
Morfogenesi
Morfogenesi
I Metazoi presentano un'enorme varietà di forme, tipologie di sviluppo e caratteristiche fisiologiche. L'analisi comparata tra le specie esistenti e i loro antenati fossili ha consentito di ordinare tale varietà in taxa e di stabilirne i rapporti filogenetici. I recenti studi di genetica dello sviluppo e l'analisi comparata delle sequenze di DNA in molte specie dimostrano la conservazione dei geni coinvolti nei processi generativi da antenati precedenti il Cambriano. Questa conservazione vale sia per i geni sia per le funzioni a essi correlate. Molti aspetti dei meccanismi alla base del comportamento cellulare durante lo sviluppo sono controllati da geni ereditati addirittura da antenati ancora più antichi: eucarioti monocellulari e persino procarioti. Questa straordinaria conservazione dei geni e delle loro funzioni sembra dovuta alle costrizioni imposte dal riconoscimento molecolare. Ciò significa che l'evoluzione di nuove forme risulta da modifiche nella regolazione dei geni, che determinano combinazioni geniche differenti nelle cellule durante lo sviluppo.
Lo studio della morfogenesi ci consente di comprendere le leggi che intercorrono, nel corso dei processi ontogenetici, tra il livello atomico-molecolare, quello dei geni, quello delle cellule, sino a quello degli organismi. Tuttavia, benché la maggior parte degli organismi abbia origine da uno zigote, la morfogenesi non inizia all'atto della fecondazione; infatti le uova, nel corso della maturazione, modificano il loro contenuto di informazione (prodotti dei geni materni e organizzazione spaziale). Inoltre, esaminando l'aspetto sorprendente della conservazione dei geni, il problema morfogenetico diviene un fenomeno diacronico. La piena comprensione della morfogenesi richiede un'analisi comparativa e un esame puntuale dei prodotti genici conservati, i quali determinano i diversi piani strutturali delle specie attuali e spiegano come questi si siano modificati nel corso dell'evoluzione. La sfida diventa ancor più evidente se si pensa che i piani strutturali fondamentali e le modalità di sviluppo dei Metazoi erano già presenti nella fauna cambriana.
In meno di 50 milioni di anni (da 580 a 530 milioni di anni fa), i Metazoi diploblastici (con due soli foglietti embrionali, endoderma ed ectoderma) hanno dato origine ai principali phyla di organismi triploblastici (con tre foglietti embrionali, ectoderma, mesoderma ed endoderma): i Protostomi, tra cui Anellidi, Molluschi e Artropodi, e i Deuterostomi, cioè Echinodermi e Cordati, un 'big bang' che ha determinato un incredibile aumento della diversità morfologica. Gli organismi fossili dei primi phyla triploblastici sono rappresentati da poche specie con organizzazione e morfologia degli adulti altrettanto complesse di quelle delle specie tuttora esistenti. La documentazione fossile cambriana, costituita da pochi campioni, ci mostra come questa fauna occupasse habitat marini piuttosto omogenei. Pertanto le novità evolutive si sono generate nonostante l'apparente omogeneità e continuità degli habitat che allora esistevano. I processi di natura genetica che hanno determinato tale trasformazione si realizzano tuttora nel corso della morfogenesi dei loro discendenti attuali e possono quindi essere studiati da un punto di vista sia molecolare sia genetico.
Diversi meccanismi del comportamento cellulare
Nel corso dell'evoluzione la conservazione delle sequenze geniche codificanti è dovuta soprattutto all'inerzia imposta dal riconoscimento molecolare, che conferisce specificità alle funzioni geniche, in base alle differenti configurazioni delle strutture tridimensionali delle proteine. I geni che codificano proteine costituite da più domini (una porzione della proteina con una peculiare struttura tridimensionale) consentono così alla stessa proteina di svolgere funzioni differenti. Tuttavia il riconoscimento molecolare, tramite la formazione di legami sia tra proteine uguali sia tra proteine diverse (omo- ed eteromeri), può aver limitato a poche dozzine il numero dei singoli domini funzionali delle proteine. La duplicazione genica, seguita dal rimescolamento dei domini in seguito a ricombinazione intragenica oppure a un accumulo di mutazioni successive e che comporta una sequenza codificante diversa da quella originaria, ha reso possibile la comparsa di famiglie di geni capaci di svolgere funzioni diverse ma correlate tra loro. Nonostante vi sia la possibilità teorica di aumentare all'infinito il numero di geni differenti, questo rimane ancora circoscritto tra 4000 e 5000 nei batteri, fino a qualche decina di migliaia nei Cordati.
La diversità morfologica dei Metazoi cambriani suggerisce come i loro geni fossero già in grado di svolgere un insieme complesso di operazioni correlate ai processi di sviluppo. I geni che controllano i processi di divisione e proliferazione delle cellule eucariotiche sono estremamente conservati nel corso dell'evoluzione e hanno a che fare con tutte le diverse fasi del ciclo cellulare. I fattori di crescita e i segnali per la riattivazione di una cellula quiescente sono simili dai lieviti all'uomo, e anche i segnali che inducono l'apoptosi (morte cellulare) sono conservati a partire dai progenitori di Insetti e Vertebrati. L'orientamento del fuso nel corso dell'ultima fase della mitosi comporta conseguenze per la morfologia della cellula e dipende in ultima analisi dalla ricezione localizzata dei segnali di comunicazione tra le cellule, come avviene durante la divisione cellulare del lievito. Nei Metazoi, le cellule epidermiche mostrano differenze per almeno tre assi cellulari: uno apicale-basale (istologicamente visibile a partire dagli Cnidari) e gli altri due, rispettivamente, antero-posteriore e medio-laterale, tutti deducibili da esperimenti di dissociazione e riaggregazione (negli Insetti) nel quadro del riconoscimento cellulare. Le due ultime specificazioni sono necessarie per le cellule perfino nei Metazoi più primitivi, dal momento che è necessario definire correttamente dimensioni e forma dei territori (organi).
Tutti gli animali triploblastici presentano una notevole diversificazione per quanto riguarda il differenziamento dei diversi tipi cellulari. I tessuti formati da cellule completamente differenziate possono essere istologicamente suddivisi in classi distinte (istotipi), tra cui, per esempio, la classe dei tessuti epiteliale (inclusi dotti e ghiandole), mesenchimale (muscoli e cartilagine) e nervoso. Questi differenti istotipi si formano come derivati dei tre strati germinali e risultano, pertanto, scarsamente correlati a una precisa discendenza da cellule embrionali (eccetto che per i tessuti che si formano dall'endoderma); il loro differenziamento costituisce pertanto il risultato di interazioni tra cellule a livello locale, in cui sono attivati gruppi particolari di geni caratteristici di ciascun istotipo. In cellule di animali diploblastici si può rivelare la presenza di caratteri appartenenti a cellule di istotipi differenti. Negli animali triploblastici, invece, le cellule perdono le proprietà caratteristiche di istotipi differenti per assumere quelle proprie di ciascun tessuto. Il fatto che i geni regolatori della trascrizione (selettori), che controllano gruppi di proteine specifiche per i diversi tipi di cellule, siano comuni a Insetti e Vertebrati attesta quanto lontano nel tempo si possa collocare il processo di differenziamento istotipico. Esistono quindi geni omologhi per numerose funzioni, anche in virtù della somiglianza strutturale che può esistere tra proteine diverse per ciò che riguarda la composizione.
L'omologia di geni coinvolti nello stesso tipo di funzione cellulare, seppure in organismi di phyla diversi, si ritrova costantemente anche in altri geni connessi allo sviluppo. Pertanto appare chiaro che non sono soltanto i geni selettori a venire conservati, ma anche molti dei membri dei rispettivi sintagmi di appartenenza. Inoltre, anche i processi coinvolti nella specificazione degli istotipi, che portano alla formazione di strutture topograficamente delimitate quali sono gli organi, sono sottoposti al controllo di geni che si sono conservati nel corso della filogenesi. Così il cuore degli Insetti e dei Vertebrati viene specificato da geni omologhi al gene tinman di Drosophila e CSX del topo. Il differenziamento del primordio dell'occhio inizia con la sintesi del fotorecettore e la determinazione delle cellule del pigmento; in molti taxa animali questo processo è sotto il controllo di geni omologhi a quelli eyeless di Drosophila e pax6 del topo.
Iterazione modulare di territori cellulari e di tipi di cellule
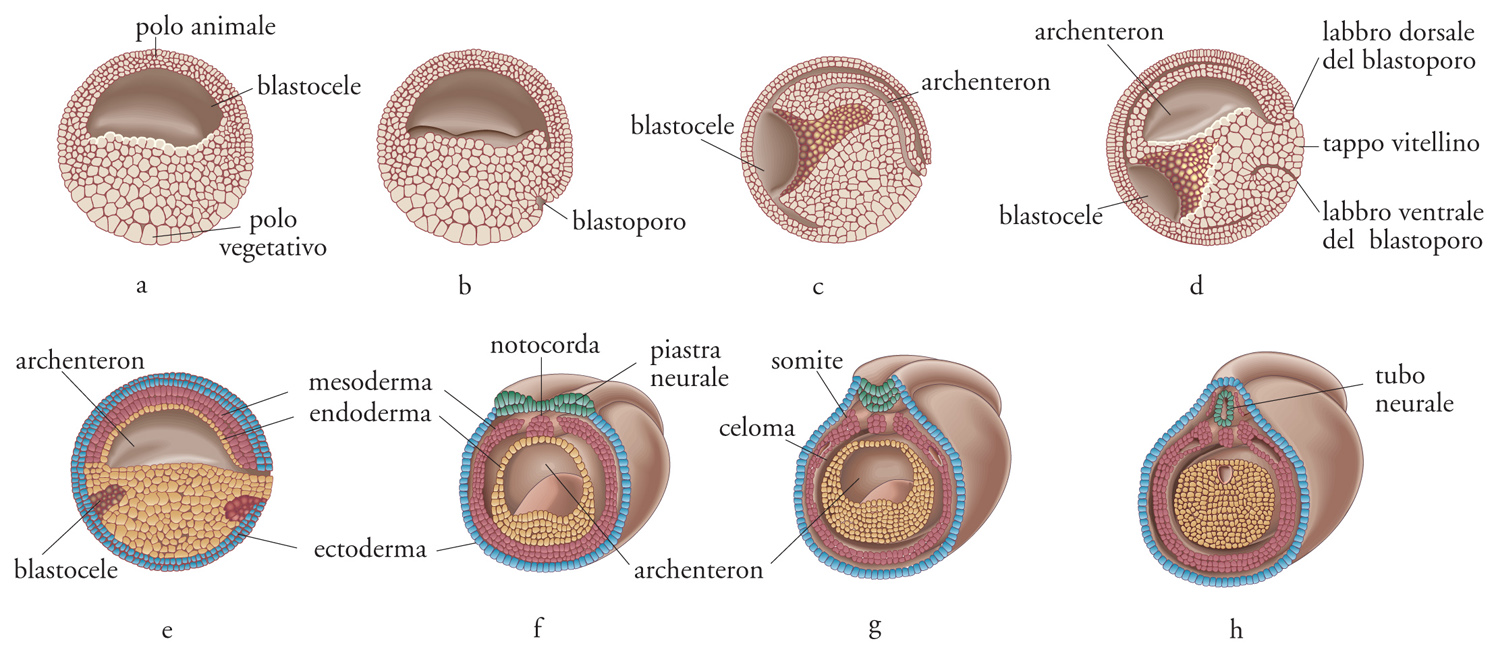
Di regola nel corso della segmentazione dello zigote si formano i blastomeri, in cui è già definita la futura topologia delle differenti parti del corpo e così pure gli assi corporei. La segmentazione può essere radiale o spirale, ma entrambe le modalità non hanno conseguenze morfogenetiche rilevanti, poiché sia nel corso dell'una sia dell'altra si possono formare organi tanto simmetrici quanto asimmetrici. La determinazione dell'asse corporeo potrebbe derivare dalla distribuzione anisotropa dei prodotti dei geni di origine materna, oppure risultare dal rimescolamento dei componenti citoplasmatici. Generalmente le anisotropie dell'uovo portano alla definizione di un asse cui segue immediatamente la definizione dell'altro, ortogonale al primo; in tal modo si definiscono i piani di simmetria antero-posteriore (A-P) e dorso-ventrale (D-V), ed è indifferente quale dei due piani appaia per primo. Negli animali a simmetria bilaterale, l'intersezione degli assi A-P e D-V definisce il piano dorso-ventrale e quindi la parte destra e quella sinistra dell'animale. In tutti gli organismi triploblastici a simmetria bilaterale, deuterostomi e protostomi, l'asse A-P è stabilito da un determinato gruppo di geni conservati che definiscono in sequenza specifica la regione cefalica, quella del tronco e quella della coda. Nel corso della gastrulazione (per invaginazione o delaminazione), i precursori del tubo digerente e degli organi di origine mesodermica si invaginano nella cavità del blastocele (fig. 2). Una seconda apertura di questo tubo digerente primitivo di origine endodermica dà luogo, rispettivamente, al futuro apparato boccale (stomodeo) nei Deuterostomi e al futuro intestino retto (proctodeo) nei Protostomi.
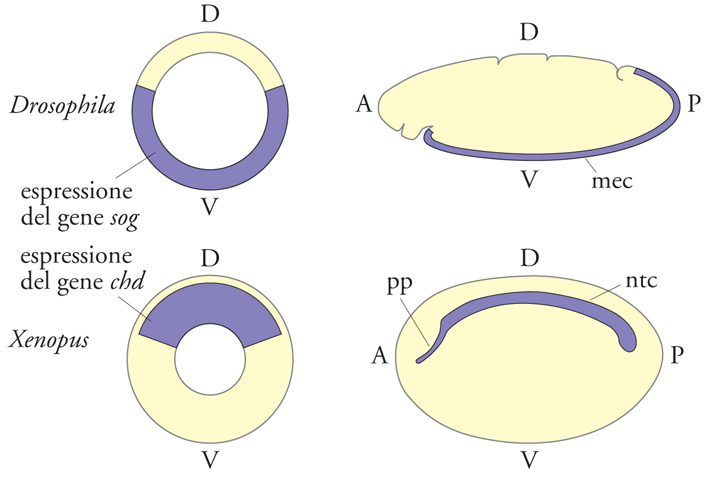
Oltre agli organi che derivano dal foglietto embrionale esterno (di origine ectodermica) e a quelli che derivano dal foglietto embrionale interno (di origine endodermica), si differenziano altri organi a partire dal foglietto di cellule compreso tra l'ectoderma e l'endoderma, il mesoderma, e che sono, appunto, organi di origine mesodermica. I tessuti neurali derivano generalmente da territori ectodermici e, nell'organismo adulto, rimarranno medio-dorsali (come nei Cordati, tra i Deuterostomi) oppure laterali e, tramite fusione ventrale successiva, anatomicamente medio-ventrali. Considerando come riferimento l'asse D-V, le posizioni anatomiche del sistema nervoso, dell'aorta e del tubo digerente risultano pertanto invertite nei Deuterostomi e Protostomi a simmetria bilaterale (fig. 3). Tuttavia, nel corso della filogenesi si sono conservati a partire dai progenitori comuni anche i geni che specificano, attraverso antagonismo reciproco, il territorio dorsale e quello ventrale nel blastoderma.
Nei progenitori triploblastici, gli stessi processi di sviluppo, quali l'invaginazione e la delaminazione che hanno luogo nel corso della gastrulazione, devono aver portato alla formazione di un'apertura verso l'esterno con funzioni inverse nei Deuterostomi rispetto ai Protostomi e con conseguenti posizioni anatomiche diverse negli uni rispetto agli altri non solo per la bocca e l'ano, ma anche per il sistema nervoso centrale e l'aorta. La conoscenza di questi particolari rapporti anatomici e dei processi che portano al differenziamento delle cellule e dei tessuti, con la formazione dei diversi organi, consente di evidenziare omologie esistenti tra i vari phyla, che all'osservazione topologica potrebbero sembrare semplici analogie. La gastrulazione al tempo stesso è determinata e influisce sulle differenze presenti lungo gli assi corporei antero-posteriore e dorso-ventrale. Nei Protostomi (così come nei Cordati fra i Deuterostomi), lungo l'asse A-P si specifica la reiterazione dei territori. Tale reiterazione si manifesta in una ripetizione dei segmenti (metameria) per tutti gli organi derivati dai foglietti embrionali (incluso, in parte, il tubo digerente).
Dalle comparazioni genetico-molecolari sta emergendo il fatto che differenze, sia a livello globale sia locale, sono controllate da geni conservati a partire dai progenitori comuni ai Protostomi e ai Deuterostomi. I primitivi geni HOX (regolatori della trascrizione) appaiono, nella loro versione più arcaica, come tre membri di una medesima famiglia, espressi rispettivamente nella regione cefalica, mediana e terminale del corpo, in una medesima sequenza cefalo-caudale. È evidente come il loro allineamento sul genoma corrispondesse esattamente alla successione delle regioni corporee specificate da questi geni lungo l'asse A-P dell'embrione e anche al loro ordine di espressione antero-posteriore. Tale schema di espressione si presenta secondo questa successione anatomica anche in organismi non segmentati, quali i Molluschi e gli Echinodermi. La successiva evoluzione molecolare mediante duplicazione genica dei membri di questa famiglia ha generato fino a dieci complessi di geni, i cui membri relativi mantengono la stessa corrispondenza spaziale lungo l'asse A-P sia nei Vertebrati sia negli Insetti, suggerendo così un'origine comune e un medesimo ruolo nei processi di sviluppo dei diversi organismi. I complessi di geni specificano l'eterogeneità tra gruppi di metameri o tra singoli metameri, perché sia in Drosophila sia nel topo le loro mutazioni causano sostituzioni omeotiche (mutazioni a seguito delle quali si ha la trasformazione di una parte del corpo in un'altra). Questo vale anche per tutti quegli organismi, sia Insetti sia Vertebrati, in cui l'espressione dei geni HOX coincide con le eterogeneità dorso-ventrali associate ai differenti istotipi in tutti i foglietti germinali di un determinato segmento.
In Drosophila l'analisi dei mutanti a mosaico dei geni HOX ha evidenziato come, per i derivati epidermici, gli effetti morfogenetici di questi geni si manifestano come cellule autonome capaci di definire il comportamento cellulare specifico di un determinato segmento (modalità di proliferazione e di differenziamento). Pertanto i geni HOX controllano, in qualità di geni selettori, l'espressione dei geni bersaglio (geni target, altrimenti detti geni realizzatori; alcuni di questi sono anche membri di sintagmi conservati) che definiscono il comportamento dei territori cellulari coordinando il legame di specifici fattori di crescita, con i rispettivi recettori. Attualmente si conoscono molti geni selettori comuni a Insetti e topi, che specificano territori cellulari e che sono conservati sin dai progenitori comuni del Cambriano. Questi includono quei geni che codificano proteine con differenti motivi strutturali, i quali controllano la trascrizione mediante il riconoscimento diretto delle sequenze promotrici del DNA. Alcuni di questi geni e i loro sintagmi possono funzionare contemporaneamente nella stessa cellula, generando mosaici di territori secondo le differenti combinazioni. Mentre i singoli processi genetici che essi controllano sono omologhi, cioè conservati, la loro combinazione differenziale nello spazio (e nel tempo dello sviluppo) dà luogo, nei differenti phyla, a strutture analoghe o completamente diverse.
L'origine della metameria
Il fenomeno della metameria suscita diverse domande, tra cui come questa si sia potuta generare, in quale modo venga fissato il numero di metameri e come venga specificata la loro sequenza spaziale. Queste domande sono comprese in un'altra domanda di ordine più generale: come si generano gli insiemi sovracellulari e come riescono questi ad assumere una forma specifica? Come possono gli organismi animali e le differenti parti di cui sono costituiti raggiungere le dimensioni proprie a seconda della specie di appartenenza? Lo studio delle genealogie cellulari e dei dati provenienti dalla genetica molecolare suggerisce alcune possibili spiegazioni. Il meccanismo che determina metameria lungo l'asse A-P è strettamente collegato a quello che determina, a sua volta, differenze cellulari istotipiche lungo l'asse D-V, con susseguente formazione di territori bidimensionali di grandezza specifica della specie di appartenenza. All'interno di questi territori si evidenziano piani progettuali di differenziamento più fine. Così nel neuroectoderma, singole cellule si differenziano in neuroblasti con una netta diversificazione dagli epidermoblasti circostanti. Lo stesso avviene per le cellule epidermiche che si differenziano in scaglie o in aculei oppure in gruppi di mesodermoblasti. Ma come spiegare l'esistenza di tali piani progettuali? Cercheremo di individuare, attraverso una descrizione generale, il modo in cui si può essere prodotta la specificazione dei segmenti negli organismi metamerici.
L'iterazione metamerica può essere rappresentata visivamente da onde con valori massimi (ampiezza dell'onda) e valori intermedi a livello delle pendenze; possiamo così definire un parametro, il cui valore numerico è correlabile al numero delle cellule dell'embrione in quell'area, che potremmo chiamare 'lunghezza d'onda'. L'analisi delle discendenze cellulari nei territori metamerici consente di evidenziare le due diverse modalità di proliferazione delle cellule. Negli Anellidi (per es., la sanguisuga) i metameri mediani sono rappresentati da strutture periodiche generate per anamorfosi: cinque teloblasti (grosse cellule che si distinguono precocemente nell'uovo in segmentazione degli Anellidi, così come nei Molluschi e in alcuni Artropodi), a simmetria bilaterale (designati MNOPQ, seguendo l'asse D-V), danno origine, mediante divisioni asimmetriche, a due bande.
Le divisioni asimmetriche dei teloblasti avvengono secondo ritmi differenti, corrispondenti alle diverse lunghezze d'onda e caratteristici di ciascun teloblasto. I gruppi risultanti di cellule (5÷10), o blastocellule primitive, costituiscono il metamero primitivo. Le due bande sono suddivise, lungo l'asse D-V, in raggruppamenti di cellule precursori di istotipi diversi (ectoderma, neuroectoderma e mesoderma), specifici a seconda del teloblasto da cui discendono e organizzati in una determinata sequenza dorso-ventrale per ciascun metamero (lo stesso avviene negli Insetti). La divisione cellulare per diamorfosi delle blastocellule originarie porta all'espansione dei metameri primitivi. Il differenziamento delle cellule muscolari e nefridiali, dei neuroni motori e sensoriali e degli epidermoblasti avviene in un tempo successivo, dando origine a modelli ad andamento periodico. Benché essi siano leggermente sfasati l'uno rispetto all'altro, il numero totale dei segmenti subesofagei è costante (32 nella sanguisuga), suggerendo così l'ipotesi che essi corrispondano a multipli di differenti lunghezze d'onda, come avviene nelle divisioni dei teloblasti. Mentre i segmenti sono molto simili l'uno all'altro (cioè omonimi), gli orifizi riproduttivi e le altre specializzazioni segmentali compaiono in segmenti riconducibili a un'identificazione numerica specifica (eteronomia).
In Drosophila la periodicità dei segmenti dipende dal modo in cui le varie e diverse lunghezze d'onda sono articolate tra loro. Le articolazioni lunghe sono quelle che coinvolgono quattro segmenti presuntivi e sono definite dall'espressione dei geni gap (geni zigotici espressi nel sincizio blastodermico, la cui trascrizione è controllata da proteine di origine materna e, a loro volta, coinvolti nella definizione sia dell'asse A-P sia della regione addominale). Le articolazioni di lunghezza media si realizzano lungo due segmenti presuntivi e sono definite dai geni chiamati pair-rule (geni anch'essi zigotici, la cui trascrizione è controllata da proteine di origine materna e da quelle codificate dai geni gap, che mediano l'ulteriore definizione dell'organizzazione dell'embrione lungo l'asse AP). Infine le articolazioni corte sono definite da geni segment-polarity (geni zigotici, espressi quando l'embrione è ormai totalmente formato da cellule, e che definiscono la polarità della distribuzione delle cellule entro un dato parasegmento).
I confini entro i quali sono espressi i geni pair-rule e segment-polarity definiscono i campi d'espressione del complesso genico Antennapedia-Ultrabithorax (ANT-C e BX-C) e dei geni HOX che specificano l'identità dei segmenti e, quindi, le strutture delle varie parti del corpo (geni selettori). Negli Artropodi i singoli segmenti sono suddivisi in due compartimenti, rispettivamente anteriore (A) e posteriore (P); quest'ultimo è specificato in tutti i segmenti dall'espressione del gene omeobox engrailed, il cui confine anteriore di espressione definisce iterativamente quello del segmento primigenio (parasegmento). Così, l'ordine di comparsa dei segmenti potrebbe seguire un meccanismo secondo il quale i geni, la cui differente area d'espressione determina varie lunghezze d'onda, inducendo combinazioni diverse tra i prodotti quando le cellule si differenziano a partire dai teloblasti, generano segmenti di numero definito, con gli istotipi in registro. La particolare specificazione dei segmenti da parte dei geni HOX si realizza in modo autonomo relativamente alla singola progenie di derivazione teloblastica e non deriva dalle interazioni cellulari, cioè da informazioni relative alla posizione tra le blastocellule primitive.
Negli Anellidi, negli Insetti e nei Cordati, i confini territoriali dell'espressione differenziale dei geni HOX (e PAX) appaiono in successione, con limiti netti e discontinui nei segmenti cefalici e del tronco, ma si sovrappongono via via nei segmenti più caudali. Nel tronco di entrambi i phyla, l'ordine di comparsa dei segmenti è rostro-caudale. Questo comporta che, sia per gli Artropodi sia per i Cordati, i nuovi segmenti pregnatali sono comparsi secondo una sequenza caudo-rostrale. Questi nuovi segmenti possono essere omologhi a strutture cefaliche della sanguisuga che non derivano dai teloblasti del tronco. Si può affermare che i geni territoriali, mentre specificano l'identità di gruppi di cellule mediante le loro relazioni regolative, potrebbero avere la funzione di definire specifiche sequenze topografiche e, di conseguenza, il numero dei segmenti. Un numero costante di segmenti è caratteristico dei Crostacei, dei Chelicerati e degli Onicofori cambriani (precursori degli Insetti). Al contrario, i Platelminti, i primi Anellidi (Polychaeta) e i Cordati possono avere un numero indefinito di segmenti.
Appendici e organi
Il fatto che i segmenti embrionali siano definiti dai geni HOX in bande della larghezza di poche cellule lungo l'asse A-P sembra costituire una regola. Queste bande si espandono in seguito in senso medio-laterale, seguendo una proliferazione cellulare di tipo simmetrico (diamorfosi), mentre possono comparire nuove periodicità di ampiezza non superiore ad alcune cellule. Il processo è reiterato, dando così origine a spazi morfologici con lunghezza d'onda sempre più corta, proprio come si osserva nell'organizzazione finale. È sempre il medesimo processo che funziona nella fase successiva dello sviluppo, durante l'organogenesi. In Drosophila, possiamo studiare dettagliatamente lo svolgimento di questo processo, durante la formazione delle appendici. Queste compaiono in campi embrionali secondari, nei quali vengono ricapitolate le diverse fasi di sviluppo già sperimentate nella formazione dei segmenti. Le appendici sono escrescenze reiterative che originano al confine A-P dei compartimenti all'interno di ciascun segmento embrionale, con primordi costituiti da poche cellule (meno di trenta), di cui circa la metà si trova nel compartimento A e l'altra in quello P. I primordi si accrescono, generando per diamorfosi una struttura bidimensionale su entrambi i lati del confine. In Drosophila, i raggruppamenti di cellule hanno un'origine policlonale, cioè derivano dall'interazione di cellule la cui genealogia non è direttamente correlata. L'attività dei geni selettori nelle cellule di ciascun compartimento fa sì che queste vengano contrassegnate da molecole di riconoscimento che impediscono alle cellule di un compartimento di mescolarsi con quelle del compartimento adiacente, attraverso un processo chiamato di restrizione clonale. I confini, quindi, corrispondono alle discontinuità nell'espressione genica dei geni selettori.
Nelle appendici, inoltre, i confini fungono da assi di simmetria e, lungo questi margini, le cellule si scambiano segnali, fattori di crescita e rispettivi ligandi (dei quali esistono versioni omologhe in Drosophila e nei Vertebrati), usati sia per limitare l'espressione dei geni selettori sia per indurre la proliferazione cellulare. In tal senso, i confini agiscono su ambedue i lati come 'organizzatori' per la proliferazione simmetrica e la polarità delle cellule. La suddivisione antero-posteriore del primordio embrionale, associata alla specificazione del gene en (engrailed), viene mantenuta durante la proliferazione successiva. In seguito a ulteriore crescita, compare un nuovo asse di simmetria che separa le cellule dorsali da quelle ventrali lungo un nuovo confine tra compartimenti che si escludono mutualmente; ciò è determinato dall'espressione di un nuovo gene selettore, apterous (ap), che specifica le cellule dorsali in opposizione a quelle ventrali. La sua funzione mobilita, ancora una volta, sia fattori di crescita sia recettori degli stessi, determinando una restrizione clonale lungo il nuovo margine, da una e dall'altra parte del quale si determina un incremento della proliferazione cellulare.
Viene così a formarsi un nuovo compartimento che separa le regioni prossimali (noto) da quelle distali (ala vera e propria). Il primordio dell'ala è costituito a questo stadio da circa duecento cellule, cinquanta per compartimento. La proliferazione cellulare simmetrica all'interno dei quattro compartimenti dell'ala è ulteriormente stimolata da differenze nell'espressione di altri insiemi di geni regolatori in senso prossimo-distale, distalles (dll), nubbin (nub), aristaless (al), e dei loro ligandi (dpp, wgl); questi, agendo ai margini dei compartimenti, trasmettono messaggi di crescita. La proliferazione cellulare continua in modo diamorfico per tutte le cellule del primordio. Con l'aumentare delle dimensioni in seguito all'ulteriore proliferazione, si verificano nuove restrizioni clonali che portano alla suddivisione di ciascun compartimento in tre bande, i cui margini corrispondono alle venature delle ali adulte. Pertanto, il primordio dell'ala, così come quelli delle appendici ventrali (antenne, parti della bocca, arti e genitali esterni), viene a suddividersi durante la crescita in territori specificati da geni diversi, i cui margini agiscono da referenti per la proliferazione.
Questo tipo diamorfico simmetrico di crescita cellulare entro confini delimitati di volta in volta da una determinata espressione genica è, probabilmente, un processo attivo in tutti i Metazoi. La rigenerazione e il comportamento di alcuni tipi di cellule mutanti nei mosaici genetici rivelano eterogeneità spaziali che riflettono 'valori posizionali' differenziali. I valori sono massimi ai margini dell'area nella quale tale espressione è circoscritta, minimi nel punto centrale e intermedi nella zona compresa tra i due. In questo panorama di valori posizionali, le varie cellule mutanti per loci diversi possono crescere nella direzione delle creste o degli avvallamenti oppure, alternativamente, nella direzione opposta, a seconda del gene considerato e della sua condizione allelica (in omozigosi o eterozigosi). Le specificazioni territoriali e il differenziamento delle cellule, secondo modelli precisi, possono limitarsi alla formazione di discendenze cellulari specifiche (monocloni) oppure rappresentare il risultato della coordinazione di cellule adiacenti mediante interazioni cellula-cellula (policloni). I confini possono essere manifesti prima che cessi la proliferazione (ed evidenziarsi così all'analisi della discendenza cellulare) oppure comparire soltanto alla fine di questa fase. Non sono comunque solo le appendici e le branchie ad avere un'origine metamerica, ma anche i neuroni (motori e sensori), i condrociti precursori dei muscoli e delle ossa, nonché le strutture tubulari, come i nefridi, le vene e la trachea. Di conseguenza, sono tutti specificati primariamente dall'azione di meccanismi di regolazione di tipo combinatorio esercitata dai geni selettori con competenza territoriale.
Bibliografia
Akam 1995: Akam, Michael, Hox genes and the evolution of diverse body plans, "Philosophical transactions of the Royal Society of London. Series B", 349, 1995, pp. 313-319.
Carroll 2005: Carroll, Sean B., Endless forms most beautiful, New York, Norton, 2005.
Conway Morris 1993: Conway Morris, Simon, The fossil record and the early evolution of the Metazoa, "Nature", 361, 1993, pp. 219-225.
Davidson, Erwin 2006: Davidson, Eric H. - Erwin, Douglas H., Gene regulatory networks and the evolution of animal body plans, "Science", 311, 2006, pp. 796-800.
De Robertis, Sasai 1996: De Robertis, Eddy M. - Sasai, Yoshiki, A common plan for dorsoventral patterning in bilateria, "Nature", 380, 1996, pp. 37-40.
García-Bellido 1985: García-Bellido, Antonio, Cell lineages and genes, "Philosophical transactions of the Royal Society of London. Series B", 312, 1985, pp. 101-128.
García-Bellido, de Celis 1992: García-Bellido, Antonio - de Celis, José F., Developmental genetics of the venation pattern of Drosophila, "Annual review of genetics", 26, 1992, pp. 277-304.
Knoll 1992: Knoll, Andrew H., The early evolution of Eukaryotes: a geological perspective, "Science", 256, 1992, pp. 622-627.
Liu, Kaufman 2005: Liu, Paul Z. - Kaufman, Thomas C., Short and long germ segmentation: unanswered questions in the evolution of a developmental mode, "Evolution and development", 7, 2005, pp. 629-646.
Minelli 2003: Minelli, Alessandro, The development of animal form: ontogeny, morphology, and evolution, Cambridge, Cambridge University Press, 2003.
Nurse 2003: Nurse, Paul, Cell cycle control, "Philosophical transactions of the Royal Society of London. Series B", 341, 1993, pp. 449-454.
Slack 1993: Slack, Jonathan M.W. - Holland, Peter W.H. - Graham, Christopher F., The zootype and the phylotypic stage, "Nature", 361, 1993, pp. 490-492.
Tannahill 2005: Tannahill, David - Harris, Laura W. - Keynes, Roger, Role of morphogens in brain growth, "Journal of neurobiology", 64, 2005, pp. 367-375.
Zhao, Emmons 1995: Zhao, Connie - Emmons, Scott W., A transcription factor controlling development of peripheral sense organs in C. elegans, "Nature", 373, 1995, pp. 74-78.