
immunità
immunità
Acquisizione da parte dell’organismo di capacità difensive specifiche dopo un’infezione, al fine di preservare l’individualità e l’integrità nei confronti di contaminazioni provenienti dall’ambiente esterno. L’i. è legata all’azione del sistema immunitario (➔), che si è evoluto per rispondere a due fondamentali esigenze: da un lato la necessità di reagire velocemente (nel giro di pochi minuti) contro particolari molecole presenti nei microrganismi (i. aspecifica), dall’altro il bisogno di sviluppare lentamente (nel giro di giorni o settimane) una risposta immunitaria diretta esattamente (i. specifica) contro quello specifico patogeno. Lo studio dell’i. è compito specifico dell’immunologia. Qualsiasi alterazione dei meccanismi fisiologici dell’i., nelle particolari manifestazioni patogenetiche, si chiama immunopatia. Lo studio delle diverse forme cliniche riguarda una branca specifica della medicina, che si chiama immunopatologia.
Immunità innata o aspecifica
La prima risposta all’infezione viene effettuata dal sistema immunitario naturale, con meccanismi di difesa aspecifici, che agiscono verso qualsiasi corpo estraneo, indipendentemente da un precedente contatto. Si possono distinguere meccanismi di i. cellulare, che si esplica soprattutto mediante la fagocitosi di una sostanza estranea da parte dei macrofagi, e meccanismi di i. umorale, che si attua attraverso l’intervento di numerosi fattori (sistema del complemento, lisozima, interferone, ecc.), i quali provocano la distruzione, diretta o indiretta, degli agenti estranei.
Immunità acquisita o specifica
Un secondo effetto, di fondamentale importanza, generato dal sistema immunitario consiste nell’induzione di una memoria immunitaria (➔) specifica. In questo caso il sistema immunitario è capace di generare popolazioni di cellule (linfociti B e T) specifiche per l’antigene, che possono sopravvivere per lunghissimi periodi di tempo. Anche i meccanismi di i. specifica sono distinguibili a livello umorale e cellulare. L’i. umorale è rappresentata dalla comparsa, nel siero degli individui immunizzati, di sostanze proteiche (anticorpi o immunoglobuline) che agiscono specificamente contro le sostanze estranee (antigeni); l’i. cellulare è invece dovuta all’azione dei linfociti, attivati e capaci di reagire specificamente con l’antigene.
Negli esseri viventi, dal più piccolo al più grande, diversi sono i meccanismi che nel processo evolutivo sono stati approntati per difendere l’integrità dell’individuo dagli aggressori. Si riscontrano meccanismi aspecifici di difesa con enzimi che bloccano la replicazione del DNA di batteriofagi nei batteri, di produzione di polipeptidi antibatterici e antifungini nei vegetali, e di riconoscimento di batteri e dei loro prodotti negli Invertebrati. Molti meccanismi messi in essere precocemente nella filogenesi si sono mantenuti nell’evoluzione; ad essi si sono aggiunti meccanismi sempre più complessi caratterizzati, innanzi tutto, da specificità nella risposta e successivamente di memoria della stessa.
Immunità innata e specifica
Nei Vertebrati tutti i meccanismi di difesa iniziale, con caratteristiche di aspecificità e ripetitività di risposta, indipendentemente se l’infezione avvenga o no per la prima volta, vengono convenzionalmente inclusi nell’immunità innata (o naturale). Invece, i meccanismi più complessi messi in essere dal sistema immunitario proprio dei Vertebrati, come la selezione clonale, la produzione di risposte immunitarie umorali e cellulari sempre più specifiche e il fenomeno della memoria immunologica, sono raggruppati nella così detta immunità specifica (anche detta acquisita o adattativa).
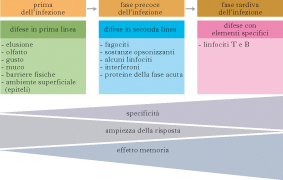
Livelli di difesa nei Mammiferi
A monte dei meccanismi dell’immunità innata nei Mammiferi si trovano le difese di barriera, con le strutture epiteliali costituite da strati di cellule vicine e tra loro strettamente collegate, che condizionano con le loro secrezioni un ambiente acido sfavorevole alla proliferazione batterica (cute, stomaco), che impediscono il contatto dei germi con la superficie e ne favoriscono l’allontanamento (secrezioni mucose, movimenti ciliari, peristalsi intestinale), o producono sostanze microbicide (lisozima, difensine). Una volta che batteri o virus riescono a penetrare le barriere, entra in gioco l’immunità innata, caratterizzata da rapidità d’azione (da alcune decine di minuti a poche ore) e dagli efficienti, ma aspecifici, meccanismi di opsonizzazione e fagocitosi. Successivamente, e grazie ai fenomeni di attivazione indotti dall’immunità naturale, si innesca, dove necessario, la risposta specifica, che impiega fino a tre giorni o più per mostrare la sua efficacia.
Risposta immunitaria innata
La risposta immunitaria innata (primaria), che avviene tramite la produzione di chemochine è la principale responsabile della formazione e della persistenza del focolaio infiammatorio, caratterizzato da arrossamento (dovuto all’iperemia), formazione di edema e infiltrazione di leucociti, e dolore per liberazione locale di sostanze della flogosi. I principali attori della risposta immunitaria innata sono le cellule fagocitarie: polimorfonucleati (PMN) e macrofagi. Entrambi sono dotati di diversi recettori di membrana capaci di riconoscere strutture molecolari tipiche dei batteri e dei virus, sia direttamente che tramite le opsonine (complemento, proteina C reattiva o PCR, ecc.). Riconosciuta la sostanza estranea, i fagociti la internalizzano in fagosomi e la digeriscono. Nei PMN la digestione porta a molecole semplici. Nei macrofagi la fagocitosi ha due conseguenze: in parte attiva il macrofago a produrre diversi tipi di chemochine (che amplificano la risposta flogistica locale) e di linfochine (che richiamano linfociti T nel focolaio); dall’altra macrofagi e cellule dendritiche assumono le funzioni di APC (cellula presentante l’antigene): frammenti antigenici delle sostanze fagocitate e digerite vengono espressi sulla membrana legate alle molecole MHC e, contemporaneamente, le membrane esprimono più recettori per i linfociti T. Altre cellule protagoniste dell’immunità innata sono i linfociti natural killer (NK), che riconoscono le cellule infettate da virus, indipendentemente dal virus infettante, determinandone la necrosi per apoptosi.
Risposta immunitaria specifica
L’attivazione dell’immunità innata è fondamentale perché si inneschino i meccanismi di quella specifica (secondaria). Infatti, l’introduzione di antigeni proteici puri porta a non reattività specifica, se non in molti casi ad anergia. Per questo, nel caso di molte vaccinazioni con antigeni proteici puri, è necessario creare un minimo di infiammazione utilizzando sostanze chiamate adiuvanti. L’immunità aspecifica, in queste condizioni, innesca la risposta immunitaria specifica, o per meglio dire selettiva. Le cellule APC migrano dal focolaio infiammatorio al linfonodo dove possono venire in contatto con i linfociti T ricircolanti. Il contatto tra APC, che presenta l’antigene riconoscibile, e il T linfocito che sa riconoscerlo in maniera specifica, innesca la risposta immune specifica. I linfociti T si attivano, producono interleuchine, si moltiplicano e attivano i linfociti B, che producono anticorpi specifici per quell’antigene. La risposta immune inizialmente coinvolge molti cloni linfocitari reattivi contro l’antigene presentato, ma poi si raffina sempre più fino a coinvolgere solo i cloni T-CD4 helper, T-CD8 citotossici e i linfociti B più selettivi, e quindi più efficaci. Gli anticorpi coniugati con antigeni solubili formano immunocomplessi che sono riconosciuti dai recettori dei fagociti. Gli anticorpi sulla superficie di cellule infettate da virus permettono l’azione litica delle cellule NK. Quindi l’immunità specifica si avvale del sistema immunitario innata per eliminare virus e batteri. Spento il focolaio infiammatorio, quando il carico antigenico si riduce, perché la risposta immune ha avuto ragione dell’aggressore, nei linfonodi permangono a lungo le APC che presentano l’antigene, come continuo stimolo per i linfociti T ricircolanti. Linfociti T e B dei cloni coinvolti nella risposta immunitaria si differenziano in cellule memoria e vanno incontro a una lunga vita, pronti a essere rapidamente attivati alla ripresentarsi dell’antigene specifico.