielo).
L'orientamento del primo tipo ha grande importanza fin dalle prime sortite. L'operaia s'imprime allora, innanzitutto, insieme ad alcune caratteristiche visuali e olfattive dell'arnia che le serviranno a riconoscerla (‛orientamento immediato'), la posizione dell'arnia stessa rispetto ai punti di repere più prossimi (elementi del paesaggio). Ampliando poi gradatamente l'ambito di volo, l'operaia prende via via nota di altri riferimenti salienti e della loro posizione rispetto all'arnia. È dimostrabile che, già in questa fase, il reticolo di punti di repere a terra viene, per così dire, correttamente inserito nel campo orientante solare: pilotaggio e bussola solare si integrano, se c'è il sole. L'orientamento sulla base del paesaggio continua ad avere comunque grande importanza anche per le foraggiatrici ormai esperte. In certi casi, elementi del paesaggio linearmente estesi lungo la rotta di volo assumono tale salienza, da escludere perfino il riferimento solare.
L'orientamento olfattivo ha una funzione dominante nel reclutamento e nel foraggiamento quando la sorgente trofica è situata entro un raggio di poche decine di metri dall'arnia (il limite varia a seconda delle razze e delle specie). La bottinatrice che l'ha scoperta e che porta con sé l'odore specifico dei fiori (o, negli esperimenti con fonti artificiali, un profumo eventualmente aggiunto al substrato) può allora eseguire - a seconda di alcune condizioni dipendenti e dal rendimento delle fonti di cibo e dalle esigenze della colonia in quel momento - una danza di forma ‛circolare' sulla superficie dei favi: questa manovra allarma le compagne che la seguono attentamente, le induce a uscire e a cercare attorno all'alveare, in tutte le direzioni, una sorgente che abbia quello stesso particolare odore. Per la difficoltà di seguire il volo individuale, la precisa traiettoria di ricerca non è ben nota, ma è dimostrato che, al ritrovamento, partecipano elementi di orientamento immediato (odore e colore dei fiori) e mediato (visione delle altre api in raccolta, messaggi olfattivi sparsi attorno alla fonte dalle compagne con l'estroflessione dell'organo odoroso addominale - formato da ghiandole localizzate sul margine anteriore del VII tergite addominale - ed eventualmente correnti aeree che convogliano gli stimoli olfattivi).
Ma la probabilità di trovare una meta localizzata attraverso una ricerca in varie direzioni, pur agevolata nel modo che si è detto, diminuisce drasticamente con la distanza. Nelle api si è evoluto un sistema di orientamento e di comunicazione che sopprime (o riduce) la ricerca delle operaie per un largo raggio attorno all'alveare e la differisce nello spazio limitandola alla regione più o meno lontana in cui si trova la fonte trofica. Il sistema comporta tre fasi: a) rilevamento delle coordinate polari della fonte trofica da parte della foraggiatrice, assumendo l'arnia come punto zero; b) comunicazione dei dati rilevati alle compagne; c) utilizzazione delle informazioni da parte di queste ultime.
Non si perda comunque di vista che non si tratta di un sistema di rigore assoluto; che in tutte e tre le fasi vi è dispersione dell'informazione, per l'ovvia variabilità nel rilevare, nel valutare e nell'eseguire; e che il successo dipende anche dal numero degli individui coinvolti.
Premettiamo che quando un'operaia esploratrice cerca più o meno a lungo e con percorsi tortuosi ‛pilotandosi' nell'ambito di volo noto, è rapidamente in grado, una volta trovata una fonte redditizia, di ricavare una rotta pressoché rettilinea tra la fonte e l'arnia e viceversa. Al pilotaggio tende a sostituirsi rapidamente l'uso della bussola solare: l'operaia assume nel viaggio di andata un certo angolo utile con il sole, inverte la rotta di 180° dopo la raccolta facendo l'angolo supplementare e ritorna all'alveare (metodo della ‛inversione di rotta').
Ora, la distanza della meta viene misurata sulla base del dispendio di energia a cui l'operaia è sottoposta soprattutto nei percorsi di andata: con vento contrario in questo tratto o dovendo superare un pendio, l'ape misura e segnala distanze maggiori. Dal momento in cui il percorso fra la meta e l'arnia e viceversa si fa rettilineo, l'operaia è in grado di riprodurre nell'interno dell'alveare l'angolo memorizzato corrispondente al viaggio di andata: poiché lo sfruttamento di una fonte trofica può durare a lungo ciò comporta compensazione del moto solare apparente. L'orientamento solare è dunque cronometrico.
La comunicazione alle compagne dell'angolo solare utile avviene con la famosa ‛danza a otto' (o scodinzolante, Schwänzeltanz, tail-wagging dance) che la foraggiatrice ripete più volte sui favi, seguita con attenzione da una piccola schiera di compagne olfattivamente allarmate. Per fonti trofiche oltre una certa distanza critica, la danza a otto, con forme di passaggio, si sostituisce alla danza circolare.
Il ritmo della danza, tanto più rapido quanto più vicina è la meta, secondo una precisa relazione, informa sulla distanza. Il tratto diametrale della danza, durante il quale l'animale scodinzola ed emette suoni udibili, contiene l'indicazione direzionale. Se la manovra viene eseguita su un substrato orizzontale e in piena vista del sole e/o di parte del cielo azzurro - come avviene, di regola, in certe api esotiche o, eccezionalmente, nelle nostrane - l'operaia, percorrendo il tratto diametrale, indica direttamente col capo l'azimut della sorgente trofica, così come l'ago della bussola indica il nord magnetico: l'angolo che essa attivamente mantiene rispetto al sole (anche se si ruota il piano su cui poggia) corrisponde a quello utile per raggiungere la meta dall'arnia ed è l'angolo del ‛viaggio di andata'. Quando, come di regola nelle nostre specie, la danza viene compiuta in oscurità sulla superficie verticale dei favi l'operaia (che ha nella regione del collo organi di senso con cui controlla la posizione rispetto alla gravità) traduce l'angolo solare in un angolo rispetto alla gravità, secondo un codice semplice: ciò si dice ‛trasposizione fotogeomenotattica' ed è - con alcune varianti - una capacità fisiologica di base presente in vari altri insetti, pur senza avere valore di comunicazione. Angolo solare e gravitazionale, che indicano una sorgente trofica determinata, variano regolarmente nel corso del giorno e col mutare della posizione apparente del sole.
Le informazioni trasmesse con la danza vengono valutate con approssimazione efficiente dalle compagne che escono e tendono a raggiungere la meta indicata dopo aver seguito qualche ciclo di danza. Ciò è dimostrato dai noti esperimenti ‛a scala' e ‛a ventaglio'. A loro volta le operaie reclutate trasmetteranno ad altre compagne le informazioni ricevute e precisate dall'esperienza personale, diffondendo ‛la notizia'.
Le prove che l'orientamento è solare sono dirette e indirette. Già l'osservazione della variazione regolare dell'angolo di danza, a mano a mano che il sole cambia posizione apparente, è un forte indizio, ma decisiva è l'osservazione che le danze sul piano orizzontale sono completamente a caso (cioè col tratto diametrale comunque orientato) se si impedisce la visione del sole e del cielo. Ancora, le danze possono essere riorientate con una luce artificiale, poiché le api assumono rispetto a essa un angolo eguale a quello efficiente col sole di quel momento. Infine, in zone intertropicali, le danze sono disorientate, tanto su substrato orizzontale che verticale, quando il sole è troppo vicino allo zenit: il valore critico è una distanza zenitale di 3° (Lindauer, 1957); negli Anfipodi (Ercolini, 1962) il valore critico è superiore (5-9°).
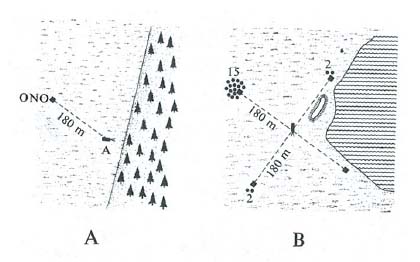
Indirette importanti verifiche vengono poi dagli esperimenti di ‛dislocazione' dell'alveare a piccola o grande distanza. Uno di questi è riportato nella fig. 13. L'esperimento è cruciale perché i presumibili riferimenti, per ritrovare la meta dall'arnia e l'arnia dalla meta, vengono esclusi tutti salvo uno: l'uso cronometrico del riferimento solare. Si può dire che la dislocazione estragga dal contesto spaziale locale in cui è stato costruito, il sistema di coordinate polari con il punto di partenza dell'animale al suo centro, liberandolo da ogni eventuale residua distorsione.
Gli esperimenti di dislocazione a grande distanza hanno poi permesso di dimostrare, analogamente a quelli compiuti da Papi (1955) per Talitrus e da Pardi (1955) per Tylos, che la compensazione del movimento solare è basata su un ritmo endogeno circadiano. Renner (1954) ha trasportato per via aerea un alveare da New York a San Francisco con un salto di circa 47° di longitudine, corrispondente a una differenza di circa 3h 15 min nel tempo medio locale. Siccome gli animali a San Francisco si regolano sul loro orologio interno (tempo di New York) e non sul tempo locale, essi hanno compiuto errori prevedibili nell'assunzione dell'angolo solare e perciò nella direzione di ricerca.
Di tutte queste complesse capacità che permettono lo orientamento e il reclutamento corretto, alcune sono strumenti comportamentali di base geneticamente fissati: orologio interno, capacità di utilizzare come riferimento il sole, la luce polarizzata e la gravità, automatica trasposizione dall'angolo solare all'angolo gravitazionale, forma e ritmo specifici (con differenze razziali) della danza sono innati. A differenza degli Anfipodi, le api debbono però imparare il particolare tipo del moto apparente solare corrispondente alla loro posizione geografica. Api trasportate in aereo da una zona in cui il sole ha moto apparente antiorario a una dove ha moto orario, si comportano come fossero di fronte al sole del luogo d'origine e compiono gli errori previsti, che imparano però a correggere dopo vari giorni. È possibile mantenere per lungo tempo una colonia in ambiente a luce artificiale: se poi si trasferisce all'aperto, le operaie che non hanno mai visto il sole dapprima non sanno valersene in alcun modo; successivamente imparano a servirsene con un orientamento fotomenotattico non cronometrico (facendo un angolo fisso col sole) e solo dopo qualche giorno sono in grado di compensare correttamente il moto apparente solare.
Il complesso delle ricerche sulle api (di cui si è potuto dare qui un'idea molto sommaria) costituisce un vero monumento dell'etologia e della fisiologia comparata. È comprensibile, perciò, l'interesse vivissimo (e quasi lo sgomento) del mondo scientifico, quando - in quest'ultimo decennio - alcuni ricercatori americani (Wenner e Johnson, 1967-1971 e altri), su periodici autorevoli e con grande rilievo, ne hanno posto in discussione la scoperta indubbiamente più significativa: il valore della danza a otto come linguaggio simbolico.
Secondo gli oppositori non vi sarebbero prove sicure che la danza comunichi direzioni e distanze: l'effetto di reclutamento attorno a una determinata sorgente di nutrimento ‛anche lontana' si spiegherebbe, per loro, altrettanto bene ammettendo che le novizie, ricevuta dalla foraggiatrice danzante un'informazione odorosa composita (odore proprio della sorgente e/o della località di raccolta), escano dall'alveare e cerchino l'odore giusto: un reclutamento, dunque, sempre olfattivo e senza indicazioni spaziali, che potrebbe eventualmente essere agevolato (ma non e molto chiaro come ciò avvenga) dagli odori che le compagne lasciano lungo la linea di traffico o sottovento a essa. In prossimità, poi, della sorgente, l'odore sparso dalle compagne e la stessa visione di altre raccoglitrici favoriscono l'approccio finale. Alcuni esperimenti a ‛scala' e a ‛ventaglio', simili a quelli di von Frisch, sembrano giustificare queste ipotesi.
La controversia tocca solo marginalmente il problema dell'orientamento, poiché nessuno pone in dubbio che le api si orientino con il sole o la luce polarizzata durante il volo e che - nell'oscuro dell'alveare - traducano l'angolo solare in un angolo con la direzione di gravità. Non si nega nemmeno che la danza abbia, nella forma, nel ritmo e nella disposizione, precise correlazioni con la distanza e la direzione della sorgente, ma solo il suo valore di informa- zione. Ora è proprio qui un aspetto singolare della vicenda, come molti hanno rilevato. Poiché si ha un bell'essere scettici sulla validità dell'argomento ‛teleologico' (se una struttura o un comportamento complesso ci sono, debbono servire a qualcosa), ma si dovrà pur riconoscere con Gould (v., 1976) l'altissima improbabilità che le specialissime correlazioni della danza siano sorte così per caso, senza che abbiano alcun valore adattativo. Né vale - ci sembra - l'obiezione formulata dallo stesso Gould che altri Apidi sociali (come le Melipone) effettuano, come hanno dimostrato Lindauer e Kerr (1958), una raccolta efficientissima con un reclutamento essenzialmente olfattivo. Verissimo, ma, per rimanere allo stesso esempio, alcune specie di Melipone hanno colonie persino più popolose delle api, pur essendo senza aculeo: gli è che si difendono in un altro modo, e non meno temibile di quello delle api domestiche. Sembra banale dover osservare che, anche in specie vicine, possono essersi evolute soluzioni diverse per lo stesso problema.
Nella sostanza, poi, come dimostra Gould, i risultati di Wenner e collaboratori sono solo apparentemente in disaccordo con la teoria di von Frisch e possono spiegarsi assai bene come dovuti alla particolare tecnica di addestramento usata da questi autori. Molti nuovi esperimenti critici, con controlli molto rigorosi circa la possibile interferenza di un reclutamento odoroso, hanno riconfermato ad abundantiam la correttezza delle vedute di von Frisch. Soprattutto probativi sono quelli in cui si inducono le foraggiatrici a dare informazioni sbagliate: con dosi subletali di parathion il ritmo della danza segnalatrice rallenta e le reclute cercano allora a una distanza minore di quella attesa. In altri esperimenti, con una luce artificiale, si riesce a dirottare a volontà la direzione del tratto rettilineo della danza, facendo compiere l'errore previsto nella direzione di ricerca delle novizie allarmate. La controversia ha avuto il merito di sottolineare l'importanza, del resto già nota, dell'orientamento olfattivo sia quando il cibo è vicino, sia nell'approccio finale di una sorgente lontana.
L'orientamento delle formiche, delle termiti, delle ve- spe pone problemi comparabili che, particolarmente per le formiche, sono stati profondamente studiati da molti autori (Jander, 1957). L'uso cronometrico del sole e della luce polarizzata sono stati accertati: l'orientamento delle formiche è peraltro dominato in larga misura dal complesso sistema delle tracce olfattive, di cui qui non possiamo occuparci.
L'orientamento delle api è ‛pluridirezionale' e ‛vettoriale': della meta (l'arnia) l'operaia conosce direzione e distanza, ma - dislocata al di fuori della rotta consueta di quel giorno - ritrova l'alveare solo nell'ambito di volo noto e con giri tortuosi che attestano la necessità di un riorientamento, senza dubbio con ricerca e poi pilotaggio su punti di repere noti (visuali, forse anche olfattivi). Dallo spazio esterno, invece, non riesce a tornare allo home range. Per quel che sappiamo, né l'ape né alcun altro insetto o invertebrato si è dimostrato mai capace di una vera ‛navigazione'.
II. L'orientamento migratorio degli Uccelli. È questo il problema forse più dibattuto nel campo dell'orientamento animale. Benché molte questioni siano tuttora insolute, negli ultimi decenni, a partire dalla scoperta dell'orientamento solare dovuta a Kramer (1950), sono stati realizzati progressi di grande rilievo: oggi sappiamo con sicurezza che i sistemi di guida fondamentali nelle migrazioni sono la bussola solare, stellare e magnetica.
Classifichiamo l'orientamento migratorio degli Uccelli come ‛pluridirezionale' e ‛vettoriale'.
Il carattere pluridirezionale è sottolineato dai fatti seguenti: 1) le rotte di migrazione appaiono in qualche caso pressoché rettilinee e uniformi, ma più spesso - soprattutto nelle migrazioni intercontinentali - presentano segmenti diversamente orientati, che sembrano ‛prevedere' a distanza le barriere geografiche, o si succedono, come accade ad es. nelle rotte dei migratori oceanici, per mutamenti direzionali in un punto pressoché costante del percorso, anche in mare aperto. I giovani di molte specie sanno assumere le corrette, successive direzioni anche alla loro prima migrazione senza gli adulti; 2) mentre i giovani dislocati sperimentalmente dalla rotta non sembrano capaci di correzione (e ciò ricorda un rigido orientamento unidirezionale), gli adulti, secondo molti esperimenti su varie specie, hanno tale possibilità (vera ‛navigazione'); 3) alla fine del viaggio migratorio, gli individui già esperti sono in grado di ritornare - con fedeltà estrema - a particolari località di nidificazione e di svernamento (homing).
Il carattere ‛vettoriale' risulta con evidenza da ricerche relativamente recenti su alcune specie (ma sembra lecita una generalizzazione): gli animali hanno meccanismi endogeni di controllo delle distanze da percorrere.
Rimandando al successivo punto III il problema della navigazione e dello homing, consideriamo ora soltanto la determinazione della direzione e della distanza nella migrazione.
a) Regolazione direzionale. I sistemi di guida fondamentali, si è detto, sono l'orientamento solare, stellare e magnetico. Il primo meccanismo è cronometrico, il secondo sembra possa esser cronometrico per alcune specie e non per altre o, nella stessa specie, in regioni diverse percorse dalla migrazione. L'orientamento magnetico non è cronometrico, essendo irrilevanti dal punto di vista direzionale le variazioni giornaliere.
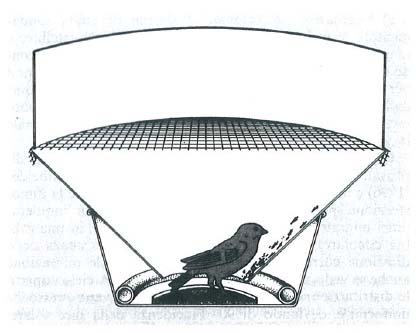
Che il sole avesse importanza nel mantenimento della direzione dei migratori diurni fu già supposto da Schneider (1906) e da Wachs (1926). Kramer (1950) ne dette la dimostrazione sperimentale. Osservando la cosiddetta ‛inquietudine' migratoria degli storni, egli si accorse che, in una gabbia circolare, gli animali saltellavano con insistenza nella direzione corrispondente a quella di normale migrazione anche se vedevano soltanto il sole e il cielo. A cielo coperto le distribuzioni dei tentativi di fuga diventavano pressoché uniformi e, deviando di 90° l'incidenza della luce solare, ruotavano di un angolo corrispondente.
Esperimenti in laboratorio con un sole artificiale ed esperimenti di sfasamento del fotoperiodo (Hoffmann, 1954) dimostrarono con chiarezza che l'orientamento era cronometrico: gli animali sapevano compensare il movimento apparente del sole.
Vi sono tuttavia specie migratrici notturne, e ancora Kramer (1950) dimostrò, nelle solite gabbie circolari, che capinere e averle (Sylvia atricapilla L. e Lanius collurio L.) sono orientate a cielo stellato, fortemente disorientate a cielo coperto. Sauer (1957) ebbe per primo l'idea brillante di eseguire gli esperimenti in un planetario e vide, per varie specie di Silvidi, che la direzione corretta autunnale o primaverile compariva anche con il cielo artificiale, mentre subentravano disorientamento e inattività con la cupola illuminata a luce diffusa. Di più, gli animali mutavano nel senso previsto la loro direzione preferenziale quando l'immagine della volta celeste veniva ruotata in modo che il nord stellare fosse, poniamo, a est o a ovest.
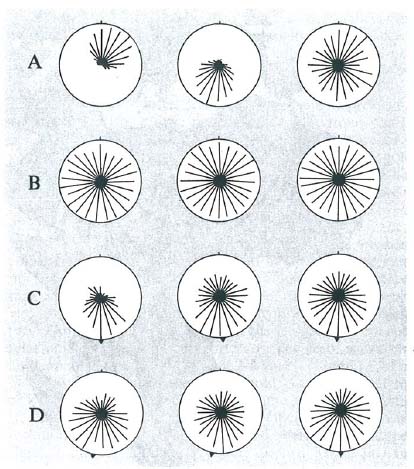
L'orientamento stellare, così dimostrato, può esser considerato un orientamento cronometrico, come quello solare dei migranti diurni o degli animali ripari? Gli animali si orientano perché compensano il moto apparente della volta celeste o di qualche stella o costellazione scelta per riferimento? La risposta, per alcune specie almeno, è negativa, poiché la loro direzione preferita permane immutata, anche se nel planetario si presentano le costellazioni nella posizione che avrebbero a ore diverse da quella dell'esperimento. E ancora, individui sottoposti a fotoperiodo anticipato o posticipato che, al sole, presentano i prevedibili errori direzionali, di notte sotto il cielo stellato naturale - sono ottimamente orientati. Per forme insensibili allo sfasamento temporale (Passerina cyanea L.) Emlen (1966; v., 1975) prospetta una seducente seppur complessa teoria: in un periodo sensibile, fra l'abbandono del nido e la prima migrazione autunnale, i giovani imparerebbero (è una sorta di imprinting) a riconoscere certe configurazioni stellari circumpolari che hanno minore velocità lineare nella loro rotazione e assumerebbero poi questa regione del cielo come punto di riferimento. Il metodo è dopo tutto assai più semplice di quello cronometrico, perché permette l'individuazione del nord stellare fintanto che quella zona del cielo è visibile, indipendentemente dall'ora e dalla stagione. La tecnica di rilevazione di Emlen è riportata nella fig. 17 e alcuni suoi esperimenti nella fig. 18. Fra gli esperimenti più probativi: la perdita di orientamento per occultazione delle stelle circumpolari nel planetario; l'assenza di orientamento nei soggetti a cui si è impedita, nella fase critica di apprendimento, la visione del cielo naturale o artificiale normalmente ruotante. Ma il risultato più impressionante si è ottenuto esponendo dei soggetti sin dalla nascita a un cielo artificiale con le stelle ruotanti attorno a Betelgeuse (la più luminosa di Orione): nell'esperimento critico, questi soggetti assumono, rispetto a Orione, la stessa direzione media che i controlli (esposti a un cielo normale) prendono rispetto alle costellazioni circumpolari.
L'esistenza di un orientamento stellare cronometrico non è tuttavia da escludersi completamente. Si suppone che ne facciano uso, in qualche modo, le specie a migrazione transequatoriale, che prima o poi devono necessariamente perdere di vista le stelle circumpolari.
L'esistenza di un terzo tipo di bussola, quella magnetica, cominciò ad apparire plausibile quando le osservazioni radar del flusso migratorio notturno accertarono che, in natura, la direzione corretta viene mantenuta, con una dispersione appena più alta, anche a cielo coperto o nell'interno delle nubi. D'altronde, la stessa analisi della distribuzione dei salti nelle gabbie circolari rivelava un residuo orientamento anche nei saggi a cielo coperto. La prima dimostrazione convincente della bussola magnetica, confermata in seguito da vari autori, si deve tuttavia a Merkel e Wiltschko (1965).
Non è una sorpresa che, anche in questo caso, i sistemi di guida siano molteplici. Al contrario è certo, più che probabile, che i tre ora ricordati non siano i soli: le grandi difficoltà del problema di orientamento che taluni migranti intercontinentali debbono risolvere, per le grandi distanze da coprire e la compensazione di numerosi fattori di disturbo, sono tali che vi sono ancora margini amplissimi per la ricerca futura di altri meccanismi di guida nonostante i risultati brillanti di questi ultimi decenni. Ad alcuni sistemi di guida che sembrano avere, per ora, una funzione solo marginale o ausiliaria, come certe regolarità nella direzione dei venti, i riferimenti topografici di varia scala, il punto di repere dato dall'azimut solare al tramonto e dalla persistente circoscritta luminosità crepuscolare, non possiamo che accennare.
Piuttosto, esiste anche in questo caso una gerarchia, come si usa dire, fra le varie strategie di orientamento?
Appare del tutto plausibile, a prima vista, una funzione semplicemente vicariante delle tre bussole: il sole per i migratori diurni, le stelle per i notturni quando il cielo è sereno, la bussola magnetica se sole e stelle sono invisibili. Ed è pure ammissibile un'integrazione fra i vari sistemi, che agevoli il risultato finale. Ma qual'è il fattore primario? In altri termini, dobbiamo ammettere un'indipendente e innata capacità di utilizzare tutti e tre i sistemi di guida - visto che anche i giovani alla prima migrazione sanno valersene - oppure solo rispetto a uno dei riferimenti gli animali ereditano la possibilità di orientamento corretto, trasferendo poi negli altri l'informazione direzionale primaria? Qui le opinioni sono divise. Per la migrazione notturna alcuni sostengono il ‛primato' dell'orientamento stellare, altri di quello magnetico. Esperimenti recenti, molto significativi, vengono a sostegno di quest'ultima opinione. Wiltschko e Wiltschko (1976) hanno dimostrato che l'inquietudine migratoria del pettirosso (Erithacus rubecula L.) è distribuita a caso di fronte a un cielo artificiale con un certo numero di ‛stelle' disposte secondo un disegno arbitrario, purché gli animali siano esposti a un campo magnetico così debole da non consentire un orientamento su quest'ultima base. Se, permanendo immutato il cielo artificiale, il campo magnetico viene portato a un'intensità adeguata, gli animali si orientano correttamente rispetto a esso e, successivamente, anche in un campo magnetico debole, con le sole stelle artificiali, saranno del pari ben orientati. L'esperimento indica con evidenza che la configurazione stellare è di per sé priva di valore, ma può acquistarlo e mantenerlo quando venga associata all'informazione direzionale data dal campo magnetico.
Il problema storico rimane: in quali circostanze si è affermata la capacità di assumere un certo orientamento efficace rispetto al riferimento primario, qualunque esso sia? Ma è un problema strettamente connesso a quello, estremamente arduo, dell'origine e dell'evoluzione del comportamento migratorio.
b) Regolazione delle distanze. È intuitivo che la migrazione, oltre a essere diretta in modo giusto, debba cessare al momento giusto: individui di specie differenti terminano il proprio viaggio in aree diverse anche alla prima migrazione e varie spiegazioni sono state addotte per questo fatto. Prospettive di grande interesse si sono aperte da quando Gwinner (1967) ha dimostrato che, in cattività e in condizioni costanti, l'inquietudine migratoria di specie diverse dura un numero di giorni più o meno fisso e adatto a coprire la distanza specifica fra quartieri nuziali e contronuziali. E ciò tanto se gli animali rimangono nell'area di partenza, quanto se vengono trasportati in quella di arrivo. Individui catturati e dislocati lungo la rotta di migrazione sono stati ricatturati oltre l'area consueta. Queste specie sembrano dunque possedere un programma endogeno e geneticamente determinato della durata di volo. Ma secondo ricerche di Gwinner e Wiltschko (1977) certe specie hanno per di più un programma interno di variazione dell'orientamento rispetto al campo magnetico, che corrisponde esattamente ai vari segmenti della rotta specifica. Il beccafico (Sylvia borin L.) ha una rotta autunnale dapprima verso SO nell'Europa centrale, poi verso SSE nella Spagna settentrionale e nel Sudafrica: registrando a intervalli regolari la direzione dell'inquietudine migratoria di giovani allevati ex ovo e tenuti in cattività, si è dimostrato che, esclusa la visione del cielo e col solo riferimento magnetico locale, la direzione prevalente ruota da SO a S a SSE in precisa corrispondenza, anche temporale, alla migrazione in natura.
Fatti corrispondenti, con la bussola solare, sono stati osservati da Johnson e Groot (1963) nella migrazione di discesa dei salmoni.
III. Homing e navigazione. Sono due termini che vengono spesso usati senza troppe esigenze di definizione: per alcuni autori, ad esempio, il mantenimento di zona basato sull'orientamento unidirezionale è homing. E spesso, particolarmente fra gli autori anglosassoni, navigazione è intesa come sinonimo di orientamento.
A nostro avviso home è il nido, il rifugio, il territorio individuale o anche, se si vuole, un'area circoscritta più estesa, ma comunque dotata di caratteri peculiari per cui è nota e preferita da uno o più individui, fra le molte possibili entro l'area ecologicamente adatta alle specie. Ogni animale che allontanato dalla home (attivamente o passivamente) vi ritorna o tende a tornarvi orientandosi verso di essa da qualunque direzione è dotato di homing. Lo homing è dunque, per definizione, ‛pluridirezionale'.
Con un esperimento di ‛dislocazione' lo homing si distingue subito dal mantenimento di zona (unidirezionale).
Di alcuni esempi di homing e dei loro meccanismi abbiamo discusso nelle pagine precedenti. A qualche altro converrà accennare, prima di esaminare il solo caso su cui siamo sufficientemente informati: lo homing del colombo viaggiatore (Columba livia L.).
Dei Mammiferi è certo largamente noto uno homing entro il normale ambito d'azione (uno homing ‛interno') basato sul riconoscimento di segnali visuali, spesso olfattivi. Tuttavia, nonostante una conoscenza sufficientemente precisa degli spostamenti naturali di certe specie nel loro ambiente, ben poco sappiamo dei loro sistemi di guida: ad esempio molti grandi Ungulati, come cervi, alci, caribù, zebre, gru e altri ancora, compiono regolari migrazioni che sicuramente comportano homing, ma a dispetto delle raffinate tecniche di radio-tracking, non molto ci hanno rivelato dei loro segreti. A maggior ragione sono spesso ancor più enigmatici gli homing che diremo ‛esterni', cioè dal di fuori dello home range. Ne sono capaci i Chirotteri, ma poco o nulla si conosce sui meccanismi di guida. Notizie di stampa su homing più o meno spettacolari di cani e gatti compaiono spesso, ma un controllo accurato della loro veridicità e delle loro condizioni manca del tutto. D'altronde, sempre per i Mammiferi, una sperimentazione più precisa su gatti e Roditori, basata sia sulla distribuzione delle direzioni assunte al momento del rilascio (libero e/o in strutture labirintiche radiali) a varia distanza dalla casa sia sugli esperimenti di radio-tracking, rivela solo deboli tendenze di ritorno ed è comunque insufficiente a fornire indizi sui meccanismi adoperati. A proposito di Roditori, Bovet (1977) parla di una ‟era prekrameriana" delle indagini.
Non sorprende che negli Uccelli, capaci di spostamenti individuali di dimensioni planetarie, si trovino i casi più impressionanti di homing presumibilmente esterno, da enormi distanze: il ritorno al nido sull'isola di Skokholm (Galles) di un esemplare di Procellaria puffinus Gm. da Boston, Mass., dopo un volo di 3.000 miglia per 12 giorni e mezzo, o quello degli albatros di Laydan (Diomedea immutabilis Rothsch.) da varie località (a distanze dell'ordine di 1.500- 4.000 miglia) all'isola Midway nel mezzo del Pacifico, sono i più spesso citati.
Per i Rettili, homing da grande distanza e di precisione estrema sono descritti nelle tartarughe marine (Chelonia mydas L.). Per riprodursi talune popolazioni, da vari tratti delle coste brasiliane, sanno raggiungere l'isola di Ascensione, un bersaglio di circa 7 miglia d'estensione in mezzo all'Atlantico, con un viaggio di circa 1.400 miglia marine: anche in questo caso abbiamo varie ipotesi ma scarsi dati di fatto sui meccanismi di guida.
Su distanze molto più modeste (qualche chilometro) sono capaci di homing Anfibi Anuri e Urodeli. I rospi (Bufo bufo L.), ad esempio, tornano fedelmente alla raccolta di acqua in cui sono nati, ma, nonostante studi accuratissimi (Heusser, 1963), il meccanismo di ritorno non è noto: i rospi ritornano persino al punto preciso della pozza natia, anche quando è stata prosciugata e livellata.
Per i Pesci, uno dei casi più profondamente studiati, quello dei salmoni, presenta tuttora - come si è detto - problemi irrisolti: per il percorso in mare aperto, dovendo l'individuo raggiungere una precisa foce fluviale, è difficile ammettere un semplice meccanismo unidirezionale.
Negli Insetti, a parte i casi profondamente analizzati di api e formiche, è molto ben nota la fase terminale dello homing al nido in vari altri Imenotteri - negli Sfecidi, ad esempio Philanthus, Ammophila, Bembix - basato su caratteristiche visuali prossime al nido (Tinbergen e Kruyt, 1938 ; Baerends, 1941) o anche relativamente remote, come caratteri cospicui dell'orizzonte ottico (van Iersel, 1978).
Uno homing è accertato anche in Crostacei: ad es., me- diante tracking sonoro si è stabilito che le aragoste Panulirus argus Latr., dislocate a qualche centinaio di metri, tornano al punto di cattura anche accecate, ma non si conosce come (Herrnkind e McLean, 1971).
Infine, nei Molluschi, un ritorno a casa su scala minima è quello delle patelle (Patella vulgata L. e altre specie) che compiono escursioni trofiche a partire da una piccola de- pressione sulla roccia (scavatasi per il logorio della conchiglia) e vi ritornano da direzioni diverse. Il meccanismo è discusso, ma un fattore di guida importante sono senza dubbio le tracce mucose lasciate dal mollusco nei suoi spostamenti (Cook e altri, 1969).
Questa breve rassegna dimostra come lo homing sia, dopo tutto, assai spesso enigmatico anche quando il ritorno implica brevi o brevissime distanze.
Quali sono i meccanismi oggi ritenuti teoricamente possibili? In un ordine che, senza troppe pretese, giudichiamo di crescente difficoltà possiamo elencare: 1) orientamento immediato; 2) pilotaggio per punti di repere noti; 3) orientamento alla bussola, pluridirezionale e vettoriale; 4) ricerca a caso o secondo configurazioni determinate; 5) navigazione vera (varie forme).
Diciamo innanzi tutto che tutti i metodi sono in teoria invocabili anche per uno homing interno, mentre uno homing esterno deve di necessità far ricorso a uno o a più d'uno dei più complessi sistemi da 3 a 5.
Prima di tentare una definizione della navigazione animale, ci sembra opportuno fare alcune brevi considerazioni sui primi quattro metodi di homing (che potremmo classificare ‛non navigazionali').
L'orientamento immediato ha un valore limitato, poiché presuppone il contatto sensoriale con la casa. Il pilotaggio sulla base di segnali visuali, olfattivi, tattili è stato largamente studiato nello homing interno di numerosi gruppi e ha sicuramente grande diffusione e importanza. L'orientamento pluridirezionale (plurivettoriale) alla bussola può realizzare assai semplicemente lo homing con la semplice inversione di rotta. In seguito a dislocamento attivo o passivo, se il riferimento (il sole, ad es.) è variabile per posizione, si ha naturalmente compensazione cronometrica della variazione di azimut. Ma abbiamo già visto quali siano i limiti del meccanismo solare direzionale nelle api: in un ambito ignoto un'eventuale dislocazione cospicua in senso trasversale rispetto alla rotta non viene compensata e l'animale manca il bersaglio. La ricerca a caso o secondo configurazioni programmate (ad es. seguendo una traiettoria a spirale a partire dal punto di lancio) ha un valore tanto più limitato con una meta circoscritta quanto più la meta è lontana, ma ha sicuramente importanza nello homing interno, perché può condurre l'animale a imbattersi in un riferimento noto: lo home range non è certo mai completamente esplorato.
Navigazione (nell'orientamento animale) è un termine - si è detto - usato con accezioni più o meno ampie. Una definizione ‛intermedia' potrebbe essere la seguente: è la capacità di pronto orientamento (mediato) verso una meta definita e circoscritta da qualunque punto (anche se non compreso entro l'ambito d'azione ‛direttamente' esplorato dall'animale e non attivamente raggiunto), ed è caratterizzata dalla correzione di ogni scostamento che abbia una componente trasversale rispetto alla rotta più economica. La definizione esclude quindi, oltre ai meccanismi 1, 2 e 4, anche l'orientamento unidirezionale o pluridirezionale di bussola, in cui può esservi correzione continua dell'angolo assunto, ma non vera correzione di rotta, se l'animale è unicamente affidato alla sua bussola.
Effetti attribuibili a capacità navigatorie nelle migrazioni aviarie sono stati rilevati molte volte con numerosi classici esperimenti di dirottamento dei migranti durante il loro viaggio. Ci è già noto l'esempio dei migratori oceanici. È fuor di dubbio che i meccanismi di navigazione, quali che siano, debbono avere un'importanza grandissima nelle migrazioni aviarie o d'altri animali su grandi distanze e verso aree relativamente circoscritte. Ma, mentre sui meccanismi di bussola nella migrazione siamo, come si è visto, sufficientemente informati, grazie soprattutto alle possibilità della sperimentazione sulla componente direzionale dell'inquietudine migratoria, i meccanismi di vera navigazione durante la migrazione ci sfuggono in gran parte, proprio per difficoltà pratiche sperimentali. È comprensibile perciò che gli studi sulla navigazione si siano rivolti allo studio molto più agevole dello homing, scegliendo soprattutto una specie domestica come il colombo viaggiatore (Columba livia L.), sempre disponibile e facilmente allevabile, meglio nota nella sua organizzazione sensoriale e neurale e per di più lungamente selezionata - per scopi pratici ben noti - proprio per le sue capacità di ritorno alla casa.
È ormai dimostrato che i colombi, anche assolutamente inesperti e senza alcun addestramento direzionale (come quello che si usa nelle gare in cui si effettuano lanci dalla medesima direzione, a distanze sempre maggiori), sanno orientarsi verso la colombaia e ritornarvi da località distanti molti chilometri, anche situate al di fuori dell'ambito sino ad allora direttamente esplorato.
La tecnica di analisi si basa principalmente su tre ordini di dati: I, la distribuzione dei punti in cui gli animali, liberati uno per uno e seguiti con un binocolo, svaniscono alla vista dopo il lancio (punti di sparizione). L'attendibilità di questo orientamento iniziale e le eventuali differenze fra lotti sperimentali e controlli vengono giudicate con i test statistici delle distribuzioni circolari; II, il numero degli arrivi in colombaia, la loro distribuzione nel tempo, il numero dei soggetti perduti (homing performance); III, la distribuzione delle località di ricattura di alcuni colombi non ritornati.
Migliaia di esperimenti non lasciano oggi il benché minimo dubbio sulla realtà del fenomeno che - abbastanza curiosamente - era stato negato nel passato da grandi ornitologi (Heinroth e Heinroth, 1941). I risultati permettono, d'altronde, di limitare l'eventuale intervento dei meccanismi 1 e 2 alla fase finale del ritorno: il contatto sensoriale con la casa è quasi sempre escluso; lo homing è, in molti casi, certamente esterno. Anche il meccanismo 4 (ricerca a caso o programmata secondo configurazioni determinate) può avere al massimo una funzione ausiliaria e - nella maggior parte dei casi - è escluso dal veloce tempo di ritorno. Torneremo sulla possibilità del meccanismo 3; consideriamo ora le principali teorie di navigazione proposte.
a) Navigazione inerziale. L'idea risale a Darwin (1873), e Barlow (1964) l'ha utilizzata per una teoria completa: gli animali sarebbero in grado di misurare e registrare, anche se spostati passivamente, ogni successiva accelerazione del movimento rettilineo o angolare durante l'andata e, integrando tutte le informazioni ricevute, stabilirebbero la propria posizione finale rispetto alla casa. La teoria incontra serie difficoltà nei risultati sperimentali: i colombi trasportati al punto di lancio in anestesia profonda o con lesioni chirurgiche degli organi vestibolari non differiscono dai controlli nell'orientamento iniziale e nel successo del ritorno.
b) Navigazione a due coordinate. Per alcuni è la navigazione stricto sensu. Parte dall'ipotesi che l'animale avverta la variazione di almeno due fattori (geofisici, astronomici) distribuiti secondo gradienti differentemente orientati, in modo che le isolinee dell'uno e dell'altro formino una sorta di griglia. Se l'animale dislocato è in grado di cogliere la differenza fra i due valori del punto di lancio e i due, memorizzati, corrispondenti alla casa, dovrebbe essere in grado di ritornare, solo che tenda a minimizzare la differenza rilevata.
La teoria di Yeagley (1947-1951) fa ricorso a due parametri geofisici, la variazione della componente verticale del campo magnetico terrestre e dell'intensità della forza di Coriolis: gli esperimenti non hanno portato alcun risultato a favore.
La più nota teoria di questo gruppo è quella di Matthews, astronomica. Kramer e Riese (1952) avevano già dimostrato che anche i colombi si valgono del sole come bussola. Matthews (1953) ipotizza un'utilizzazione assai più complessa del riferimento solare, simile a quella di un navigante. Requisiti necessari sarebbero un cronometro interno molto preciso e la capacità di estrapolare il punto di culminazione meridiana del sole, osservandone il movimento per un arco pur piccolo. Supponiamo, considerando due esempi opponibili, che l'animale sia stato trasportato (nel nostro emisfero) verso SE (o verso NO). Osservando il sole l'animale dovrebbe rilevare: 1) che il punto di culminazione è più alto (o più basso) di quello ricordato corrispondente a casa (in quella stagione); 2) che il sole, nella sua traiettoria, è un po' più alto (o più basso) di quanto dovrebbe essere, secondo l'orologio interno che segna l'ora di casa.
Lo spostamento latitudinale rilevato con 1) può essere annullato volando via dal (o rispettivamente verso il) punto di culminazione, cioè verso nord (o verso sud); quello longitudinale rilevato con 2) volando nello stesso senso del moto solare apparente (o in senso contrario), cioè verso ovest (o, rispettivamente, verso est). La combinazione vettoriale delle due tendenze dovrebbe dare la direzione almeno approssimativa di casa: nei due esempi opposti è NO o SE.
L'ipotesi presume una precisione talmente rigorosa di rilievi spaziali e temporali da risultare a priori poco credibile. Oltre a serie considerazioni teoriche vari esperimenti la contraddicono, ma uno appare cruciale: la manipolazione dell'orologio interno, con lo sfasamento del fotoperiodo (Schmidt-Koenig 1960 e 1972), di cui conosciamo già i risultati per l'orientamento solare alla bussola.
Un esempio: supponiamo che quattro lotti di colombi vengano sottoposti per alcuni giorni a un giorno artificiale ritardato di 6 ore (alba alle 12). Rilasciati a mezzogiorno da quattro direzioni diverse (N, S, E, O), secondo Matthews essi dovrebbero in tutti i casi dirigersi verso ovest (frecce bianche grandi): il sole nel luogo di lancio è infatti molto più alto di quanto essi si attendono sulla base dell'ora interna (6 del mattino) e ciò corrisponde, secondo la teoria di Matthews, a una dislocazione longitudinale di 90° verso oriente. In realtà i colombi si comportano in modo ben diverso: in tutte le località assumono un orientamento iniziale (frecce nere) che è ruotato di circa 90° in senso orario rispetto a quello teoncamente corretto verso casa (frecce bianche piccole). Una sola interpretazione sembra ammissibile: in ciascuna delle località essi hanno determinato, in qualche modo, la loro posizione rispetto alla colombaia e, valendosi del sole unicamente come bussola, hanno preso l'angolo giusto per tornare, come se il sole fosse nella posizione che l'ora interna suggerisce, cioè a est: di qui l'errore: ad es. il lotto dislocato a O va verso il sole (angolo d'orientamento 0°). Se il sole fosse a E, un angolo di 0° li riporterebbe a casa.
c) L'ipotesi di ‛mappa e bussola'. In contrapposizione a Matthews, Kramer (1953) aveva già supposto che la navigazione consistesse di due operazioni indipendenti nel punto di lancio: 1) stabilire la posizione del punto rispetto alla casa e quindi la direzione di casa (‛operazione di mappa'); in realtà, come ha fatto notare Papi (1974), la direzione di casa è di per sé sufficiente; 2) assumere e mantenere quella direzione valendosi di una bussola (‛operazione di bussola'). L'uso della bussola solare era già noto e gli esperimenti di sfasamento che abbiamo ora riferito convalidarono pienamente l'ipotesi krameriana. Nel 1971, Keeton dimostrò che la bussola solare poteva all'occorrenza esser sostituita, anche nel colombo, da quella magnetica. Animali lanciati a cielo completamente coperto si orientano molto bene verso casa, anche se il loro orologio interno è stato anticipato o ritardato. L'orientamento scompare se si assicura al collo una sbarretta magnetica o se l'animale viene deviato in modo prevedibile da piccole bobine di Helmholtz (applicate al collo e al capo) in cui la corrente fluisce in senso orario o antiorario. La componente ‛mappa' rimaneva tuttavia assolutamente enigmatica.
d) La teoria olfattiva. È merito di F. Papi e della sua scuola aver dimostrato di qual tipo di mappa gli animali possono effettivamente servirsi, grazie alla scoperta della navigazione olfattiva.
Individui sottoposti a resezione dei nervi olfattivi hanno orientamento iniziale fortemente disturbato e nettissima riduzione della capacità di ritorno, a differenza dei controlli (sottoposti a una falsa operazione). La resezione unilaterale con l'occlusione della narice ipsilaterale non disturba l'orientamento, mentre il disturbo compare, netto, se viene occlusa la narice controlaterale: l'effetto è quindi dovuto proprio alla deprivazione olfattiva e non già a un'influenza generale e aspecifica sul comportamento, come - poniamo - una soppressione della motivazione a tornare o una minorazione delle capacità di volo. D'altronde, animali operati bilateralmente rientrano ottimamente da località situate in una direzione determinata, da cui sono stati precedentemente addestrati a tornare, ma si perdono (a differenza dei controlli) se vengono liberati in una località sconosciuta a eguale distanza. L'olfatto appare dunque necessario solo per lo homing da località in cui gli animali non sono mai stati, mentre il ritorno lungo una rotta consueta può compiersi anche senza olfatto, con uno dei sistemi di bussola e con il pilotaggio entro un paesaggio visuale già noto. Infine, sostanze fortemente odorose applicate sulle narici prima del lancio hanno spesso effetti evidenti sull'orientamento iniziale e la capacità di ritorno. Tutti questi dati sperimentali dimostrano concordemente l'importanza dell'olfatto nello homing.
Era, a priori, assai poco plausibile che i colombi percepissero a distanze anche considerevoli (e persino sottovento) gli odori della voliera o della regione della voliera. La teoria prevede perciò un'utilizzazione assai più complessa dei riferimenti olfattivi che si fonda sulle proposizioni seguenti: 1) gli animali - stando in voliera o volando nei dintorni - imparano a conoscere non solo gli odori delle zone vicine direttamente esplorate, ma acquisiscono altresì - con i venti che spirano da direzioni diverse - una conoscenza indiretta degli odori che dominano nelle varie zone circostanti per un certo raggio più o meno ampio. Le informazioni olfattive portate dai venti vengono associate alla direzione da cui provengono mediante l'uno o l'altro dei sistemi di bussola (il vento di nord porta odori diversi da quello di sud, ad esempio, e così via): i colombi costruiscono così poco a poco una sorta di ‛mappa' del paesaggio olfattivo circostante, che non vi è motivo di supporre estremamente dettagliata; 2) quando i colombi vengono dislocati, riconoscendo l'odore dell'area in cui vengono a trovarsi, possono determinare approssimativamente la direzione di casa e pertanto assumere, mediante uno dei sistemi di bussola, la rotta utile per il ritorno; 3) informazioni olfattive di localizzazione nota per gli animali vengono raccolte anche durante il dislocamento.
Circa la natura degli odori la teoria dà una prevalente importanza a quelli di origine vegetale. Riassumiamo da una sintesi recente (v. Papi, 1976) alcuni degli esperimenti più significativi.
Prevenzione dell'associazione direzionale vento-odori. Un lotto di colombi (controlli) soggiorna per tre giorni ogni settimana in una gabbia a stecche orizzontali embricate che non consentono la visione del paesaggio ma lasciano passare i venti. Il lotto sperimentale, negli stessi giorni, è in una gabbia schermata che non consente nemmeno il passaggio dei venti. Negli altri quattro giorni, i soggetti sperimentali e i controlli stanno insieme in una grande voliera metallica o volano nei dintorni, portando (gli uni e gli altri) maschere che - comprimendo le narici - impediscono ogni percezione olfattiva. Soggetti sperimentali e controlli, che hanno perciò identiche esperienze visuali e di volo e identico stress, differiscono unicamente perché solo i controlli possono rilevare la direzione da cui provengono le informazioni olfattive. Al rilascio, i controlli sono ben orientati verso casa e rientrano assai velocemente, gli sperimentali hanno orientamento iniziale erroneo, rientrano più lentamente e alcuni si perdono.
Informazioni olfattive artificiali di direzione controllata. In un particolare corridoio orientato in senso N-S due lotti di giovani colombi (A e B) ricevono soltanto venti artificialmente prodotti con un ventilatore e aromatizzati con olio d'oliva e con trementina: il gruppo A riceve odore di olio d'oliva da sud e di trementina da nord, il gruppo B odore di olio d'oliva da nord e di trementina da sud. Dopo un certo periodo di questo pretrattamento, ambedue i gruppi vengono rilasciati a una certa distanza dalla voliera in direzione ENE, previa applicazione di olio d'oliva sulle narici di una metà degli individui di ciascun lotto e su quelle dell'altra metà di trementina. Nessuno dei due gruppi è mediamente orientato verso casa: i soggetti del lotto A si orientano prevalentemente verso nord se hanno subito l'applicazione d'olio d'oliva, preferibilmente verso sud con l'applicazione di trementina. Gli individui del lotto B si orientano in modo pressoché opponibile. Il risultato è in ottimo accordo con la previsione teorica 2. Esempio: per gli animali A, che hanno associato la direzione sud con l'odore di olio d'oliva e quella nord con la trementina, la percezione del primo odore, al momento del lancio, dà l'informazione direzionale ‛voliera a nord', quella del secondo ‛voliera a sud': di qui l'orientamento speculare dei due sottogruppi.
Rotazione controllata delle informazioni olfattive naturali. Con speciali deflettori applicati alla voliera si fa in modo che la direzione dei venti apportatori delle informazioni olfattive venga ruotata di 90° in senso orario o antiorario. Le differenze fra i controlli (voliera senza deflettori) e i due gruppi sperimentali, orario e antiorario, sono evidenti. Il procedimento ha provocato la rotazione della ‛mappa' olfattiva e le prevedibili deviazioni.
Détour. Due lotti di equivalente esperienza vengono trasportati, escludendo la visione del paesaggio, in un luogo lontano di lancio, presumibilmente al di fuori dell'ambito olfattivo noto, partendo dalla voliera in direzioni fortemente divergenti e facendo un lungo giro in senso opposto. In molti esperimenti siffatti, l'orientamento iniziale verso casa devia più o meno fortemente da quello atteso in senso orario o antiorario, a seconda del percorso seguito. L'effetto si può interpretare ammettendo che i colombi non raccolgano informazioni olfattive solo nel luogo di liberazione (come dimostrano gli esperimenti precedenti) ma anche durante il viaggio di dislocamento, soprattutto nelle zone più vicine alla voliera e perciò meglio note olfattivamente. Esperimenti simili con soggetti a narici occluse o trasportati in contenitori senza accesso dell'aria esterna, confermano la tesi: l'effetto détour manca e i colombi presentano, in esperimenti diversi, un orientamento che non differisce statisticamente dal casuale.
È evidente ormai (anche sulla base di recenti indipendenti conferme) che la possibilità di vera navigazione su base olfattiva è dimostrata: anzi è, questa, l'unica veramente dimostrata. Vi sono d'altronde indicazioni che altre specie posseggano capacità simili (Fiaschi e altri, 1974, esperimenti sui rondoni) e che la stessa alta fedeltà dei migratori a una specifica dimora, per cui si è generalmente fatto ricorso a un finale pilotaggio visivo, possa esser facilitata dall'orientamento olfattivo. Un'altra conseguenza importante degli esperimenti di Papi e altri, infine, è quella di aver attirato nuovamente l'attenzione sulla necessità di controllare le informazioni di varia natura che gli animali raccolgono quando si allontanano o vengono allontanati da casa.
Può un solo sistema navigazionale come quello olfattivo render pienamente conto dello homing del colombo? Se per la soluzione di problemi più semplici sono di norma disponibili meccanismi di guida multipli, a maggior ragione è da attendersi, in questo comportamento tanto più complesso, la validità della regola. Entro l'area direttamente o indirettamente nota, meccanismi ausiliari come la ricerca a caso di elementi noti del paesaggio olfattivo o visuale sono comunemente ammessi. Un pilotaggio su elementi visuali può essere invocato per i rari ritorni di animali anosmatici, così come a uno homing esclusivamente olfattivo si fa appello quando tale possibilità è ragionevolmente esclusa: Schlichte e Schmidt-Koenig (1971) e altri hanno dimostrato uno homing molto preciso, da brevi distanze, di animali che portavano lenti a contatto opache tali da rendere impossibile il riconoscimento di forme consuete da pochi metri.
Ma esperimenti recenti hanno aggiunto un elemento ulteriore di grande importanza. Ricerche di Papi e altri, 1977, Kiepenheuer, 1977, Wiltschko e Wiltschko, 1977, suggeriscono concordemente e con evidenza che i colombi - durante il dislocamento - usano anche informazioni derivanti dalla sensibilità al campo magnetico. È prematuro però inquadrare questi reperti recenti nei loro rapporti con la navigazione olfattiva e gli altri meccanismi di homing.
Infine un problema dibattuto è quello dello homing da località remote. Un'opinione critica diffusa tende a limitare la validità di una navigazione olfattiva solo entro un breve raggio (100 km circa). Ma nel colombo un orientamento iniziale verso casa sembra accertato e, per giovani inesperti, anche da varie centinaia di chilometri (Wallraff, 1970). Wallraff e Graue (1973) sostengono, pur con qualche incertezza, che persino colombi europei negli Stati Uniti (e americani in Europa) hanno tendenza iniziale giusta. Questi reperti - come del resto gli homings transoceanici di altre specie - suggeriscono subito l'esistenza di altri sistemi di navigazione a validità ‛planetaria'. È un'ipotesi che, se non può escludersi, attualmente non è obbligatoria. Mancava, difatti, in questi esperimenti a distanza, un controllo oggi necessario: un colombo potrebbe orientarsi anche in località remote, ricordando gli odori delle zone attraversate. E - d'altra parte - l'utilizzazione delle informazioni magnetiche durante il dislocamento potrebbe anche essere - come già si suggerisce - un orientamento alla bussola con inversione di rotta analogo, mutatis mutandis, a quello delle api.
4. Conclusione
Abbiamo considerato alcuni aspetti di ‛morfologia ecologica' dell'orientamento animale, escludendo deliberatamente i problemi strettamenti fisiologici. Pur con questa limitazione, la conclusione che si può trarre oggi da un esame anche sommario e per esempi è - come spesso accade - che i problemi irrisolti sono soverchianti. Nonostante i progressi notevoli, talvolta sorprendenti, degli ultimi decenni, si può affermare con Enright (1977) che ‟la ricerca moderna ha veramente scalfito appena la superficie [del vastissimo campo di studio che si occupa] delle prodezze diverse di cui sono capaci gli animali quando siano posti di fronte al problema critico di raggiungere un habitat favorevole o di rimanervi" e - aggiungiamo - di ritrovare la propria casa.
L'asserzione è presto giustificata; infatti: 1) solo per una esigua minoranza di specie abbiamo dati d'osservazione e sperimentali sufficienti sull'orientamento mediato in natura e in laboratorio; per la maggior parte dei casi osservazioni frammentarie in natura ed esperimenti spesso non conclusivi ci hanno rivelato ben poco; 2) abbiamo più di una prova che forme simili, in ambienti appena diversi, possono usare strategie differenti, e più di una prova - come si è rilevato più volte - che la stessa specie si vale di riferimenti diversi a seconda delle circostanze. Differenze costanti nell'uso prevalente dell'uno o dell'altro meccanismo per popolazioni diverse sono pure accertate. L'estensione delle ricerche rivela di continuo novità e impone revisioni e limitazioni; 3) per le stesse specie ‛predilette' dalla ricerca non possiamo assolutamente escludere che vi siano altri canali di informazione oltre a quelli noti (Kreithen, 1977). L'orientamento magnetico era tabù per il fisico, fino a qualche anno fa. Ora, a parte il problema irrisolto della recezione (su cui si stanno cimentando vari laboratori), il fenomeno in sé è incontrovertibile. Ne abbiamo viste le prove per la migrazione e per lo homing degli Uccelli. Nelle api la trasposizione fotogeomenotattica è influenzata in modo caratteristico dalle variazioni giornaliere del campo magnetico (Lindauer e Martin, 1968), che vengono utilizzate anche come segnatempo (Zeitgeber) dell'orologio interno (v. Lindauer, 1976); la costruzione orientata dei favi si perde alterando il campo magnetico. Nessuno avrebbe sospettato, qualche anno fa, che anche i Vertebrati (Pesci, Rettili, Anfibi, Uccelli) percepissero la luce polarizzata, che i colombi vedessero nell'ultravioletto, che avvertissero infrasuoni (Kreithen, 1977), che la loro mucosa olfattiva fosse uno dei principali rivelatori di raggi X (Keeton, 1974). Quale funzione possono avere questi parametri nell'orientamento non lo sappiamo, ma sarebbe un errore trascurarli ; 4) ancora per le forme ‛predilette', da quando lo zoologo ha smesso di cercarne ‛il' meccanismo d'orientamento e si è accorto che ne avevano diversi, il problema più urgente, spesso controverso, è divenuto quello di comprendere l'integrazione dei vari meccanismi in un sistema, di studiarne i rapporti e la variazione nel tempo: rapporti di vicarianza, di competizione, di cooperazione, di successione o di simultaneità, di dipendenza o di indipendenza. Studiare la variazione di tali rapporti significa allora studiare anche l'ontogenesi dell'orientamento, non sempre evidente, e ciò coinvolge la discriminazione fra componenti innate e apprese, una questione intricata che - per molte specie - dev'essere ancora chiarita. Solo con queste premesse potrà essere finalmente affrontato il problema dell'evoluzione dell'orientamento.
bibliografia
Batschelet, E., Statistical methods for the analysis of problems in animal orientation and certain biological rhythms, Washington 1965.
Carthy, J. D., An introduction to the behaviour of Invertebrates, London 1962.
Creutzberg, F., Orientation in space: animals. Invertebrates, in Marine ecology (a cura di O. Kinne), vol. II, part 2, London 1975, pp. 555-655.
Emlen, S. T., Migration: orientation and navigation, in Avian biology (a cura di D. S. Farner, J. R. King e K. C. Parkes), vol. V, New York 1975, pp. 129-219.
Fraenkel, G. S., Gunn, D. L., The orientation of animals, Oxford 1940.
Frisch, K. von, The dance language and orientation of bees, Cambridge, Mass., 1967.
Gould, L. S., The dance language and orientation of bees, Cambrige, Mass., 1967.
Gould, L. S., The dance language controversy, in ‟The quarterly review of biology", 1976, II, pp. 211-244.
Griffin, D. R., Bird navigation, in ‟Biological reviews", 1952, XXVII, pp. 359-390.
Hasler, A. D., Orientation and Fish migration, in Fish physiology (a cura di W. S. Moar e D. S. Randall), vol. VI, New York 1971, pp. 429-519.
Herrnkind, W. F., Orientation in shore living Arthropods, especially the sand fiddler crab, in Behavior of marine animals, vol. I, Invertebrates (a cura di H. E. Winn e E. L. Olla), New York 1972, pp. 1-55.
Holst, E. von, Mittelstaedt, H., Das Reafferenzprinzip (Wechselwirkungen zwischen Zentralnervensystem und Peripherie), in ‟Naturwissenschaften", 1950, XXXVII, pp. 113-132.
Jander, R., Die Phylogenie von Orientierungsmechanismen der Arthropoden, in ‟Verhandlungen der deutschen zoologishen Gesellschaft", 59. Jahresversammlung, 1965, pp. 266-306.
Jander, R., Ecological aspects of spatial orientation, in ‟Annual review of ecology and systematics", 1975, VI, pp. 171-188.
Johnson, C. G., Migration and dispersal of insects by flight, London 1969.
Keeton, W. T., The orientational and navigational basis of homing in Birds, in ‟Advances in the study of behavior", 1975, V, pp. 47-132.
Kramer, G., Experiments on Bird orientation, in ‟Ibis", 1952, XCIV, pp. 265-285.
Kühn, A., Die Orientierung der Tiere im Raum, Jena 1919.
Lindauer, M., Allgemeine Sinnesphysiologie: Orientierung im Raum, in ‟Fortschritte der Zoologie", 1963, XVI, pp. 57-140.
Lindauer, M., Evolutionary aspects of orientation and learning, in Function and evolution in behaviour (a cura di G. Baerends, C. Beer e A. Manning), Oxford 1975, pp. 229-242.
Lindauer, M., Orientation in animals. Old and new problems, in ‟Verhandlungen der deutschen zoologischen Gesellschaft", 69. Jahresversammlung, 1976, pp. 156-183.
Loeb, J., Forced movements, tropisms and animal conduct, Philadelphia-London 1918.
Mast, S. O., Light and behavior of organisms, New York-London 1911.
Matthews, G. V. T., Bird navigation, Cambridge 1968.
Mittelstaedt, H., Grundprobleme der Analyse von Orientierungsleistungen, in ‟Jahrbuch 1966 der Max-Planck-Gesellschaft zur Förderung der Wissenschaften", 1966, pp. 121-151.
Newell, R. C., Biology of intertidal animals, London 1972.
Papi, F., The olfactory navigation system of the homing pigeon, in ‟Verhandlungen der deutschen zoologischen Gesellschaft", 69. Jahresversammlung, 1976, pp. 184-205.
Pardi, L., L'orientamento astronomico degli animali: risultati e problemi attuali, in ‟Bollettino di zoologia", 1957, XXIV, pp. 473-523.
Pardi, L., Innate components in the solar orientation of littoral Amphipods, in ‟Cold Spring Harbor symposia on quantitative biology", 1969, XXV, pp. 395-401.
Pardi, L., Papi, F., Kinetic and tactic responses, in The physiology of Crustacea (a cura di T. H. Waterman), New York 1961, pp. 365-399.
Schmidt-Koenig, K., Current problems in Bird orientation, in Advances in study of behaviour (a cura di D. Lehrman, E. Hinde ed E. Shau), vol. I, New York 1965, pp. 217-278.
Schmidt-Koenig, K., Migration and homing in animals, Berlin 1975.
Schöne, H., Orientation in space: animals. General introduction, in Marine ecology (a cura di O. Kinne), vol. II, part 2, London 1975, pp. 499-553.
Schöne, H., Forms and mechanisms of orientation in space, in Encyclopaedia of ethology (a cura di K. Immelmann), New York 1977.
Wiltschko, W., Kompassysteme in der Orientierung von Zugvögeln, in Informationsaufnahme und Informationsverarbeitung im lebenden Organismus (a cura di M. Lindauer), Wiesbaden 1973.