
funzioni cerebrali superiori
funzioni cerebrali superiori
Le funzioni cerebrali superiori sono funzioni complesse quali la memoria, il linguaggio, la capacità di ragionamento, di pianificazione, di risolvere un problema, nonché la percezione e l’azione. La conoscenza dei substrati cerebrali implicati in tali processi è ancora incompleta. È però noto che il segnale informativo viene recepito dalle aree sensoriali primarie per giungere, con un processo a tappe che coinvolge le aree associative polimodali, fino alle aree motorie. Le aree associative polimodali sono ritenute il substrato anatomico delle funzioni superiori e sono particolarmente sviluppate nei primati e nell’uomo. Le funzioni cerebrali superiori più complesse emergono probabilmente dalla coordinazione delle attività delle diverse aree associative. In partic. le associazioni fra la corteccia parietale e la corteccia prefrontale sembrerebbero le principali responsabili delle funzioni superiori più vicine a quella che definiamo intelligenza. [➔ apprendimento; cervello, evoluzione del; corteccia cerebrale; intelligenza; intelligenza artificiale; limbico, sistema; linguaggio; memoria; sensazione e percezione]
Le f. c. s. utilizzano l’informazione sensoriale elaborata dalle aree sensoriali per formulare concetti complessi che possono essere comunicati, ricordati, a breve o a lungo termine, usati per creare nuove idee, associazioni e scopi, e che possono essere sviluppati in una azione. Si tratta di funzioni complesse, dette anche funzioni corticali superiori, quali memoria, linguaggio, capacità di ragionamento, di pianificazione, di risolvere un problema; anche la percezione (➔ sensazione e percezione) e l’azione possono essere considerate funzioni superiori, in considerazione della complessità del compito di riconoscere uno stimolo o di mettere in atto il piano motorio adatto a ottenere uno scopo. Per es., il poter udire e distinguere tra loro una serie di suoni quando ascoltiamo qualcuno parlare, dipende dalle funzioni uditive di base; è invece il frutto di f. c. s. la capacità di riconoscere nella sequenza di questi suoni parole e frasi di significato compiuto, e di collegarle a concetti già noti, per poi formulare una nuova idea che potremo comunicare al nostro interlocutore, e successivamente ricordare e combinare con altre idee in base alle quali agire.
Meccanismi delle funzioni cerebrali superiori
I meccanismi responsabili delle funzioni sensoriali di base, così come quelli responsabili delle funzioni motorie di base sono in genere abbastanza ben compresi. Invece, per quel che riguarda le f. c. s. e, in partic., i substrati cerebrali implicati in tali processi, le conoscenze sono ancora (2010) molto incomplete, anche se la maggior parte degli studi indica le aree corticali associative come il substrato neurale di tali funzioni. Già Carl Wernicke aveva ipotizzato che le f. c. s., e in partic. il linguaggio, fossero processi derivanti dalla interconnessione di sistemi che investono più aree funzionali cerebrali, emergenti quindi da connessioni associative che legano fra loro aree diverse. In contemporanea, John Hughlings Jackson aveva proposto che la corteccia cerebrale avesse una organizzazione gerarchica, e che alcune aree corticali non svolgessero funzioni esclusivamente sensoriali o motorie ma funzioni integrative superiori. L’ipotesi corrente è che le f. c. s. emergano dall’integrazione di informazioni che provengono da numerose strutture corticali che possono risiedere in entrambi gli emisferi e che tale integrazione venga svolta dalle aree associative. Le f. c. s. sono altamente interconnesse e interrelate, basti pensare alla inscindibile relazione tra memoria dichiarativa (memoria episodica e memoria semantica) e linguaggio, tra percezione e memoria, anche se, per motivi di semplicità di analisi, queste funzioni vengono in genere trattate separatamente.
Flusso delle informazioni
Il segnale informativo viene recepito dalle aree sensoriali primarie (per es., visiva primaria, acustica primaria, somatosensoriale primaria) che proiettano ad aree sensoriali adiacenti. La funzione di queste aree, che integrano informazioni all’interno di una singola modalità (aree associative unimodali), è quella di consentire rappresentazioni progressivamente più complesse dello stimolo sensoriale. Le aree associative unimodali proiettano ad aree associative sensoriali polimodali, che integrano informazioni da più di una modalità. Le connessioni sono a doppio senso: le aree associative unimodali e polimodali proiettano alle aree sensoriali primarie, consentendo il controllo della loro attività in modalità top-down, controllo importante per gli effetti dei processi attentivi e di apprendimento sulla percezione. Le aree associative sensoriali polimodali proiettano alle aree motorie associative polimodali, localizzate anteriormente nei lobi frontali. Tali aree pianificano una uscita comportamentale appropriata per il contesto e per gli scopi del soggetto ed elaborano i programmi per i movimenti da eseguire. Questi programmi sono inviati alle aree motorie, premotoria e motoria primaria, che inviano i segnali ai motoneuroni per l’attuazione dell’uscita motoria.

Le aree associative polimodali sono ritenute il substrato anatomico delle funzioni superiori. Emerse gradatamente durante l’evoluzione, esse sono molto sviluppate nei primati e nell’uomo. Tre aree associative polimodali sono particolarmente rilevanti:
• area associativa posteriore (area occipito-temporo-parietale), al confine tra i lobi occipitale, temporale e parietale: integra informazioni provenienti da modalità diverse ed è implicata nell’attenzione, nel linguaggio, nell’orientamento spaziale, nel riconoscimento del sé e dell’ambiente e partecipa all’organizzazione di movimenti complessi;
• area associativa anteriore (corteccia prefrontale): è associata con le funzioni esecutive del comportamento, quali la risoluzione di problemi, la pianificazione di una strategia di azione che conduce allo scopo prefisso, il monitoraggio delle prestazioni, la capacità di cambiare strategia nel momento in cui le circostanze lo richiedono, la valutazione delle conseguenze delle proprie e altrui azioni, il pensiero astratto, la memoria di lavoro;
• area associativa limbica: situata lungo le facce mediali degli emisferi cerebrali, è implicata nella formazione della memoria dichiarativa a lungo termine e nel comportamento emotivo.
Aree associative
Le funzioni delle aree associative sono state dedotte inizialmente dalla osservazione di pazienti con lesioni corticali selettive e, più recentemente, dall’utilizzo di tecniche di imaging cerebrale, che consentono di visualizzare l’attività cerebrale in soggetti impegnati in compiti tali da richiedere funzioni superiori. Inoltre, l’utilizzo di lesioni selettive e di registrazioni elettrofisiologiche da specifiche aree cerebrali nella scimmia ha dato un contributo alla conoscenza dei processi svolti in queste aree. I diversi tipi di informazione si integrano molto bene: per es., lesioni delle aree associative unimodali visive nel lobo temporale possono produrre una selettiva incapacità di riconoscere una classe di oggetti presentati visivamente, senza influenzare la capacità di riconoscerli attraverso il tatto. Esperimenti condotti nelle scimmie hanno mostrato che lesioni in aree corrispondenti provocano anch’esse danni al riconoscimento visivo degli oggetti, e che i neuroni di queste aree rispondono a stimoli complessi quali le facce o la mano, suggerendo fortemente il loro coinvolgimento in compiti di riconoscimento visivo degli oggetti e delle immagini. A partire dal 2000, molte ricerche hanno mostrato che durante lo svolgimento di compiti che coinvolgono le f. c. s. è evidente la presenza di una rete di aree associative attive. Le f. c. s. più complesse potrebbero quindi emergere dalla coordinazione delle attività delle diverse aree associative. È stato proposto che la sincronizzazione delle oscillazioni dell’attività neuronale tra queste aree sia un modo per coordinare tali reti neurali distribuite: per es., in un difficile compito di riconoscimento visivo di immagini frammentate, si osserva che quando il soggetto riesce a riconoscere l’immagine, le oscillazioni dell’attività neuronale su specifiche bande di frequenza risulta sincronizzata per tre gruppi di aree, temporali, occipitali e frontali; questo non si verifica quando il soggetto fallisce nel compito di riconoscimento. È come quando più computer lavorano in rete per elaborare lo stesso problema, raggiungendo così un potere computazionale maggiore di quello di ogni singolo computer. L’alterazione delle oscillazioni locali (nelle singole aree) o della sincronizzazione delle oscillazioni locali tra aree diverse, viene ritenuta una possibile causa di disturbi delle f. c. s., quali si possono riscontrare per es. nella schizofrenia. Le associazioni fra la corteccia parietale e la corteccia prefrontale sembrerebbero le principali responsabili delle f. c. s. più vicine al nostro concetto di intelligenza: per es., è stato proposto che l’intelligenza generale emerga dall’attività di una rete distribuita di aree associative frontoparietali che integrano funzioni di memoria verbale, visuospaziale e di lavoro con funzioni esecutive.
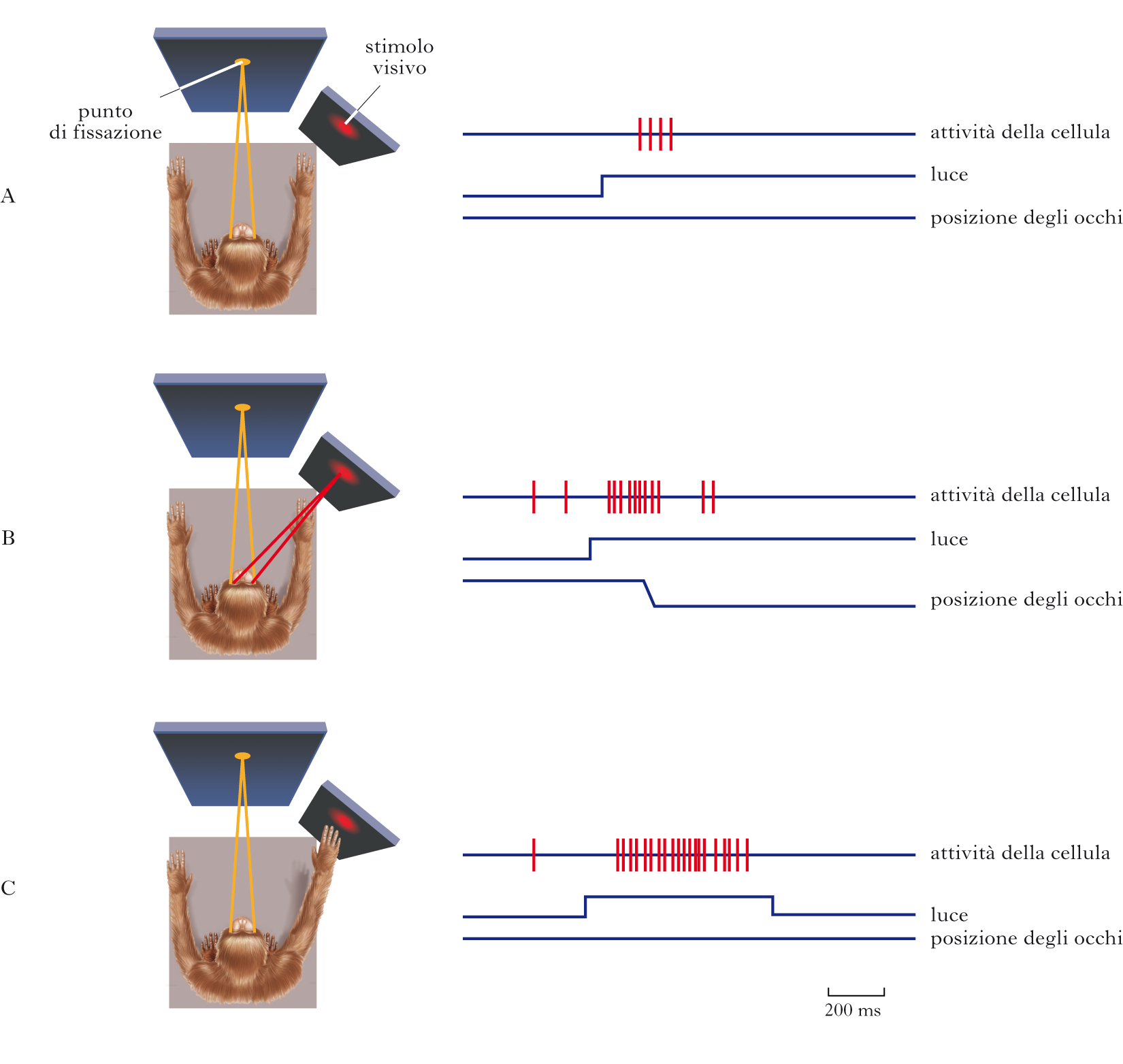
Evidenze sperimentali del ruolo delle specifiche aree associative
Corteccia parietale posteriore (CPP). Pazienti con lesioni nella CPP destra mostrano una incapacità a percepire, esplorare e agire nello spazio controlaterale alla sede della lesione cerebrale, pur evidenziando una condizione di normalità per quanto riguarda i canali sensoriali. Il paziente si comporta come se non fosse più in grado di percepire e concepire l’esistenza di un lato dello spazio e vi è anche una mancanza di percezione del proprio emisoma controlaterale; tali pazienti possono per es., non mangiare il cibo nella metà sinistra del piatto, o non lavare la metà sinistra del corpo. Quindi, l’attività delle cortecce sensoriali primarie non è sufficiente per la percezione cosciente se manca l’attività della CPP. In un famoso esperimento, Edoardo Bisiach ha mostrato l’esistenza di una eminegligenza che coinvolge anche la memoria dello spazio extrapersonale secondo un sistema di riferimento centrato sul corpo. La conclusione tratta è che la CPP sia cruciale per la consapevolezza della struttura e della dimensioni del nostro corpo e dello spazio che lo circonda e per la loro rappresentazione mentale. Studi effettuati registrando l’attività di singoli neuroni nella CPP della scimmia hanno suggerito che essa sia implicata nei meccanismi di attenzione selettiva: la risposta a uno stesso stimolo è molto maggiore quando l’animale presta a esso attenzione in quanto è il bersaglio di un futuro movimento, sia effettuato con gli occhi che con la mano. La CPP destra è stata implicata nella cognizione numerica, parte delle capacità cognitive matematiche. La CPP sembrerebbe essere la sede della ‘linea mentale dei numeri’, ossia della visualizzazione della successione dei numeri lungo una linea (numeri più piccoli a sinistra e quelli più grandi a destra). L’esistenza della linea dei numeri viene interpretata come evidenza del fatto che i numeri sono codificati spazialmente nel cervello. Il coinvolgimento dell’area associativa posteriore di sinistra nel linguaggio è legato alla presenza in tale area del giro angolare, dell’area di Wernicke (➔) e di altre aree perisilviane coinvolte nella comprensione della parola letta (visivamente o tattilmente, in Braille) o udita.
Corteccia parietale inferiore (CPI). Quest’area gioca un ruolo nella memoria di lavoro (quella particolare forma di memoria utilizzata per mantenere le informazioni nel corso dell’attuazione di un comportamento); in partic., la CPI sinistra sembra essere il substrato neurale del ‘magazzino fonologico’ per la memoria di lavoro verbale. Il corrispondente del magazzino per la memoria di lavoro non verbale (il ‘taccuino visuo-spaziale’) sembrerebbe essere situato nella CPI destra. La memoria di lavoro emergerebbe quindi dal coordinamento dell’attività di zone parietali (magazzino fonologico e taccuino visuospaziale) e zone prefrontali (esecutivo centrale).
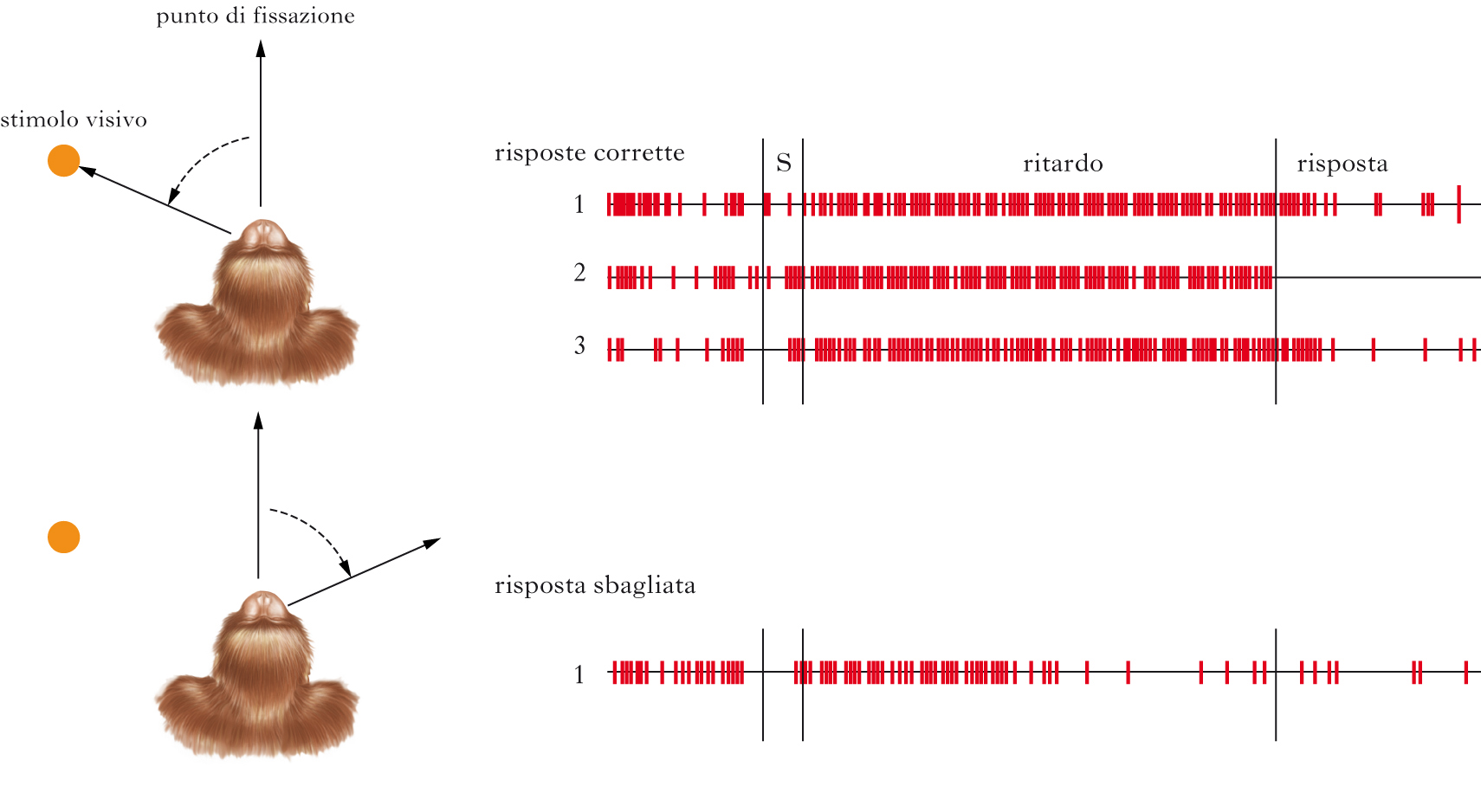
Corteccia prefrontale (CPF). È l’area che maggiormente differisce fra l’uomo e i primati non umani, suggerendo che sia responsabile delle f. c. s. che maggiormente ci differenziano da essi. La CPF è suddivisibile in tre regioni, dorsale, mediale e orbitofrontale (spesso indicata come ventromediale). Osservazioni in pazienti con lesioni a sezioni diverse della CPF, esperimenti di imaging cerebrale in soggetti normali impegnati in compiti diversi, esperimenti di lesione e di registrazione dell’attività di singoli neuroni nelle scimmie, hanno suggerito che aree diverse della CPF giocano ruoli diversi nelle f. c. s..; per es., il ruolo principale della corteccia prefrontale dorsolaterale (CPFDL) sembra essere quello di esecutivo centrale nella memoria di lavoro. Infatti, lesioni in questa area producono nell’animale e nell’uomo gravi deficit di memoria di lavoro; per es., non appena si introduce un ritardo temporale fra il mostrare all’animale in quale contenitore, fra due o più, viene nascosto il cibo e il momento in cui l’animale può selezionare il contenitore per recuperare il cibo, in modo che esso scelga sulla base di un’informazione che deve mantenere in memoria per qualche secondo (compito di scelta ritardata), la prestazione dell’animale lesionato diventa molto scadente. La CPFDL nell’uomo si attiva durante lo svolgimento di compiti che implicano la memoria di lavoro. Se si registra l’attività di singole cellule dalla CPFDL mentre una scimmia esegue un compito di scelta ritardata, si osserva che la scarica neuronale si mantiene elevata durante tutto l’intervallo temporale in cui l’animale tiene in memoria l’informazione necessaria per eseguire poi la scelta. È come se queste cellule mantenessero attiva l’informazione; in effetti, nelle prove in cui la loro attività decade, durante l’intervallo, l’animale effettua una scelta scorretta. Come accennato precedentemente, la CPF sembra reclutata per lo svolgimento di compiti di riconoscimento visivo complessi, quali il riconoscimento di oggetti visti da una prospettiva inusuale; il reclutamento delle aree prefrontali, in aggiunta a quello delle aree associative posteriori, consentirebbe processi di rotazione mentale, che permettono il riconoscimento dell’oggetto in base all’esperienza di esso fatta sotto una prospettiva canonica. Il reclutamento di aree prefrontali come ausilio nello svolgimento di compiti ‘difficili’ viene suggerito anche da osservazioni in soggetti anziani che mantengono buone prestazioni nei compiti di memoria di lavoro o di memoria a lungo termine: in tali soggetti, ma non in soggetti di pari età con prestazioni non buone, si osserva l’attivazione bilaterale di aree prefrontali, mentre in soggetti giovani l’attivazione è lateralizzata a un emisfero. La CPF ha anche un ruolo nel recupero delle tracce di memoria dichiarativa formatesi da lungo tempo: la CPF si attiva infatti durante la codifica e il recupero di memorie episodiche; il disturbo di tale funzione per mezzo di stimolazione transcranica magnetica, interferisce con la prestazione di soggetti impegnati in compiti di memoria episodica. Inoltre, lesioni alle aree associative prefrontali causano la cosiddetta amnesia della fonte, ossia l’incapacità di ricordare quando e dove un nuovo fatto è stato appreso. Danni al settore ventromediale della corteccia prefrontale sconvolgono il comportamento sociale: individui ben inseriti nella società diventano incapaci di osservare le regole sociali e di decidere in maniera per loro vantaggiosa mentre mantengono prestazioni normali in compiti di memoria di linguaggio e di attenzione. Il primo caso documentato in questo senso è stato quello di Phineas Gage, vittima di un grave incidente che lesionò la parte ventrale e mediale della CPF; in seguito all’incidente Gage, da persona affidabile e laboriosa, divenne inaffidabile, scioperato, incapace di organizzarsi nel lavoro e nella vita. Lo studio di pazienti di questo tipo è stato portato avanti, a partire dal 1994, da Antonio Damasio utilizzando test di laboratorio che simulano un gioco d’azzardo: i risultati mostrano che fra un mazzo di carte la cui sequenza darà un guadagno certo e uno in cui avranno una perdita certa, i soggetti con lesioni alle aree ventromediali, che includono la corteccia orbitofrontale, continuano a campionare il mazzo svantaggioso. Queste osservazioni hanno condotto Damasio a proporre che la incapacità dei soggetti con lesioni alle CPF ventromediali di prendere decisioni per loro vantaggiose è causata dal danno a un meccanismo emozionale che immagazzina e segnala il valore delle conseguenze future di una azione. Risulta evidente che la CPF ventromediale è implicata anche nei meccanismi decisionali e nei meccanismi alla base del comportamento emotivo e della qualità di ’attrattivo’ che alcuni stimoli hanno per noi (➔ piacere).
Corteccia frontale mediale. Quest’area sembra particolarmente responsabile della flessibilità del comportamento e del controllo cognitivo su di esso. La individuazione di conseguenze sfavorevoli, di errori di risposta, di risposte conflittuali e di incertezze decisionali, attivano zone nella corteccia frontale mediale ed evocano attività neuronale in un’ampia parte della corteccia frontale mediale posteriore. Questa include anche la corteccia cingolata anteriore, e ciò si correla con la successiva correzione della prestazione comportamentale. La corteccia frontale mediale posteriore sembrerebbe quindi implicata nell’attività di monitoraggio di comportamenti in contesti in cui si anticipa la presenza di una ricompensa, mentre la CPF laterale sembrerebbe implicata nell’implementazione delle correzioni alle strategie comportamentali.
Strutture mediali del lobo temporale. Queste strutture svolgono un ruolo cruciale nella memoria dichiarativa. Lo studio delle strutture coinvolte nella memoria dichiarativa inizia praticamente con le osservazioni di William B. Scoville e Brenda Milner negli anni Cinquanta. Il caso più interessante è stato quello del paziente H.M., cui vennero asportati bilateralmente i lobi temporali mediali. Dopo l’operazione, H.M. mostrava una normale memoria di lavoro ma mostrava una completa amnesia anterograda: la capacità di formare nuove memorie a lungo termine per fatti (memoria semantica) ed eventi (memoria episodica) era scomparsa. H.M. mostrava anche amnesia retrograda limitata, ossia la perdita di memoria per fatti ed eventi accaduti prima dell’intervento che si estendeva all’indietro per qualche anno (amnesia retrograda). La memoria di fatti ed eventi avvenuti diversi anni prima dell’intervento era intatta. Questo implica che l’ippocampo e le altre strutture del lobo temporale mediale sono cruciali per la formazione iniziale di una traccia di memoria dichiarativa a lungo termine, alla cui formazione contribuiscono informazioni che vengono da aree associative differenti. La traccia di memoria, poi, sarebbe lentamente trasferita nel magazzino definitivo, costituito da aree associative neocorticali, tra cui le aree prefrontali. Nicoletta Berardi