
movimento
movimento
Il movimento è un aspetto fondamentale della vita degli organismi. Lo studio del movimento nell’ambito delle neuroscienze riguarda principalmente gli atti motori muscolari. I movimenti muscolari sono classificabili in riflessi, movimenti ritmici e movimenti volontari e sottostanno a precise leggi psicofisiche. Dal punto di vista neurale, il movimento è regolato da un sistema gerarchico a tre livelli: il midollo spinale, il tronco dell’encefalo e la corteccia cerebrale. L’azione di questi centri ha come unica uscita l’attivazione dei motoneuroni e infine la contrazione muscolare. L’esistenza di questa struttura gerarchica permette alle strutture di ordine superiore di utilizzare un repertorio di movimenti coordinati realizzabili dai circuiti della struttura gerarchicamente inferiore in modo indipendente. All’organizzazione del movimento contribuiscono anche i nuclei della base e il cervelletto. La conoscenza dell’organizzazione del sistema motorio permette di comprendere i gravissimi sintomi di neuropatologie legate a specifiche aree del sistema motorio, quali la malattia di Parkinson, la corea di Huntington o le lesioni cerebrali. [➔ cervelletto; corteccia cerebrale; midollo spinale; movimento, disturbi del; nuclei della base] Chiamiamo m. un’azione che porta un corpo a spostarsi da una posizione a un’altra, da un luogo a un altro, o a mutarsi da una configurazione a un’altra. Dai batteri all’uomo, il m. è un aspetto fondamentale dell’esistenza, in quanto permette sia di reagire a stimoli esterni sia di mettere in atto comportamenti volontari. A livello cellulare, il m. è molto importante per varie funzioni dell’individuo, dalla circolazione alla digestione. Negli organismi viventi possiamo definire tre tipi di m.: protoplasmatico, ciliare e muscolare. Il m. protoplasmatico si manifesta sotto forma di correnti nella massa semifluida del citoplasma, riconoscibili per lo spostamento del materiale citoplasmatico. Il m. ciliare è basato sul m. di specifici componenti cellulari dette cilia. Nel sistema nervoso, il m. protoplasmatico è coinvolto nei fenomeni di migrazione neuronale presenti durante lo sviluppo e nella protrusione di filopodi, tipica della crescita assonale e della sinaptogenesi. Il m. delle cilia nelle cellule ciliate dell’orecchio interno è coinvolto nella meccanotrasduzione alla base della percezione acustica e del funzionamento del sistema vestibolare. Il m. muscolare è sicuramente il più studiato nell’ambito delle neuroscienze, in quanto è alla base di tutti i comportamenti attivi dell’individuo. Inoltre, le patologie che colpiscono a vari livelli i meccanismi di controllo ed esecuzione dei m. costituiscono un sottoinsieme molto importante delle neuropatologie.
Tipi di movimento e meccanismi di controllo
Una prima suddivisione categoriale dei m. è tra riflessi, m. ritmici e m. volontari. I riflessi (➔) sono schemi coordinati di contrazioni muscolari attivati in genere dalla stimolazione periferica. Se delle lesioni isolano i centri del midollo spinale dai centri di controllo superiori, i riflessi si presentano come risposte isolate e altamente stereotipate. I riflessi costituiscono il livello più semplice di organizzazione del m., utilizzato dai sistemi motori gerarchicamente superiori per coordinare l’attività dei muscoli che prendono parte all’esecuzione del movimento. I m. ritmici comprendono deglutizione, masticazione e le contrazioni alternate di muscoli flessori ed estensori alla base della deambulazione; come i riflessi, hanno alla base circuiti posti nel midollo spinale o nel tronco dell’encefalo la cui attività è modulata dai centri superiori. I m. volontari sono atti motori diretti a uno scopo specifico, il cui raggiungimento può avvenire anche in presenza di ostacoli o perturbazioni della posizione del corpo, grazie a meccanismi correttivi di tipo anticipatorio o a feedback negativo. I meccanismi anticipatori preparano l’esecuzione del m. in modo che non venga alterato da fattori esterni, percepiti grazie ai sistemi sensoriali e valutati anche grazie all’esperienza precedente. Un esempio tipico è l’aggiustamento posturale che precede il sollevarsi di un braccio, in modo da non perdere l’equilibrio. Il controllo a feedback prevede che i segnali sensoriali vengano confrontati con una copia del segnale di riferimento, che rappresenta la condizione che si vuole mantenere; dal confronto si genera un eventuale segnale di errore capace di attivare meccanismi di compensazione del m. tali da annullare l’errore.
Psicofisica del movimento
Le caratteristiche di esecuzione di un m. sottostanno a tre leggi principali: in primo luogo il risultato finale di un m. viene prefigurato a prescindere da quale gruppo di muscoli o arto viene impiegato per ottenerlo. Per es., è possibile imparare a scrivere con entrambe le mani o addirittura con i piedi, eppure la calligrafia sarà sempre simile. In secondo luogo, il tempo necessario a eseguire una risposta motoria è direttamente proporzionale alla quantità di informazione sensoriale che deve essere analizzata, ossia alla complessità della via neurale che mette in relazione lo stimolo con la risposta motoria. Per es., stimoli propriocettivi che arrivano con poche sinapsi al sistema motorio possono dare risposte in 80÷120 ms, mentre la risposta a stimoli visivi, che richiede una via neurale più lunga, può avvenire dopo intervalli dell’ordine dei 150÷180 ms. In terzo luogo, velocità di esecuzione e accuratezza sono caratteristiche del m. legate tra loro da una proporzionalità inversa.
Azione dei muscoli
Già Charles S. Sherrington agli inizi del 20° sec. aveva mostrato che, per quanto complesso, il comando nervoso relativo a un m. doveva attivare una via finale comune costituita dai motoneuroni spinali e dai muscoli da essi innervati. La contrazione muscolare è l’ultimo passaggio nella catena di eventi che genera il movimento. Il muscolo è costituito da migliaia di fibre polinucleate poste in serie e, nei muscoli più lunghi, in parallelo. Ciascuna fibra riceve l’innervazione di un singolo motoneurone (➔), tuttavia ciascun motoneurone è in grado di innervare circa 100÷1.000 fibre muscolari, sebbene questo numero possa variare tra muscolo e muscolo. L’insieme del motoneurone con le fibre da esso innervate prende il nome di un’unità motrice. L’attivazione del motoneurone si propaga alla fibra muscolare per mezzo del rilascio del neurotrasmettitore acetilcolina che, aprendo i canali associati ai recettori postsinaptici di tipo nicotinico, determina nella fibra muscolare un potenziale postsinaptico eccitatorio (EPSP, Excitatory PostSynaptic Potential). Questo EPSP normalmente è sufficiente a far insorgere un potenziale di azione che si propaga lungo la fibra muscolare. La sinapsi neuromuscolare è molto estesa, inoltre la fibra muscolare può essere assimilata dal punto di vista biofisico a un assone amielinico di grande spessore. Questi due fattori fanno sì che i flussi di ioni coinvolti nella generazione dei segnali elettrici muscolari siano di grande entità. Dato che ciascuna unità motrice si attiva insieme, e che per contrazioni progressivamente maggiori vi è l’attivazione contemporanea di un numero di unità motrici maggiore, i flussi ionici relativi all’attivazione delle singole fibre muscolari si sommano generando forti differenze di potenziale elettrico tra zone attive e zone inattive: il potenziale di azione muscolare determina una variazione del potenziale elettrico tra l’interno di strutture tubulari, che fanno parte del reticolo sarcoplasmatico della fibra muscolare, e il citoplasma della fibra stessa. Questa variazione di potenziale elettrico determina il rilascio nel citoplasma muscolare dello ione calcio, da parte di cisterne (cisterne terminali) poste all’interno della fibra muscolare.
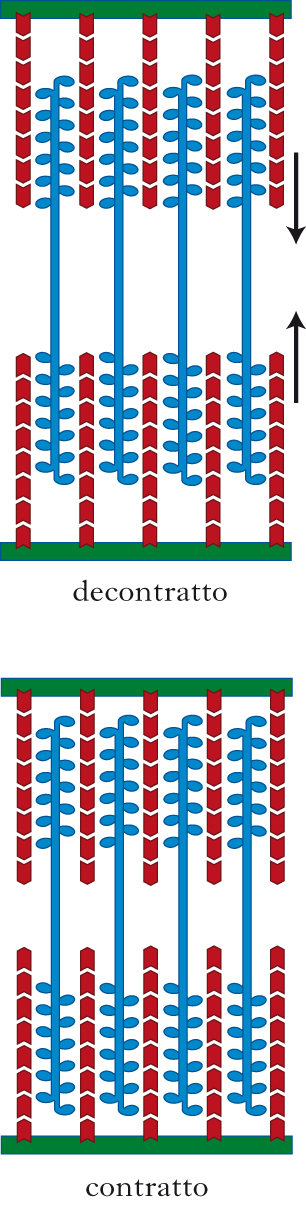
Gli ioni calcio agiscono in modo che i filamenti di actina, posti parallelamente a filamenti di miosina, possano legarsi alla testa globulare di questi ultimi. L’avvenuto legame determina un cambio conformazionale nella testa globulare che si piega, provocando uno scorrimento del filamento di actina rispetto a quello di miosina e sovrapponendo così i due filamenti; in questo modo si accorcia la lunghezza della fibra (contrazione muscolare). Il cambio conformazionale della testa della miosina fa sì che essa risulti in grado di legare ATP, il quale viene idrolizzato liberando l’energia necessaria per il ritorno della testa della miosina nella sua posizione iniziale. Se sono ancora presenti ioni calcio e ATP il ciclo può ripetersi incrementando la contrazione della fibra. All’interno della fibra muscolare sono presenti molti filamenti di actina e miosina allineati lungo l’asse maggiore, posti in parallelo e in serie. Sommando i loro scorrimenti, i filamenti di actina e miosina determinano la contrazione della fibra e del muscolo. L’allungamento della fibra è invece passivo e causa uno scorrimento delle fibre di actina e miosina in senso opposto. La contrazione ripetuta porta alla riduzione dell’ATP, e quindi a un rallentamento o a un blocco del ciclo di contrazione che si riflette in una minor forza e una minor velocità di contrazione del muscolo (affaticamento).
Tipi di contrazione
Il fenomeno alla base della contrazione è uguale in tutte le fibre, tuttavia esistono tre tipi di unità motrici suddivisibili sulla base della velocità di contrazione e della resistenza alla fatica. I muscoli rossi a scossa lenta danno luogo a lente contrazioni e hanno un metabolismo che, garantendo una continua produzione di ATP, permette loro di resistere all’affaticamento. I muscoli bianchi danno invece origine a una rapida scossa e si dividono a loro volta in due sottotipi, a seconda della loro maggiore o minore resistenza alla fatica. I vari tipi di unità motrici vengono reclutati secondo un ordine prestabilito: per una contrazione di modesta entità vengono attivate solo le unità motrici lente resistenti alla fatica; al crescere della forza di contrazione vengono poi reclutate unità motrici veloci resistenti alla fatica; per contrazioni ancora maggiori sono infine reclutate le unità veloci meno resistenti alla fatica. Una maggiore o minore forza di una contrazione muscolare dipende quindi da quanto ciascuna unità motrice viene attivata e da quante unità motrici vengono reclutate.
Controllo neurale del movimento
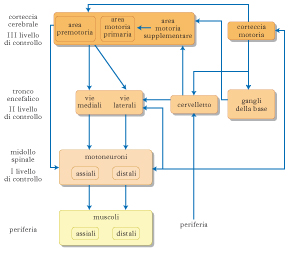
L’attivazione dei motoneuroni costituisce il terminale comune controllato dalle vie neurali che comandano e controllano il movimento. Questa azione viene svolta da circuiti organizzati gerarchicamente e posti a vari livelli del sistema nervoso centrale. In primo luogo, l’analisi delle informazioni sensoriali e dei comandi da convogliare ai motoneuroni viene eseguita all’interno di circuiti del midollo spinale. La loro azione è sufficiente all’esecuzione di riflessi e di movimenti ritmici relativi al corpo, mentre per quanto riguarda il viso e la bocca i circuiti relativi si ritrovano a livello troncoencefalico. I circuiti che mediano i riflessi possono essere anche estremamente semplici. Per es., il riflesso da stiramento, o patellare, viene mediato da una connessione eccitatoria diretta tra i neuroni sensoriali che trasmettono l’informazione relativa alla lunghezza del muscolo e i motoneuroni che innervano il muscolo stesso. In questo modo, ad allungamenti del muscolo dovuti all’azione di forze esterne (come la forza di gravità per i muscoli estensori delle gambe), viene opposta una contrazione muscolare capace di mantenere il muscolo contratto. Questo riflesso ha una funzione importante nel conservare la postura controbattendo l’azione della forza di gravità, e viene comunemente valutato dal neurologo a scopo diagnostico. Il riflesso da stiramento mostra anche come i riflessi possono dare origine ad attivazioni coordinate di gruppi di muscoli. Infatti, l’attivazione sensoriale derivante dall’allungamento muscolare viene anche trasmessa a interneuroni inibitori posti nel midollo spinale che, a loro volta, innervano i muscoli antagonisti a quello allungatosi. In questo modo la ricontrazione del muscolo allungato non troverà opposizione da parte dei muscoli antagonisti che verranno rilasciati. Esistono altri riflessi, oltre a quello da stiramento, la cui funzione è determinata dal tipo di segnale sensoriale che attiva il riflesso (per es., il riflesso di retrazione generato da stimoli nocicettivi), e dal circuito spinale che lo media (il riflesso di retrazione può influenzare in modo inibitorio o eccitatorio l’azione di muscoli posti su più arti). Alcuni riflessi sono osservabili solo durante alcuni stadi dello sviluppo (per es., il riflesso di Moro e il riflesso di Babinski) e sono quindi utilizzati in neuropsichiatria infantile e in pediatria per valutare lo sviluppo neurale del bambino. Il fatto che questi riflessi scompaiano con lo sviluppo non indica necessariamente che i circuiti alla base non siano più presenti; infatti, alcuni di questi riflessi neonatali possono ricomparire nell’adulto a seguito di lesioni (per es., il riflesso di Babinski). Il successivo livello di controllo del m. è il tronco dell’encefalo. Due sistemi, quello mediale e quello laterale, proiettano al midollo spinale: il primo regola principalmente la postura integrando informazioni somatosensoriali, visive e vestibolari, il secondo controlla principalmente la muscolatura distale degli arti ed è particolarmente coinvolto in movimenti del braccio e della mano. Esistono inoltre altri circuiti troncoencefalici che controllano i movimenti del capo e degli occhi.
Ruolo della corteccia cerebrale
Il terzo livello di controllo è costituito dalla corteccia cerebrale. La corteccia motoria primaria e altre aree premotorie proiettano al midollo spinale, direttamente attraverso il tratto corticospinale, e indirettamente mediante proiezioni ai nuclei da cui originano i tratti motori del tronco dell’encefalo. L’attivazione dei neuroni della corteccia motoria primaria precede m. semplici di piccoli gruppi di muscoli e codifica la forza di contrazione e la direzione del movimento. Le aree premotorie sono importanti per la pianificazione e la coordinazione di sequenze complesse di movimenti. A loro volta esse ricevono informazioni da aree associative parietali e prefrontali. Il compito di pianificazione svolto dalla corteccia è semplificato dall’esistenza di un ricco repertorio di m. riflessi codificati già a livello sottocorticale. In questo modo la corteccia può agire limitandosi a facilitare alcuni circuiti e inibendone altri, facendo così in modo che siano i circuiti spinali che integrano gli stimoli sensoriali a definire i dettagli temporali dell’attivazione dei muscoli agonisti e antagonisti. È possibile che sia questa modalità organizzativa a permetterci di eseguire molti m. senza pensare, in modo semiautomatico.
Ruolo del cervelletto e dei nuclei della base
Oltre ai tre livelli gerarchici sopra menzionati, due altre strutture cerebrali hanno un ruolo importante nella pianificazione e nell’esecuzione del m.: i nuclei della base (➔) e il cervelletto (➔). Entrambe queste strutture non inviano tratti nervosi diretti al midollo spinale, tuttavia esse ricevono afferenze dalle aree corticali e vi rinviano segnali. Ciascuna di queste due strutture riceve da zone corticali distinte e rinvia a zone corticali diverse attraverso specifici nuclei talamici. I nuclei della base agiscono primariamente influenzando le aree motorie primarie, premotorie e supplementari motrici, che agiscono a loro volta attraverso la via corticospinale diretta. I sintomi di pazienti con malattie dei nuclei della base come la malattia di Parkinson e la corea di Huntington hanno dato importanti indicazioni sul ruolo dei nuclei della base nel controllo motorio. In partic., si è osservato che alterazioni funzionali a carico dei nuclei della base determinano alterazioni sia in aumento sia in diminuzione dell’inizio dei m. e della loro ampiezza e velocità. Questi studi, insieme a risultati ottenuti mediante registrazione dell’attività di singoli neuroni, hanno dimostrato che i nuclei della base regolano l’inizio del m. e la sua gradualità attraverso due vie di controllo poste in parallelo e aventi azione opposta. Lo sbilanciamento tra l’attività di queste vie che si verifica nelle neuropatologie dei nuclei della base sarebbe responsabile della marcata sintomatologia motoria di queste malattie. Al contrario delle lesioni a carico dei nuclei della base, le lesioni del cervelletto non provocano acinesia o ipercinesia, bensì errori nella contrazione di molti muscoli. L’organizzazione delle connessioni in entrata e in uscita dal cervelletto indica che esso opera un confronto a feedback negativo tra i segnali sensoriali in ingresso che trasmettono informazioni relative al m. eseguito e una copia del programma motorio. Quando il m. viene ripetuto il cervelletto è in grado di segnalare l’errore operando quindi una correzione anticipatoria del m. attraverso le sue connessioni con la corteccia motoria e i nuclei motori troncoencefalici. Il segnale di errore prodotto dal cervelletto può anche essere memorizzato e costituire un substrato per l’apprendimento motorio. Le oscillazioni dei m. dei soggetti con lesioni cerebellari sarebbero dovute all’assenza di questi meccanismi di correzione. Tommaso Pizzorusso
Apprendimento motorio
I movimenti oculari