meccanotrasduzione
meccanotrasduzione
Nella meccanotrasduzione un segnale meccanico viene convertito in un segnale nervoso tramite i meccanorecettori. Alla base di questa forma di trasduzione ci sono i canali ionici a controllo meccanico. Nel caso della trasmissione del suono, la meccanica degli elementi dell’orecchio interno trasmette alle cellule ciliate acustiche la vibrazione causata dall’arrivo di onde sonore; il movimento del fascetto di cilia e la sua particolare struttura fanno sì che la vibrazione determini una variazione periodica della tensione esercitata sui canali ionici, che si aprono e si chiudono causando cicli di depolarizzazione e iperpolarizzazione che costituiscono il segnale nervoso. Le cellule ciliate esterne svolgono un ruolo di amplificazione della vibrazione e quindi del segnale nervoso. Nella recezione tattile l’infossamento della cute innervata dai recettori tattili, causato da uno stimolo, determina una deformazione della loro membrana con conseguente apertura dei canali ionici e depolarizzazione. La sensibilità alla deformazione meccanica è una caratteristica dei meccanorecettori, mentre il tipo di risposta che viene data alla deformazione, e quindi la funzione fisiologica che essi svolgono, dipende dalla struttura specializzata, che differisce tra i vari meccanorecettori tattili, con cui essi sono associati.n [➔ recettore nervoso; tatto; trasduzione; udito] La m. fa parte del processo di trasduzione attraverso il quale recettori sensoriali, detti meccanorecettori (MR), convertono un segnale meccanico in un segnale nervoso, ossia in variazioni del potenziale di membrana. La m. è effettuata dalle cellule ciliate dell’orecchio interno, per l’udito e per l’equilibrio, e dalle cellule del sistema somatosensoriale, per il tatto, la propriocezione e la nocicezione; è effettuata anche da cellule non sensoriali, che fungono da sensori per la pressione arteriosa (➔ pressocettore), per mantenere sotto controllo questa variabile cruciale alla sopravvivenza dell’organismo.
Caratteristiche
La capacità di convertire uno stimolo meccanico esterno in variazioni del potenziale di membrana è data dai canali ionici a controllo meccanico (CICM) presenti sugli MR; i CICM, formati da proteine di membrana, sono normalmente chiusi e vengono aperti da stimoli meccanici. Anche se possono differire tra loro per struttura e funzione, tutti i CICM condividono la caratteristica di aprirsi in risposta a stimoli che deformano meccanicamente la membrana su cui sono inseriti. La m. consiste quindi nella deformazione della membrana degli MR, con conseguente apertura dei CICM, passaggio di ioni e variazione del potenziale di membrana (potenziale di recettore, PDR), segno dell’avvenuta meccanotrasduzione. L’efficacia della m. dipende dal tipo di stimoli meccanici in grado di determinare una deformazione o un cambiamento di tensione nella membrana di un tipo di MR. Per es., le cellule ciliate (CC) dell’orecchio interno, poste nella coclea, sono sensibili agli effetti delle onde di pressione nell’aria e, tramite m., danno inizio al processo che conduce alla percezione uditiva; invece le CC dell’orecchio interno, poste nell’apparato vestibolare, sono sensibili alla posizione e al movimento della testa e, tramite m., danno origine al processo che media l’equilibrio e il suo controllo. La differenza fra i due tipi di sensibilità (onde di pressione e movimenti della testa), non è dovuta a differenze tra le CC cocleari o vestibolari o ai rispettivi processi di m., praticamente identici, ma semplicemente alla collocazione delle cellule: la porzione acustica dell’orecchio interno è progettata per causare il movimento delle cilia delle CC cocleari, che determina un cambiamento di tensione sulla membrana, in risposta alle onde di pressione, mentre la porzione vestibolare è progettata per causare il movimento delle cilia delle CC vestibolari in risposta ai movimenti della testa. Qui esemplificheremo due tipi di m.: quella delle CC cocleari e la m. dei recettori tattili, come esempio di m. somatosensoriale.
Meccanotrasduzione acustica
Il movimento delle cilia delle CC cocleari dipende dalla meccanica degli elementi dell’orecchio. Gli stimoli acustici sono rappresentati dalle onde di pressione; sono quindi definibili come una successione di compressioni e rarefazioni che si propaga in un mezzo.

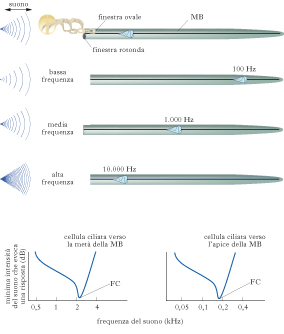
Trasmissione del suono. Solo le onde di pressione la cui frequenza è tale da evocare una risposta nelle CC cocleari evocano una sensazione uditiva (➔ udito). L’onda sonora è catturata dal padiglione auricolare e convogliata nel canale uditivo o meato acustico, chiuso dalla membrana del timpano. La vibrazione della membrana del timpano si trasmette alla catena degli ossicini dell’orecchio medio, di cui il primo, il martello, poggia sul timpano e l’ultimo, la staffa, poggia sulla membrana della finestra ovale, che è in contatto con i liquidi endococleari dell’orecchio interno. Nell’orecchio medio c’è una seconda finestra, la finestra rotonda, che separa l’aria dell’orecchio medio dai fluidi endococleari. Quando la conduzione del suono da parte della catena degli ossicini è difettosa, c’è una perdita di udito, detta sordità conduttiva. L’orecchio interno, scavato all’interno dell’osso temporale, contiene la coclea e il labirinto vestibolare (utricolo, sacculo e canali semicircolari). L’interno della coclea, a sua volta, è diviso in tre compartimenti: la scala vestibolare, che è il compartimento superiore e alla cui base c’è la finestra ovale, la scala media, o dotto cocleare, e la scala timpanica, il compartimento inferiore, alla cui base c’è la finestra rotonda. La scala vestibolare e quella timpanica sono riempite dello stesso fluido, la perilinfa. La scala media è completamente separata dalle altre due scale ed è ripiena di un liquido, l’endolinfa, la cui composizione ionica è diversa da quella della perilinfa e, in partic., è molto più ricca di ioni K+. La membrana che separa la scala media dalla scala timpanica è la membrana basilare, su cui poggia l’organo del Corti, che contiene le CC cocleari.
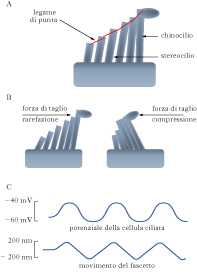
Ruolo delle cellule cocleari. Esistono tre file di cellule ciliate esterne (CCE) e una fila di cellule ciliate interne (CCI, ca. 16.000 cellule ciliate interne nell’uomo), le cui cilia sono a contatto con la membrana che funge da ‘tetto’all’organo del Corti (membrana tectoria). Le CC, sia quelle esterne sia quelle interne, possiedono all’apice un fascetto di cilia la cui lunghezza cresce passando da un cilio all’altro del fascetto, fino al cilio più alto. Nella fase di compressione del ciclo dell’onda sonora, la staffa spinge sulla membrana della finestra ovale e questa affonda nella perilinfa della scala vestibolare. Questo determina un movimento della perilinfa che causa la deflessione della membrana della finestra rotonda verso l’orecchio medio, spingendo verso il basso la membrana basilare. Nella fase di rarefazione avviene l’opposto e la membrana basilare si muove verso l’alto. Quindi, l’oscillazione avanti-indietro della membrana del timpano a ogni ciclo dell’onda sonora si traduce in una oscillazione altobasso della membrana basilare, la quale determina una forza di taglio fra la membrana tectoria e le cilia delle CC, che si piegano, alternativamente, verso il cilio più alto quando la membrana basilare deflette verso l’alto e verso il cilio più basso quando la membrana basilare deflette verso il basso. Ogni cilio è legato al successivo da una connessione filamentosa, detta legame di punta, le cui alterazioni causano sordità. La tensione esercitata dai legami di punta regola l’apertura dei CICM espressi dalle CC cocleari : la permeabilità di questi canali consente il passaggio di una corrente depolarizzante trasportata dagli ioni K+, molto più concentrati nella endolinfa che all’interno della CC, e dagli ioni Ca2+. In condizioni di riposo, la tensione esercitata dai legami di punta è piccola e pochi CICM saranno aperti: di conseguenza, la corrente trasportata dagli ioni K+ sarà molto piccola. Quando il fascetto di cilia viene deflesso nella direzione del cilio più alto, la tensione esercitata dai legami di punta aumenta, un maggior numero di CICM si apre e una più ampia corrente depolarizzante fluisce all’interno della cellula; quando il fascetto viene deflesso nella direzione opposta, i legami di punta si afflosciano, la tensione locale diminuisce, la maggior parte dei canali si chiude e la cellula si iperpolarizza. Così, mentre la pressione alla membrana del timpano oscilla, per es., mille volte al secondo per l’arrivo di un suono di frequenza 1.000 Hz, il potenziale di membrana delle CC depolarizza e iperpolarizza 1.000 volte al secondo. L’ampiezza della risposta depolarizzante e iperpolarizzante aumenta progressivamente con l’intensità del suono fino a una ampiezza di risposta massima, oltre la quale ulteriori aumenti dell’intensità del suono non provocano aumenti dell’ampiezza di risposta (saturazione); la minima intensità del suono che provoca una risposta nelle CC è invece detta soglia. Per potersi aprire e chiudere affidabilmente fino a 20.000 volte al secondo, il meccanismo di apertura dei canali di m. deve essere rapidissimo: alcune stime danno tempi di apertura dell’ordine di pochi microsecondi. A fronte del lento processo di fototrasduzione (➔), la rapidità e la semplicità della m. acustica sono impressionanti. Del resto, la velocità e l’accuratezza nel dominio del tempo sono la caratteristica principale di lavoro del sistema acustico e vengono curate fin dal processo di trasduzione.
Amplificazione del segnale da parte delle cellule cocleari esterne. Sulla base delle caratteristiche della membrana basilare, più larga ed elastica all’apice, più stretta e rigida alla base, già Hermann von Helmholz aveva ipotizzato che essa funzionasse come una serie di risonatori, in grado di scomporre un suono dalla forma d’onda complessa nelle frequenze componenti (mappa tono-topica). Tuttavia la sensibilità delle CC della coclea è troppo grande e la selettività per le diverse frequenze acustiche troppo elevata per dipendere soltanto dalle proprietà passive della membrana basilare: la coclea deve possedere un modo attivo di amplificare l’energia sonora. Numerose evidenze indicano che la sorgente dell’amplificazione attiva della coclea, che contribuisce sia alla selettività per la frequenza del suono sia alla sensibilità cocleare, sono le CCE, che hanno la peculiare capacità di cambiare lunghezza in funzione della depolarizzazione. Le CCE attivate si accorciano e si allungano in modo periodico, con frequenza uguale a quella del suono. Le CCE attive possono quindi amplificare l’oscillazione locale della membrana basilare attraverso i loro movimenti oscillatori. Dato che l’ampiezza dell’oscillazione delle CCE dipende dall’entità della loro attivazione, tale ampiezza sarà massima nella ristretta regione della membrana basilare che corrisponde alla frequenza del suono a cui le locali CCE sono massimamente sensibili. In questo modo le CCE contribuiscono ad amplificare selettivamente la risposta oscillatoria, e quindi il segnale delle CCI, in una ristretta zona della membrana basilare. Se si viene esposti a un suono di grande intensità, l’ingresso massiccio di Ca2+ attraverso i CICM innesca processi di adattamento, che riducono la sensibilità delle CC determinando lo slittamento dei CICM verso il basso nelle cilia, con conseguente riduzione della tensione dei legami di punta e chiusura dei canali stessi. Riduzione di sensibilità vuol dire aumento della soglia: un suono di debole intensità, in grado di evocare una risposta nelle CC, e quindi per noi udibile prima dell’adattamento, non lo è più dopo. Questo è il motivo per cui, dopo essere stati esposti a un suono molto forte, si ha una sensazione di ottundimento delle capacità acustiche. Il processo di adattamento, come sempre nelle cellule recettoriali, serve a conciliare la possibilità di rispondere a stimoli di piccola intensità, che richiedono una sensibilità elevata, con la necessità di evitare la saturazione già a bassi livelli di intensità dello stimolo, che sarebbe appunto conseguenza di una sensibilità elevata fissa. L’adattamento non va confuso con gli effetti meccanici di una esposizione prolungata a suoni di intensità molto elevata, che possono provocare danni irreversibili alle CC, determinando perdite gravi e permanenti dell’udito. Le CC inviano il loro messaggio alle cellule del nervo acustico, il cui corpo cellulare è nel ganglio del Corti, nell’orecchio interno. Le cellule del nervo acustico sono dotate di assone e generano potenziali d’azione che trasporteranno l’informazione verso le successive strutture lungo le vie acustiche. Le cellule del nervo acustico mostrano una selettività per la frequenza del suono: cellule che innervano la base della coclea hanno la massima sensibilità per le frequenze alte, quelle che innervano l’apice per le frequenze basse. Questo arrangiamento spaziale ha consentito lo sviluppo di protesi (impianti cocleari), che possono supplire con successo a situazioni di sordità dovute alla degenerazione o al mancato sviluppo delle CC. La protesi consta di un microfono per captare i suoni e trasdurli in segnali elettrici, (sostituendo così le CC mancanti), un processore per scomporli in frequenze semplici e una serie di elettrodi per stimolare le fibre del nervo acustico.
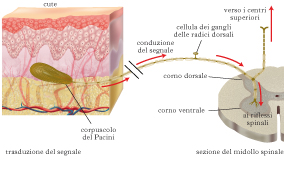
Meccanotrasduzione somatosensoriale
La sensibilità somatica ha origine da recettori distribuiti nella pelle, nei muscoli e nelle articolazioni, negli organi interni. Il sistema somatosensoriale segnala stimoli meccanici molto differenti fra loro, da uno stimolo doloroso come una puntura con un ago (modalità nocicettiva), a un lievissimo sfioramento della pelle con una piuma (modalità tattile, ➔ tatto), al cambiamento di posizione della mano rispetto al polso (modalità propriocettiva). Il corpo cellulare dei neuroni dei gangli delle radici dorsali si trova nei gangli delle radici dorsali di un nervo spinale. L’assone ha due branche, una periferica e una centrale, che entra nel midollo spinale dalle radici dorsali. Alla fine della branca periferica si trova il terminale recettoriale, che svolge il processo di m., generando il PDR. Se l’ampiezza del PDR supera la soglia per la generazione di potenziali d’azione, questi trasportano il segnale verso il midollo spinale, per i riflessi, e verso i centri superiori, per la percezione cosciente.
I recettori tattili. Sono stati identificati quattro tipi principali di MR tattili nella pelle glabra (priva di peli). Due tipi di MR sono collocati negli strati superficiali della pelle, il recettore di Merkel e il recettore di Meissner, e due nel tessuto sottocutaneo, i recettori di Pacini e i recettori di Ruffini. Una seconda distinzione fra questi quattro tipi di MR si basa sulle proprietà della loro risposta a stimoli mantenuti costanti nel tempo: gli MR di Meissner e di Pacini sono a rapido adattamento, cessando rapidamente di rispondere a uno stimolo costante, gli MR di Merkel e di Ruffini sono invece a lento adattamento. Lo stimolo meccanico che eccita gli MR tattili è una pressione esercitata sulla pelle. Tale pressione infossa la pelle e deforma meccanicamente il terminale periferico degli MR tattili, determinando l’apertura dei CICM e quindi la depolarizzazione del terminale stesso (PDR). Anche se la sensibilità alla deformazione meccanica è una caratteristica della membrana degli MR, il tipo di risposta che viene data alla deformazione, e quindi la funzione fisiologica che essi svolgono, dipende dalla struttura specializzata (capsula) con cui tale terminale è associato, e che differisce fra i diversi MR tattili. Per es., la caratteristica dei corpuscoli di Pacini di rispondere a stimoli vibratori rapidi deriva dal loro essere MR a rapido adattamento, che a sua volta dipende dalla presenza della capsula che riveste il terminale. Il contributo di tali strutture specializzate alla m. tattile è ancora controverso così come ancora controversa è la natura dei CICM coinvolti nella m. tattile. A tutt’oggi (2010), solo uno studio ha mostrato una relazione fra la delezione di geni codificanti le proteine associate con specifici CICM, la perdita di meccanosensibilità di MR tattili sensibili al tocco lieve e la mancanza di preferenza comportamentale per superfici ruvide verso superfici lisce. Nicoletta Berardi