
Genetica del comportamento
Genetica del comportamento
La convinzione che i caratteri comportamentali si trasmettano da una generazione alla successiva è molto antica. Per secoli gli uomini hanno sfruttato questa conoscenza nell'allevamento degli animali domestici e ipotizzato che ciò spiegasse, sotto molti aspetti, anche il nostro comportamento. Più recentemente, con l'avvento della genetica, la questione dell'ereditarietà del comportamento si è trasformata nella ricerca di quali geni influenzino un dato comportamento e di come ciò avvenga. Per quanto riguarda l'uomo, l'influenza genetica sul comportamento è un problema ancora fortemente controverso e si conosce ben poco dei meccanismi che la regolano. In particolare, non si sa ancora come distinguere l'influenza dei geni da quella dell'ambiente. Per quanto invece riguarda gli animali da laboratorio, lo studio dei geni che influenzano il comportamento si focalizza su quegli aspetti dello stesso che si possono manipolare e studiare geneticamente, in condizioni in cui sono minimizzate le differenze ambientali tra gli individui.
Lo studio delle influenze genetiche sul comportamento è stato affrontato in due modi diversi: inattivando l'espressione di particolari geni e studiando le variazioni naturali del comportamento. La prima strategia utilizza le mutazioni per stabilire il ruolo di un gene nel comportamento e i suoi meccanismi di funzionamento, mentre la seconda cerca di mappare e identificare i geni responsabili di variazioni naturali nel comportamento degli animali che vengono tenuti in laboratorio. Le principali informazioni ottenute dagli studi condotti su animali in laboratorio hanno rivelato che la relazione tra geni e comportamento è complessa: più geni contribuiscono a un singolo aspetto del comportamento e ogni gene partecipa a molteplici aspetti di esso. Il comportamento, a sua volta, è una proprietà emergente dall'interazione di molti sistemi all'interno dell'organismo: i geni codificano la sintesi delle proteine, le proteine costruiscono i costituenti delle cellule, le cellule formano il sistema nervoso e altri organi, il sistema nervoso, in ultimo, dà origine al comportamento. I geni possono così influenzare il comportamento partecipando a tutti i maggiori processi biologici quali lo sviluppo, la fisiologia e la biochimica dei segnali intracellulari. Qui si esaminerà, come esempio di funzione di tali geni, il moscerino della frutta Drosophila melanogaster, organismo di cui si conosce ormai molto, per poi discutere in dettaglio le implicazioni relative alla comprensione dell'azione genica sul comportamento.
Corteggiamento e accoppiamento
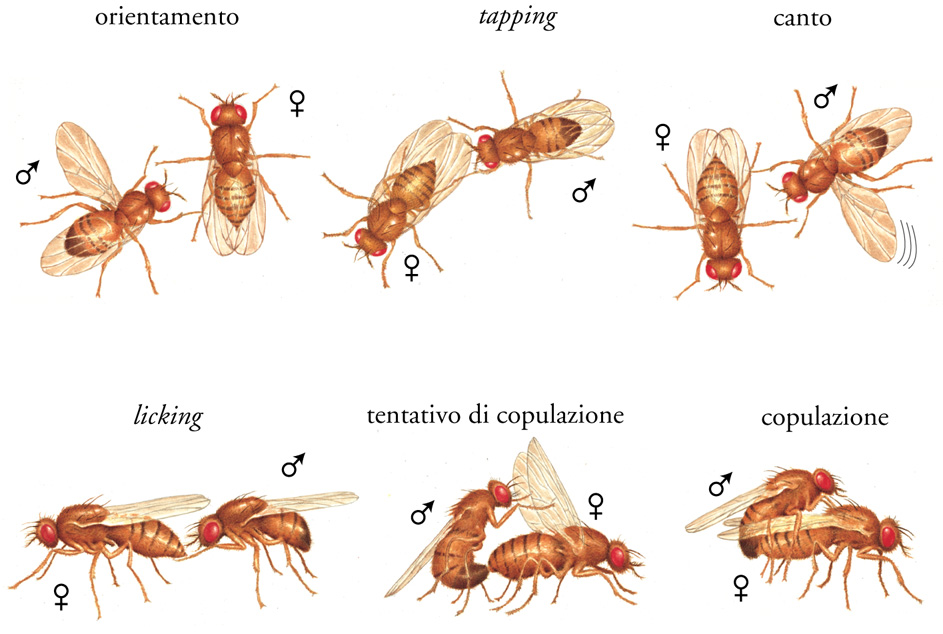
Per un organismo, il comportamento riproduttivo ha una grande importanza evolutiva e varia considerevolmente tra le specie. Il corteggiamento e l'accoppiamento in Drosophila sono comportamenti geneticamente programmati, nel senso che il maschio e la femmina non necessitano di esperienze precedenti: il comportamento riproduttivo consiste in una sequenza di azioni stereotipate il cui normale adempimento richiede uno scambio di segnali olfattivi, visivi e uditivi ben definiti tra maschi e femmine. Ciascun segnale, a sua volta, aumenta il livello di interesse dell'altro partecipante e provoca un comportamento guidato da regioni del sistema nervoso centrale specifiche a seconda del sesso. I maschi, durante il corteggiamento, sono molto più attivi delle femmine (fig. 2), poiché emettono una complessa sequenza di movimenti e posture specie-specifiche, che fungono da segnale di riconoscimento tra individui di sesso diverso ma della stessa specie. Al contrario, le femmine restano ferme per tutta la durata della sequenza. Questo comportamento di corteggiamento è controllato da fattori nervosi e genetici insieme.
Una parte dell'apparato neuronale si forma, durante lo sviluppo, in modo differente nel maschio e nella femmina. Tali differenze sono sotto il controllo dei geni che regolano la determinazione del sesso. In Drosophila, a differenza dei Vertebrati, la determinazione del sesso non è controllata dagli ormoni (quali testosterone, estrogeni, ecc.), ma è dettata da quanti cromosomi X possiede la cellula rispetto agli autosomi. Il fatto che i geni che determinano il sesso regolino il comportamento sessuale non emerge soltanto dalla differenza di comportamento del maschio rispetto alla femmina, ma, soprattutto nell'ambito di esperimenti in cui in un moscerino parte del cervello si sviluppa come maschio e il resto come femmina (in quello che è denominato mosaico genetico), si è visto che il comportamento dipende da quali porzioni del cervello sono maschili e quali femminili. In alcuni casi, il moscerino mostrerà comportamenti caratteristici di entrambi i sessi; in altri, i moscerini maschi proveranno a corteggiare senza distinzione sia maschi che femmine. In particolare, studi condotti sul gene fruitless hanno permesso di stabilire una connessione tra comportamento nel corteggiamento e sviluppo sessuale. Mutazioni di questo gene influenzano soltanto il comportamento maschile inibendo il corteggiamento; mutazioni drastiche lo bloccano del tutto, mentre mutazioni non drastiche lo frenano nelle fasi finali, cosicché i mutanti maschi non arrivano mai alla copulazione. Essi, inoltre, corteggiano insistentemente altri maschi. Coerentemente con questa idea, il prodotto del gene fruitless si trova soltanto in porzioni ristrette del cervello del maschio, molte delle quali corrispondono a siti precedentemente mappati nei mosaici genetici.
Esistono peraltro geni che controllano il comportamento di corteggiamento in maniera più sottile. Quando un moscerino maschio presenta una mutazione che influenza i ritmi biologici, il corteggiamento, all'osservazione, è del tutto normale, ma a una registrazione amplificata il canto di corteggiamento risulta anomalo. Il canto, che viene prodotto dalle vibrazioni dell'ala del maschio, è costituito da una serie di pulsazioni con una frequenza di circa 30 al secondo. Il ritmo è dato da una leggera, ma regolare, variazione nell'intervallo tra le pulsazioni. Vale a dire che il maschio aumenta gradualmente e successivamente diminuisce questo intervallo per produrre un canto lievemente oscillante, con un periodo di circa 60 secondi. Sebbene non sia assolutamente l'unico gene coinvolto in questo fenomeno, sembra che il gene period rappresenti l'elemento centrale del meccanismo di misura del tempo dell'orologio interno e, in tal senso, svolge un ruolo molto specifico nella regolazione del canto. D'altra parte, i ritmi influenzano molti aspetti differenti del comportamento di un moscerino, cosicché le mutazioni nel gene period hanno effetti tanto vasti quanto il numero dei processi ritmici. Ciò fornisce una prospettiva diversa all'idea secondo la quale un tratto comportamentale è controllato dall'azione di molti geni differenti. Ciascun gene può influenzare molteplici comportamenti e ciascun comportamento a sua volta può essere il prodotto dell'interazione di molti geni.
Le femmine rispondono al canto del maschio mostrandosi più ricettive alle proposte di corteggiamento. Se a una femmina solitaria viene fatta ascoltare una registrazione del canto e subito dopo essa viene messa in presenza di un maschio, si accoppierà più rapidamente rispetto a una femmina che non ha ascoltato il canto nei minuti precedenti. I maschi che posseggono una mutazione nel gene period (per0) producono un canto in cui manca il normale ritmo. Se si riproduce un canto aritmico utilizzando il computer e lo si fa ascoltare alla femmina come prestimolo, si noterà che esso ha meno effetto nell'accelerare l'accoppiamento. Un'altra mutazione nel gene period (pers) causa un ritmo con un periodo più corto, pari a 30 secondi invece di 55. Anche questo canto è meno efficace nello stimolare la femmina all'accoppiamento. Infine, va detto che le influenze genetiche fanno sì che i moscerini mostrino preferenze, in certi momenti della giornata, per alcuni processi biologici, come per esempio lo stato generale di attività, la schiusa delle pupe dei moscerini dopo la metamorfosi e la formazione della membrana vitellina attorno agli oociti delle femmine. Normalmente essi regolano il loro orologio interno in modo analogo all'uomo, cioè basandosi sull'alternanza del giorno e della notte ma, anche se sottoposti a buio costante, sono in grado di mantenere il ritmo di 24 ore sotto l'influenza del loro orologio interno.
Al contrario, quando i moscerini presentano mutazioni nel gene period, sono in grado di avere un normale ritmo ciclico se esposti a un ciclo di buio e luce di 24 ore, ma non riescono a mantenerlo quando sono tenuti in condizioni di buio costante. Il mutante per0, invece, non possiede un ciclo distinguibile, il mutante pers ha un ciclo di 19 ore e un terzo mutante perl ha un ciclo di 29 ore. Gli effetti delle mutazioni nel gene period sul ritmo del canto di corteggiamento ricalcano i difetti osservati nei ritmi circadiani. Ciò è tanto più evidente se si considera che il ritmo del canto impiega soltanto un minuto circa per compiere un ciclo. Sembra che il ruolo molecolare del prodotto del gene period sia quello di controllo di altri geni. Tale proteina è localizzata nel nucleo di molte cellule del sistema nervoso del moscerino e, in quelle che presumibilmente governano il ritmo circadiano, il livello del prodotto genico oscilla con un ciclo di 24 ore.
Variazioni genetiche naturali del comportamento
In che misura gli esempi citati finora, riguardanti geni che influenzano il comportamento, sono applicabili alla diversità di comportamenti presente in natura? La misura della variazione esistente in natura si può osservare nelle differenze tra animali domestici e animali selvatici. Il processo di addomesticazione consiste nel selezionare, generazione dopo generazione con incroci controllati, particolari caratteri, molti dei quali comportamentali. In assenza di variazione genetica non c'è possibilità di selezione. Così, l'esistenza di specie domestiche e la presenza di differenze tra le razze di una stessa specie indicano che c'è una naturale e significativa variazione spontanea nel comportamento.
Un buon metodo per studiare il meccanismo con cui i geni regolano questi caratteri è quello di ricorrere all'incrocio di razze che sono state selezionate per differenze comportamentali. Due razze differenti vengono incrociate, la progenie (F1) viene osservata per le caratteristiche comportamentali e, successivamente, i singoli individui vengono incrociati l'uno con l'altro. La progenie risultante (F2) rivelerà se la differenza comportamentale è dovuta a un singolo gene, a pochi o a molti geni a seconda di quanti differenti fenotipi presenta. In questo modo sono stati effettuati incroci tra cani per determinare la base genetica di caratteri come il puntamento, la paura del colpo di fucile e l'adattamento al guinzaglio. In tutti i casi, i caratteri analizzati presentano un tipo complesso di ereditarietà, ossia sono ereditabili, e tuttavia non sono mai controllati da uno o pochi geni, essendo chiaramente poligenici. Lo stesso tipo di risultati è stato ottenuto in esperimenti sull'ereditarietà di differenze comportamentali nell'ambito di una razza. Sono state ottenute informazioni su ratti, api e moscerini, selezionati per la loro abilità ad apprendere più o meno rapidamente. Dopo numerosi anni di studio si può concludere che tali caratteri hanno alcune componenti ereditarie prevalenti, ma sicuramente non sono controllati da uno o da pochi geni.
Una delle poche eccezioni a questa tendenza è stata ottenuta mediante un esperimento analogo realizzato sul moscone, Phormia regina, operando una selezione per differenze nello stato eccitatorio centrale (CES). Il moscone protrude la proboscide quando i suoi peli sensoriali sono stimolati con una soluzione di saccarosio, ma non con la sola acqua. Durante la stimolazione di un solo pelo con il saccarosio, il moscone estrofletterà la proboscide in risposta allo stimolo con l'acqua di un pelo diverso. L'accoppiamento selettivo fra individui che mostrano livelli alti oppure bassi (o assenti) di risposta a questo tipo di stimolazione porta solitamente alla generazione di due linee pure caratterizzate da risposte deboli o forti e dimostra che tale carattere è prevalentemente controllato da un singolo locus principale.
Una soluzione alternativa per identificare variazioni comportamentali naturali è quella di cercarle in un organismo che sia già a un livello sofisticato per quanto riguarda l'analisi di genetica molecolare, come nel caso del moscerino della frutta. Le larve del moscerino della frutta presentano un polimorfismo nel comportamento locomotorio associato alla ricerca e all'assunzione di cibo (foraging). Questo polimorfismo comportamentale che si osserva in natura si ritrova parimenti sia nei ceppi di Drosophila cresciuti in laboratorio sia in quelli selvatici. Il comportamento di foraging viene valutato misurando la lunghezza della traccia del percorso fatto dalle larve durante l'esplorazione per il cibo su una piastra di agar coperta di lievito.

Le larve si dividono in due categorie: erranti e sedentarie (rover e sitter). Le larve erranti, in un determinato intervallo di tempo, lasciano una traccia più lunga rispetto alle larve sedentarie (fig. 4). Quando si analizzano caratteri quantitativi non è rara una distribuzione bimodale di questo tipo, anche se non c'è stata alcuna selezione per quanto riguarda i fenotipi. Come per la maggior parte delle varianti comportamentali isolate da popolazioni naturali, i due tipi presentano distribuzioni quantitative sovrapposte. Tuttavia, quando sono stati prodotti ceppi omozigoti per il fenotipo errante o sedentario, essi presentavano una sovrapposizione minima e si è dimostrato che la differenza tra erranti e sedentarie è causata dall'azione di un singolo gene, denominato successivamente foraging, in cui il fenotipo errante è geneticamente dominante su quello sedentario. In seguito il gene foraging è stato mappato, ne è stata determinata la localizzazione cromosomica ed è stato sottoposto a un tipo di mutagenesi tradizionale che produce mutazioni più drastiche di quelle che si trovano in natura (come fruitless).
Questi studi hanno così dimostrato che il fenotipo dominante rover poteva essere mutato nel fenotipo recessivo sitter, aprendo così una serie di interrogativi che normalmente vengono sollevati nel caso dell'analisi di mutazioni indotte più severe, ossia stabilire se il gene è essenziale. Alcune delle nuove mutazioni del gene foraging erano letali, indicando che quella naturale era una forma lieve di mutazione. Successivamente sono state identificate numerose altre mutazioni vitali, la cui analisi ha mostrato un effetto sulla locomozione dell'adulto. Procedendo in questo modo, il gene foraging è stato trovato e identificato come una blanda mutazione comportamentale naturale; attraverso la mutagenesi è stato possibile creare alleli più drasticamente letali e questi, a loro volta, hanno reso possibile l'identificazione e il clonaggio del frammento di DNA che contiene il gene. In base alla sequenza di DNA, si è visto che il foraging codifica la sintesi di una proteina-chinasi cGMP-dipendente. Come per altre chinasi note in letteratura, anche la chinasi cGMP-dipendente è implicata in modificazioni di vario tipo dell'attività neuronale. La differenza errante/sedentario è precisamente di tipo enzimatico: il fenotipo errante presenta un'attività enzimatica maggiore rispetto al fenotipo sedentario. Non si conosce tuttavia ancora in quale parte del cervello si manifesti tale differenza.
Ritmi biologici naturali
Non esiste una netta distinzione tra i geni identificati in seguito a mutazioni indotte in laboratorio e quelli identificati tramite i polimorfismi naturali. Così, una volta che il gene period è stato identificato e clonato in laboratorio, sono state cercate variazioni nella sequenza del DNA in natura e ne sono state trovate sia tra specie diverse sia all'interno della stessa specie D. melanogaster. Trovata una variazione nella sequenza nucleotidica, si può determinare se questa differenza causi una modificazione nel comportamento. Quando la sequenza è stata confrontata con la specie strettamente correlata D. simulans, sono state riscontrate alcune differenze. Queste erano per lo più contenute in una regione localizzata a circa metà del gene, distinguibile in quanto codifica una lunga ripetizione dei due amminoacidi treonina e glicina. Tale mutazione di 'origine spontanea' è sembrata particolarmente interessante perché D. melanogaster e D. simulans hanno lo stesso ritmo circadiano di 24 ore, ma differiscono per il ritmo del canto durante il corteggiamento del maschio: maschi della specie D. simulans presentano infatti un ritmo oscillante ogni 40 secondi, mentre maschi di D. melanogaster ogni 60 secondi.
Le differenze tra D. simulans e D. melanogaster suggeriscono che lo stesso gene mantiene costante la temporalità di una funzione (ritmo circadiano), mentre permette all'altra funzione (ritmo del canto) di variare. Con tecniche di genetica molecolare è possibile individuare le regioni del gene responsabili della variazione del ritmo del canto. Per mezzo dell'ingegneria genetica è stato prodotto un gene period misto, una nuova versione che codificava la sintesi di una proteina in cui la regione centrale del gene period di D. simulans sostituiva la regione corrispondente del gene period di D. melanogaster. La porzione centrale trasferita conteneva le ripetizioni treonina-glicina (T-G) e le sequenze adiacenti. Questo gene 'chimerico' è stato successivamente introdotto in un ceppo di D. melanogaster in cui mancava il gene period; in tal modo questo moscerino aveva come unica fonte funzionale il gene period funzionante. I moscerini risultanti presentavano il solito ritmo circadiano di 24 ore comune a entrambe le specie. Inoltre, essi producevano un canto con l'autentico ritmo di D. simulans. Il gene reciproco in cui la regione centrale di D. melanogaster sostituisce quella corrispondente nel gene di D. simulans ha dato il risultato opposto: un canto di tipo melanogaster, ma nessun cambiamento del ciclo di 24 ore.
Quando sono stati raccolti campioni di DNA da ceppi selvatici di moscerini catturati in una vasta area geografica che si estende da Bristol, in Inghilterra, a Casablanca, in Marocco, sono state trovate differenze nella stessa regione centrale del gene che include le ripetizioni treonina-glicina e le sequenze adiacenti. Di questa sequenza si sono trovate tre versioni principali (alleli), e in ciascuna area geografica esse erano rappresentate tutte e tre nella popolazione di moscerini. L'allele predominante, tuttavia, non era lo stesso per ciascuna zona geografica, ma presentava una transizione graduale da nord a sud. Il fatto che la prevalenza allelica si correli con differenze climatiche induce a pensare che ciascun allele conferisca un vantaggio selettivo leggermente differente. Infatti, successive analisi comportamentali di queste varie popolazioni naturali hanno rivelato che le popolazioni con più lunghe ripetizioni T-G erano più costanti nel mantenere la periodicità del ritmo circadiano, in seguito a fluttuazioni della temperatura. Ciò è particolarmente importante negli animali che, come i moscerini, non regolano autonomamente la loro temperatura corporea. Si è infatti osservato che la regione con ripetizioni T-G più lunghe predominava nelle zone climatiche più a nord. I differenti alleli mostrano anche piccole differenze nel ritmo del canto, ma non è chiaro quanto queste variazioni siano rilevanti. Sarebbe veramente suggestivo se si trovasse che le femmine manifestano una preferenza per il canto di maschi con l'allele period più appropriato alla loro zona climatica.
L'idea che in natura un gene possa variare in modo così circostanziato da aggiustarsi in accordo con le differenti nicchie ecologiche comporta enormi implicazioni evolutive. Più in generale, il gene period dimostra che studiando l'ecologia comportamentale possono essere scoperti aspetti interessanti della struttura e funzione genica. Questo significa sia che possiamo imparare molto su come i geni influenzano il comportamento, ricercando versioni naturali dei geni identificati in laboratorio, sia che questa influenza non è di tipo specialistico. In sintesi, per i geni che durante lo sviluppo hanno un ruolo talmente importante da influenzare il comportamento, come abbiamo visto per quelli che determinano il sesso, la mancanza di specificità deriva dalla promiscuità dell'azione genica durante lo sviluppo. La maggior parte dei geni implicati nello sviluppo adempie a molte funzioni distribuite in varie parti dell'animale e in vari stadi dello sviluppo. I geni per la determinazione del sesso influenzano un'ampia gamma di caratteri sessuali dimorfici. Anche gli effetti più ristretti di fruitless si estendono oltre il comportamento del corteggiamento e il gene omb è coinvolto nello sviluppo di ampie porzioni del cervello, non soltanto in quello delle cellule del movimento nei lobi ottici. Molti dei geni che interessano il comportamento codificano enzimi coinvolti nella trasduzione del segnale, quali la proteina-chinasi cGMP-dipendente dal gene foraging.
Queste osservazioni hanno importanti implicazioni per ciò che pensiamo riguardo all'influenza dei geni sul comportamento. Esse mostrano infatti che non ci può essere una semplice corrispondenza tra un particolare gene e un dato comportamento, ossia i geni non controllano il comportamento direttamente, anche in quei casi in cui un certo gene è assolutamente richiesto. È invece necessario cominciare a pensare nei termini di una rete di geni che forma una matrice interconnessa in grado di produrre i circuiti e la capacità di trasmissione di segnali del sistema nervoso che, a sua volta, dà origine al comportamento. Tale modo di pensare rappresenta un allontanamento dal paradigma che ha guidato la biologia molecolare dagli anni Sessanta del XX sec. fino a oggi: la scomposizione di un processo nelle sue varie componenti e lo studio di queste singole parti fino a ottenerne una descrizione completa. Il fallimento di questo paradigma nella risoluzione dei problemi a livello di sistemi e di reti sta diventando sempre più evidente negli studi sulla trasduzione del segnale nelle cellule e nel funzionamento delle reti neurali, così come nel comportamento.
È possibile una genetica del comportamento umano?
Per quanto riguarda l'uomo, la questione di quali siano le componenti ereditarie del comportamento è stata molto controversa, a causa delle sue pesanti implicazioni politiche. All'inizio del XX sec., quando la genetica come scienza era ancora agli albori, c'è stata una corsa nell'attribuire tutti i caratteri umani, inclusi i tipi complessi di personalità e addirittura lo status socioeconomico, agli effetti di singoli geni ereditati secondo il modello mendeliano. Questa posizione estremista di assoluto determinismo alimentò il movimento eugenetico volto al miglioramento genetico della specie umana. A partire dalla metà del secolo si verificò un netto cambiamento come risultato di diversi fattori: le puntuali critiche dei nuovi risultati ottenuti da alcuni genetisti, la nascita delle moderne scienze sociali con il loro riconoscimento delle influenze culturali e l'avversione per l'attuazione dell'ideologia eugenetica nella Germania nazista.
Il punto critico di questa controversia è il ruolo dell'ambiente. I fautori di uno stretto determinismo genetico negano che l'ambiente influenzi in modo significativo il comportamento, mentre i loro oppositori asseriscono che l'ambiente è tutto ciò che conta. La chiave per riconoscere e, in ultimo, identificare le componenti genetiche del comportamento è controllare le differenze ambientali. Questo approccio ha consentito di gettare le basi per gli attuali studi sul ruolo dei geni nel comportamento degli animali. Da questo punto di vista, gli studi sull'uomo appaiono irrealizzabili, poiché non è possibile (o accettabile) controllare l'ambiente di un singolo individuo per fini sperimentali, a eccezione di alcuni casi, molto limitati, di soggetti volontari. C'è il rischio che i risultati degli esperimenti genetici qui esaminati possano servire per diffondere la nozione che il comportamento, incluso quello dell'uomo, è geneticamente determinato. È invece probabile che la predisposizione genetica abbia influenza soltanto su una piccola parte del nostro comportamento e che un'altra limitata frazione risulti determinata esclusivamente da fattori ambientali relativamente semplici. È però molto importante ricordare che la grande maggioranza dei fattori deterministici risiede verosimilmente in una moltitudine di interazioni tra fattori genetici.
Tenendo presenti queste osservazioni, è possibile valutare criticamente quanto gli studi genetici condotti sui moscerini e sui topi siano attinenti all'uomo. La conservazione dei geni tra le specie, comprese specie così lontane come il moscerino e l'uomo, è stata una delle scoperte più incredibili della biologia molecolare contemporanea. Per esempio, il gene che codifica la proteina precursore dell'amiloide, principale componente delle placche associate alla malattia di Alzheimer, si trova anche in Drosophila e i geni sono talmente simili che i mutanti di Drosophila possono essere completamente recuperati per mezzo del gene umano. Tuttavia le sequenze del DNA e delle proteine non sono le sole caratteristiche a essere conservate. C'è anche da tenere presente una conservazione significativa della funzione complessiva di un gene, come mostrato nel ruolo giocato dal gene period anche nei ritmi circadiani umani. Da simili dati si sarebbe tentati di estrapolare le conclusioni direttamente da una specie all'altra; tali generalizzazioni acritiche non sono tuttavia giustificate. Il principale valore di queste ricerche è di tipo euristico e serve da guida per sviluppare nuove teorie, alle quali dovrà sempre seguire la necessaria verifica sperimentale. Per esempio, non c'è alcuna indicazione che i meccanismi per la determinazione del sesso che operano nel moscerino possano essere rilevanti anche per noi. Questo dovrebbe indurre alla prudenza, per esempio, chi volesse tout court estendere all'uomo le conoscenze in merito agli effetti dei geni sull'orientamento sessuale del moscerino. Le conoscenze acquisite con gli studi che hanno preso in esame i geni e il comportamento degli animali di laboratorio saranno certamente utili quando i dati sulle influenze genetiche sul comportamento umano saranno più consistenti. Gli studi di mappatura del genoma umano hanno condotto all'isolamento di nuovi geni coinvolti nel comportamento con una velocità stupefacente. È diventato così sempre più evidente che i geni permettono la vita ma non la determinano. Tuttavia, anche se la genetica ci informa sui meccanismi della vita, essa non cancella la nostra essenza profonda di essere umani e cioè la nostra specifica capacità di riflettere sulle cose, di valutarle e di compiere scelte.
Bibliografia
Colot 1988: Colot, Hildur V. - Hall, Jeffrey C. - Rosbash, Michael, Interspecific comparison of the period gene of Drosophila reveals large blocks of non-conserved coding DNA, "European molecular biology organization journal", 7, 1988, pp. 3929-3937.
Greenacre 1993: Greenacre, Mary L. e altri, Female song preference and the period gene in Drosophila, "Behavior genetics", 23, 1993, pp. 85-90.
Greenspan, Tully 1994: Greenspan, Ralph J. - Tully, Tim, How do genes set up behavior?, in: Flexibility and constraint in behavioral systems, edited by Ralph J. Greenspan and Charalambos P. Kyriacou, Chichester-New York, Wiley, 1994, pp. 65-80.
Hall 1994: Hall, Jeffrey C., The mating of a fly, "Science", 264, 1994, pp. 1702-1714.
Lee 2000: Lee, Gyunghee e altri, Spatial, temporal, and sexually dimorphic expression patterns of the fruitless gene in the Drosophila central nervous system, "Journal of neurobio_logy", 43, 2000, pp. 404-426.
Luo 1992: Luo, Laiyi - Tully, Tim - White, Kalpana, Human amyloid precursor protein ameliorates behavioral deficit of flies deleted for Appl gene, "Neuron", 9, 1992, pp. 595-605.
Osborne 1997: Osborne, Kathleen A. e altri, Natural behavior polymorphism due to a cGMP-dependent protein kinase of Drosophila, "Science", 277, 1997, pp. 834-836.
Poeck 1993: Poeck, Burkhard - Hofbauer, Alois - Pflugfelder, Gert O., Expression of the Drosophila optomotor-blind gene transcript in neuronal and glial cells of the developing nervous system, "Development", 117, 1993, pp. 1017-1029.
Sawyer 1997: Sawyer, Leigh A. e altri, Natural variation in a Drosophila clock gene and temperature compensation, "Science", 278, 1997, pp. 2117-2120.
Sokolowski 1992: Sokolowski, Marie B., Genetic analysis of behavior in the fruit fly, Drosophila melanogaster, in: Techniques for the genetic analysis of brain and behavior: focus on the mouse, edited by Daniel Goldowitz, Douglas Wahlsten and R.E. Wimer, Amsterdam-London, Elsevier, 1992, pp. 497-512.
Tully, Hirsch 1982: Tully, Tim - Hirsch, Joy, Behavior-genetic analysis of Phormia regina. II. Detection of a single, major-gene effect from behavioral variation for central excitatory state (CES) using replicate hybrid crosses, "Animal behavior", 30, 1982, pp. 1193-1202.
Wheeler 1991: Wheeler, David A. e altri, Molecular transfer of a species-specific behavior from Drosophila simulans to Drosophila melanogaster, "Science", 251, 1991, pp. 1082-1085.
Young 2000: Young, Mark W., The tick-tock of the biological clock, "Scientific American", 282, 2000, pp. 64-71.