
Neuroimaging dei processi cognitivi
Neuroimaging dei processi cognitivi
Storicamente, lo studio della mente e lo studio del cervello sono stati a lungo separati. La psicologia si avvicinava alla mente come se fosse una scatola nera, un'entità il cui lavorio interno risultava nascosto, ma il cui funzionamento poteva essere esplorato esaminando le trasformazioni tra i dati in entrata e i dati in uscita e misurando i parametri caratteristici di tali trasformazioni, quali i tempi di reazione e l'accuratezza nello svolgimento di un compito. Lo studio del cervello, viceversa, era il regno della biologia. Le tecniche classiche utilizzate per studiare l'anatomia e la fisiologia del tessuto nervoso hanno svelato molti aspetti importanti della struttura e del funzionamento del cervello, grazie all'analisi del sistema nervoso di topi, gatti, scimmie e persino lumache di mare. Le implicazioni di queste scoperte neurofisiologiche per il funzionamento del cervello umano dovevano però essere sempre dedotte, in quanto la natura invasiva delle tecniche a disposizione impediva una loro diretta applicazione all'esame dell'attività cognitiva umana.
Per oltre un secolo, gli studi sul comportamento dei cerebrolesi e quelli anatomici del cervello umano post mortem sono stati pertanto i metodi neuroscientifici predominanti nell'approccio diretto al problema. La situazione è cambiata radicalmente negli ultimi decenni, perché gli straordinari progressi teorici e tecnologici hanno favorito lo sviluppo di alcune metodiche non invasive che sono in grado di investigare il funzionamento cerebrale: le tecniche di neuroimaging. La vera forza di queste tecniche nel delucidare le più profonde attività del cervello durante i processi cognitivi si evidenzia soprattutto quando esse sono combinate con approcci psicologici adeguati.
Si tratteranno qui alcune questioni teoriche fondamentali per apprezzare le potenzialità di tali tecniche nel chiarimento delle basi neuronali dei processi cognitivi, e sarà fornito materiale di riflessione a chi intenda valutare criticamente gli studi che stanno fiorendo in letteratura. I punti presi in esame comprendono una descrizione delle tecniche usate per l'imaging cerebrale, un'analisi a livello sistemico del funzionamento del cervello e uno schema per valutare la funzione cognitiva a un livello appropriato per gli studi di imaging. Dopo una breve visione d'insieme delle più importanti tecniche di neuroimaging attualmente in uso, esamineremo in particolare quelle di visualizzazione mediante risonanza magnetica funzionale, discutendone sia le capacità sia i limiti. Soprattutto, ampio spazio verrà dato ai principî logici e metodologici che consentono di progettare esperimenti di neuroimaging da cui sia possibile estrarre dati attendibili sui rapporti tra funzionamento del cervello e processi cognitivi. Attraverso i progressi della psicologia cognitiva è possibile una corretta scomposizione e analisi degli stadi di elaborazione delle informazioni e di costruzione delle rappresentazioni mentali: senza tali conoscenze, infatti, diventa difficile interpretare i pattern di attivazione metabolica rilevati con le moderne tecniche di neuroimaging; ma è anche vero che in molti casi sono state queste ultime a dare contributi decisivi per dirimere questioni difficili che i metodi psicologici di per sé non potevano risolvere.
Le tecniche di neuroimaging
La maggior parte delle tecniche neurofisiologiche classiche fornisce informazioni sul funzionamento del sistema nervoso a livello cellulare o subcellulare. Quasi tutte producono un qualche tipo di rappresentazione visiva del funzionamento neuronale, che si tratti di istogrammi delle scariche cellulari, di videomicroscopie al rallentatore di cellule in migrazione, oppure del tracciato oscilloscopico di un singolo potenziale d'azione. Tutte potrebbero essere considerate tecniche di neuroimaging. Senza entrare nel merito della definizione del termine, tuttavia, in questa sede considereremo tecniche di neuroimaging quelle che forniscono informazioni sul funzionamento del sistema nervoso su larga scala e a livello sistemico.
La discussione sarà in gran parte limitata all'uso di queste tecniche negli studi di attivazione, ossia negli studi messi a punto per attribuire ruoli funzionali a specifiche regioni del cervello, nelle quali sono stati identificati cambiamenti elettrici o corrispondenti cambiamenti emodinamici; a proposito di tali cambiamenti è fondamentale sapere quanto accuratamente la tecnica di neuroimaging rifletta il segnale biologico sottostante. Delineare i correlati neuronali dei processi cognitivi significa migliorare la nostra conoscenza sia delle componenti costitutive di tali processi sia dell'organizzazione cerebrale. In alcuni casi, gli studi di attivazione delineano 'l'anatomia funzionale' di un compito cognitivo; in altri, tentano di determinare in modo più dettagliato il tipo di computazione che una regione compie.
Nei paradigmi di attivazione, sono tipicamente misurate le risposte a due o più compiti che vengono assegnati durante una serie di scansioni condotte in una singola seduta di sperimentazione. Si ritiene che le differenze a livello regionale nel segnale misurato durante i vari compiti riflettano le differenze nell'attività neuronale locale associata allo svolgimento di quei compiti. Tali differenze regionali possono essere determinate in vari modi. Nella maggior parte degli esperimenti di neuroimaging si usa un metodo chiamato 'sottrazione' per individuare le regioni di variazione emodinamica tra due diversi compiti. I paradigmi di sottrazione possono impiegare il confronto diretto tra il compito bersaglio e il compito di comparazione, rivelando le regioni differentemente attivate dai due, o alternativamente ciascun compito di interesse può essere confrontato con gli altri rispetto a un comune compito di base, che rivela sia le analogie sia le differenze tra i vari compiti. Le immagini mediate e normalizzate di una condizione di base o di comparazione vengono sottratte dalle immagini mediate e normalizzate di un compito bersaglio: le regioni a intensità positiva nell'immagine risultante riflettono le regioni cerebrali maggiormente attive nel compito bersaglio; le regioni a intensità negativa riflettono quelle maggiormente attive nel compito di comparazione. Le differenze nell'attivazione locale tra le diverse condizioni di scansione si possono attribuire alle differenze tra i compiti svolti. Le implicazioni della sottrazione di immagine non possono andare oltre, in quanto essa è solo un mezzo in grado di rilevare le differenze di attività tra due condizioni diverse.
Elettroencefalogramma e magnetoencefalogramma
L'elettroencefalogramma (EEG) utilizza un apparato di elettrodi localizzati sul cuoio capelluto per rivelare le variazioni dei campi elettrici che sono generate dall'attività neuronale. Analogamente, il magnetoencefalogramma rivela le perturbazioni del campo magnetico sulla superficie del capo che vengono causate dagli effetti induttivi delle variazioni dei campi elettrici nel cervello. Entrambe le tecniche possono essere utilizzate insieme a manipolazioni cognitive allo scopo di studiare determinati aspetti del processo cognitivo. I metodi di EEG, quando sono usati in questo modo, sono spesso chiamati 'potenziali evocati' (EP, Evoked potentials) o 'potenziali correlati a eventi' (ERP, Event related potentials). I segnali elettrici o magnetici rilevati forniscono un'immagine in tempo reale dell'attività cerebrale in termini di tracciati dinamici dei segnali in vari punti del cuoio capelluto. Tuttavia, con questo tipo di dati la localizzazione della fonte risulta problematica: il tentativo di determinare la collocazione del tessuto neuronale attivo può dar luogo a un numero pressoché infinito di ipotesi. Allo scopo di localizzare le fonti bisogna aggiungere ulteriori limitazioni. Così, mentre la risoluzione temporale di queste tecniche è di gran lunga superiore a quella di altri metodi di visualizzazione del funzionamento del cervello, in molti casi esse sono inadeguate per la localizzazione spaziale di segnali multipli.
Tecniche di imaging ottico
Alcune tecniche di imaging ottico utilizzano agenti di contrasto esogeni, come coloranti la cui fluorescenza dipende dalla differenza di potenziale, dalla concentrazione di calcio o da altre variabili regolate fisiologicamente; esistono anche tecniche che si basano su fattori endogeni, come la diffusione della luce in risposta al volume cellulare. La maggior parte di questi metodi è invasiva o comunque inadatta agli studi sull'uomo, eccetto che in casi limitati, come per esempio in neurochirurgia. Ciononostante, la cosiddetta 'spettroscopia nel vicino infrarosso' (NIRS, Near infrared spectroscopy), che misura attraverso il cranio lo spettro di assorbimento della luce da parte dell'emoglobina ossigenata, dell'emoglobina deossigenata e della citocromo c ossidasi, si sta dimostrando una promettente tecnica di imaging funzionale non invasiva. Al momento, i suoi principali limiti sono costituiti dalla luce, che non riesce a penetrare a fondo nei tessuti, e dal metodo, che misura le risposte emodinamiche relative al flusso sanguigno in siti discreti, anzichè in tutto il cervello.
Tomografia a emissione di positroni
La tomografia a emissione di positroni (PET) è stata la prima tecnica di neuroimaging dell'intero cervello a offrire una buona localizzazione della fonte dell'attività neuronale. Essa lavora localizzando gli eventi di decadimento di traccianti marcati radioattivamente, che vengono somministrati al soggetto per endovena o inalazione. I rivelatori PET, sistemati in anelli attorno allo scanner, individuano i fotoni emessi dalle annichilazioni elettrone-positrone che sono causate dal decadimento radiattivo, e li localizzano secondo un principio chiamato 'rilevamento per coincidenza'. Gli eventi di decadimento individuati e registrati durante una scansione dell'emissione sono ricostruiti in un'immagine della distribuzione spaziale dei segnali mediante un algoritmo matematico basato su una retroproiezione filtrata, un algoritmo tomografico che è stato sviluppato per la prima volta per la TC (Tomografia computerizzata).
La versatilità della PET è dovuta al fatto che essa permette di usare traccianti specifici per 'etichettare' molte variabili fisiologiche differenti. Tra i marcatori (marker) fisiologici collegati al funzionamento del cervello che la PET può misurare vi sono il metabolismo del glucosio e dell'ossigeno, il volume sanguigno, la distribuzione di diversi recettori neurochimici e il flusso sanguigno cerebrale. Queste misurazioni sono importanti sia per comprendere le basi fisiologiche di vari metodi di neuroimaging sia per scopi clinici. Le prove di legame dei recettori si sono rivelate di valore incalcolabile, in quanto hanno permesso di svelare alcuni 'segreti' farmacologici del cervello, comprese la distribuzione e l'affinità dei recettori e dei siti di legame di un farmaco, e di mostrare l'eventuale perdita funzionale di una certa classe di recettori associata a sindromi cliniche specifiche.
Nei paradigmi di attivazione con la PET, l'attività neuronale regionale è dedotta dalle misurazioni del flusso sanguigno cerebrale locale. Come il flusso sanguigno aumenta per fornire energia ai muscoli quando vengono utilizzati, allo stesso modo esso aumenta per portare i substrati metabolici (glucosio e ossigeno) alle regioni localizzate del cervello che sono coinvolte nell'attività di elaborazione. Usando traccianti che incorporano l'isotopo dell'ossigeno 15O, dotato di un'emivita di 122 sec, per misurare le variazioni del flusso sanguigno durante lo svolgimento di determinati compiti, sono stati fatti considerevoli passi avanti nella comprensione dei processi cognitivi, quali per esempio la visione, l'attenzione, il linguaggio, l'apprendimento motorio e la memoria.
Risonanza magnetica funzionale
Fino al recente sviluppo della risonanza magnetica funzionale (fMRI, Functional magnetic resonance imaging), la PET non aveva rivali per la capacità di fornire informazioni circa l'organizzazione funzionale del cervello umano su larga scala. Tuttavia, per una serie di motivi tecnici, la fMRI è in grado di offrire una risoluzione spaziale e temporale decisamente superiore. Lo sviluppo dell'MRI ha reso possibile visualizzare, in modo tridimensionale e non invasivo, la struttura interna di oggetti composti da materiali chimicamente diversi, con una risoluzione molto più alta rispetto a quella che si ottiene attraverso le tecniche basate sui raggi X come la TC. Inoltre, poiché non richiede la somministrazione di sostanze radioattive, la MRI non è invasiva quanto la PET e ogni soggetto può essere esaminato più e più volte. Fino a poco tempo fa, la risonanza magnetica era usata soprattutto in sede clinica per visualizzare la struttura di patologie cerebrali come tumori ed edemi, e solo negli ultimi anni è diventato evidente che il segnale MR può riflettere anche le variazioni emodinamiche provocate dall'attività neuronale.
Il segnale della risonanza magnetica deriva dalle caratteristiche intrinseche dei nuclei dotati di momenti di dipolo magnetico in un campo magnetico esterno. La precessione od oscillazione dei dipoli magnetici in un campo magnetico costituisce il segnale base MR. Nel cervello esso proviene principalmente dai nuclei di idrogeno delle molecole d'acqua, e il suo tempo di decadimento dipende dalla composizione del tessuto: il segnale decade più lentamente nella materia grigia piuttosto che nella materia bianca, e ancora più lentamente in molte lesioni, come i tumori. Il contrasto tra un tessuto e un altro osservabile in un'immagine MR è dovuto principalmente a questa differenza.
La sequenza dettagliata degli impulsi, che influenza in maniera cruciale il segnale ed è essenziale per qualsiasi resoconto completo dell'MR, è notevolmente complessa. Il gran numero di parametri coinvolti in ogni esperimento di imaging MR e l'ampio spettro di valori che si possono scegliere per ciascuno di essi rendono la tecnica molto stimolante e versatile.
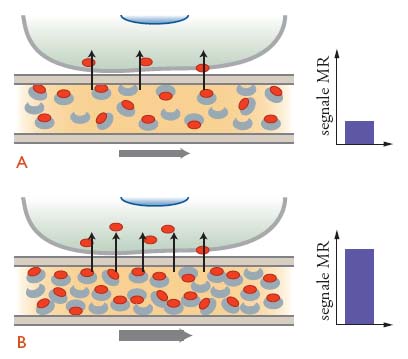
Sin dalle prime dimostrazioni della potenzialità dell'MR come tecnica per misurare l'attività cerebrale, sono stati sviluppati parecchi tipi di sequenze di impulsi sensibili ai parametri correlati al funzionamento del cervello, come la perfusione, il volume sanguigno e l'ossigenazione del sangue. La tecnica BOLD (Blood oxygenation level dependent), che è la più usata, trae vantaggio dal fatto che le proporzioni relative di emoglobina ossigenata e deossigenata nel sangue cambiano in seguito all'aumento dell'attività neuronale (fig. 3). Durante l'attivazione, gli incrementi del consumo di ossigeno sono infatti inferiori a quelli del metabolismo del glucosio e del flusso sanguigno. Di conseguenza, l'incremento di ossiemoglobina dovuto all'aumentato flusso sanguigno supera di gran lunga la conversione dell'emoglobina ossigenata in deossigenata dovuta al consumo di O2. L'ossiemoglobina e la deossiemoglobina possiedono differenti proprietà magnetiche: il netto decremento nella concentrazione relativa di deossiemoglobina risulta in un aumento del segnale BOLD.
In teoria, non c'è un limite alle capacità della fMRI di risolvere l'informazione spaziale; tuttavia, come per tutte le metodiche di imaging, va tenuto conto del rapporto segnale/rumore (SNR, Signal-to-noise ratio). Esso, per la fMRI, è proporzionale al segnale intrinseco (inclusa la forza del campo), al volume del voxel e alla radice quadrata del tempo di imaging. Riducendo la dimensione lineare del voxel alla metà (per es., il voxel di un fattore 8) occorre aumentare il tempo di imaging 64 volte per mantenere lo stesso SNR. Poiché la modulazione dell'intensità del segnale BOLD durante l'attivazione funzionale è relativamente modesta, superando in genere di appena il 2-5% i valori a riposo, l''arte' dell'MR consiste nel massimizzare il segnale rispetto al rumore. Le dimensioni del voxel scelte negli esperimenti fMRI riflettono dunque la necessità di questo bilanciamento, e non la risoluzione spaziale della tecnica in sé. Per migliorare l'SNR i dati sono frequentemente mediati su numerose sequenze relative a uno stesso soggetto. Occasionalmente essi sono mediati su diversi soggetti, come nella PET, per aumentare ulteriormente l'SNR ma, a causa della variabilità anatomica e funzionale tra gli individui, ciò va a scapito della risoluzione spaziale dell'immagine finale.
Quanto sia accurata la localizzazione dell'attivazione cerebrale da parte della fMRI resta un problema aperto e, in una certa misura, la risposta dipende dalla sequenza di impulsi usata: si ritiene che alcune sequenze evidenzino le vene di deflusso, le quali possono trovarsi ad alcuni millimetri dal sito dell'attività neuronale che provoca i cambiamenti di ossigenazione, mentre altre mirano in modo più specifico ai letti capillari del tessuto attivo. Inoltre, la scansione con un voxel di piccole dimensioni dirige la tecnica verso l'identificazione di piccole regioni con grande variazione di ossigenazione, che di nuovo possono corrispondere alle localizzazioni delle vene di deflusso, e non del neuropilo. Queste misurazioni potrebbero offrire una rappresentazione fuorviante della localizzazione e dell'intensità dell'attività neuronale sottostante. Nella maggior parte degli studi fMRI i voxel misurano almeno alcuni millimetri di lato. Sebbene i dettagli delle caratteristiche temporali della reazione emodinamica non siano ben compresi, è noto che un cambiamento rilevabile del flusso sanguigno ha luogo molti secondi dopo l'attività neuronale che lo produce, e che il segnale ha bisogno di pochi secondi per aumentare fino al massimo e decadere. Così, sebbene la fMRI possa misurare i cambiamenti del flusso sanguigno quasi in tempo reale, ciò che essa misura corrisponde all'attività neuronale avvenuta diversi secondi prima. Gli studi attuali suggeriscono tuttavia che i dati relativi all'inizio e allo sviluppo della reazione emodinamica, rilevata in una data area rispetto a un dato stimolo, sono del tutto attendibili, il che permette di fare alcune deduzioni sulle proprietà temporali delle risposte misurate.
Con l'invenzione della fMRI ecoplanare (EPI, Ecoplanar imaging) e il contemporaneo sviluppo di una strumentazione che consente una variazione rapidissima dei gradienti del campo magnetico, una singola immagine MR può essere acquisita in meno di 100 msec, e i dati relativi all'intero cervello in 1-2 secondi. Tuttavia, la qualità del rapporto tra il segnale e il rumore di una singola immagine è troppo scadente perché questa sia di per sé stessa utile. Le scansioni EPI sono tipicamente mediate su esperimenti simili, per produrre dati fMRI con un SNR adeguato.
Un'altra tecnica molto efficace che è stata di recente sviluppata è l'adattamento alla fMR. Tale metodo consente di marcare efficacemente un sottoinsieme di neuroni in un'area eterogenea e di studiare le loro proprietà funzionali. Ciò si realizza adattando una popolazione di neuroni ed esponendola prima a un singolo stimolo ripetitivo, e successivamente a uno stimolo che differisce dall'originale per un singolo parametro. I dati relativi al ritorno allo stato iniziale indicano quanto essa sia sensibile a tale parametro.
Il ruolo della psicologia cognitiva negli studi di imaging
Una prospettiva sistemica rivela che, durante l'esecuzione di compiti cognitivi, l'attività neuronale del cervello è localizzata fortemente a livello delle regioni corticali, per quanto esse possano essere ampiamente distribuite. Nel determinare quali regioni siano attive, e durante quali compiti, si comincia a comprendere come il cervello suddivida le operazioni cognitive complesse in componenti più semplici. Il fatto che gli esperimenti di neuroimaging abbiano fornito informazioni attendibili e riproducibili sui centri dell'attività neuronale durante compiti che vanno dalla vista al linguaggio, alla memoria e all'abilità di apprendimento, dimostra che il cervello non è un organo omogeneo, nel quale tutti i processi cognitivi impiegano gli stessi gruppi di neuroni, ma è piuttosto una struttura altamente organizzata in unità funzionali specifiche. Perciò, per progettare o interpretare gli studi di neuroimaging funzionale, è necessario conoscere non solo i limiti degli strumenti di misurazione e la biologia del cervello, ma anche i meccanismi e i processi mentali di elaborazione delle informazioni.
Negli studi di attivazione vengono utilizzati tre tipi di compiti psicologici: i compiti bersaglio, comprendenti un compito o un sottocompito di interesse; i compiti di comparazione, simili ai precedenti ma che manipolano o mantengono costante una componente di interesse; i compiti di base, di livello più basso, usati per rappresentare uno stato inattivo. Le differenze nell'attivazione regionale tra scansioni di compiti attivi, o fra scansioni di compiti attivi e di base, riflettono differenze nelle richieste associate ai compiti. La selezione dei compiti è estremamente importante, poiché se non si è sicuri di che cosa il soggetto sperimentale stia effettivamente pensando diventa impossibile interpretare il pattern di attivazione fisiologica rilevato da una qualsiasi delle tecniche di imaging. È fondamentale tenere presente che i compiti cognitivi complessi sono quasi sempre costituiti da numerose sotto-operazioni: per esempio, nella maggior parte degli esperimenti di attivazione, un soggetto riceve istruzioni, percepisce stimoli, svolge determinate operazioni cognitive e risponde apertamente in un modo prescritto. Si capisce, quindi, quanto sia rilevante l'analisi funzionale del compito in tutti i suoi stadi di elaborazione. Sebbene questo fatto non riceva generalmente una sufficiente attenzione, una scomposizione funzionale attenta e appropriata è alla base del successo degli esperimenti di neuroimaging.
Il metodo più diretto per affrontare un esperimento consiste nel caratterizzare un paradigma del compito bersaglio tenendo conto delle informazioni introdotte, delle necessità di elaborazione e dei risultati. L'esperimento ideale mantiene fisse due di queste tre componenti nei vari compiti di comparazione, e manipola il più basso numero possibile di variabili per volta. Se è disponibile una documentazione, di carattere psicologico o neurologico, riguardante la natura del compito, essa può essere usata per indirizzare la scomposizione funzionale. Quando si progettano i compiti psicologici per un esperimento di neuroimaging, occorre tenere presente che variazioni apparentemente piccole nei loro tratti superficiali possono avere effetti sorprendenti sui corrispondenti schemi di attivazione. Parametri come la frequenza di presentazione visiva, la frequenza delle risposte motorie, la frequenza o familiarità dei vocaboli e così via, possono influenzare notevolmente l'emodinamica e confondere l'interpretazione dei risultati sperimentali. Di conseguenza, è importante cercare di adattare per quanto possibile questi parametri alle condizioni sperimentali, a meno di verificarne esplicitamente gli effetti. Per esempio, nell'esaminare le differenze di elaborazione fra vocaboli ad alta e bassa frequenza, le parole dovrebbero essere presentate nella stessa misura, essere della stessa lunghezza e dello stesso numero di sillabe, e la soluzione migliore le vorrebbe il più possibile corrispondenti anche nella composizione fonetica. Infine, va sottolineato che sebbene una data scomposizione funzionale possa sembrare giustificata e completa, il suo ruolo è quello di avanzare un'ipotesi di lavoro che rappresenta una soltanto delle numerose alternative possibili. La validità di una particolare scomposizione funzionale può sempre essere messa in discussione, specialmente se le prove empiriche ottenute dall'esperimento o da altre fonti non si conformano al modello ipotizzato.
La scelta dei compiti di base e di comparazione
Molti studi di attivazione sono volti a chiarire quali regioni del cervello siano coinvolte in un determinato compito. Per fornire una risposta, l'ideale sarebbe confrontare i livelli di attività durante lo svolgimento del compito con i livelli di attività di quando il cervello 'non sta facendo nulla' o è a riposo. Ciò è impossibile, in quanto il cervello non è mai veramente a riposo, dovendo presiedere alle funzioni omeostatiche, regolare il livello di attenzione o di allerta, e così via. È però possibile individuare condizioni che prevedono un'elaborazione cognitiva minima: i cosiddetti 'compiti di base'. Questi ultimi sono solitamente scelti per la loro semplicità e per il fatto che non condividono importanti caratteristiche o esigenze con il compito di interesse. Si deve sempre tener presente, tuttavia, che il compito scelto come presumibilmente semplice potrebbe in realtà non essere tale, o potrebbe avere in comune con il compito di interesse alcune componenti, senza che lo sperimentatore ne sia consapevole. Molti ricercatori utilizzano compiti di base standard in tutti i loro studi, indipendentemente dall'obiettivo dell'indagine. Ciò rende più facile trovare schemi comuni di attivazione in molte ricerche diverse e apre la possibilità a future meta-analisi. A prescindere dalla scelta del compito di base, è di primaria importanza essere consapevoli delle operazioni cognitive che esso richiede, così come delle altre in cui il soggetto potrebbe impegnarsi.
I compiti di comparazione sono più complessi, spesso di alto livello, e sono scelti perché differiscono in qualche modo specifico dal compito primario di interesse (nel qual caso sono a volte chiamati 'compiti di contrasto'), o perché condividono con esso importanti caratteristiche, oppure per entrambi i motivi. La scomposizione funzionale del compito primario di interesse può identificare caratteristiche che dovrebbero essere condivise oppure assenti nel potenziale compito di comparazione, allo scopo di verificare un'ipotesi specifica sul ruolo funzionale di una o più aree oppure su una particolare scomposizione. Si possono in tal modo sviluppare compiti che soddisfino questi criteri. La scelta dei compiti di comparazione o di contrasto è una discriminante fondamentale per l'effettiva pregnanza e validità di uno studio. Una loro attenta selezione può infatti essere decisiva per svelare il ruolo di una o più regioni del cervello, mentre una loro scelta inappropriata può produrre uno studio che fallisce nel centrare qualsiasi specifico problema cognitivo.
La misura del comportamento
Un requisito minimo per uno studio ben progettato è che i soggetti svolgano effettivamente il compito mentale che lo sperimentatore crede stiano svolgendo. Per tale motivo, nel corso delle sedute esplorative vengono acquisite determinate misure comportamentali. Per esempio, dati relativi ai tempi di reazione, all'accuratezza, alla frequenza di azione e così via, possono essere usati per valutare lo svolgimento del compito e le strategie usate, che hanno un impatto significativo sull'intepretazione dei risultati di neuroimaging. È ovvio che una caratterizzazione fortemente comportamentale del compito e dei metodi di valutazione del compito stesso è fondamentale per impostare correttamente ed eseguire con successo un esperimento di questo tipo.
In certe condizioni, specialmente quelle della fMRI, in cui il movimento facciale che accompagna le risposte verbali può creare serie interferenze, non è possibile misurare il comportamento palese durante la scansione. Si possono comunque usare alcune strategie per valutare se il compito venga svolto come previsto. Per esempio, il ricordo degli oggetti può essere condizionato manipolando il modo in cui essi vengono codificati: un compito a codificazione profonda, come un giudizio semantico, produce un ricordo migliore rispetto a un compito a codificazione debole, come un giudizio ortografico. Si possono presentare ai soggetti alcuni oggetti e chiedere loro di svolgere segretamente compiti che comportano una manipolazione in profondità dell'elaborazione. Se essi svolgano o meno questi compiti secondo le istruzioni ricevute lo si può accertare indirettamente al termine della seduta, attraverso test sulla memoria.
I rischi dell'attribuzione funzionale
Poiché il neuroimaging cerca di attribuire ruoli funzionali a regioni cerebrali specifiche o a gruppi di regioni, non è sorprendente che le pubblicazioni sull'argomento spesso affermino, in modo più o meno esplicito, che un certo studio dimostra che l'area X attiva la funzione Y. È raro che ciò sia vero. È virtualmente impossibile interpretare la rilevanza funzionale di una qualsiasi regione attivata sulla base di una o di alcune scansioni. Quanto si può dire con certezza è che il compito complesso A, realizzato con i parametri usati nell'esperimento specifico, induce un'attività nella regione X, e che tale compito può essere provvisoriamente scomposto in senso funzionale nei sottocompiti W, Y, Z, ecc. Al massimo, un esperimento di neuroimaging può fornire forti indizi che quella regione sia coinvolta in quel particolare processo; ma nessuno studio singolo può dimostrare la funzione di una particolare regione cerebrale. Perché si possa con forza stabilire un'attribuzione funzionale è necessario un processo estensivo e interattivo definito 'triangolazione funzionale'. Non diversamente dalla triangolazione nelle rilevazioni, la definizione del panorama anatomico-funzionale richiede molteplici misurazioni nel corso di una varietà di compiti e un continuo affinamento dell'interpretazione del ruolo che una particolare area svolge nella cognizione. L'osservazione dell'attivazione di una specifica regione cerebrale in un'ampia gamma di compiti caratterizzati da una stessa sottocomponente funzionale costituisce una buona prova per l'attribuzione; l'assenza di quell'attivazione in compiti simili, ma privi di quella componente funzionale, rafforza ancor più l'inferenza (sebbene, visto che l'analisi deve servire a ridurre al minimo i falsi positivi, sia necessario essere cauti con le affermazioni circa la mancanza di attivazione). La sfida della triangolazione funzionale risiede nell'individuazione di compiti appropriati da un punto di vista psicologico a verificare le ipotesi funzionali. Poiché ogni funzione cognitiva è multidimensionale, ovvero comporta l'attivazione seriale e parallela di numerosi sottoprocessi di elaborazione, possono essere necessarie molte misurazioni per accumulare prove sufficienti a triangolare una funzione in una qualche area, con la clausola che qualsiasi attribuzione funzionale è incrementale e aperta a revisioni di fronte a dati che eventualmente risultassero incoerenti.
Anche utilizzando la triangolazione funzionale, resta inoltre la possibilità che una singola regione sia responsabile di alcuni o persino di tanti tipi diversi di operazioni cognitive. Questa conclusione non riflette necessariamente un'incoerenza biologica o interpretativa. In certe aree cerebrali, come la corteccia visiva, differenti classi di cellule situate in una piccola regione sono responsabili di diverse operazioni; ciò accade probabilmente in tutto il cervello. Poiché il flusso sanguigno è misurato (e forse regolato) a livello regionale (nell'ordine dei millimetri) e non a livello cellulare (nell'ordine dei micron), l'attivazione di differenti popolazioni neuronali nella stessa area può portare a schemi di flusso sanguigno identici, almeno secondo la risoluzione e la sensibilità delle attuali tecniche di neuroimaging. Di conseguenza, l'attivazione della stessa regione in compiti diversi non implica necessariamente l'operatività di identiche componenti funzionali.
I contributi del neuroimaging alla psicologia cognitiva
Nella psicologia cognitiva si verificano spesso situazioni in cui due o più teorie contrastanti spiegano ugualmente bene un fenomeno. In alcuni casi, gli esperimenti di neuroimaging possono essere usati per fornire prove che aiutino a scegliere una teoria piuttosto che un'altra. Per esempio, essi hanno fornito importanti evidenze che permettono di scegliere tra due modelli psicologici, prima indifferenziabili, riguardanti i ruoli che svolge l'attenzione spaziale nei compiti di ricerca visiva. Gli studi psicologici concernenti i paradigmi della ricerca visiva riportano curve con un aumento del tempo medio di scoperta dell'obiettivo in presenza di un aumento di fattori distraenti in concomitanza con i compiti di ricerca visiva. È stato ipotizzato che questi dati risultino dal decrescere dell'efficienza della ricerca parallela con più fattori distraenti, oppure dal tempo aggiuntivo richiesto per realizzare cambi seriali di attenzione rispetto agli oggetti a disposizione. I soli studi comportamentali non erano in grado di distinguere tra i due modelli. Con una serie di esperimenti PET, Maurizio Corbetta e i suoi collaboratori hanno individuato le aree della corteccia parietale superiore che sono coinvolte nei cambiamenti di attenzione spaziale. In uno studio successivo, tali autori hanno scoperto che queste stesse regioni corticali erano fortemente attivate nel corso di un compito di ricerca sulla congiunzione, e ciò ha fornito una prova evidente che per questi compiti di congiunzione il cervello impiega una strategia di ricerca seriale e non una ricerca parallela. L'imaging ha fornito un valido aiuto anche ai seguenti problemi: se l'attenzione sia basata sull'oggetto o sulla caratteristica; se i giudizi riguardanti il contesto in cui qualcosa è appreso (memoria della fonte), piuttosto che i giudizi di familiarità (memoria degli oggetti), coinvolgano differenti operazioni di recupero; se l'immagine mentale implichi o meno la riattivazione di aree corticali primitive.
I miglioramenti nella precisione e nella velocità con cui si svolge un compito dopo un certo periodo di pratica sono stati attribuiti al perfezionamento, con la pratica stessa, dell'efficienza operativa del sistema neuronale, oppure a un cambiamento nei circuiti neuronali che svolgono compiti ben noti o praticati. È probabile che un'analisi attenta della letteratura sulle lesioni avrebbe potuto consentire agli scienziati di discriminare tra queste due possibilità, tuttavia il tipo di cambiamento sotteso agli effetti della pratica è divenuto chiaro soltanto quando tali effetti sono stati esaminati con l'uso di metodi di neuroimaging. Numerosi studi sull'apprendimento cognitivo e su quello motorio hanno permesso di rilevare che differenti circuiti cerebrali si attivano in relazione all'automaticità di un compito e che questi percorsi neuronali possono cambiare nel giro di pochi minuti. Marcus E. Raichle e i suoi collaboratori hanno dimostrato che un compito di risposta verbale non sperimentato attiva sicuramente le regioni del cingolo anteriore prefrontale sinistro e delle cortecce temporali posteriori sinistre. Dopo un certo allenamento, la situazione risulta modificata: le regioni menzionate sopra diminuiscono la loro attivazione e la corteccia silviana insulare mostra un'attivazione più alta; lo schema dell'attività non è distinguibile da quello osservato in un semplice compito di ripetizione di parole. Di fronte a un nuovo stimolo, gli schemi di attivazione si riconvertono in quelli tipici di un compito non sperimentato.
Di recente sono stati prodotti risultati simili, che mostrano un cambiamento nell'architettura neuronale sottesa allo svolgimento di compiti semplici e sperimentati, a proposito dell'apprendimento motorio e visuomotorio. L'ulteriore esplorazione dei tempi di cambiamento degli schemi e delle correlazioni con le variabili comportamentali getteranno nuova luce sulla plasticità neuronale e sulla variabilità dei percorsi di elaborazione che accompagnano le operazioni cognitive più complesse. L'imaging ha inoltre permesso di compiere grandi progressi nella comprensione della memoria umana. Uno dei risultati più recenti e più sorprendenti ottenuti usando gli schemi connessi a eventi è l'aver scoperto che l'attivazione di diverse regioni della corteccia prefrontale e paraippocampale è correlata con la qualità del ricordo verbale e visivo: il tracciato dell'attività neuronale durante la codifica può predire se in seguito uno stimolo sarà ricordato oppure dimenticato.
La fMRI può anche essere usata per indagare i correlati neuronali della percezione conscia. I compiti di antagonismo binoculare presentano a ciascun occhio immagini tra loro incompatibili: sebbene la stimolazione retinica rimanga costante, i soggetti percepiscono alternativamente soltanto un'immagine per volta. Frank Tong e i suoi collaboratori hanno evidenziato che le risposte di ciascuna delle due aree visive dimostratesi in precedenza selettive nei confronti dei volti e delle case variavano periodicamente in opposizione, a seconda che il soggetto riferisse di vedere lo stimolo del volto o della casa, dimostrando così che un'aumentata attività neuronale in queste regioni selettive nei confronti dello stimolo era correlata con la percezione conscia. Infine, oltre a contribuire a chiarire problemi che la psicologia e gli studi sulle lesioni non erano stati in grado di risolvere, il neuroimaging ha fornito risultati del tutto imprevisti, ma sicuramente attendibili, le cui implicazioni per la teoria psicologica e neuronale restano oscure. Per esempio, in contrasto con le aspettative della letteratura neuropsicologica, gli studi di imaging hanno mostrato in modo coerente che le regioni della corteccia prefrontale sono importanti nel recupero della memoria episodica. Essi hanno suggerito che il coinvolgimento della corteccia prefrontale dorsolaterale nel recupero dell'informazione durante compiti di memoria episodica era lateralizzato, anche se studi più recenti indicano che la lateralizzazione della funzione durante il recupero è influenzata da numerosi fattori, tra i quali la natura dell'informazione che deve essere recuperata e il suo livello di dettaglio. Una così chiara dissociazione del recupero della memoria episodica da altri tipi di recupero della memoria non era stata mai anticipata prima degli studi di neuroimaging e, sebbene le implicazioni funzionali e cognitive di questa scoperta debbano essere ancora determinate, senza dubbio esse influenzeranno in modo significativo le conoscenze sull'organizzazione funzionale dei processi mnemonici.
Bibliografia
Buxton 2001: Buxton, Richard B., Introduction to functional magnetic resonance imaging: principles and techniques, Cambridge, Cambridge University Press, 2001.
Corbetta 1995: Corbetta, Maurizio e altri, Superior parietal cortex activation during spatial attention shifts and visual feature conjunction, "Science", 270, 1995, pp. 802-805.
Doyon 1996: Doyon, Julien e altri, Functional anatomy of visuomotor skill learning in human subjects examined with positron emission tomography, "European journal of neuroscience", 8, 1996, pp. 637-648.
Fox, Raichle 1984: Fox, Peter T. - Raichle, Marcus E., Stimulus rate dependence of regional cerebral blood flow in human striate cortex, demonstrated by positron emission tomography, "Journal of neurophysiology", 51, 1984, pp. 1109-1120.
Fox, Raichle 1986: Fox, Peter T. - Raichle, Marcus E., Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects, "Proceedings of the National Academy of Sciences USA", 83, 1986, pp. 1140-1144.
Grill-Spector, Malach 2001: Grill-Spector, Kalanit - Malach, Rafael, fMR-adaptation: a tool for studying the functional properties of human cortical neurons, "Acta psychologica", 107, 2001, pp. 293-321.
Kanwisher, Wojciulik 2000: Kanwisher, Nancy - Wojciulik, Ewa, Visual attention: insights from brain imaging, "Nature reviews. Neuroscience", 1, 2000, pp. 91-100.
Miezin 2000: Miezin, Francis M. e altri, Characterizing the hemodynamic response: effects of presentation rate, sampling procedure, and the possibility of ordering brain activity based on relative timing, "NeuroImage", 11, 2000, pp. 735-759.
Posner, Raichle 1994: Posner, Michael I. - Raichle, Marcus E., Images of mind, New York, Scientific American Library, 1994.
Roland 1993: Roland, Per E., Brain activation, New York, Wiley-Liss, 1993.
Tong 1998: Tong, Frank e altri, Binocular rivalry and visual awareness in human extrastriate cortex, "Neuron", 21, 1998, pp. 753-759.
Tulving 1994: Tulving, Endel e altri, Neuroanatomical correlates of retrieval in episodic memory: auditory sentence recognition, "Proceedings of the National Academy of Sciences USA", 91, 1994, pp. 2012-2015.