
Ontogenesi dell'apprendimento
Ontogenesi dell'apprendimento
Per un neonato di mammifero la capacità di apprendere, ossia di stabilire relazioni causali tra eventi e di modificare il proprio comportamento in base a tali esperienze, riveste un enorme valore adattativo a fronte delle possibili e imprevedibili variazioni dell'ambiente. A partire dai primi anni Settanta l'affermazione di una teoria ecologica dell'apprendimento animale, che riconosceva le limitazioni specie-specifiche ed età-dipendenti nei processi associativi, ha dato un grande impulso agli studi ontogenetici di tali processi. La scelta di procedure di condizionamento adattate alle caratteristiche motorie e sensoriali dell'animale immaturo ha rivelato insospettate capacità di apprendimento e memoria nell'animale neonato, che appare in grado di apprendere e conservare la memoria di eventi significativi prima che le strutture cerebrali superiori, come la corteccia e l'ippocampo, abbiano raggiunto la piena maturità funzionale. Anche nella specie umana, l'apprendimento neonatale, se non addirittura prenatale, riveste un'importanza determinante per l'organizzazione futura del sistema nervoso centrale e del combortamento.
Introduzione
In molte specie di mammiferi, il neonato dipende, per un periodo relativamente lungo, dalle cure parentali prima di poter sopravvivere autonomamente. Lo sviluppo ontogenetico è un processo graduale e continuo nel corso del quale, dalla nascita al raggiungimento dell'età adulta, il comportamento di un individuo va incontro a cambiamenti più o meno rapidi, in una interazione dinamica tra fattori di maturazione endogeni e sollecitazioni dell'ambiente circostante. In tale processo, la capacità di apprendere, ossia di stabilire relazioni causali tra eventi e di modificare il proprio comportamento in base a tali esperienze, riveste un enorme valore adattativo perché consente al giovane animale di rispondere con una relativa flessibilità comportamentale a possibili anche se imprevedibili variazioni dell'ambiente (Klopfer, 1988). La capacità di apprendere si basa sulle straordinarie caratteristiche di plasticità del sistema nervoso centrale dei mammiferi, che già nelle prime fasi postnatali è in grado di svolgere funzioni molteplici e complesse, dalla selezione degli stimoli significativi alla trasduzione sensoriale di questi stessi stimoli, dalla formazione e l'immagazzinamento di un ricordo fino al recupero di quello stesso ricordo e alla esecuzione della appropriata risposta motoria.
L'apprendimento precoce ha un'importanza fondamentale nel porre le basi della organizzazione futura del comportamento di un individuo. Tuttavia, fino agli anni Settanta gli aspetti ontogenetici non hanno ricevuto una grande attenzione da parte degli studiosi dei processi di apprendimento e memoria. Nella seconda metà del 20° secolo, lo studio dell'apprendimento animale ha risentito fortemente dei contrasti fra la scuola etologica europea e quella comportamentista cui facevano riferimento molti psicologi sperimentali statunitensi. Gli psicologi comportamentisti sostenevano l'utilità dell' analisi del comportamento animale effettuata in laboratorio in un numero limitato di specie (ratti, piccioni, scimmie) e in ambienti controllati (labirinti, gabbie di Skinner, e così via). In questo contesto, essi tentavano di categorizzare i processi di apprendimento in base al fatto che la modificazione del comportamento fosse indotta dall'associazione tra uno stimolo e un rinforzo (come nel condizionamento classico o pavloviano), o tra una risposta e un rinforzo (come nel caso del condizionamento operante o strumentale). L'obiettivo della psicologia sperimentale era quello di stabilire leggi generali che spiegassero tutti i processi di apprendimento associativo e che fossero applicabili a tutte le specie animali. La formulazione di queste leggi poteva essere garantita solo dall'impiego di procedure sperimentali le più artificiali possibile, che minimizzassero il contributo dei comportamenti tipici della specie in esame. Nello stesso periodo gli etologi si interessavano invece alla descrizione funzionale del comportamento animale in un contesto naturale. La premessa teorica della scuola etologica europea di cui K. Lorenz è stato il rappresentante più noto, era quella che l'evoluzione, attraverso la selezione naturale, favorisse la trasmissione dei fenotipi comportamentali nello stesso modo in cui favoriva la trasmissione della caratteristica anatomica più adatta. L'etologia classica, perciò, si concentrò sullo studio dei cosiddetti moduli fissi di azione (FAP, Fixed Action Patterns). Tali moduli sono considerati comportamenti innati e stereotipati che vengono attivati in un dato stadio dello sviluppo dell' animale in seguito all'esposizione a un particolare stimolo o serie di stimoli. La capacità di alcune specie di apprendere particolari risposte rappresentò un importante settore di studio per gli etologi, ma, tranne poche eccezioni (imprinting nel pulcino, apprendimento canoro negli uccelli, abituazione), gli esempi di apprendimento in natura non furono mai sottoposti a una rigorosa analisi sperimentale. Per gli etologi l'apprendimento era un processo che non poteva essere separato da altre caratteristiche biologiche della specie, consistendo in un insieme di abilità specializzate evolutesi indipendentemente in ciascuna specie per svolgere una determinata funzione adattativa. Pertanto, era considerato assurdo il tentativo di ricondurre tutti gli esempi di apprendimento a leggi generali valide per tutte le specie animali (Zolman, 1983). Queste due posizioni teoriche rimasero per molto tempo distanti. Alla fine degli anni Sessanta tuttavia, alcuni psicologi sperimentali suggerirono che dovesse essere adottato un approccio più naturalistico allo studio dell'apprendimento animale (Shettleworth, 1988). l risultati di alcuni esperimenti di condizionamento classico e strumentale condotti in laboratorio indicavano infatti l'esistenza di limiti biologici alla associabilità tra stimoli, rinforzi e risposte, dal momento che la storia filetica di una data specie influenza profondamente ciò che un animale può apprendere e ricordare. In particolare, venivano messi in discussione due dei fondamenti teorici della teoria comportamentista: il principio di equipotenzialità, secondo il quale un animale può apprendere qualsiasi associazione tra eventi, e il principio della contiguità temporale, per cui l'associazione tra eventi ha luogo solo se essi si presentano quasi simultaneamente. Nel 1966 vennero pubblicati i risultati di un esperimento di J. Garcia e R.A. Koelling sull'avversione gustativa condizionata nel ratto. Questi due autori dimostrarono che i ratti sviluppavano un'avversione per una soluzione di acqua e saccarina se l'ingestione era concomitante con l'esposizione alle radiazioni o con l'iniezione di apomorfina o cloruro di litio (trattamenti che producono nell'animale i sintomi di un avvelenamento da cibo), ma che tale avversione non si stabiliva se il rinforzo negativo era uno stimolo di natura esterocettiva come uno shock elettrico. Inoltre, nell'avversione per uno stimolo gustativo, l'associazione tra lo stimolo condizionato e quello incondizionato si verificava anche in assenza di contiguità temporale tra i due stimoli. Perciò, lo sviluppo di un'avversione condizionata dipendeva dal normale comportamento alimentare della specie ratto che, in natura, è in grado di apprendere a evitare cibi che possano risultare tossici anche a distanza di ore dall'ingestione. Veniva così dimostrata l'importanza delle predisposizioni specie-specifiche nei processi di apprendimento, per le quali un animale apprende con maggiore facilità ad associare due eventi che abbiano maggiore probabilità di presentarsi secondo una relazione causale anche nel mondo reale. Successivamente R.e. Bolles (1970) dimostrò che la capacità di apprendere, in un test di evitamento attivo o passivo che richiede all'animale di inibire una risposta di fuga oppure di spostarsi da un compartimento all'altro di un'apparecchiatura per evitare una punizione, era profondamente influenzata dal repertorio innato di risposte difensive del ratto. Più in particolare, quando il paradigma sperimentale richiedeva l'emissione di risposte appartenenti al repertorio specie-specifico del ratto per evitare una punizione, come correre o saltare, l'apprendimento della risposta di evitamento si verificava con successo. Al contrario, quando il comportamento richiesto per evitare la punizione era quello di abbassare una leva, l'apprendimento della risposta di evitamento era difficile da ottenere.
L'esistenza di limiti biologici all'apprendimento ha rappresentato una delle maggiori sfide alla teoria tradizionale dell'apprendimento, e ha costretto molti sperimentatori a riconsiderare la metodologia dei test impiegati in laboratorio in modo tale da tenere conto della storia evolutiva e delle caratteristiche ecologiche della specie studiata. D'altra parte la tradizione etologica ha dovuto riconoscere l'utilità degli studi di condizionamento classico e strumentale condotti in laboratorio, che hanno rivelato una flessibilità insospettata nelle capacità associative delle specie con un repertorio comportamentale apparentemente limitato. In definitiva, l'incontro tra l'approccio psicologico e quello etologico all'apprendimento nel decennio compreso tra gli anni Sessanta e Settanta ha dato un grande impulso alla biologia dell'apprendimento e della memoria e ha stimolato l'interesse per l'ontogenesi di tali processi (Zolman, 1983).
L'approccio ecologico all'ontogenesi dell'apprendimento
Il riconoscimento dell'importanza delle predisposizioni specie-specifiche nei processi di apprendimento ha favorito un approccio sostanzialmente diverso anche allo studio dello sviluppo del comportamento. Il punto di vista tradizionale, che considerava l'animale giovane come una versione incompleta e inferiore dell'adulto con un repertorio comportamentale semplificato, aveva relegato gli aspetti ontogenetici ai margini delle scienze del comportamento. Infatti, fino all'inizio degli anni Settanta il problema centrale della ricerca ontogenetica era stato quello di determinare in quale stadio dello sviluppo le prestazioni di apprendimento e memoria dell'animale raggiungono quelle dell'adulto, e se le esperienze precoci potessero influenzare o no le capacità cognitive dell'animale adulto. In tale prospettiva, le capacità associative dell' animale in via di sviluppo venivano saggiate con i paradigmi normalmente usati nell'adulto, con l'ovvio risultato che le capacità di apprendimento del neonato e del giovane risultavano limitatissime o addirittura inesistenti.
Apparve invece chiaro ad alcuni sperimentatori che l'animale immaturo si confronta con problemi sostanzialmente differenti da quelli incontrati dall'adulto. In particolare, nelle specie di mammiferi a prole inetta, il piccolo nasce cieco e con un sistema nervoso ancora largamente immaturo, non è in grado di regolare la propria temperatura corporea, di muoversi efficientemente né di nutrirsi autonomamente, e necessita, pena la sopravvivenza, delle cure materne fino all'età dello svezzamento. L'universo sensoriale di un ratto o di un gattino appena nato è molto diverso da quello dell'adulto, e ovviamente il suo repertorio comportamentale è funzionale a risolvere i problemi specifici di un organismo immaturo, come per esempio restare in contatto con il corpo materno, fonte di nutrimento e di calore, mediante stimoli termotattili e olfattivi. In un tempo relativamente breve, le relazioni ecologiche dell'animale in via di sviluppo cambiano radicalmente: tali cambiamenti si accompagnano spesso a una completa riorganizzazione dell'organismo e del comportamento, così come accade, secondo l'efficace metafora usata da P.P.G. Bateson (1981), nella metamorfosi da bruco in farfalla. Così come, sostiene Bateson, sarebbe assurdo considerare un bruco come la versione incompleta di una farfalla, è fuorviante ricercare ogni volta le origini di un elemento comportamentale dell'adulto nel suo corrispettivo infantile. Per esempio, il comportamento alimentare di un mammifero si modifica radicalmente con lo svezzamento, ma la suzione non può essere considerata una versione 'inferiore' del comportamento alimentare dell'adulto.
Visto in tale prospettiva, lo sviluppo del comportamento non è più un processo attraverso il quale il piccolo diventa via via più 'capace' o più 'abile', ma piuttosto un processo dinamico nel corso del quale l'individuo vede il mondo da una prospettiva che si modifica progressivamente con il modificarsi delle proprie relazioni ecologiche e con il maturare dei propri sistemi sensoriali e motori (Spear, 1984). L'ontogenesi neurocomportamentale non risulta più basata solo su processi sequenziali di maturazione, come sostenuto dai precedenti modelli biodeterministi, ma anche, e in egual misura, sulle opportunità di apprendimento che si presentano al neonato nel microambiente iniziale del nido già al momento della nascita, se non addirittura durante la vita prenatale.
Gli studi condotti a partire dall'inizio degli anni Settanta, che hanno utilizzato in larga parte specie di roditori a prole inetta come il ratto e il topo, hanno rivelato tutta la ricchezza e la complessità del repertorio comportamentale dei neonati di mammiferi. Capacità di apprendimento e ritenzione assai complesse sono emerse con la scelta di procedure di condizionamento che tenessero nel dovuto conto sia l'ecologia della specie in esame che le caratteristiche motorie e sensoriali dell'animale immaturo. L'approccio ecologico all'ontogenesi dell'apprendimento, sottolineando il valore adattativo del repertorio comportamentale del neonato, ha prodotto una vera e propria rivoluzione metodologica: i ricercatori hanno messo a punto paradigmi sperimentali adattati alle specifiche capacità del piccolo, nei quali gli stimoli olfattivi e termotattili sostituiscono, nelle procedure di condizionamento classico, gli stimoli uditivi o visivi. Una cura analoga è stata posta nella scelta sia delle risposte comportamentali con le quali misurare l'apprendimento tra quelle normalmente espresse dal neonato nelle prime fasi postnatali quali approach (avvicinarsi, in genere alla madre), sniffing (annusare l'ambiente), probing (ricercare il capezzolo), sia dei rinforzi con i quali indurre un condizionamento strumentale (Zolman, 1983; Campbell, 1984; D'Udine e Alleva, 1984).
Ontogenesi dell'apprendimento nei roditori
La maggior parte degli studi ontogenetici sull'apprendimento e la memoria è stata condotta sulle due specie di roditori a prole inetta più utilizzate dalle scienze comportamentali, il ratto e il topo. l neonati di queste due specie, a somiglianza di altre specie di mammiferi, nascono con un sistema nervoso immaturo e con un aspetto poco dissimile da quello fetale. Il roditore neonato vive in un universo olfattivo e tattile, e sulla base di questo tipo di stimoli localizza la superficie del ventre materno per attaccarsi a uno dei capezzoli e cominciare ad alimentarsi. Durante le due prime settimane di vita postnatale la maggior parte delle attività del piccolo è organizzata in relazione alle funzioni nutritive. Con la maturazione del senso della vista e dell'udito e lo sviluppo dei meccanismi di termoregolazione il repertorio comportamentale del piccolo si arricchisce notevolmente nel corso della terza settimana di vita, fino ad assomigliare, allo svezzamento (21⁰ giorno), a quello di un adulto di dimensioni più ridotte. Nonostante l'aspetto fetale, nelle prime ore dopo la nascita il piccolo di roditore apprende e memorizza informazioni determinanti per la propria sopravvivenza, principalmente di carattere olfattivo, e tali processi di apprendimento presentano un notevole grado di flessibilità.
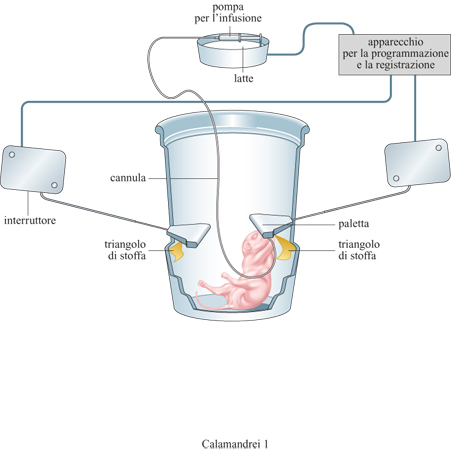
Un numero rilevante di studi ontogenetici ha dimostrato l'importanza delle esperienze olfattive precoci nella maturazione neurocomportamentale dei roditori. Il segnale olfattivo che guida il neonato verso il capezzolo è l'odore del liquido amniotico, che la madre, leccandosi, distribuisce sui capezzoli durante il parto. Questo stimolo olfattivo viene riconosciuto dal piccolo come familiare, in quanto appreso durante la vita intrauterina. P.E. Pedersen e collaboratori (1983) dimostrarono che la preferenza per l'odore delliquido arnniotico è appresa e non rigidamente determinata. Infatti, se nel corso della gravidanza viene iniettata nell 'utero una piccola quantità di essenza di limone, i piccoli alla nascita sembrano preferire i capezzoli marcati con questo stesso odore rispetto a quelli marcati con liquido amniotico. Ratti neonati di un solo giorno di età ai quali sia stata impiantata una cannula intraorale attraverso la quale viene somministrato il latte, imparano a orientarsi verso una paletta per ricevere il latte (fig. 1) e a distinguere con successo tra due palette che sono state contrassegnate da differenti odori (Johansone Hall, 1979). A una sola settimana di vita, i piccoli di ratto imparano a percorrere un labirinto a Y e, sulla base di segnali olfattivi, scelgono la direzione giusta per raggiungere la madre anestetizzata posta al termine del labirinto.
Lo sviluppo e la ritenzione a lungo termine di una preferenza olfattiva per un odore artificiale sono stati ottenuti in seguito all'associazione di particolari segnali olfattivi con l'infusione intraorale di latte. È inoltre possibile (Rudy e Cheatle, 1977) indurre in ratti di soli 2 giorni di età un'avversione condizionata per un odore, qualora l'esposizione a tale odore sia associata a un malessere gastrointestinale indotto da una sostanza tossica, come il cloruro di litio. L'avversione condizionata per un odore artificiale si sviluppa anche nella vita prenatale, come dimostrato dai primi studi di Pedersen e collaboratori (1983), e sembrerebbe mantenersi fino all'età adulta.
Come si è visto, l'utilizzazione di stimoli olfattivi e di risposte comportamentali tipiche del repertorio del neonato, come la suzione e l'orientamento verso la madre e il nido, ha permesso di rivelare capacità associative addirittura nella fase prenatale. L'impiego di test di condizionamento più complessi, che richiedano al piccolo di utilizzare informazioni spaziali per orientarsi in labirinti a Y, T, o radiali, o di test di evitamento attivo o passivo, evidenziano capacità di apprendimento inferiori nell'animale immaturo rispetto all'adulto, date le ovvie limitazioni motorie. Ma qualora le procedure sperimentali vengano modificate tenendo nel dovuto conto le caratteristiche sensoriali e motorie del piccolo, anche in questo tipo di test si osservano buone prestazioni di apprendimento. Per esempio, prima dello svezzamento i ratti e i topi hanno più difficoltà dell'adulto nell'apprendere una risposta di evitamento attivo o passivo. Questo tipo di paradigma, comunemente impiegato per saggiare le capacità associative nell'adulto, richiede all'animale di inibire una risposta di fuga, oppure di spostarsi da un compartimento all'altro di una specifica apparecchiatura per evitare una punizione, di solito un leggero shock elettrico.
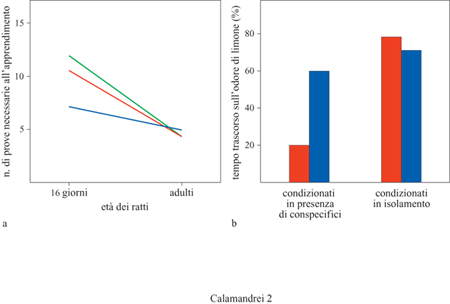
Le difficoltà nell'apprendere un compito di evitamento sono in larga parte relative all' espressione della risposta condizionata, ostacolata sia dall' elevato livello di attivazione provocato nell'animale immaturo dallo shock elettrico, sia dalle ridotte capacità di orientarsi per individuare la direzione di fuga. L'eliminazione della componente direzionale della risposta di fuga (il piccolo deve evitare di toccare le pareti di un'arena circolare per evitare la punizione), e la sostituzione dello shock elettrico con stimoli avversivi meno stressanti quali una corrente di aria fredda, riducono notevolmente l'età alla quale si manifesta l'apprendimento (Stehouer e Campbell, 1980). Anche il contesto nel quale vengono saggiate le capacità associative del giovane riveste un'importanza determinante: è stato osservato che un ambiente di test reso più simile alla nicchia ecologica del piccolo favorisce l'espressione di capacità associative a un'età più precoce. G.J. Smith e N.E. Spear (1978) hanno dimostrato il valore incentivante degli stimoli familiari, come l'odore del nido o la presenza dei conspecifici, nel modulare sia le prestazioni di apprendimento di ratti di 16 giorni in un test di evitamento passivo sia la formazione di un'avversione condizionata per un odore (fig. 2).
L'amnesia infantile
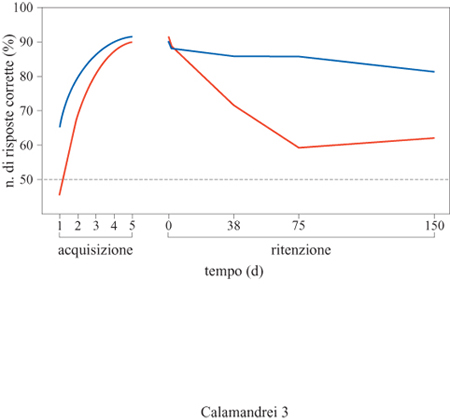
I risultati sperimentali degli ultimi vent'anni hanno dimostrato che i neonati di mammifero hanno una straordinaria capacità di apprendere basata su un sistema nervoso plastico e pronto a essere modificato dall' esperienza. Da questi stessi studi emerge che nel corso dello sviluppo il neonato e in genere il giovane hanno capacità di memoria inferiori a quelle dell'adulto, e che la capacità di conservare ed esprimere una risposta condizionata migliora progressivamente con l'età dell'animale, un fenomeno che è stato definito come amnesia infantile (fig. 3). Come si vedrà più avanti, il fenomeno dell' amnesia infantile è stato ed è tuttora un problema molto dibattuto anche dagli psicologi interessati allo sviluppo cognitivo umano. Molti autori hanno ipotizzato che questa ridotta capacità di memoria sia conseguenza dell'immaturità del sistema nervoso centrale. Essi hanno suggerito che i meccanismi di plasticità neuronale, che hanno luogo nel corso dei processi di apprendimento e di immagazzinamento dell'informazione appresa, non siano ancora perfettamente funzionali durante lo sviluppo e che le capacità di memoria analoghe a quelle dell'adulto siano strettamente dipendenti dalla maturazione più tardiva di strutture cortico-limbiche, come l'ippocampo e la corteccia (Nadel e Zola-Morgan, 1984). L'ipotesi dell'immaturità neurologica dei sistemi psicofisiologici di memorizzazione sarebbe sostenuta dal fatto che nelle specie di roditori a prole precoce, nei quali il sistema nervoso centrale è già maturo alla nascita, come la cavia (Cavia porcellus), le capacità di apprendimento e memoria non differiscono nel giovane e nell' adulto (Campbell, 1984).
Altri autori sostengono che più che nella immaturità delle strutture nervose, la spiegazione delle inferiori capacità di memoria del neonato e del giovane sia da ricercare nella rapidità con la quale cambiano le sue relazioni con l'ambiente durante il processo di crescita. L'animale in sviluppo sarebbe caratterizzato da una minore selettività nell'apprendere, e presterebbe una diversa attenzione agli stimoli presenti nell'ambiente in virtù di disposizioni dipendenti dall'età e della particolarità dei propri canali sensoriali (Spear, 1984). Lo stesso contesto viene recepito in modo completamente diverso a età differenti, e questo può costituire un fattore che può fuorviare la valutazione delle capacità di ritenzione. Questo punto di vista sembrerebbe essere sostenuto dal fenomeno della riattivazione (reinstatement) in base al quale le memorie del neonato e del giovane non sono perse una volta per tutte, in quanto comportamenti appresi che sono normalmente dimenticati rapidamente dall'animale immaturo possono essere conservati se gli animali sperimentano ripetizioni periodiche e parziali dell' esperienza iniziale, di per sé insufficienti per ristabilire l'apprendimento. Le procedure di riattivazione si sono dimostrate efficaci anche per favorire la ritenzione dell'esperienza appresa nel neonato umano.
Verosimilmente, tutti i meccanismi descritti contribuiscono alle minori capacità mnemoniche del giovane. Negli anni Ottanta la psicobiologia dello sviluppo si è concentrata, in particolare, sull'analisi dei meccanismi neuronali che regolano l'apprendimento precoce con l'obiettivo di trovare correlati neurofisiologici che spiegassero le differenze tra le capacità di memoria a lungo termine del neonato e quelle dell'adulto. Ciò che appare chiaro dai risultati degli studi ontogenetici è che lo sviluppo della memoria a lungo termine non è un processo unitario mediato dalla progressiva maturazione di strutture corticali. Piuttosto, ciascun sistema sensoriale ha la capacità di decodificare e conservare informazioni indipendentemente dalla maturità di altri sistemi sensoriali e centrali. Come si vedrà nei due esempi che seguono, differenti sistemi di memoria maturano in momenti diversi nel corso dello sviluppo, sulla base dei requisiti relativi alla fase ontogenetica dell'animale. Alla formazione di specifiche capacità associative concorrerebbero perciò canali sensoriali diversi, nonché aree cerebrali e regolazioni neurochimiche anch'esse differenti.
Basi neuronali delle preferenze olfattive precoci
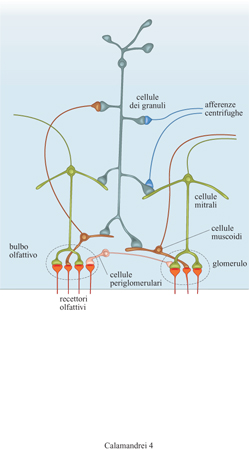
Come precedentemente descritto, nei roditori la memoria per alcune esperienze olfattive precoci viene conservata fino all'età adulta e riveste un ruolo importante per lo stabilirsi delle relazioni sociali, sessuali e parentali nell'adulto. l meccanismi plastici cruciali per la formazione delle preferenze olfattive avrebbero luogo nel bulbo olfattivo, una struttura telencefalica particolarmente sviluppata nelle specie macrosmatiche come i roditori, a uno stadio dello sviluppo nel quale le strutture superiori, come l'ippocampo e soprattutto la corteccia, sono ancora in un relativo stato di immaturità funzionale. Il bulbo olfattivo riceve un input sensoriale dai recettori olfattivi situati nella cavità nasale, che trasducono i segnali chimici in segnali neuronali. Nel bulbo gli assoni dei recettori olfattivi stabiliscono contatti sinaptici con i dendriti di due particolari classi di neuroni, le cellule mitrali e le cellule muscoidi, formando strutture approssimativamente sferiche, dette glomeruli. Un singolo glomerulo, o gruppi di glomeruli, costituirebbero unità funzionali in risposta a particolari stimoli olfattivi. Esternamente allo strato dei glomeruli, un terzo tipo cellulare, le cellule dei granuli, forma contatti sinaptici con le cellule mitrali e quelle muscoidi modulandone l'attività, ed è a sua volta il terminale di varie afferenze centrifughe al bulbo (fig. 4).
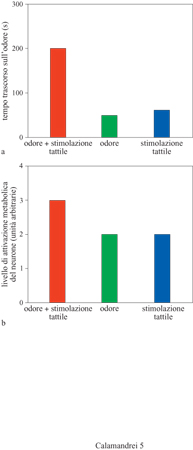
Quando nel corso della prima settimana di vita piccoli di ratto vengono ripetutamente esposti a un odore artificiale, per esempio a quello di menta, tendono a sviluppare una preferenza per quello stesso odore. Questa preferenza si stabilisce più facilmente se, contemporaneamente all'esposizione all'odore, i piccoli ricevono una stimolazione tattile simile a quella normalmente ricevuta dalla madre. A 20 giorni di età i piccoli preferiscono l'odore precedentemente associato con la stimolazione tattile rispetto a un altro odore anch'esso familiare, e mostrano segni evidenti di attivazione comportamentale (fig. 5). M. Leon e collaboratori hanno osservato che tale preferenza comportamentale era associata ad alterazioni fisiologiche e neuroanatomiche dello strato glomerulare del bulbo. In particolare, in seguito all'esperienza olfattiva associata alla stimolazione tattile si verifica un incremento sia nel numero che nell'attività metabolica dei neuroni glomerulari. Un'analisi più dettagliata ha inoltre dimostrato che le relazioni sinaptiche tra i diversi tipi di neuroni del bulbo olfattivo si modificano, cosicché la presentazione successiva dell'odore al quale l'animale è stato condizionato evoca un segnale elettrico alterato in uscita da una particolare regione del bulbo verso altre strutture cerebrali coinvolte nel processo associativo, come l'amigdala e la corteccia piriforme o entorinale. Tali variazioni neurofisiologiche sembrerebbero essere permanenti, ma hanno luogo solo se l'esposizione all'associazione odore/stimolazione tattile si verifica nella prima settimana di vita postnatale, e sarebbero modulate dall'innervazione noradrenergica al bulbo olfattivo attraverso un'azione inibente delle cellule dei granuli sugli altri tipi cellulari del bulbo (Wilson e Sullivan, 1994).
Questo modello di plasticità neurocomportamentale presenta molte similitudini sia con altri fenomeni di plasticità neurocomportamentale nel corso dello sviluppo, come per esempio lo sviluppo della dominanza oculare nel sistema visivo del gatto, sia con alcuni esempi di apprendimento olfattivo nel roditore adulto, suggerendo così che i meccanismi di plasticità coinvolti nell'apprendimento olfattivo sono simili a età diverse. Una struttura relativamente semplice, come il bulbo olfattivo, può consentire la formazione di memorie olfattive a lungo termine prima che i circuiti cortico-limbici coinvolti nella formazione di associazioni nell'adulto diventino pienamente funzionali.
Maturazione del controllo colinergico del comportamento
Alcuni tipi di apprendimento, che implicano l'integrazione di informazioni più selettive sull'ambiente e la costruzione di una rappresentazione spaziale del contesto ambientale in cui avviene il test, maturano relativamente più tardi nel corso dello sviluppo. Tra i substrati neuronali potenziali candidati alla regolazione di questi processi di apprendimento c'è il sistema colinergico centrale. l neuroni colinergici centrali utilizzano il neurotrasmettitore acetilcolina e sono raccolti in una serie di nuclei sottocorticali (nuclei della base, nuclei del setto e nucleo caudato) che forniscono la maggiore innervazione colinergica alla corteccia e all'ippocampo, due aree cerebrali considerate critiche nei processi di apprendimento e memoria (O'Keefe e Nadel, 1978).
Il sistema colinergico centrale raggiunge la maturità funzionale intorno al 21⁰ giorno di vita sia nel topo che nel ratto, ossia al momento dello svezzamento, anche se già intorno alla fine della seconda settimana di vita tale sistema va incontro a un salto maturativo da un punto di vista neurochimico e anatomico, segnalato dallo stabilirsi delle connessioni setto-ippocampali e dall'incremento della sintesi del neurotrasmettitore acetilcolina. Parallelamente, tra la seconda e la terza settimana postnatale i test che richiedono l'inibizione di una risposta, come l'evitamento passivo, evidenziano capacità di ritenzione per almeno 24 ore, mentre migliora notevolmente la capacità di apprendere compiti che richiedono all'animale di integrare le informazioni spaziali provenienti dall'ambiente. Infatti, è solo intorno all'età dello svezzamento che si evidenzia un profilo di abituazione simile a quello dell'adulto quando l'animale si trova in un ambiente sconosciuto, profilo caratterizzato da un'intensa attività esplorativa all'inizio del test e da un successivo decremento di tale attività. In generale, la maturazione del sistema colinergico sembrerebbe coincidere con la maturazione di meccanismi di attenzione più selettiva, che consentono all'animale di integrare quante più informazioni è possibile sulle contingenze del test, e successivamente di richiamare tali informazioni una volta posto nello stesso contesto. Non è un caso che nei paradigmi di condizionamento classico il fenomeno della inibizione latente si verifichi solo a partire dal18÷20° giorno di età: se un animale deve associare uno stimolo con un rinforzo avversivo (per esempio, un suono con uno shock elettrico) in un dato contesto ambientale (per esempio, una gabbia per l'evitamento) la pre-esposizione a quell'ambiente di test in assenza della punizione interferirà con il successivo apprendimento dell'associazione fra i due eventi. L'inibizione latente presuppone perciò che le caratteristiche dell'ambiente siano registrate, conservate e richiamate. In definitiva, tutti quei paradigmi di apprendimento che richiedono informazioni sul 'quando' e sul 'dove' uno specifico evento si possa verificare sembrerebbero diventare possibili solo dopo la maturazione delle afferenze colinergiche alla corteccia e all'ippocampo e raggiungerebbero livelli adulti con la maturazione dell'ippocampo stesso (Nadel e Zola-Morgan, 1984).
La manipolazione sperimentale del sistema colinergico, tramite somministrazione di farmaci che ne bloccano l'attività, o lesioni delle connessioni setto-ippocampali o dell'ippocampo stesso, induce infatti anche nell'animale adulto difficoltà nell'apprendere e ricordare un compito di memoria spazi aie. Recentemente è stato osservato come la maturazione neurochimica e funzionale dei neuroni colinergici centrali sia regolata in fasi critiche dello sviluppo dall' azione trofica, tropica e differenziativa dell'NGF (Nerve Growth Factor, fattore di crescita nervoso) e di altre neurotrofine. Queste molecole hanno un ruolo chiave nella regolazione dello sviluppo ontogenetico dei vertebrati, controllando la crescita e il differenziamento cellulare in vari sistemi dell'organismo. Nel sistema nervoso centrale, in particolare, l'NGF sintetizzato in momenti critici dello sviluppo dai neuroni ippocampali e corticali sembrerebbe dirigere la crescita delle afferenze colinergiche dal setto e dai nuclei basali. La manipolazione sperimentale dei livelli endogeni di questo fattore neurotrofico ha l'effetto di rallentare o accelerare la maturazione della funzione colinergica, e produce effetti acceleranti o peggiorativi anche sulla maturazione delle capacità di ritenzione di un test di evitamento passivo (Calamandrei e Alleva, 1996).
Ontogenesi dell'apprendimento nei primati umani e non umani
La teoria ecologica dell'apprendimento animale ha influenzato significativamente anche lo studio dello sviluppo cognitivo umano. La psicologia dello sviluppo tentava di applicare al bambino i principi dell'apprendimento e della memoria derivati da studi condotti sull'adulto, senza un vero interesse per lo sviluppo in sé. Gli psicologi comportamentisti consideravano l'ontogenesi comportamentale umana un processo determinato e organizzato in larga misura dall'apprendimento, dal condizionamento e dall'esposizione a modelli sociali, e prestavano scarsa attenzione ai processi maturativi endogeni. Si deve al grande psicologo svizzero J. Piaget, che non a caso aveva avuto una formazione biologica e zoologica, l'aver sottolineato la specificità dei processi cognitivi del bambino. Le teorie di Piaget, osteggiate in quanto troppo deterministe, hanno molto in comune con le teorie ecologiche dell'apprendimento animale: nello sviluppo umano si procede per stadi continui, ma distinti l'uno dall'altro, e soprattutto distinti dallo stadio adulto. Perciò, gli stadi cognitivi dell'infanzia sono considerati da Piaget unici e non mere versioni incomplete delle abilità cognitive adulte, e le capacità associative del bambino sono determinate da proprietà endogene e specifiche dello stadio maturativo dell'organismo.
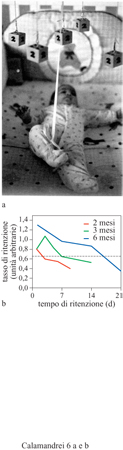
l primi studi sistematici sui processi associativi nel neonato umano risalgono ai primi anni Settanta, e consistettero in esperimenti sull' attenzione e l'abituazione agli stimoli visivi che dimostravano la capacità di riconoscere stimoli familiari già alla nascita. Successivamente, anche sotto la spinta dei risultati ottenuti in modelli animali grazie all'evoluzione delle metodologie sperimentali, sono stati utilizzati test di condizionamento classico od operante adattati alle specifiche capacità sensoriali e motorie dei neonati. Risposte comportamentali come la suzione, la vocalizzazione o l'orientamento della testa possono essere condizionate utilizzando diversi rinforzi. Per esempio, a sole 24 ore dalla nascita i neonati umani sono in grado di apprendere a modulare l'intensità e la frequenza di alcuni comportamenti (per esempio, la suzione) al fine di ascoltare il suono della voce materna (DeCasper e Fifer, 1980). C. Rovee-Collier e collaboratori (1990) hanno messo a punto un test di condizionamento operante nel neonato umano, nel quale è il neonato stesso, con il proprio comportamento, a determinare l'intensità del rinforzo. In questo test il neonato, a partire dai due mesi di vita, impara a muovere una giostra colorata sospesa sopra la culla e collegata con un nastro alla propria caviglia, determinandone la frequenza e l'intensità del movimento (fig. 6). Con il procedere delle sessioni i movimenti della gamba a cui è collegata la giostra diventano sempre più efficaci, essendo il premio rappresentato dal movimento stesso della giostra. Il test di ritenzione, che consiste nel misurare la reazione del bambino in termini di frequenza dei movimenti della gamba alla presentazione della giostra ferma e non collegata alla caviglia del neonato, ha luogo da 1 a 42 giorni dopo la sessione di apprendimento. l neonati di 2 mesi dimenticano l'esperienza in un intervallo di 1÷ 3 giorni, mentre neonati di 3 mesi mostrano di ricordare la risposta operante per 6÷8 giorni dopo l'apprendimento. Le capacità di memoria a lungo termine aumentano con l'età dei neonati fino a raggiungere circa i 20 giorni nei bambini di 6 mesi.
Anche nel neonato umano, le capacità di memoria sembrerebbero migliorare con l'età così come descritto nell'animale. Analogamente, la tecnica della riattivazione sembra funzionare anche nel neonato umano: la presentazione per soli 3 minuti di un elemento del test, 24 ore prima del test di ritenzione, nello stesso contesto nel quale era avvenuto il condizionamento (per esempio, la stessa giostra mossa dallo sperimentatore) migliora le prestazioni dei neonati di 3 mesi fino a 4 settimane dopo la sessione di acquisizione. Perciò, come nell'animale, la memoria di eventi appresi non sembrerebbe definitivamente persa nel neonato umano, ma potrebbe essere riattivata dalla presentazione di alcuni particolari del contesto.
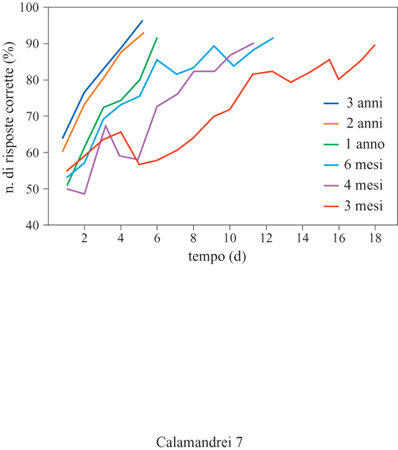
Nella specie umana è stata prestata una particolare attenzione al fenomeno dell'amnesia infantile, intesa come l'impossibilità di ricordare nella vita adulta eventi precedenti ai primi 2 ÷ 3 anni di vita. Analogamente a quanto è avvenuto nel caso delle inferiori capacità di memoria mostrate durante lo sviluppo in modelli animali, nel dibattito sull' origine dell' amnesia infantile si confrontano due diverse posizioni. L'approccio più organicista sostiene che le strutture neurologiche (essenzialmente quelle del sistema limbico) preposte all'immagazzinamento della memoria a lungo termine non sono pienamente funzionali durante i primi due anni di vita. Il ruolo della immaturità di questi sistemi neurali è stato ampiamente saggiato in primati non umani come il macaco (Macaca mulatta). Esiste una buona corrispondenza tra lo sviluppo cognitivo di macachi e quello dei bambini in test cognitivi classici (Bachevalier, 1990), che presuppongono capacità di discriminazione sia visiva che spaziale (fig. 7). Mentre la capacità di discriminare visivamente tra oggetti è già presente sia nel macaco che nel bambino nei primi mesi di vita, quella di discriminare tra più oggetti sulla base della minore o maggiore familiarità matura molto più tardivamente. Nei primati non umani lo sviluppo delle capacità cognitive è parallelo alla maturazione sinaptica della corteccia prefrontale e dell'ippocampo. Sulla base di questi risultati alcuni autori (Bachevalier, 1990) sostengono l'esistenza, nei primati umani e non umani, di due sistemi di memoria dissociabili ontogeneticamente: un sistema non cognitivo (non cognitive habit system), maturo alla nascita, mediato da strutture cortico-striatali, e un sistema cognitivo vero e proprio o di rappresentazione, che utilizzerebbe un circuito cortico-limbico-diencefalico, a maturazione più tardiva, e che sarebbe responsabile dell'attuazione di paradigmi di condizionamento che richiedono al soggetto l'uso del contesto spazio-temporale. La dicotomia tra questi due sistemi viene criticata dalla stessa Rovee-Collier (1990), che ha dimostrato in neonati umani ottime capacità di memoria a lungo termine anche nel corso del primo anno di vita. Le strutture neurologiche necessarie per la percezione, l'apprendimento e la memoria (per esempio, quelle del sistema visivo) sono già funzionali alla nascita, anche se continuano a maturare considerevolmente nei primi due anni di vita. Le differenze tra le prestazioni di apprendimento e memoria del bambino e quelle dell'adulto sarebbero perciò quantitative piuttosto che qualitative. Nei primi 2 ÷ 3 anni di vita cambierebbero la percezione, l'attenzione e la selezione degli stimoli nel contesto. Inoltre, lo sviluppo del linguaggio sarebbe uno dei maggiori cambiamenti di stadio che hanno luogo in questa fase ontogenetica. Lo sviluppo cognitivo potrebbe essere sinteticamente caratterizzato dall'incremento della capacità di generalizzare, ossia di ampliare la varietà dei contesti nei quali le esperienze apprese precocemente possono essere richiamate ed espresse. Il dibattito sull' amnesia infantile è tuttora aperto; M.L. Howe e M.L. Courage (1993), in un'approfondita rassegna, ipotizzano che la maturazione del concetto di sé segni la comparsa della memoria autobiografica, e quindi della capacità di ricordare eventi anche molto distanti nel tempo. Dati molto recenti indicano che alcune esperienze fatte nel corso del primo anno di vita avrebbero ripercussioni importanti per lo sviluppo di capacità complesse quali l'apprendimento del linguaggio (Saffran et al., 1996). Addirittura, alcune caratteristiche del linguaggio nativo potrebbero essere apprese nella vita intrauterina. Lo studio del comportamento fetale sembrerebbe indicare che durante la vita prenatale, in particolare nel corso del terzo trimestre di gravidanza, il feto è già in grado di ricevere ed elaborare informazioni, principalmente di carattere uditivo e olfattivo.
Conclusioni
Negli ultimi trent'anni del 20° secolo la psicobiologia dello sviluppo ha conquistato un ruolo importante nel settore delle scienze del comportamento e, più in generale, delle neuroscienze. Gli studi ontogenetici hanno infatti rivelato la ricchezza del repertorio comportamentale del neonato nonché le sue insospettate capacità di apprendere e ricordare eventi anche in fasi molto precoci dello sviluppo, evidenziando la grande importanza delle esperienze precoci per l'organizzazione successiva del comportamento e del sistema nervoso. La scoperta di analogie tra i meccanismi plastici alla base di esperienze precoci di tipo diverso in specie differenti di vertebrati ha contribuito al superamento della vecchia distinzione tra innato e appreso, evidenziando come la modificazione del comportamento in base all'esperienza sia sempre la risultante della interazione dialettica tra i limiti specie-specifici e la flessibilità del sistema nervoso. Lo studio dell'apprendimento neonatale rappresenta un'opportunità unica per chiarire come circuiti e vie nervose complessi e tra loro in competizione si organizzino nel corso dell'ontogenesi cerebrale per dar luogo alle funzioni cognitive dell'adulto.
Bibliografia citata
BACHEVALIER, J. (1990) Ontogenetic development of habit andmemory formation in primates. In The development and neural bases of higher cognitive functions, a c. di Diamond A., vo1.608, New York, New York Academy of Sciences, pp. 457-485.
BATESON, P.P.G. (1981) Ontogeny of behaviour. British Med. Bull., 37, 159-164.
BOLLES, R.C. (1970) Species-specific defense reactions and avoidance learning. Psychol. Rev., 77, 32-48.
CALAMANDREI, G., ALLEVA, E. (1996) Neuronal growth factors, neurotrophins and memory deficiency. Behav. Brain Res., 66, 129-132.
CAMPBELL, B.A. (1984) Reflections on the ontogeny of leaming and memory. In Comparative perspectives on the development of memory, a c. di Kail R., Spear N.E., Hillsdale, Lawrence Erlbaum Associates, pp. 23-35.
DECASPER, A.J., FIFER, W.P. (1980) Of human bonding: newboms prefer their mothers' voices. Science, 208, 1174-1176.
D'UDINE, B., ALLEvA, E. (1984) The ontogeny of leaming capability in rodents: comparative models. In Contemporary psychology: biological processes and theoretical issues, a c. di McGaugh l, voI. I, Amsterdam-New York, North Holland, pp. 131-143.
GARCIA, J., KOELLING, R.A. (1966) Relation of cue to consequence in avoidance leaming. Psychon. Sci., 4, 123-124.
HOWE, M.L., COURAGE, M.L. (1993) On resolving the enigma of infantile amnesia. Psychol. Bull., 113, 305-326.
JOHANSON, I.B., HALL, W.G. (1979) Appetitive leaming in I-dayold rat pups. Science, 205, 419-421.
KLOPFER, P. (1988) Metaphors for development: how important are experiences early in life? Dev. Psychobiol., 21, 671-678.
NADEL, L., ZOLA-MORGAN, S. (1984) Infantile anmesia: a neurobiological perspective. In Infant memory, a c. di Moscovitch M., New York, Plenum Press, pp. 145-172.
O'KEEFE, J., NADEL, L. (1978) The hippocampus as a cognitive map. New York-Oxford, Clarendon Press.
PEDERSEN, P.E., STEWART, W.B., GREER, C.A., SHEPHERD, G.M. (1983) Evidence for olfactory function in utero. Science, 221, 478-480.
ROVEE-COLLIER, C. (1990) The 'memory system' of prelinguistic infants. In The development and neural bases of higher cognitive functions, a c. di Diamond A., New York, New York Academy of Sciences, pp. 517-543.
RUDY, J.W., CHEATLE, M.D. (1977) Odor-aversion leaming in neonatal rats. Science, 198, 845-846.
SAFFRAN, J.R., ASLIN, R.N., NEWPORT, E.L. (1996) Statistical leaming by 8-month-old infants. Science, 274, 1926-1928.
SHETTLEWORTH, S.J. (1988) Varieties of leaming and memory in animals. J. Exp. Psychol., 19, 5-14.
SMITH, G.J., SPEAR, N.E. (1978) Effects ofthe home enviroument on withholding behaviors and conditioning in infant and neonatal rats. Science, 202, 327-329.
SPEAR, N.E. (1984) Ecologically determined dispositions control the ontogeny of leaming and memory. In Comparative perspectives on the development of memory, a c. di Kail R., Spear N.E., Hillsdale, Lawrence Erlbaum Associates, pp. 325-358.
STEHOUER, G.J., CAMPBELL, B.A. (1980) Ontogeny of passive avoidance: role of task demands and development of speciestypical behaviours. Dev. Psychobiol., 13, 385-398.
WILSON, D.A., SULLIVAN, R.M. (1994) Neurobiology of associative learning in the neonate: early olfactory leaming. Behav. Neur. Biol., 61, 1-18.
ZOLMAN, J.F. (1983) Ontogeny of leaming. In Perspectives in ethology, a c. di Bateson P.P.G., Klopfer P.H., voI. 5, Plenum Press, pp. 275-323.
Bibliografia generale
HALLIDAY, T.R., SLATER, P.J.B. Genes, development and learning. Oxford, Blackwell Scientific Publications, 1983.
JOHNSON, M.H. Brain development and cognition: a reader. Oxford-Cambridge, Mass., Blackwell, 1993.
KRASNEGOR, N.A., BLASS, E.M., HOFER, M.A., SMOTHERMAN, W.P. Perinatal development: a psychobiological perspective. Orlando, Academic Press, 1987.
PIAGET, J. The construction of reality in the child. New Y ork, Basic Books, 1954.
SPEAR, N.E., CAMPBELL, B.A. Ontogeny of learning and memory. Hillsdale, Lawrence Erlbaum Associates, 1979.