
Ecologia del paesaggio
Ecologia del paesaggio
II concetto di paesaggio è nato nel periodo protoagricolo. Sebbene la sua formulazione scientifica esplicita risalga all'inizio del 19° secolo, esso è rimasto a lungo escluso dalla teorizzazione in ecologia: l'ecologia generale tratta solo gli organismi, le popolazioni e le comunità, e riconduce ogni sistema ecologico nell'ambiguo concetto di ecosistema. Soltanto la recente riformulazione dell'ecologia alla luce di nuovi paradigmi scientifici ha portato alla nascita dell'ecologia del paesaggio che studia struttura, funzioni e trasformazioni dei sistemi interagenti di ecosistemi. Ciò apre nuove prospettive teoriche nello studio della natura e consente nuovi importanti sviluppi applicativi. II concetto di integratività dei livelli di organizzazione biologica implica l'impossibilità di studiare il paesaggio con parametri derivati dal livello di ecosistema, e ne riconosce l'importanza come specifico livello di organizzazione biologica. L'ecologia del paesaggio si presenta come transdisciplina che indirizza gli altri contributi di tipo applicativo relativi alla pianificazione ecologica del territorio, alla progettazione e al recupero dell'ambiente, alla conservazione e alla valutazione ambientale.
Introduzione
L'ecologia del paesaggio e delle regioni studia struttura funzioni e trasformazioni di sistemi interagenti di ecosistemi: ciò apre nuove prospettive nello studio della natura e permette nuovi e importanti sviluppi applicativi. A livelle teorico, si deve subito notare che il concetto di integratività dei livelli di organizzazione biologica implica che non s possa studiare il paesaggio utilizzando parametri derivat dal livello di singolo ecosistema; al contrario, va riconosciuta l'esistenza di una struttura e una dinamica proprie del paesaggio come specifico livello di organizzazione biologica. Il paesaggio mostra tutti i caratteri che definiscono l'organizzazione della vita a ogni livello: infatti, se ne possono descrivere non solo la struttura, ma anche i confini, i sistem di comunicazione e di movimento della biomassa, i process di riproduzione, il livello di metastabilità e lo specifice comportamento dinamico. Di conseguenza, un nuovo settore di ricerca come l'ecologia del paesaggio apre la porta alle studio dell'ecologia integrata o unified ecology (Allen e Hoekstra, 1992), dall'organismo alla biosfera, in cui anche la componente antropica trova la sua corretta collocazione.
A livello delle applicazioni assume importanza innanzitutto il metodo transdisciplinare. Esso nasce dall'osservazione che le discipline interessate allo studio dell'ambiente sono varie ma non tutte forniscono lo stesso contributo. Il principio organizzativo dei vari livelli in cui si manifestano le forme di vita resta essenzialmente biologico, quindi solo tale principio può integrare tutti gli altri. Per tale ragione l'ecologia del paesaggio si presenta come transdisciplina capace di indirizzare altri contributi di tipo applicativo, in particolare nel campo della pianificazione ecologica del territorio, nella progettazione e nel recupero dell'ambiente, nella conservazione della natura, nella valutazione ambientale. Problematiche cruciali, come la frammentazione del territorio e la pianificazione del suo uso, il controllo delle reti ecologiche, persino l'influenza del territorio sul comportamento degli organismi, necessitano oggi sempre più delle conoscenze derivate dall'ecologia del paesaggio.
Paesaggio ed ecologia
Sviluppo del concetto di paesaggio nella scienza
Il concetto di paesaggio è molto antico: esso è nato con i primi insediamenti stabili delle popolazioni umane nel periodo protoagricolo, quando le esigenze di esplorazione funzionale del territorio diventarono questione di sopravvivenza. L'uomo doveva acquisire informazioni sull'intero mosaico di biogeocenosi che formava il proprio territorio, cioè sul paesaggio naturale, conditio sine qua non per poterlo sviluppare in senso agricolo e insediativo, ma anche per gestirlo e difenderlo. Questo approccio, che si basava su un'operazione di cosciente mutualismo con la natura, portò la religione verso il culto solare, causò una prima forma di regionalizzazione della cultura e dette inizio alle maggiori trasformazioni ambientali su scala territoriale ampia. Questo processo è testimoniato da diversi fatti. Reperti archeologici risalenti ai primi villaggi protoagricoli dell'inizio del Neolitico mostrano che gli uomini sapevano come combinare e trasformare diversi elementi del paesaggio naturale per creare un paesaggio antropico dove vivere in modo ottimale (Forman e Godron, 1986). Le raffigurazioni di mappe che risalgono a 4000 anni fa nel territorio dei Camuni rappresentano un'indubbia acquisizione del concetto di paesaggio (lngegnoli, 1993). Nei popoli di più antica civiltà, come quello ebraico, il termine paesaggio (noff) compare già nel Libro dei Salmi, 3000 anni fa (Naveh e Lieberman, 1990).
Nel mondo romano il paesaggio veniva denominato regio (regione) da Columella, noto agronomo del l° secolo d.C., sottolineandone l'aspetto geografico; il pittore paesaggista, che mostrava già buon senso prospettico, veniva denominato "pictor topiarius qui regiones formas pingit", mentre il panorama veniva chiamato prospectus. Nel Rinascimento iniziò a essere usato in Italia il termine attuale, derivato dal francese paysage (quindi dal latino pagus), ma soprattutto in ambito letterario e artistico (da Petrarca a Tiziano), come riassuntivo dei caratteri propri di un paese. Il senso di questi caratteri si nota bene negli scritti di Leonardo da Vinci, dato che in lui la curiosità scientifica e la ricerca della forma in tutte le sue molteplici espressioni procedevano di pari passo. Egli considerava "le membra de' paesi, sassi, piante e simili", paragonandole a quelle degli animali non soltanto in senso strutturale, ma anche funzionale. Non a caso, con i suoi studi sui paesaggi prealpini egli iniziò l'analisi della geomorfologia e propose una prima formulazione del concetto di vegetazione.
Dopo Columella e Leonardo, un secondo passo verso l'utilizzazione del termine "paesaggio" (landscape) in senso scientifico fu compiuto dagli architetti paesaggisti inglesi nel 18° secolo. Le prime grandi trasformazioni del territorio in senso industriale avevano spinto questi ultimi a porre fine al formalismo francese nella creazione dei giardini e dei primi parchi pubblici, per dirigersi invece verso una progettazione più conforme alla natura. Rimaneva tuttavia ancora preminente una concezione percettiva del paesaggio. Solo all'inizio del 19° secolo, con la scuola di A. von Humboldt in Germania, il termine "paesaggio" entrò pienamente nel vocabolario scientifico. Egli definiva il paesaggio come "Der Totalcharakter einer Erdgegend", cioè l'insieme di tutti i caratteri propri di un determinato territorio. Ciononostante, dai primi anni di quel secolo fino alla metà del secolo successivo, gli ecologi non furono propensi ad adottare il termine "paesaggio" né a trattare tale argomento. Dopo l'introduzione del concetto di ecologia, dovuto al naturalista (e pittore) tedesco E.H. Haeckel intorno alla metà del 19° secolo, e dopo la formulazione del concetto di bioceno si da parte di K. Moebius (1877), solo nel 1921 il geomorfologo tedesco S. Passarge (che era anche un medico) cominciò a parlare di fisiologia del paesaggio secondo la tradizione risalente a von Humboldt, esponendo concezioni assai moderne basate su una geomorfologia climatica integrata con le scienze naturali. Tali tentativi non furono seguiti dagli ecologi fino a quando, nel 1939, il biogeografo K. Troll propose finalmente il termine Landschaflsoekologie, ecologia del paesaggio (Troll, 1939).
Limiti dell'ecologia tradizionale
L'ecologia è nata di fatto come 'autoecologia', cioè come studio del rapporto fra organismi e ambiente, e si è evo Iuta sviluppandosi per capitoli separati, quando non addirittura contrapposti. È nota la diatriba fra ecologi di popolazione e di ecosistemi: i concetti di comunità (insieme di popolazioni) e di ecosistema (insieme di processi) sono stati ricomposti con un compromesso solo da E.P. Odum (1971). D'altra parte, R.V. O'Neill e collaboratori (1986) ricordano come tali punti di vista non siano mai del tutto conciliabili. Infatti, come scrivono T.F.H. Allen e T.W. Hoekstra (1992), la conservazione di materia ed energia (punto centrale per il concetto di ecosistema) risulta irrilevante per il concetto di comunità, e inoltre una parte di un ecosistema, per esempio una pathway (o sequenza funzionale di specie), può corrispondere a più parti in una comunità, perché ogni organismo può essere un membro autonomo di una comunità. Pure l'ecologia delle grandi scale, cioè quella dei biomi e quella della biosfera, si articola in capitoli separati: la prima è legata soprattutto alla biogeografia, la seconda ha portato alla global ecology (ecologia globale), a partire dalla nota ipotesi Gaia.
Inoltre, l'ecologia tradizionale si trova in difficoltà a trattare problemi di applicazione su scala territoriale, perché il concetto di ecosistema non può essere valido per ogni scala, specialmente quando sono in gioco diversi schemi spaziali. Si rammenti che l'ecologia degli ecosistemi considera l'eterogeneità dell'ambiente come un rumore di fondo (BIondel, 1986) al quale annettere un'importanza secondaria. Ma la realtà è ben diversa: l'eterogeneità e la variabilità del mosaico ambientale costituiscono una componente anche evolutivamente molto importante nel determinare la distribuzione degli organismi e le loro interazioni. Bisogna anche ricordare che l'ecologia degli ecosistemi non ha fatto nulla per ostacolare la logica dell'impatto fra l'uomo e la natura, una logica deterministica, che scaturisce in realtà da preconcetti, che non ha alcun senso scientifico e che impedisce di gestire il territorio in modo ecologicamente equilibrato.
l tentativi di superamento delle suddette difficoltà, attraverso metodi ecologici per studiare l'ambiente su scala paesistica presi a prestito da altre discipline, trovano d'altra parte limiti intrinseci nel concetto stesso di integratività dei livelli di organizzazione biologica. Con ciò si vuole intendere che non ha senso analizzare i sistemi ecologici a livello di paesaggio utilizzando le caratteristiche proprie del livello ecosistemico, perché questo costituisce un livello di organizzazione più semplice. Non ci si deve dimenticare che Odum, già nel 1971, scriveva che nello 'spettro biologico' sussiste un palese principio di integratività, per cui i risultati ottenuti a un certo livello di organizzazione aiutano a studiare il livello superiore, ma non possono mai spiegare completamente i fenomeni che si verificano in esso.
Riassumendo, ancora oggi gran parte dell'ecologia generale si limita a trattare i temi dell'ecologia degli organismi, delle popolazioni, delle comunità, e riconduce ogni sistema ecologico nell'ambiguo concetto di ecosistema. Va osservato infine che il principio di integratività evidenziato da Odum non è che un corollario del più importante principio delle proprietà emergenti (Lorenz, 1978): un tutto organico è superiore alla somma delle sue parti, per cui basta cambiare l'assemblaggio strutturale degli elementi di un sistema per ottenere un comportamento non solo diverso, ma spesso imprevedibile. È indispensabile, di conseguenza, analizzare le caratteristiche di un livello di organizzazione biologica studiando il comportamento intrinseco del livello stesso, perché le informazioni provenienti dai livelli inferiori sono tutt'al più parziali. È però nello stesso tempo indispensabile riferirsi a nuovi paradigmi scientifici, alla cui luce la disciplina ecologica è spinta a rinnovarsi.
Nuovi paradigmi scientifici
Sistemi complessi adattativi
Come scrive M. Gell-Mann (1994), un esempio meraviglioso della semplicità dei principi naturali che sono alla base di tutti i fenomeni che osserviamo è la legge di gravitazione. Tale legge è in grado di descrivere i processi di aggregazione della materia che portarono, nel corso dell'evoluzione dell'Universo, alla formazione di galassie e poi di stelle e di pianeti, compresa la Terra. Fin dal tempo della loro formazione, questi corpi manifestavano caratteristiche di complessità, diversità e individualità. Tali proprietà assunsero però nuovi significati con l'emergere di sistemi complessi adattativi. Sulla Terra questo sviluppo è legato all'origine della vita e al processo dell'evoluzione biologica che ha prodotto la sorprendente varietà delle specie, il comportamento degli organismi in sistemi ecologici, l'apprendimento e il pensiero negli animali e nell'uomo, l'evoluzione delle società umane (compreso l'aspetto economico- tecnologico), la formazione e l'evoluzione dei paesaggi.
La nostra specie, che almeno sotto qualche aspetto è la più complessa evolutasi sulla Terra, è riuscita a scoprire molto sui sistemi più semplici, ma gli studi sui sistemi complessi adattativi sono ancora all'inizio. Il carattere comune a tutti i processi evolutivi è che in ciascuno di essi un sistema complesso adattativo acquisisce informazione sul suo ambiente e sulla propria interazione con esso; l'esame di tale informazione gli permette di identificare regolarità che vengono condensate in una sorta di modello capace di fornire una guida per agire nel mondo reale. In ogni caso esistono vari schemi in competizione fra loro e i risultati dell'azione nel mondo reale vengono reintrodotti nel sistema, influenzando la competizione tra schemi mentre il sistema stesso viene trasformato.
Lo studio dei sistemi biologici, in quanto sistemi complessi adattativi collocati su una scala superiore a quella dell'organismo-individuo, è senza dubbio compito dell'ecologia. Va sottolineato però che il tentativo dell'ecologia tradizionale di rimanere di fatto nell'ambito del determinismo cartesiano non permette di affrontare compiutamente tale questione. Per esempio, il problema della scala è travisato, visto che tanto una pozza temporanea, quanto un'intera regione possono essere definiti come ecosistemi; di conseguenza, il paesaggio è relegato (se mai) in ambito geografico, e si pretende di studiarlo semplicemente conoscendo organismi, popolazioni e comunità. Inoltre, la mancanza di una freccia del tempo (il tempo inteso come irreversibile) porta a considerare la natura senza storia, quindi meccanicistica, sotto il dominio dell'uomo che se ne fa ingegnere.
Del resto, una descrizione della natura secondo leggi deterministiche, in cui il fenomeno è reversibile rispetto al tempo, è incapace di considerare novità e creatività, quindi è incapace di definire la vita al di fuori di un organismo o di un insieme di organismi. Quindi, è impensabile che un sistema interagente di ecosistemi (il paesaggio) possa realmente essere definito nell'ecologia tradizionale come un livello di organizzazione biologica. Peraltro, oltre a ostacolare l'incontro con la realtà, si può notare che il determinismo pone in discussione addirittura la libertà umana (Lorenz e Popper, 1985; Popper, 1996), che presuppone invece l'idea di creatività. È quindi indispensabile riferirsi a nuovi paradigmi scientifici.
I nuovi paradigmi di riferimento per l'ecologia
La più grande spinta verso una nuova concezione della natura viene forse da K. Popper, K. Lorenz e I. Prigogine, che hanno osservato come la natura realizzi le strutture più delicate e più complesse grazie a processi irreversibili associati alla dimensione temporale; questi autori sostengono che l'indeterminismo è compatibile con il realismo, che un tutto organico è più complesso della somma delle sue parti, e che la vita è possibile solo in un Universo lontano dall'equilibrio.
Lo studio di ogni ramo della realtà, che appare in sé continua, inizia con una separazione in senso sistemico. Dalla formulazione, nel corso del 20° secolo, di importanti teorie epistemologiche quali la teoria della complementarità, il teorema di Goedel, la teoria dell'informazione e la teoria dei sistemi gerarchici, sappiamo che per prevedere il comportamento di un sistema compatibile con le leggi della natura non sono più sufficienti i paradigmi meccanicistici, per i quali la conoscenza di uno stato di un sistema è ottenibile mediante la determinazione delle sue coordinate (Ingegnoli, 1971).
Per conoscere lo stato di un ecosistema o di un paesaggio, al contrario, non è possibile limitarsi a un paradigma meccanicistico. Non possiamo più supporre che lo stato di un oggetto sia indipendente dal modo in cui viene osservato, perché le sorgenti di informazione sull'oggetto vengono alterate dall'osservazione stessa. Inoltre, i sistemi complessi contengono e includono i disturbi passati nelle componenti di livello più basso. Tutto ciò pone in evidenza l'importanza della storia, della scala e del contesto quali basi di conoscenza e di integrazione.
Prigogine (Prigogine et al., 1972; Prigogine e Nicolis, 1977; Prigogine, 1996) osserva che è necessaria una nuova oggettività, diversa da quella della termodinamica classica, che identifica il conoscibile con il controllabile, e da quella dinamica, che tenta di risolvere l'evoluzione di un sistema partendo da uno stato istantaneo. Una struttura prodotta da una successione di fluttuazioni amplificate non può essere compresa che in riferimento al suo passato. Tale passato, prodotto da eventi imprevedibili, deve essere considerato unico e non riproducibile. Tale assunto concorda con il principio delle proprietà emergenti e rivaluta in modo inequivocabile l'importanza della storia, anche in assenza dell'uomo, nello studio dell'ecologia. Sulle orme dei pionieri di questa accezione più complessa e completa della storia, che risale tra l'altro a C. Cattaneo e F.-P. Braudel, interessa rilevare il concetto di localizzazione della storia espresso da L. Zanzi (1995) come "avvenimentale", in cui i diversi protagonisti (alberi, animali, uomini, configurazioni e assetti del terreno e così via) evolvono interagendo vicendevolmente.
l sistemi dinamici sono considerati instabili se piccole modifiche delle condizioni iniziali producono effetti cospicui, che si amplificano nel tempo. Per esempio, i sistemi caotici, dove traiettorie quasi combacianti al momento iniziale divergono esponenzialmente nel tempo, sono instabili. La differenza fra i sistemi dinamici stabili e quelli instabili è dunque fondamentale. Di solito, lo stato dinamico di un sistema è rappresentato da un punto nello spazio delle fasi (cioè lo spazio individuato da posizioni e quantità di moto). Volendo considerare anche i sistemi instabili, si nota invece con Prigogine (1996) che le condizioni iniziali non sono più assimilabili a un punto in tale spazio, ma corrispondono a una regione descritta da una distribuzione di probabilità. Si tratta quindi di una descrizione non puntuale, ma paesaggistica. Tutto ciò comporta, inoltre, una rottura nella simmetria temporale a seguito della quale, nella formulazione statistica, passato e futuro vengono a svolgere ruoli diversi, in quanto l'instabilità distrugge l'equivalenza tra il livello individuale e quello statistico e di conseguenza le probabilità assumono un significato intrinseco, non riducibile al caso delle traiettorie individuali. La distribuzione di probabilità alle condizioni iniziali, quindi, permette di includere nella descrizione dinamica la microstruttura complessa dello spazio delle fasi. Tale informazione, addizionale rispetto alla descrizione individuale, permette di prevedere l'evoluzione dell'insieme. Solo se si devono descrivere sistemi stabili si può tornare alla dinamica tradizionale. Ma in biologia, di solito, i sistemi stabili non sono di interesse.
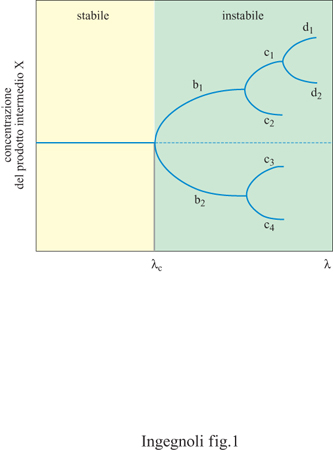
Ogni sistema dinamico può essere caratterizzato da un'energia cinetica, che dipende dalla velocità delle componenti, e da un' energia potenziale, che dipende dall'interazione fra le componenti. In un mondo isomorfo a un insieme di corpi non soggetti ad alcuna interazione, non c'è posto per la freccia del tempo, né per l'autoorganizzazione, né per la vita. Tuttavia, come ha osservato l-H. Poincaré (1906), l'esistenza di risonanze tra i gradi di libertà del sistema produce condizioni di non equilibrio. In queste condizioni si producono tra le componenti correlazioni di grande portata. Si verifica cioè una sensibilità nuova della materia a sé e al suo ambiente, associata alla dissipazione e ai processi irreversibili. Il più noto esempio in tal senso, riportato da Prigogine, è quello della concentrazione del prodotto intermedio X di una reazione chimica: oltre il ramo termodinamico, stabile, si ha un campo di instabilità con la comparsa di biforcazioni successive (fig. 1). Nei punti di biforcazione diventano importanti i disturbi, cioè le fluttuazioni che permettono al sistema di scegliere uno dei due rami di nuova stabilità relativa. L'evoluzione di un simile sistema ha in sé un elemento storico. Per esempio, il fatto che il sistema venga osservato nello stato d2 (v. figura 1), implica che esso è passato per gli stati b1 e c1. Inoltre, se applichiamo il concetto di Prigogine dell'autoorganizzazione delle strutture dissipative alla società umana, è possibile dimostrare, fra l'altro, che l'attività dell'uomo, creativa e innovatrice, non è estranea alla natura, ma può anzi essere considerata un'amplificazione e un'intensificazione di aspetti già presenti nel mondo fisico che la scoperta dei modelli per i processi lontani dall'equilibrio aiuta a decifrare.
Ecologia del paesaggio
Alle frontiere dell'ecologia
Alla luce delle teorie scientifiche sopra esposte, è possibile mostrare i limiti degli approcci dell'ecologia tradizionaIe. l problemi di scala, di complessità, di contesto, di rapporto uomo-ambiente, di autoorganizzazione e persino di definizione dei livelli biologici sono stati finora poco considerati. Per arrivare alle frontiere dell'ecologia, è importante partire dal fatto che l'organizzazione biologica dipende dalla dinamica delle strutture dissipative, associata ai processi irreversibili. La teoria delle strutture dissipative e i concetti di metastabilità e di incorporazione dei disturbi permettono così di giungere alla storicità dei sistemi biologici e alla loro evoluzione gerarchica (O'Neill et al., 1986).
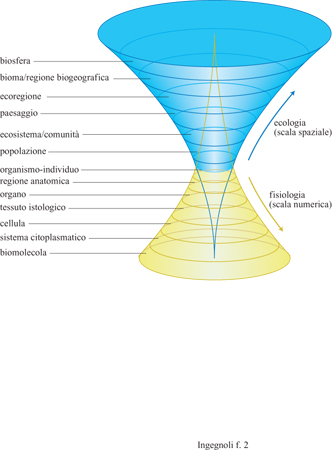
L'ecologia generale (Whittaker, 1975) definisce il paesaggio come contesto spaziale per le comunità (o gli ecosisterni), cioè come un gradiente geografico capace di influire sulle strutture e sui processi ecologici a livello di organismi, popolazioni e comunità. Questa definizione è senza dubbio necessaria, ma non sufficiente, proprio perché da essa il paesaggio non risulta come specifico livello di organizzazione della vita. Come abbiamo già notato, basterebbe accettare lo 'spettro biologico' (fig. 2) come derivato dalla teoria dei sistemi gerarchici, e il principio delle proprietà emergenti, per arrivare a considerare il paesaggio come un sistema ecologico non del tutto descrivibile con i caratteri dei precedenti livelli (popolazioni, comunità o ecosistemi). Il paesaggio come 'meta-eco sistema' deve infatti possedere anche suoi specifici caratteri e comportamenti.
Comunque, per definire il paesaggio nel senso che si vuole sostenere qui, è necessario richiamare il concetto di ecosistema, la cui componente biotica è la comunità (Odum, 1971; 1993; Whittaker, 1975), e la teoria della termodinamica di non equilibrio (Prigogine et al., 1972; Prigogine, 1996).
Il paesaggio come sistema di ecosistemi
Bisogna tenere presente che, quando un intenso flusso di energia attraversa un sistema, appaiono solitamente strutture dissipative caratterizzate da notevole instabilità. I sistemi viventi, auto organizzanti, catturano l'energia che viene poi utilizzata per produrre nuove strutture: "order through fluctuations", scrive Prigogine. Da questo punto di vista, un ecosistema è un sistema aperto, lontano dall'equilibrio termodinamico, e quindi dissipativo; di conseguenza esso è caratterizzato dalla medesima importanza degli scambi sia interni che esterni che possono essere descritti dalla termodinamica dei sistemi lontani dell'equilibrio. In un sistema aperto la diseguaglianza di Clausius-Carnot prende la forma:
dS=deS+diS⋛0
dove deS è la variazione di entropia dovuta agli scambi con l'ambiente, diS è la produzione di entropia dovuta ai processi irreversibili entro il sistema. Poiché deS può assumere valori negativi, ne risulta che, durante la sua evoluzione, un sistema può raggiungere uno stato in cui l'entropia è minore di quella iniziale, cioè uno stato più ordinato. Ciò è valido sia per una zona di foresta sia per un'area urbanizzata, e necessita la presenza di confini ('superfici' o 'membrane') per ogni unità ecosistemica. È necessaria inoltre l'esistenza di un sistema di ecosistemi, cioè di un contesto esterno di unità similari interagenti, e l'esistenza di popolazioni, cioè di strutture interne per ogni ecosistema. Il contesto esterno, lo specifico sistema di ecosistemi, è ciò che noi definiamo paesaggio. Questo contesto è un sistema di ecosistemi, quindi interagisce direttamente con i suoi componenti, scambiando specie, energia, materia e informazione e seguendo il principio delle proprietà emergenti, quindi creando una propria struttura e proprie dinamiche. Un'importante osservazione deriva subito da quanto detto: un elemento del paesaggio assume uno specifico ruolo funzionale nel suo complesso mosaico ecologico.
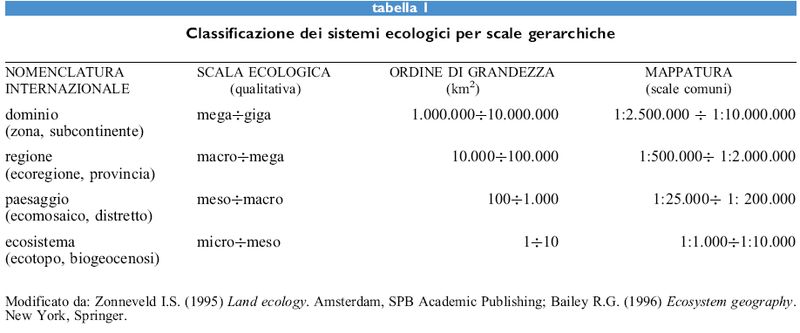
Certo, un sistema di ecosistemi fra loro interagenti rappresenta una struttura molto complessa, difficile da comprendere in modo intuitivo e quindi difficile da rappresentare. Può essere utile ricordare che ogni sistema biologico mostra una struttura formata da insiemi funzionali ben definiti in un contesto di substrati variabili nello spazio e nel tempo; ciò vale dalle cellule fino agli ecosistemi, ai paesaggi, alle regioni e all'intera biosfera. Il paesaggio mostra tutti i principali caratteri che definiscono ciascuno dei livelli di organizzazione della materia vivente; infatti, se ne possono descrivere la struttura, ma anche i confrni, i sistemi di comunicazione e di movimento della biomassa, i processi di riproduzione, il livello di metastabilità e lo specifico comportamento dinamico (v. oltre). Il paesaggio è inoltre individuabile in una ben determinata gamma di scale (tab. I), poiché il suo livello inferiore è proporzionato all'insieme di specie che formano una comunità, e il suo livello superiore è contenuto nella regione di appartenenza biogeografica.
Va ricordato a questo proposito che non si può studiare un ecosistema al di sotto della scala propria della comunità che lo forma e senza conoscere le popolazioni che lo compongono. Quando, allargando la scala, si riscontrano altri elementi che sono al di fuori della struttura e delle funzioni di un ecosistema, è inevitabile parlare di paesaggio. Per esempio, raggiunta la scala di un ecomosaico, appaiono nuove strutture e nuovi processi: reti ecotonali, connettività fra ecosistemi, tipo di porosità della matrice paesistica, tipi di apparati paesistici, dinamica di paesaggio, nuove strategie di metastabilità, e così via. Analogamente, altre caratteristiche appaiono su scala regionale e non si possono studiare a livello di paesaggio: l'ordinamento dei suoli, la formazione delle foreste, i cambiamenti nella fauna, i gradienti biologici nei fiumi, il ruolo climatico nella differenziazione ecologica e così via.
Bisogna notare che, con l'allargamento della scala nei sistemi ecologici, cambiano non solo molti caratteri del livello precedente, ma anche molti fattori dominanti di controllo, secondo quanto previsto dalla teoria dei sistemi gerarchici. Anche i fattori di controllo antropico cambiano palesemente con il crescere della scala, come si può notare osservando le differenze di struttura nei livelli di organizzazione ecologica che appaiono da un rilevamento aerofotogrammetrico usuale (circa 1:25.000) in confronto a un telerilevamento da satellite (circa 1:1.000.000). Nel primo caso, l'influenza dell'uomo è solitamente rilevante, nel secondo è spesso più rilevante quella geomorfologica e topografica. Notiamo comunque che la presenza di ecosistemi antropizzati può essere un carattere distintivo, ma non certo un carattere essenziale di un paesaggio anche se analizzato su una scala di dettaglio.
La definizione di paesaggio viene così a precisarsi; di conseguenza, si può definire l'ecologia del paesaggio come quella parte dell'ecologia che studia il paesaggio nel senso dell'organizzazione biologica, e con ciò contribuisce all'avanzamento dell'ecologia integrata e della pianificazione del territorio. L'ecologia del paesaggio diventa così indispensabile per la conservazione biologica e deve essere considerata come uno dei capitoli di base dell'ecologia integrata.
Caratteri propri di un sistema di ecosistemi
Caratteristiche strutturali proprie
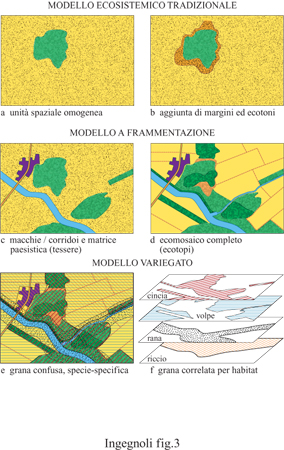
l principali modelli strutturali del paesaggio sono due: quello cosiddetto a frammentazione e quello cosiddetto variegato (fig. 3). Tali modelli rappresentano due visioni opposte: una statica e una dinamica. Non sembra però del tutto corretto contrapporre un mosaico formato da tessere giustapposte di elementi del paesaggio (modello a frammentazione) a un mosaico a geometria variabile di tessere speciespecifiche (modello variegato). In realtà, ogni sistema biologico mostra una struttura formata da insiemi funzionali ben definiti in un contesto di substrati variabili nello spazio e nel tempo. È meglio invece parlare di tessuto ecologico o ecotessuto, cioè di un mosaico multi dimensionale individuato da un mosaico di base e da una serie gerarchica di mosaici correlati:
Ect=Mb ⋃ (⋂)M₁⋃ (⋂)M₂ ... ⋃(⋂)Mk
dove: Ect è l'ecotessuto, Mb il mosaico di base, M₁ ... Mk i mosaici correlati e i simboli insiemistici di unione e di intersezione possono essere intercambiabili o addirittura coesistere, trattandosi spesso di insiemi eterogenei.
Si noti che il mosaico di base è solitamente individuabile come quello formato dalle biogeocenosi vegetali, in quanto esse sono i più importanti controllori del flusso di energia e di materia, hanno la capacità di costruire l'ambiente e di interagire col suolo e di formare un microclima. Oltre al termine "tessera" si può utilizzare, per un elemento del paesaggio, il termine "ecotopo". L'ecotopo va però inteso non solo come biogeocenosi (secondo la scuola tedesca e russa), cioè con denotazione ecosistemica, sia pure integrata, ma anche come espressione di un ruolo paesistico, cioè come nicchia territoriale con funzioni individuabili nel contesto del paesaggio di cui fa parte.
Fra i primi studi effettuati sul paesaggio vi sono quelli che riguardano l'individuazione di un disegno strutturale proprio e di una fisionomia propria. Dato che la geometria della natura è caotica e di rado si identifica con quella euclidea, essa si può descrivere con la geometria dei frattali. Per geometria frattale si intende, secondo B.B. Mandelbrot (1975), lo studio degli oggetti geometrici di dimensione euclidea frazionaria: per esempio una linea di costa, che ha dimensione compresa fra 1 e 2, dato che la lunghezza tra due punti, passando di scala, tende a infinito; oppure la chioma di un albero, che presenta dimensione frattalica ed è assimilabile visivamente al concetto euclideo di volume ma non è formalizzabile come tale con la geometria. Due gruppi di forze sembrano preponderanti nel modellare il paesaggio e sono rivelati dalla dimensione frattale media: su piccola scala agiscono le influenze umane, su grande scala quelle topo grafiche. La dimensione frattale dei mosaici sia antropizzati sia naturali rimane inoltre costante per una gamma decisamente ampia di scale, e ciò è spiegabile perché i modelli strutturali di base, componibili e ripetibili in un intorno, sono soltanto cinque o sei in tutto.
La distribuzione, il tipo della grana e la frammentazione dei mosaici ecologici aiutano a studiare la fisionomia dei paesaggi, per esempio con osservazioni sulla porosità della matrice paesistica. Ma anche la struttura degli apparati paesistici, cioè di insiemi di ecotopi di ruolo funzionale simile, serve a capire il disegno del paesaggio e agevola la classificazione.
Strato di delimitazione e filtro
Margini e fasce ecotonali acquistano in questo contesto una nuova importanza perché, mentre nell'ecologia delle comunità erano visti come un'interfaccia fra l'omogeneità di un ecosistema e l'intera eterogeneità dell'ambiente circostante, nell'ecologia del paesaggio essi costituiscono un sistema analogo a quello delle membrane cellulari in un tessuto istologico (Forman e Moore, 1992). Va tuttavia sottolineato che, mentre gli strati di biomassa vegetale costituiscono la delimitazione più evidente degli ecotopi, a livello di paesaggio assumono maggior peso le barriere geomorfologiche e antropiche. L'azione dell'uomo ha portato spesso a definire in modo netto confrni che naturalmente erano rimasti sfumati per tempi assai lunghi, ma ciò non va considerato sempre negativamente perché può aiutare a comprendere la funzionalità degli ecomosaici. Si può anzi ipotizzare che, fra le funzioni naturali dello sviluppo così imponente delle popolazioni umane, vi sia anche quella di mettere in evidenza la rete di confrni fra le diverse comunità. Anche altre popolazioni animali (per esempio, i castori o, su scala minore, le formiche e le termiti) usano trasformare confrni incerti in margini più netti, influendo così sull'eterogeneità e sulla dinamica dei mosaici ecologici.
Sistema interno di comunicazioni
Catene interagenti di organismi si comportano come reti di comunicazione interna in un ecotessuto, tanto che attraverso di esse è possibile il mantenimento di un certo livello di metastabilità. A tale sistema va riferito in particolare il concetto di coevoluzione, che si esplica in pieno solo comprendendo anche il livello di paesaggio; infatti, a seconda delle scale di interazione con altri livelli dell'organizzazione biologica, un organismo reagisce di solito in modo diverso. Per esempio, l'interazione di un organismo con un altro organismo può portare alla competizione; l'azione dell'ambiente su di una comunità di organismi può portare al mutualismo; infine, l'azione di un ecotopo su di un altro ecotopo porta di solito alla coevoluzione.
Per tali ragioni il concetto classico (darwiniano) di adattamento si deve allargare per comprendere un concetto nuovo, in cui il principio guida della strutturazione dei sistemi biologici non è solo quello di causa, ma anche quello di coazione (o azione integrata o complementare), e le forze non operano solo in termini di conflitto e impatto, ma anche di cooperazione e integrazione. Ciò è coerente con quanto sostiene Popper (1996): "Semplicemente non è vero che noi veniamo plasmati dall'ambiente. Siamo noi che cerchiamo l'ambiente e che lo plasmiamo attivamente. Il gene nudo ha cercato un ambiente di proteine e con queste si è costruito un mantello [ ... ]. Noi cerchiamo di cambiare e modificare il nostro ambiente immediato, poi quello più lontano e, infine, l'intero mondo". Se anche il rapporto uomo-natura si pone in quest'ottica, si può notare come esso non sia in contrasto con i principi dell'evoluzione.
Superato così il preconcetto della contrapposizione uomonatura, si osserva che un ruolo di primaria importanza nel sistema di comunicazioni e informazioni su scala paesistica è svolto dagli ecosistemi antropici, che sembrano essersi evoluti con un compito primario di tipo informativo-gestionaIe (Lorenz, 1978; Godron, 1984; lngegnoli, 1993; 1997). La capacità di pianificare l'ambiente ha quindi un significato ecologico ed etologico di notevole rilevanza naturalistica e si esplica a livello di paesaggio.
Sistema interno di spostamento di biomasse, materia ed energia
La configurazione di un ecotessuto influisce in modo caratteristico su diversi flussi: dal movimento degli organismi ai flussi biogeochimici, a quelli genetici. Anche i movimenti delle acque e del vento o delle particelle minerali sono legati alla configurazione dei paesaggi. Molti studi sono stati compiuti in proposito, spesso utilizzando modelli neutrali, come quello di percolazione (derivato dalle modalità di filtrazione dell'acqua nel suolo), anche se diverse questioni rimangono irrisolte. La configurazione di un ecotessuto è legata al sistema ecotonale poiché in entrambi i contesti assumono importanza i problemi di filtro, barriera e corridoio. In particolare, i corridoi sembrano le strutture più interessanti per lo studio dei movimenti di biomassa.
Grande rilevanza assume il concetto di connettività fra elementi del paesaggio, in cui si possono distinguere fattori di connessione e circuitazione, ma anche differenze fra corridoi propri, impropri ed ecotopi a funzione di connettivo. Può essere inserito in questo capitolo anche il concetto di source and sink (sorgente e scarico), che conduce ai concetti di metaclimax e di metapopolazione, cioè insiemi interconnessi di subunità a stadi evolutivi diversi. Anche il reticolo fluviale rappresenta un sistema di spostamento di notevole importanza, perché nel paesaggio svolge il compito di apparato escretore. Infatti, la capacità di depurazione dei cataboliti provenienti dagli ecomosaici circostanti è dovuta alla struttura e alle funzioni del corridoio complesso formato dal fiume e dalla vegetazione di ripa (lngegnoli, 1993).
Autoriproduzione
Ai livelli organizzativi di ecotopo e di paesaggio la riproduzione risulta ben diversa da quella degli organismi o delle cellule, anche se rimangono validi alcuni fattori di base. Le informazioni da trasmettere vengono circoscritte nello spazio e nel tempo, per esempio in banchi di propaguli, e deve avvenire un processo scatenante, cioè un evento zero (zero event), come il fuoco, che permetta ai propaguli di sostituirsi alle precedenti strutture organiche (Oldeman, 1990).
Al concetto di evento zero è legata una diversa valutazione dei disturbi nei mosaici ecologici: sono strutturanti persino gradi elevati di perturbazione, in quanto necessari per l'autoriproduzione delle tessere di un mosaico ecologico. Infatti, senza perturbazioni capaci di azzerare le strutture esistenti di un ecotopo, sarebbe impossibile il rinnovamento riproduttivo dell'ecotopo stesso.
Gioca un ruolo importante anche la ricolonizzazione: va ricordato infatti che gli stadi pionieri tendono a essere stocastici, ma le dinamiche pioniere sono orientate dalle stesse condizioni di vincolo che si verificano in un dato ecotessuto.
Mantenimento della metastabilità
L'interpretazione gerarchica dei sistemi biologici indica come le interazioni fra i componenti di livello inferiore siano controllate da interazioni più lente ai livelli superiori. Ne consegue che i sistemi di ecosistemi tendono ad avere maggiore omeostasi (cioè maggiore capacità di incorporazione dei disturbi) degli ecosistemi componenti, e che questi a loro volta possiedono una maggiore omeostasi delle popolazioni componenti. Alcune interazioni a breve termine sono invece cicliche o oscillanti (per esempio, il rapporto preda-predatore). Tutto ciò comporta che anche nei paesaggi vengano mantenuti caratteristici livelli di metastabilità. L'organizzazione dei livelli biologici può procedere solo se lo stato finale di un sistema biologico è meno instabile (più metastabile) dello stato iniziale. Ma la massima metastabilità per un sistema di ecosistemi non corrisponde alla somma delle massime metastabilità dei suoi componenti. Ciò significa che in un paesaggio, in cui ogni ecosistema tenda al suo climax, non si raggiunge la massima metastabilità. Lo stesso vale per la biodiversità paesistica (BIondel, 1986; lngegnoli, 1993; Odum, 1993).
Il ridimensionamento conseguente del concetto di climax è di notevole rilevanza anche nel campo applicativo. Nella trattazione teorica, inoltre, la successione ecologica non risulta più un processo lineare (orientato), dato che nel paesaggio ciò condurrebbe a una minore capacità di incorporazione dei disturbi.
Comportamento dinamico proprio
Esistono diverse varietà di comportamento dinamico proprio, e qui di seguito verranno illustrati alcuni esempi tratti da contesti specifici.
Processi paesaggistici generali. - l processi paesaggistici generali sono responsabili della formazione dei fisiotopi, cioè dei substrati geomorfologici degli ecotopi, e interessano le interazioni fra clima, vegetazione, suolo, animali e uomo. Si può studiare l'evoluzione dei tipi di paesaggi in rapporto alla loro comparsa nella bio sfera, a partire dalle prime colonizzazioni paleozoiche delle terre emerse. Viene analizzata anche l'integrazione dei processi morfologici: ambienti geodinamici stabili, intermedi e instabili, e loro distribuzione territoriale. Per integrare le azioni morfogenetiche nel sistema ecopaesistico, esse si devono definire secondo il duplice criterio spaziale (come fenomeni localizzati o diffusi) e temporale (come fenomeni intermittenti e discontinui). Anche i processi pedologici, in particolare le distribuzioni catenali, sono di notevole importanza, come aveva già evidenziato nel 1898 V.V. Dokucaev per la legge della zonalità (Dokucaev, 1951). Infine, i processi idrologici, soprattutto di erosione e ruscellamento per tipo di ecotopo e di regime fluviale, sono quelli che maggiormente caratterizzano gli ecotessuti.
Funzioni di configurazione di elementi e relativi movimenti ecologici. - Le caratteristiche delle macchie rapportate in funzione dei tipi di ecotessuto mostrano variazioni particolari, la cui conoscenza è utile per valutare lo stato di un paesaggio. Fenomeni come quello dell'interdigitazione a penisola fra due tipi di elementi paesistici producono specifici processi. L'influenza dell'effetto margine in rapporto alla grandezza e alla frammentazione delle macchie e dei corridoi è pure una funzione tipica dei paesaggi. Riguardo ai movimenti che dipendono dalla struttura del paesaggio, basti dire che essi si verificano per l'effetto filtrante dei margini degli elementi, per il fenomeno source and sink e per i differenti stadi evolutivi degli elementi, oltre che per effetti geomorfologici. Tali movimenti dipendono da un numero ristretto di vettori, quali vento, acqua, animali volanti, terricoli e acquatici, uomini e loro mezzi. Le forze in gioco sono diffusione, trasporto, locomozione.
Funzioni di popolazioni permeanti e antropico-ecologiche. - Le funzioni svolte dalle popolazioni sono particolarmente numerose. Si può ricordare il processo del dimorfismo, mantenuto da pressioni di selezione antagoniste che si esercitano su scala di paesaggio (metapopolazione). Altri processi sono legati al paesaggio per le differenti utilizzazioni fatte da specie che necessitano di grande eterogeneità di habitat e che individuano e trasformano località di pascolo, abbeveraggio, rifugio, incontro, ritualizzazione e così via. Anche estinzione, isolamento e ricolonizzazione rientrano nel rapporto funzionale fra paesaggi (o loro parti) e popolazioni permeanti (ovvero popolazioni che utilizzano abitualmente diversi tipi di ecosistemi).
Più specifici forse sono i recenti concetti di complementazione e supplementazione. Quando una specie necessita di risorse differenti presenti in ecotopi di tipo diverso, si verifica un processo di complementazione se l'entità della popolazione è maggiore in ecotessuti in cui i due tipi di habitat sono frammisti. Nel caso della supplementazione, la popolazione di un ecotopo può risultare maggiore se tale unità di paesaggio è collocata in un ecotessuto che disponga di risorse addizionali. L'andamento della densità di popolazione di una specie in un determinato paesaggio dipende anche dall'occupazione sequenziale di un habitat ottimale, uno subottimale e uno marginale.
Dato che il maggiore agente permeante è l'uomo, è importante accennare alle funzioni paesaggistiche dell 'uomo secondo i principi ecologici. Con opportuni indicatori (Turner e Gardner, 1990; Ingegnoli, 1991; 1993; 1997) si possono evidenziare i processi di formazione dei tipi di paesaggi antropici in rapporto all'habitat agricolo pro capite, alle esigenze residenziali e industriali, al verde seminaturale protettivo, al rapporto habitat umano/habitat naturale di un territorio, alla capacità biologica territoriale (Btc) e così via, e individuare, per ogni regione ecologica, i limiti della struttura dei paesaggi urbano denso e rado, suburbano, rurale, agricolo, agricolo-protettivo. È anche possibile individuare l'habitat standard ecopaesistico minimo, teoricamente necessario per operazioni di compensazione nelle trasformazioni antropiche su scala locale.
Dinamica dei disturbi e di trasformazione. - Tutti i paesaggi mostrano modalità specifiche di trasformazione, che rimangono valide per una gamma di scale piuttosto ampia. Tali modalità possono comprendere anche processi omeoretici (capaci cioè di mantenere un regime dinamico), come nel caso di paesaggi mediterranei mantenuti da meccanismi di retro azioni cibernetiche che controllano la pressione di pascolo. Anche i paesaggi antropici presentano modalità di trasformazione guidate da leggi ecologiche, come quella della complementarità dei fenomeni di trasformazione ambientale, che sono capaci persino di modificare la cultura e, di conseguenza, il comportamento umano in modo da mantenere un equilibrio metastatico, cioè che oscilla intorno a un attrattore stabile ma capace di spostarsi verso un altro attratto re nei paesaggi in forte modificazione.
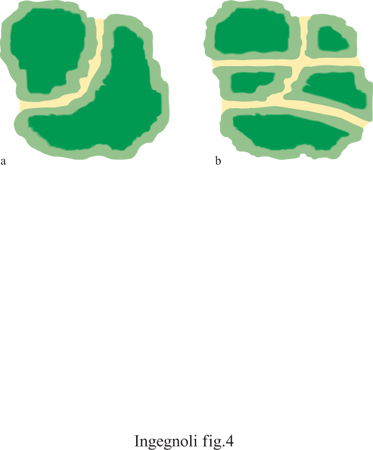
Processi di alterazione e patologia. - Eventuali segni di degrado del paesaggio si avvertono quando un'alterazione si manifesta a livello di ecotessuto. Se gli ecotopi alterati vengono sostituiti da altri ecotopi si assiste a una trasformazione; ma se l'ecotessuto non è in grado di incorporare i disturbi manifestati nel suo mosaico, allora inizia il degrado. Di solito sono i disturbi fuori scala che rompono l'organizzazione dei sistemi ecologici, con conseguenze anch'esse fuori scala. Si noti però che in certi casi, per esempio nella frammentazione (fig. 4), basta una trasformazione relativamente limitata per procurare alterazioni sensibili. È importante rilevare a questo proposito che il degrado dei paesaggi ha influenza sulla fisiologia di molti organismi che vivono in essi, uomo compreso. Lo studio dei paesaggi permette infine di formare un quadro delle patologie più ricorrenti a essi collegate, quali l'alterazione strutturale degli ecotessuti, le alterazioni funzionali, le sindromi di trasformazione impropria, il degrado da inquinamento, le distruzioni per disturbi catastrofici.
Nuove prospettive in ecologia
L'ecologia integrata
Si cercherà ora di illustrare quali sono gli apporti più significativi dell'ecologia del paesaggio allo sviluppo della teoria ecologica. Il determinismo, legato al concetto di reversibilità del tempo, ha prodotto l'ecologia tradizionale; come abbiamo visto, nuovi paradigmi scientifici hanno sancito il superamento dei limiti di quella ecologia fino a portare alla nascita dell'ecologia del paesaggio. Ciò ha permesso di completare lo studio dello 'spettro biologico', ridimensionando il concetto di ecosistema. È dunque ora possibile, come notano già alcuni studiosi (Allen e Hoekstra, 1992), tendere verso un'ecologia integrata più coerente e con maggiori prospettive applicative.
L'esigenza avanzata dagli ecologi oggi è che l'ecologia sia una disciplina predittiva e che gli studiosi siano in grado di considerare i sistemi complessi di grande scala. La risposta a questa esigenza passa sia per una consistente validazione epistemologica, come si è già sottolineato, sia per una più corretta interpretazione della gerarchia dei livelli di organizzazione biologica, unica strada per arrivare finalmente a un'integrazione dei diversi capitoli dell'ecologia.
La teoria dei sistemi gerarchici è stata introdotta da O'Neill e collaboratori (1986). Si sta superando oggi il modello gerarchico dell'ecologia tradizionale (organismo, popolazione, comunità, ecosistema, bioma, biosfera) con l'introduzione dell'ecologia del paesaggio e delle regioni. Si sta superando anche il concetto deterministico di fattore limitante, con l'introduzione del più olistico concetto di coazione. Tuttavia, nelle definizioni dei principali criteri di studio vi sono ancora delle inesattezze.

Si può osservare che una parte delle ambiguità del concetto di ecosistema deriva dal fatto che esso è inserito in una gerarchia di tipo prettamente biotico, pur avendo significato ambientale (funzionale). L'aggiunta ulteriore del concetto di paesaggio rende tale gerarchia ancora più incoerente. Infatti, procedendo dalla scala più dettagliata verso quella globale, si può notare (tab. 2) che vi sono tre gerarchie parallele, basate rispettivamente sul criterio biotico, relativo all'organismo o a insiemi di organismi, su quello ambientale, relativo agli aspetti geografici o ecologici di settore, e su quello ecologico, capace di integrare i due precedenti.
Odum (1971; 1993) elenca otto livelli di scala. Tuttavia, i livelli di scala in ecologia possono essere limitati a sei (sette se si considerano grandi unità di biosfera), corrispondenti a sei scale di osservazione legate a sei livelli di organizzazione biologica (v. tabella 2). L'ambiguità ancora presente nella figura 2 deriva dai criteri adottati. Il criterio biotico è adeguato anche in ecologia solo per le prime due scale mostrate in tabella 2, perché in tal caso una piena integrazione fra criterio biotico e ambientale è impossibile, essendo questi livelli non auto sufficienti in senso ecologico. Il criterio integrato è invece obbligatorio su scala maggiore, perché è l'unico in grado di considerare l'intera organizzazione biologica senza le incompatibilità fra comunità ed ecosistema, fra bioma e regione biogeografica, fra biosfera ed ecosfera, e con in più l'esplicito riconoscimento del livello di paesaggio.
Si noti che se il paesaggio fosse ridotto a criterio ambientale, esso servirebbe solo a porre in evidenza la contiguità spaziale degli ecosistemi o delle comunità. Solo con il criterio integrato si può sottolineare la definizione di paesaggio come livello specifico di organizzazione biologica, così come è stato già esposto in precedenza. Da ciò si ricava che l'ecologia si può sviluppare in pieno solo seguendo il criterio integrato.
Il modello di ecotessuto come esempio di ecologia integrata
Lo studio stesso del paesaggio rimane parziale senza un'interpretazione integrata. Si è già accennato al modello a frammentazione e a quello variegato, parlando di struttura del paesaggio, ed è stato presentato il concetto di ecotessuto. Ora è utile ripercorrere lo sviluppo dei modelli interpretativi dell'eterogeneità ecologica nello studio dei territori, per vedere come il criterio integrato sia davvero necessario.
Sia il concetto di comunità che quello di ecosistema sono in sé e per sé dipendenti dalla scala, eppure l'ecologia degli ecosistemi non ha mai considerato tali livelli in rapporto all'eterogeneità di un contesto ecologico su scala superiore: al massimo veniva messo in evidenza (v. figura 3b) un margine fra due diversi ecosistemi o, quando si presentava un gradiente, una fascia ecotonale (Odum, 1971; Whittaker, 1975). Quando si è visto che sarebbe stato un errore trascurare il livello di paesaggio, si è ricorso alla sottolineatura di configurazioni di elementi (ovvero di ecosistemi, intesi in senso integrato come biogeocenosi), nel senso di macchie e corridoi (v. figura 3c) su una matrice paesistica che poteva rimanere anche indistinta (Naveh e Lieberman, 1984; Formane Godron, 1986).
Con il concetto di ecotopo, si è potuti passare compiutamente al modello a frammentazione (v. figura 3d), un modello capace di rappresentare, per una data gamma di scale, un vero e proprio ecomosaico di frammenti (tessere) giustapposti (Haber, 1990, Leser, 1997). Più di recente, diversi studiosi hanno osservato che non tutti gli organismi percepiscono la struttura del paesaggio nello stesso modo: esistono taxa stenotopici ed euritopici, cioè legati ad habitat locali limitati (anche a una sola tessera) oppure ad habitat eterogeni (anche più di due ecotopi). Ne deriva di conseguenza un modello variegato (v. figura 3e, 3f), perché il mosaico ecologico si può intendere composto da tessere a conformazione più o meno variabile o, se si preferisce, come una serie sovrapposta di diversi ecomosaici, a seconda dei livelli scalari di specie evidenziati (Wiens, 1989; Farina, 1997).
Per molte analisi ecologiche sembra che i due modelli (frammentazione e variegato) debbano essere entrambi validi, per esempio quando si studiano le tessere delle biocenosi vegetali di una località e in seguito si rilevano gli indici faunistici a esse correlati. Il ragionamento che ha condotto all'elaborazione di questi due modelli, porta però, se estremizzato, a due paradossi, come evidenziato da T.F.H. Allen e T.W. Hoekstra (1992). Secondo questi autori "se per uno stesso sito coesistono organismi rispondenti a diverse scale di paesaggio, perde senso l'unità organismo come livello scalare di organizzazione biologica; se, al contrario, il paesaggio cambia conformazione strutturale per tipo di organismo, esso diventa spiegabile solo con una curvatura dello spazio". In uno spazio 'curvo', infatti, quale quello invocato nella teoria della relatività, due osservatori ottengono misure diverse sugli stessi oggetti. In sintesi, sembrerebbe che i due livelli di organizzazione biologica individuo e paesaggio non possano coesistere. Ovviamente, nella realtà che trattiamo le cose non stanno così, ma i paradossi aiutano a chiarire le idee, come spesso avviene in matematica con i ragionamenti per assurdo. La questione è molto difficile da risolvere, ma permette di fare alcune considerazioni interessanti. L'organismo rimane senza dubbio un livello di organizzazione biologica, riferito a un proprio insieme di scale, come si può dimostrare con la teoria dei sistemi gerarchici. D'altra parte, il paesaggio non è interpretabile come uno spazio 'curvo', poiché non siamo certamente nelle condizioni richieste dalla relatività per la distorsione dallo spazio!
La teoria della complementarità insegna che la percezione della realtà può essere dipendente dalle modalità di osservazione, ma da ciò non consegue che la realtà sia di per sé non misurabile. Quando sorgono difficoltà nella comprensione di una realtà complessa è quindi necessario arrivare a una descrizione capace di tenere conto della sua molteplicità, anche eventualmente per vie diverse, purché tra loro in relazione. In realtà molte osservazioni relative al modello variegato non sono del tutto veritiere. Una specie euritopica, per esempio una locusta, avrà sicuramente una percezione della grana del mosaico ecologico diversa da quella di una specie stenotopica, poniamo un carabide. La specie euritopica troverà indifferente cibarsi in una macchia a prato come nella zona limitrofa a bosco; tuttavia, le risposte fisiologiche e comportamentali non saranno le stesse indifferentemente, perché il microclima è diverso, la luce cambia, la quantità e la qualità di amminoacidi, glucidi, lipidi e fibre contenuti in media nelle foglie non sono le stesse, certi predatori cambiano, e così via. Esistono di conseguenza delle gerarchie di fattori: alcuni rimangono legati al modello a frammentazione, altri, in gradi diversi, al modello variegato. Inoltre, come già accennato, molte specie animali sono defmibili come permeanti (Odum, 1971), proprio per indicare il fatto che utilizzano abitualmente diversi tipi di ecosistemi. Tali popolazioni permeanti si spostano con ritmi stagionali che arrivano a formare addirittura 'sentieri' rilevabili e usano tessere diverse con un ritmo spesso giornaliero: per abbeverarsi, per ripararsi dal sole, per il pascolo o per la caccia, per l'allevamento della prole. Inoltre, si può osservare che in natura in un certo ecotopo coesistono, con funzioni diverse e spesso interagenti, sia le specie fisse che quelle temporanee (ma non estranee).
Tenendo presente quanto accennato finora, non sembra del tutto corretto contrapporre un mosaico formato da tessere giustapposte (modello a frammentazione) con un mosaico a livelli sovrapposti di tessere diverse (modello variegato). Meglio parlare di tessuto ecologico o ecotessuto (v. figura 3), cioè di un mosaico multidimensionale individuato da un mosaico di base e da una serie gerarchica di mosaici correlati, come già esposto in precedenza.
Chiarito il senso integrato del modello interpretativo di paesaggio, è possibile ora riprendere i sei livelli di organizzazione biologica secondo l'ecologia integrata: organismo, popolazione, biogeocenosi, paesaggio, ecoregione, biogeosfera. Si può osservare che l'insieme dei suddetti livelli organizzativi si presenta più o meno interagente per ogni livello della scala operativa, anche se le espressioni di ognuno di essi e le influenze reciproche cambiano per gamme di scale locali differenti. L'integrazione è senza dubbio una legge imperante in ecologia, ma non procede in senso antigerarchico: le asimmetrie sono maggiori delle simmetrie.
Ciò significa che in determinate risoluzioni di scala locale non ci si può aspettare di trovare certe strutture e certe funzioni proprie di uno o più livelli diversi di organizzazione, mentre in altre risoluzioni è possibile. Perciò, nessun argomento può essere compiutamente studiato a un solo livello di scala, né a un solo livello di organizzazione.
Applicazioni dell'ecologia del paesaggio
Approccio transdisciplinare
Per trattare delle applicazioni dell'ecologia del paesaggio e delle metodologie relative, è indispensabile tenere conto del ruolo gestionale svolto dall 'uomo nei confronti della natura. Tale ruolo è derivato dalla constatazione che gli ecotopi umani e quelli naturali sono integrati a livello di paesaggio nei mosaici complessi rilevabili come ecotessuti. Ne derivano nuovi paradigmi scientifici e culturali, perché l'ecologia del paesaggio si dimostra indispensabile per un approccio transdisciplinare e per tutti i nuovi criteri metodo logici di applicazione dell'ecologia: per il ripristino, la rinaturazione, il risanamento, la progettazione dell'ambiente, e (soprattutto) per la conservazione biologica e la pianificazione ecologica del territorio.
L'approccio transdisciplinare nasce dall'osservazione che le discipline interessate allo studio dell'ambiente sono tante, ma, come già accennato, non rivestono tutte la stessa importanza. Il principio organizzativo dei vari livelli in cui si manifestano le forme di vita resta principalmente biologico, e solo esso può fungere da integratore. Popper sottolineava che la natura stessa ha creato, attraverso il linguaggio e attraverso la possibilità di formulare le nostre teorie in forma linguistica, una nuova dimensione che riconcilia natura e cultura. Si osservi che l'interdisciplinarità non è più sufficiente per affrontare lo studio dei sistemi adattativi complessi, perché essa è legata ancora a basi riduzioniste e meccanicistiche di divisione per settori chiusi e ricomposizione lineare per somma. È necessario invece seguire una metodologia gerarchica a rete spaziale aperta, su basi in prevalenza olistiche, con principi integratori raggiungibili da ogni disciplina concorrente; in altre parole, una metodologia transdisciplinare.

Come ha più volte affermato Z. Naveh (Naveh e Lieberman, 1990), una disciplina che gode di tali caratteristiche è appunto l'ecologia, quando però essa abbia acquisito i principi dell'ecologia del paesaggio, ovvero ciò che noi chiamiamo più opportunamente ecologia integrata. Si può notare inoltre che l'ecologia del paesaggio riesce a tradurre le informazioni provenienti da altre discipline in dati integrabili con i principi ecologici (Ingegnoli, 1991; 1993). La nuova metodologia ecologica dovrà essere in qualche modo analoga a quella medica: le valutazioni ambientali dipenderanno dal confronto fra le condizioni del sistema ecologico in esame e quelle di uno stato considerato come 'normale'. In altre parole, sarà il rapporto tra 'patologia' ed 'ecologia' dei sistemi che permetterà una diagnosi in senso clinico del paesaggio da studiare o pianificare (fig. 5).
Da quanto detto poc'anzi, è possibile rendersi conto che lo stesso concetto di ambiente viene ad assumere una nozione di unitarietà in precedenza impossibile per l'assenza di un principio integratore, quale, per esempio, l'integrabilità fra uomo e natura. Ne seguono nuove interpretazioni, talvolta sorprendenti, dei principali problemi ambientali, visto che i concetti di ecosistema e di paesaggio risultano assai diversi da quelli finora formalmente utilizzati nella gestione e nel controllo dell'ambiente. I concetti di danno per la salute, di valutazione ambientale, di pianificazione territoriale, e di conservazione biologica vengono decisamente modificati.
Danno per la salute
La salvaguardia della salute è oggi principalmente basata sul controllo dell'inquinamento dell'aria, delle acque e del suolo. Non viene invece preso in considerazione alcun danno dovuto alle disfunzioni strutturali e spaziali dei sistemi ecologici, disfunzioni sicuramente gravi e ben più difficili da correggere (Ingegnoli, 1993). Si pensi, per esempio, che funzioni anche molto elevate del comportamento umano sono controllate a livello genetico e fisiologico e possono essere influenzate dalle condizioni dell'ambiente, soprattutto nei primi anni di vita dell'individuo. Lo sviluppo delle sinapsi e delle reti dei neuroni della corteccia cerebrale dell 'uomo si completa solo intorno ai due anni e mezzo di vita, proprio per raggiungere un adeguato rapporto interattivo con l'ambiente. Se l'ambiente però si degrada anche solo nelle sue componenti spaziali e strutturali (senza inquinamenti), i danni potrebbero essere di notevole gravità e colpire non solo la psiche ma anche il sistema ormonale. Le conseguenze si aggravano, soprattutto quando si aggiunge un ambiente familiare (e sociale) che non offre stimoli adeguati per lo sviluppo. Inoltre, nell'ambito della patologia dei sistemi di ecosistemi, le sindromi dovute agli inquinamenti sono malgrado tutto le più curabili, anche se per certi versi le più immediatamente nocive per la salute. In tal senso va data più forza ai controlli sullo stato dell'ambiente secondo i principi dell'ecologia del paesaggio, anche per la protezione della salute.
Valutazione ambientale
Come è noto, la valutazione ambientai e è attualmente basata sul concetto di impatto, derivato da una contrapposizione preconcetta fra l'uomo e la natura. Il concetto di impatto è quindi in contraddizione con l'ecologia integrata. La minimizzazione dei disturbi sarebbe da riservare solo per quelli fuori scala, dato il ruolo strutturante degli stessi nei sistemi ecologici e i processi di incorporazione in gioco. l regolamenti della Valutazione di Impatto Ambientale (VIA) sono inoltre limitati a poche categorie di opere (la pianificazione ne è esclusa!), e il concetto di paesaggio non è inteso in senso ecologico (d.p.c.m. del 27 dico 1988, par. l). Manca ogni riferimento esplicito all'erfolgskontrolle, cioè al controllo del successo, dopo che l'operazione di trasformazione ambientale è stata ultimata; mancano riferimenti metodologici legati a un criterio clinico-diagnostico, unico criterio valido per valutare lo stato di un sistema biologico. La valutazione ambientale è evidentemente assai importante, essendo necessaria sia nella progettazione e nella pianificazione ambientale sia nei controlli. Alla luce dei principi di ecologia del paesaggio, gli odierni criteri devono cambiare.
Pianificazione territoriale
Il principale problema che la pianificazione territoriale deve oggi affrontare dipende da una predominanza ancora pesante dei metodi e dei concetti urbanistici, anche se gli esperti di ecologia vengono sempre più spesso consultati dai pianificatori in fase di analisi di un piano. Tuttavia le scelte politico-economiche continuano a guidare la pianificazione, le suddivisioni amministrative sono indifferenti agli ambiti ecologici, il senso estetico-percettivo rimane la principale chiave di lettura del paesaggio e, infine, le infrastrutture tecnologiche dominano ogni esigenza di funzionalità biologica del territorio. Oggi in molti paesi, fra cui l'Italia, la pianificazione è quasi totalmente in mano agli architetti e agli ingegneri, che spesso non hanno conoscenze approfondite né sufficiente sensibilità per le componenti biologiche del paesaggio che, lo ricordiamo, non sono solo quelle naturali, ma anche quelle antropiche re interpretate in chiave biologica. Invece sarebbe necessario invertire la gerarchia dei valori decisionali, dando priorità alle componenti di pianificazione ecologico-paesaggistiche. Il passaggio dal concetto di territorio come mero supporto geografico-economico alle attività umane, al concetto di paesaggio come entità biologica alla quale l'uomo deve adeguarsi, porta grandi sconvolgimenti nei metodi stessi di pianificazione, oltre che negli obiettivi. Per questo, molti studiosi oggi cercano di cambiare metodologia, utilizzando sempre più spesso strumenti matematici più moderni, quali per esempio i sistemi informativi geografici (GlS, Geographical lnformation Systems) e l'analisi statistica multivariata. Ma ciò non è sufficiente se i principi sottesi a tali cambiamenti non sono riferiti a un'ecologia più avanzata e più consona allo studio dei sistemi territoriali. Per esempio, se non si considera il cambiamento ecologico dei comportamenti dinamici, nel passaggio di scala da un singolo ecosistema a un intero paesaggio si possono anche trarre errate conclusioni, malgrado l'uso dei più avanzati strumenti di formalizzazione matematica e di informatica.

Mancano inoltre gli operatori professionali preparati per coordinare una pianificazione avanzata. D'altra parte va sottolineato che oggi assistiamo a una svolta epocale: la maggioranza delle popolazioni umane non abita più nelle campagne, ma nelle aree urbane e nelle città. La crescita di megacittà e di vasti paesaggi suburbani (tab. 3) che, secondo stime attendibili, arriveranno nel 2025 ad assorbire circa due terzi della popolazione dell'intera bio sfera può portare a sconvolgimenti ambientali enormi quali inquinamento, epidemie, frammentazione spinta di aree naturali, aumento della temperatura e della piovosità, abbandono di coltivi, distruzione di corridoi ecologici e così via. Va ricordato che la degradazione degli habitat, sia naturali che umani, rimane la prima causa di estinzione delle specie. Eppure sembra che la nostra società non si accorga ancora di queste problematiche e l'area metropolitana di Milano, a cui manca una struttura amministrativa adeguata al paesaggio urbano, ne è un esempio. Basta sorvolare il territorio fra i laghi e il Po per vedere che la città comprende oggi almeno 50 comuni, con un'estensione di decine di chilometri. La perdita di popolazione di Milano a partire dagli anni Ottanta del 20° secolo è un processo che sembra andare contro quanto detto sull'aumento dell 'urbanizzazione a livello mondiale, ma ciò è solo apparente. In realtà Milano continua, sia pure lentamente, ad aumentare la sua popolazione; per accorgersene, basta considerare il fenomeno urbano nei suoi giusti confrni. Come si può notare dalla tabella (tab. 4), la città ha perso circa 400.000 abitanti, ma la sua provincia ha aumentato del 28,6% la densità di ab/km²; il minor numero di abitanti è dovuto al distacco del territorio di Lodi. Tutto ciò indica piuttosto il completamento dell'urbanizzazione dell'intera provincia, la quale peraltro è meno estesa della conurbazione milanese, che comprende anche la parte sud delle province di Varese e Como, e supera sicuramente i 4.000.000 di abitanti. Milano è ormai diventata la city di una metropoli fra le maggiori di Europa, ma non riesce ad avere adeguate strutture e servizi anche perché non viene riconosciuto il fenomeno urbano come fenomeno di trasformazione ecologica del territorio.
Da quanto detto sopra segue che la futura pianificazione urbanistica dovrà necessariamente essere preceduta da quella ecologica, ovvero da un indispensabile controllo dello stato del territorio che potrà fornire una base seria alle scelte amministrative. Tale pianificazione dovrà essere: elaborata su diverse scale spazio-temporali; riferita a unità di paesaggio e non a singole aree comunali; capace di aggregare le macchie isolate; volta a creare poche macchie di grandi dimensioni che ospitino una vegetazione naturale; capace di mantenere una rete di corridoi naturali che proteggano i corsi d'acqua e che garantiscano connettività per i movimenti di specie chiave tra le macchie di grandi dimensioni; tesa a progettare piccole macchie e corridoi che provvedano ad assicurare una elevata eterogeneità ambientale; infine, capace di conservare la sua traiettoria storica, dato che ogni trasformazione che non proceda in congruenza rispetto alla storia del paesaggio apporta profonde alterazioni all'ecotessuto.
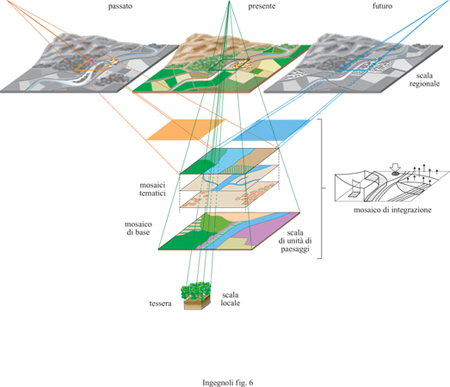

Mediante l'analisi e la diagnosi basate sull'ecologia del paesaggio, le strutture di carattere naturalistico potranno essere integrate con quelle di carattere antropico, ma dovrà trattarsi di una integrazione di reti, anzi di apparati paesaggistici con molteplicità funzionale, non di una zonizzazione di tessere con rigidità di vincoli. Oggi la conservazione biologica è fatta esclusivamente per macchie isolate residuali; poco importa che si tratti di un biotopo locale o di un grande parco, è solo questione di scala. Invece, con il paradigma dell'ecologia del paesaggio, la conservazione si articola per reti di ecotopi in un contesto di mosaici complessi, e tiene conto della complementarità degli ecosistemi umani anche in regioni molto popolate come le nostre, dove per millenni si è svolto un processo di coevoluzione dei due tipi di ecosisterni. Diventa quindi centrale lo studio degli ecotessuti, che si effettua attraverso tre integrazioni: la prima sottolinea i legami fra un insieme di scale che vanno da quella regionale a quella locale; la seconda sottolinea la scala di unità di paesaggio (o distretto) composta da un insieme di ecomosaici tematici; la terza riguarda l'aspetto temporale (fig. 6). Le analisi di ecologia del paesaggio necessitano in genere di un'interpretazione preliminare dell'ecotessuto, del tipo di quella offerta nella tabella (tab. 5).
Oggi, generalmente, gli ecologi lavorano su diverse scale, e il concetto di ecotessuto li costringe a utilizzare nuovi appropriati indicatori. Per questo occorre aggiungere almeno quattro criteri, sempre derivati dall'ecologia del paesaggio: la definizione di capacità biologica del territorio o biopotenzialità territoriale, la definizione di apparati paesistici, la definizione di habitat standard, la teoria source and sink.
Conservazione biologica
I biotopi, come per esempio i parchi e le riserve, sono tutelati come isole sacre, e ciò di fatto fornisce l'alibi per il degrado dell'intero paesaggio circostante di cui essi invece dovrebbero essere parte integrante. La distruzione degli habitat (e non solo quelli estranei all'uomo) senza dubbio è il primo dei problemi della distruzione della biodiversità e viene troppo sovente messo in secondo piano dalle questioni relative all'inquinamento.
D'altra parte, disturbi e frammentazione dell'ambiente vengono visti solo come fattori negativi se dovuti all'azione umana, dimenticando il ruolo strutturante dei disturbi (anche antropici) quando non fuori scala. È necessario quindi sviluppare nuovi concetti di biodiversità e di naturalità, e disporre di principi di ecologia integrata capaci di controllare la dinamica dei mosaici ecologici su varie scale. È necessario inoltre non cristallizzarsi solo sulle specie in via di estinzione, ma su tutte le alterazioni dei livelli dello 'spettro biologico'. Per sottolineare come il rapporto fra ecologia del paesaggio e conservazione biologica possa cambiare le vecchie concezioni, è necessario precisare meglio i seguenti punti basilari: biodiversità, naturalità, e criteri di conservazione (Ingegnoli e Massa, in corso di stampa).
Biodiversità. - Considerata l'importanza concettuale degli episodi di estinzione di massa, il concetto di biodiversità rimane quasi esclusivamente riferito alle specie. Il tentativo di estensione del concetto di biodiversità a comunità e habitat, cioè il riferimento alle diversità a, α, β e γ di R.R. Whittaker (1975), sembra tuttavia limitato e marginale. Inoltre si continua a identificare l'aumento di biodiversità con l'aumento di stabilità dei sistemi ecologici, indipendentemente dalla scala spazio-temporale.
Questo assunto, e le sue conseguenze, rappresentano per l'ecologia integrata una distorsione della realtà. È quindi necessario sottolineare l'importanza della diversità biologica a ogni livello di organizzazione, dal patrimonio genetico alle specie, dalle popolazioni ai paesaggi e alla bio sfera. È importante ricordare che l'evoluzione dei sistemi complessi adattativi comporta fasi ricche di componenti ridondanti, ma anche fasi in cui l'aumento di ordine e organizzazione dei sistemi viventi esige una diminuzione di tale ridondanza. Per esempio, l'evoluzione dalla macchia mediterranea alla foresta di lecci porta a una diminuzione di biodiversità di quasi 1'80% (Pignatti, 1994).
Secondo Prigogine, l'ordine attraverso le fluttuazioni (cioè i disturbi) sembra essere uno dei pochi processi generali del nostro Universo. Vanno dunque enfatizzate le biodiversità ecologiche e non la biodiversità specifica, e va ricordato che la questione di base non è quella della conservazione in sé, ma nel processo che implica flussi e fasi di biodiversità, cioè nella capacità creativa ed evolutiva della vita. Anche se a livello non di singoli individui, ma di specie, di comunità e di paesaggi, la morte può essere necessaria: però, come notano M.E. Soulé e B.A. Wilcox (1980), una cosa è la morte, ben altra è la fine delle nascite!
Naturalità. - Nel vocabolario della lingua italiana dell'Istituto dell'Enciclopedia ltaliana, la naturalità è definita come "qualità, condizione di ciò che è naturale", ma dai conservazionisti è intesa di solito come assenza di antropicità: ancora una contrapposizione preconcetta fra uomo e natura. In tal modo si dimentica che il ruolo dell 'uomo nella natura è principalmente gestionale, dato che egli possiede molti degli attributi propri del processo che l 'ha creato, come sostengono Lorenz e Popper (1985): "L'umana sensibilità ai valori corrisponde all'evento evolutivo in quanto anche nel nostro cervello ha luogo un giuoco di tutto con tutto come nel mondo esterno. [ ... ] il modello della soluzione razionale dei problemi resta lo stesso in tutti i campi: la combinazione di creatività e critica. È essenzialmente un modello evoluzionistico".
Fra le rivoluzioni che si sono verificate nella storia della vita si annoverano la comparsa dei procarioti, quella degli eucarioti, lo sviluppo degli organismi pluricellulari, lo sviluppo dei vertebrati superiori e quello di una specie (l'uomo) potenzialmente capace di comprendere il processo evolutivo. L'uomo sta arrivando oggi a una fase del suo sviluppo nella quale egli dovrebbe rendersi consapevole di tutto ciò. È necessario quindi un concetto nuovo di naturalità diffusa che, secondo V. Giacomini (1965), implica un riequilibrio secondo le leggi naturali fino ad arrivare agli habitat umani. E non basta ancora. Il concetto odierno di naturalità deve abbandonare ogni riferimento 'cartesiano', nel quale la natura è meccanicistica, poiché non viene considerata in prospettiva storica e può essere dominata dall'uomo che se ne fa ingegnere, contrapponendovisi. La realtà naturale è invece una realtà evolutiva, storica, in cui ordine e disordine giocano un eguale ruolo: la natura non è determinata una volta per tutte, perché aperta a prospettive creative imprevedibili.
Quindi la naturalità è la consistenza prevalente della natura, derivata dal concetto di wilderness secondo H.D. Thoreau (1849) per il quale "nella naturalità è la preservazione del mondo" o di J. Muir, il quale scriveva che "andare in montagna è come tornare a casa; la naturalità è una necessità; i parchi e le riserve di montagna sono utili non solo come fonti di legname e acque di irrigazione, ma come fonti di vita". Le nostre Alpi, laddove mostrano ancora valli non sconvolte dal consumismo, e in cui ancora si possono trovare grandi predatori e specie rare, ampi paesaggi di foreste e praterie, sono fra gli esempi di una naturalità che è capace di comprendere anche l'uomo.
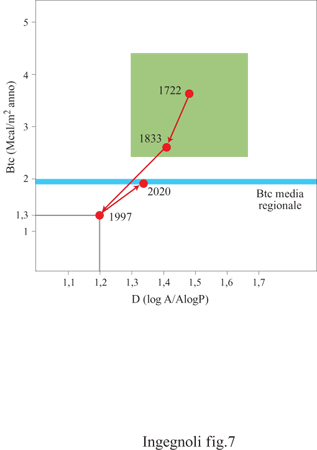
Criteri di conservazione. - Nei criteri di pianificazione a scopo conservativo è sicuramente più opportuno salvaguardare la possibilità di radiazione delle specie piuttosto che focalizzare l'attenzione su singole specie rare e magari endemiche. È bene ricordare inoltre il paradosso della gestione nella conservazione della natura (Forman e Collinge, 1996): la probabilità di avere un effetto immediato è maggiore nelle piccole aree, ma la probabilità di successo a lungo termine è maggiore nelle grandi aree. Ne consegue che i nostri sforzi di conservazione sono da compiere su scale decisamente più ampie di quanto in genere si usa fare, fino ad arrivare a comprendere i livelli di paesaggio e di regione. Oltre a essere elaborati su diverse scale, è necessario però che gli studi siano riferiti a unità di paesaggio, non a confmi amministrativi. La metodologia deve mirare innanzitutto a verificare che le zone riservate a parco naturale siano funzionalmente tali nel contesto del sistema paesistico regionale (fig. 7), e il processo di analisi ecologica dovrebbe svolgersi in almeno tre fasi principali: l) fase degli studi diagnostici: disturbi, stato della vegetazione, rapporto habitat umano/habitat naturale, tipo di trasformazione dell'ecotessuto, variazione delle frammentazioni, rete ecotonale, source and sink, specie chiave, problemi genetici, ecc; 2) fase delle proposte di intervento terapeutico: poche larghe macchie, corridoi di vegetazione lungo i corsi d'acqua, distribuzione di biotopi nella matrice, connettività, punti strategici, aree di nuovo sviluppo, rinaturazione, ingegneria naturalistica, ecc; 3) fase dei controlli ecologici e progettazione fmale: controlli sulle trasformazioni dell'ecotessuto, verifica delle nicchie di rigenerazione, incorporabilità dei disturbi, aree di interferenza fra le reti naturali e antropiche, monitoraggi, ecc.
Per concludere, bisogna ricordare che a tale metodologia è sempre necessario affiancare concetti più generali di pianificazione del territorio. Ciò per motivi di integrazione con gli altri obiettivi di tutela, quali per esempio la conservazione delle preesistenti testimonianze storico-architettoniche e il loro ambientamento, o le esigenze di salvaguardia e sostenibilità economico-sociale di popolazioni locali. L'integrità ecologica e le esigenze umane di base devono infatti svilupparsi in una reciproca armonia resa possibile da riequilibri compensativi, da opere di ingegneria naturalistica, da adeguamenti costruttivi di infrastrutture e insediamenti, sempre retti dai principi ecologici.
Ringraziamenti
Desidero ringraziare Ze Naveh e Richard Forman per l'incoraggiamento scientifico, Hirgen Ott per le osservazioni critiche al manoscritto originario e infine Elena Giglio per l'aiuto nella revisione del testo.
Bibliografia citata
ALLEN, T.P.H., HOEKSTRA, T.W. (1992) Toward a unified ecology. New York, Columbia University Press.
BLONDEL, J. (1986) Biogéographie évolutive. Parigi, Masson.
DOKUCAEV, V.V. Writings, voI. 6. Mosca, Akad. Nauk, 1951.
ERMER, K, HOFF, R., MOHRMANN, R. (1996) Landschaftsplanung in der Stadt. Stoccarda, Ulmer.
FARINA, A. (1997) Principles and methods in landscape ecology. Londra, Chapman & Hall.
FORMAN, R.T.T., COLLINGE, S.K (1996) The spatial solution to conserving biodiversity in landscapes and regions. In Conservation of faunal diversity in forested landscapes, a c. di DeGraaf R.M., Miller R.I., Londra, Chapman & Hall.
FORMAN, R.T.T., GODRON, M. (1986) Landscape ecology. New York, Wiley.
FORMAN, R.T.T., MOORE, P.N. (1992) Theoretical foundations for understanding boundaries in landscapes mosaics. In Landscapes boundaries. Consequences for biotic diversity and ecological flows, a c. di Hansen A.J., di Castri F., New York, Springer-Verlag.
GELL-MANN, M. (1994) The quark and thejaguar. Adventures in the simple and the complex. New York, Freeman & C. GIACOMINI, V. (1965) Significato e funzione dei parchi nazionali. In I parchi nazionali in Italia. Roma, Istituto di Tecnica e Propaganda.
GODRON, M. (1984) Écologie de la végétation terrestre. Parigi, Masson.
HABER, W. (1990) Basic concept oflandscape ecology in planning and management. Phys. Ecol. Jpn., 27, 131-146.
INGEGNOLI, V. (1971) Ecologia territoriale e progettazione: significati e metodologia. In L'ingegnere di fronte alla sopravvivenza umana. Coli. Ing. voI. I, pp. 398-400.
INGEGNOLI, V. (1991) Human influences in landscape change: thresholds of metastability. In Terrestrial and aquatic ecosystems: perturbation and recovery, a c. di Ravera O., New York, Ellis Horwood, p. 303-309.
INGEGNOLI, V. (1993) Fondamenti di ecologia del paesaggio. Milano, Cittastudi.
INGEGNOLI, V., a c. di (1997) Esercizi di ecologia del paesaggio. Milano, Cittastudi.
LESER, H. (1997) Landschaftsoekologie. Stoccarda, Ulmer.
LORENZ, K. (1978) Vergleichende Verhaltensforschung: Grundlagen der Ethologie. Vieuna, Springer. LORENZ, K, POPPER, K. (1985) Il futuro è aperto. Milano, Rusconi.
MANDELBROT, B.B. (1975) Les objects fractals: forme, hasard et dimension. Parigi, Flammarion.
MARIANO, C. (1958) Nuovo dizionario italiano-latino. Milano, Società Editrice Dante Alighieri.
MOEBIUS, K. (1877) Die Auster un die Austernwirtschaft. Berlino.
NAVEH, Z., LIEBERMAN, A. (1984) Landscape ecology. Theory and application. New York, Springer-Verlag. NA VEH, Z., LIEBERMAN, A. (1990) Landscape ecology: theory and application, 2a ed., New York, Springer-Verlag.
ODUM, E.P. (1971) Fundamentals of ecology, 3a ed., Filadelfia, Saunders.
ODUM, E.P. (1993) Ecology and our endangered life-support systems. Sunderland, Sinauer Ass. Inc. Pub.
OLDEMAN, R.A.A. (1990) Forests: elements of sylvology. New York-Berlino, Springer.
O'NEILL, R.V., DE ANGELIS, D.L., WAIDE, J.B., ALLEN, T.F.H. (1986) A hierarchical concept of ecosystems. Princeton, Princeton University Press.
PIGNATTI, S. (1994) Ecologia del paesaggio. Torino, UTET.
POINCARÉ, J.-H. (1906) La science et l'hypothèse. Parigi, Flammanon.
POPPER, K (1996) Tutta la vita è risolvere problemi: scritti sulla conoscenza, la storia e la politica. Milano, Rusconi.
PRIGOGINE, I. (1996) La fin des certitudes. Temps, chaos et lois de la nature. Parigi, Odile Jacob (trad.it. Lafine delle certezze. Il tempo, il caos e le leggi della natura. Torino, Bollati-Boringhieri, 1997).
PRIGOGINE, I., NICOLIS, G. (1977) Self-organization in non-equilibrium systems: from dissipative structures to order through jluctations. New York, Wiley (trad. it. Auto-organizzazione dei sistemi termodinamici in non-equilibrio. Firenze, Sansoni, 1982).
PRIGOGINE, I., NICOLIS, G., BABLOYATZ, A. (1972) Thermodinamics of evolution. Physiscs Today, 25, 23-28.
SOULÉ, M.E., WILCOX, B.A. (1980) Conservation biology. Its scope and its challenge. In Conservation biology. An evolutionary-ecological perspective, a c. di Soulé M.E., Wilcox B.A., Sunderland, Sinauer Ass. Inc. Pub.
THOREAU, H.D. (1849) A week on the Concord and Merrimack riverso Boston, James Munroe and Co. Ristampa Frauklin Center, Frauklin Library, 1983.
TROLL, K. (1939) Luftbildenplan und ökologische Bodenforschung. Berlino, Zeitschrift & Gesellschaft Erdkunde.
TURNER, M.G., GARDNER, R.H. (1990) Quantitative methods in landscape ecology. New York-Berlino, Springer.
WHITTAKER, R.H. (1975) Communities and ecosystems. New York, Macmillan.
WIENS, J.A. (1998) Spatial scaling in ecology. Funct. Ecol., 3, 385-397.
ZANZI, L. (1995) Viatico per una avventura nella storia della Val Grande, In Val Grande, storia di una foresta, a c. di Alberti L., Rainaldi G., Rizzi E., Anzola d'Ossola, Fondazione Arch. Enrico Monti.
Bibliografia generale
COLUMELLA, L.J. De re rustica. Cambridge, Harvard University Press, 1977.
FORMAN, R.T.T., GODRON, M. Landscape ecology. New York, Wiley, 1986.
LESER, H. Landschaftsoekologie. Stoccarda, Ulmer, 1997.
NAVEH, Z., LIEBERMAN, A. Landscape ecology. Theory and application. 3a ed., New York, Springer-Verlag, 1990.
TURNER, M.G., GARDNER, R.H. Quantitative methods in landscape ecology. New York-Berlino, Springer, 1990.
ZONNEVELD, I.S. Land ecology: an introduction to landscape ecology as a base for land evaluation, land management and conservation, Amsterdam, SPB Academic Publishing, 1995.