
Biodiversita
Biodiversità
C'è il rischio di una nuova
estinzione di massa?
Un pericolo per il futuro della Terra
di W. Wayt Gibbs
26 agosto-4 settembre
Si tiene a Johannesburg, in Sudafrica, il Vertice Mondiale sullo Sviluppo sostenibile. Il dibattito verte sulle strategie da adottare per assicurare alla Terra un futuro che consenta alle popolazioni di soddisfare i propri bisogni senza danneggiare l'ambiente. Fra i cinque temi chiave posti all'attenzione internazionale - acqua e igiene, energia, salute, produttività agricola e biodiversità - l'ultimo assume rilievo in considerazione della minaccia rappresentata per l'equilibrio del pianeta dall'aumento delle specie animali e vegetali in via di estinzione, tale da fare parlare di rischio di una prossima estinzione di massa.
Un allarme giustificato?
Gli allarmi degli ecologisti sul rischio di una prossima estinzione di massa sono contestati dagli scettici e spesso ignorati dai politici. Almeno in parte, queste reazioni sono la conseguenza della straordinaria difficoltà che gli stessi studiosi trovano a stabilire le reali dimensioni del fenomeno della scomparsa delle specie, ad appurarne le cause e, tanto più, a identificare i modi per porvi rimedio.
Nella conferenza tenuta nell'agosto 2001 in occasione del congresso annuale della Society for Conservation Biology, Sir Robert M. May ha espresso il senso di profondo sconforto che pervade attualmente chi si occupa di conservazione della biodiversità. Secondo la sua ultima stima, dichiaratamente approssimativa, il tasso di estinzione, ovvero la rapidità con cui le specie spariscono, si è accelerato nel corso degli ultimi cento anni, fino a raggiungere un valore di circa 1000 volte superiore a quello precedente la comparsa dell'uomo sulla Terra. Vari elementi fanno inoltre prevedere che il tasso sia destinato a decuplicarsi nei prossimi cento anni. Edward O. Wilson, professore di zoologia alla Harvard University, e molti altri eminenti ecologisti concordano con May nel ritenere che ci troviamo alla vigilia della sesta grande estinzione di massa nella storia del pianeta.
Fino a che punto sono giustificate tali fosche previsioni? Intorno a questa domanda si è acceso un aspro dibattito. In The skeptical environmentalist, un controverso volume pubblicato nel 2001 da Cambridge University Press, lo statistico danese Bjørn Lomborg sostiene che le cifre sull'imminente scomparsa della biodiversità sono state fortemente esagerate. Di fronte a questo scetticismo, all'incertezza degli stessi ricercatori e all'apatia dell'opinione pubblica, molti scienziati hanno iniziato a contestare l'enfasi eccessiva posta da alcuni gruppi ambientalisti sulla conservazione delle specie rare e delle nicchie ecologiche in cui esse vivono. Forse, sostengono, dovremmo piuttosto concentrare la nostra attenzione sul salvataggio di un elemento ugualmente a rischio, ma ancora più prezioso: l'evoluzione stessa.
May sostiene che gli esseri umani starebbero causando l'estinzione di massa più imponente dopo quella che portò alla scomparsa dei dinosauri, 65 milioni di anni fa. Questa affermazione, che può apparire probabilmente sconcertante a quanti non hanno seguito da vicino l'evoluzione del dibattito sulla biodiversità, non coglie certo impreparati i biologi conservazionisti. Da un sondaggio del 1998 risulta infatti che il 70% dei biologi è convinto che un processo di estinzione di massa sia già in corso e che nei prossimi trent'anni un quinto delle specie attuali sia destinato a scomparire.
Si deve tuttavia osservare che le stime sul tasso di estinzione sono state notevolmente ridimensionate negli ultimi anni. In The sinking ark, pubblicato nel 1979, Norman Myers valutava che ogni anno pervenissero all'estinzione 40.000 specie e che entro il 2000 il numero di specie estinte avrebbe superato il milione. Negli anni Ottanta Thomas Lovejoy prevedeva la scomparsa di una percentuale variabile dal 15 al 20 di tutte le specie allora presenti entro il 2000 e Paul Ehrlich, biologo alla Stanford University, arrivava addirittura al 50%. Per fortuna, queste prime previsioni si sono rivelate eccessivamente pessimistiche. Si valuta attualmente che almeno 1100 specie siano scomparse durante gli ultimi 500 anni. Anche se il numero effettivo di estinzioni potrebbe essere notevolmente superiore a questo, esso è certamente molto lontano dal milione.
Le proiezioni più recenti indicano un lieve rallentamento di questo processo, dato che certe specie considerate in via di estinzione hanno resistito più a lungo del previsto e alcune sono perfino, per così dire, 'ritornate dalla tomba': nel 2001, per es., i biologi hanno scoperto l'esistenza di alcune popolazioni superstiti di un piccolo roditore, il microto bavarese, che era considerato l'unico mammifero eurasiatico estinto negli ultimi 500 anni. I tassi di estinzione a cui si fa riferimento nella recente letteratura scientifica sono quelli resi noti da Wilson, secondo il quale in ogni decennio si estinguerebbe dall'1 al 10% delle specie, ossia almeno 27.000 specie ogni anno. In The skeptical environmentalist, tuttavia, Lomborg sostiene che queste stime non sono suffragate da prove sufficienti e cita altri modelli che prevedono un tasso di estinzione dello 0,15% ogni dieci anni.
Gli scienziati si chiedono dunque se il fenomeno dell'estinzione di massa si debba considerare veramente già in atto. Per rispondere a questa domanda, è necessario conoscere tre dati: il tasso di estinzione naturale (o di fondo), il tasso di estinzione corrente e, infine, la velocità di estinzione, che può essere costante o soggetta a cambiamenti.
Tasso di estinzione naturale, tasso di estinzione corrente e velocità di estinzione
Per calcolare il tasso di estinzione naturale, Wilson e May hanno considerato le stime della durata dell'esistenza di diverse specie elaborate dai paleontologi e ne hanno ricavato una media; il tasso di estinzione naturale corrisponde all'inverso di questa durata media. Se le specie hanno origine in modo casuale e hanno tutte un'esistenza media di circa un milione di anni, ne consegue, secondo Wilson, che ogni anno si estinguerà per cause naturali una specie su un milione. In un articolo pubblicato nel 1995, che viene tuttora citato nella maggior parte dei lavori su questo argomento, May si serviva di un metodo analogo per calcolare il tasso di estinzione naturale, basandosi però su stime secondo le quali la durata media dell'esistenza di una specie si collocherebbe tra cinque e dieci milioni di anni. Si giungerebbe così a un tasso da cinque a dieci volte inferiore a quello proposto successivamente da Wilson e dallo stesso May. Secondo il paleontologo David M. Raup, che ha pubblicato alcuni dei dati di cui si sono serviti Wilson e May, i loro calcoli sarebbero tuttavia seriamente inficiati da tre falsi presupposti. Il primo è che tutte le specie, siano esse appartenenti a piante, mammiferi, insetti, invertebrati marini o ad altri gruppi, abbiano una durata di esistenza sulla Terra molto simile tra loro. Di fatto, il tempo di sopravvivenza può variare tra i diversi gruppi di oltre dieci volte. Poiché le specie di mammiferi, per es., sono tra le meno longeve, il loro tasso di estinzione naturale risulta molto più elevato della media. In secondo luogo, si presume che tutti gli organismi abbiano le stesse possibilità di essere rappresentati nella documentazione fossile, ma in realtà i paleontologi valutano che meno del 4% delle specie esistite si siano conservate allo stato fossile. I fossili più comuni appartengono naturalmente alle specie più ampiamente diffuse, cioè a quelle che hanno avuto maggiore successo. Le specie rare, per es. quelle confinate su un'altura o su un'isola, sono anche più deboli e tendono a estinguersi prima di potersi fossilizzare. Il terzo problema è di ordine statistico: May e Wilson si servono di una durata media, mentre avrebbero dovuto utilizzarne una mediana. Dato che la grande maggioranza delle specie ha avuto un'esistenza breve, la media è distorta dalle poche specie che hanno avuto una durata molto lunga. Tutte e tre queste semplificazioni portano a una sottovalutazione del tasso di estinzione naturale, facendo apparire di conseguenza la situazione attuale più grave di quanto in effetti non sia.
Nel 2001, la biomatematica Helen M. Regan e molti suoi colleghi hanno tentato di correggere queste incertezze, prendendo in considerazione esclusivamente i mammiferi, il gruppo studiato più approfonditamente. Essi hanno valutato quanti dei mammiferi attualmente esistenti e quanti di quelli estinti da poco tempo potrebbero in futuro comparire tra i fossili. Inoltre, non limitandosi a fornire le stime più plausibili, hanno quantificato il margine di incertezza delle diverse cifre. Il risultato di queste ricerche ha evidenziato che il tasso di estinzione attuale, o corrente, è da 17 a 377 volte più elevato di quello naturale; l'ipotesi più probabile è quella che vede l'aumento del tasso compreso tra 36 e 78 volte.
Tuttavia, anche questo nuovo metodo non è del tutto privo di difetti. Confrontare gli ultimi 400 anni con i 65 milioni di anni precedenti equivale a sostenere che il tasso di estinzione corrente rimarrà invariato per milioni di anni, il che appare piuttosto improbabile. Il paleontologo John Alroy ha recentemente proposto un metodo per misurare la velocità di estinzione che non risente di tale presupposto e ha dedotto che negli ultimi 200 anni il tasso di estinzione dei mammiferi è stato circa 120 volte più elevato di quello naturale.
I tentativi di quantificare il tasso di estinzione corrente si sono scontrati con difficoltà, se possibile, ancora maggiori. L'organizzazione internazionale IUCN (International union for conservation of nature and natural resources) redige una Lista Rossa degli organismi ritenuti estinti, ma i metodi di cui si serve sono considerati da alcuni scienziati non sufficientemente rigorosi. Un gruppo di esperti dell'American museum of natural history ha dato vita a un Comitato sugli organismi recentemente estinti, con il compito di rivedere la Lista Rossa per identificare le specie decisamente uniche e di cui non si è trovata più traccia in natura o nei giardini zoologici dopo una ricerca ragionevolmente lunga. Il Comitato ha confermato l'estinzione di 60 delle 87 specie di mammiferi presenti negli elenchi dell'IUCN, ma solo 33 delle 92 specie di pesci ritenute estinte dalla stessa organizzazione lo sarebbero veramente.
Per ogni specie erroneamente ritenuta scomparsa, ve ne potrebbero però essere centinaia che si estinguono senza che la scienza ne sappia nulla. Le stime delle specie attualmente esistenti, fornite dagli zoologi, variano tra loro di circa un fattore dieci. Le valutazioni più attendibili indicano un numero di specie compreso tra 5 e 15 milioni, microrganismi esclusi. Quanto più elevato è il numero delle specie, tanto più numerose saranno in corrispondenza quelle che scompariranno ogni anno per cause naturali e quelle che, al contrario, si origineranno naturalmente.
I tassonomisti hanno dato un nome a circa 1.800.000 specie, ma della maggior parte di esse i biologi non sanno praticamente nulla, soprattutto per quanto riguarda gli insetti, i nematodi e i crostacei, che costituiscono la parte preponderante del regno animale. Infatti la grande maggioranza delle specie multicellulari è costituita dagli insetti, dei quali si ritiene sia stato documentato non più del 10-20%. Circa il 40% delle 400.000 specie conosciute di coleotteri è stato individuato in un'unica località, ma se non si conosce l'areale di distribuzione di una specie non vi è modo di appurarne l'estinzione. Perfino invertebrati senza dubbio estinti non sono mai stati registrati: per es., quando la colomba migratrice si estinse negli anni Venti, a causa della caccia spietata di cui era oggetto, scomparvero insieme a lei due specie di parassiti che però tuttora non figurano negli elenchi dell'IUCN.
La probabilità di osservare un'estinzione è rara quanto quella di assistere a un disastro aereo: è difficile che gli scienziati si trovino nel posto giusto, al momento giusto e che guardino nella giusta direzione. Gli ecologisti ritengono che la grande maggioranza delle estinzioni sia avvenuta nelle regioni tropicali del continente americano. I più vulnerabili sono i pesci d'acqua dolce; infatti oltre un quarto delle loro specie è classificato a rischio. Poiché in Venezuela vivono molti più pesci di acqua dolce che in tutta l'America Settentrionale, si potrebbe pensare che i biologi che svolgono un lavoro di ricerca in quella regione abbiano avuto la possibilità di assistere all'estinzione di numerose specie, ma dopo trent'anni di catalogazione della diversità ittica in questa regione, non è stato documentato un solo caso di estinzione. Del resto, una situazione analoga si verifica anche nel caso di altri gruppi di organismi. Non esistono dunque prove certe dell'estinzione di decine o centinaia di specie ogni anno. È senza dubbio possibile che il programma di calcolo dei dati sia inadeguato, ma resta valida l'ipotesi che il tasso di estinzione corrente sia in realtà meno elevato di quanto temono gli ecologisti.
Siamo alla vigilia di un'estinzione di massa?
La possibilità e la rapidità di una prossima estinzione di massa dipendono dalla velocità con la quale aumenta il tasso di estinzione, di cui numerosi parametri, indipendenti tra loro, segnalano una rapida crescita. La prova più ampiamente accettata riguarda il rapporto tra specie e territorio. Da un confronto tra il numero di specie (S) di uccelli esistenti su isole di diversa grandezza, Wilson ha dedotto più di trent'anni fa che, quando l'estensione (A) di un habitat diminuisce, il numero di specie che vi vivono decresce secondo una legge di potenza, con esponente compreso tra 1/2 e 1/6: in termini matematici, S=cAz, dove c è una costante e il valore di z è in genere di 0,25 (ovvero la radice quarta). Questo significa che la distruzione del 90% di un habitat porterà alla scomparsa della metà delle specie che vi vivono.
Secondo i calcoli di Wilson, all'attuale tasso di distruzione delle foreste tropicali, che è pari a circa l'1% all'anno, un quarto dell'1% delle specie che vivono in questo ambiente si sta estinguendo nel medesimo tempo. Considerando un bacino di circa 10 milioni di specie, dovremmo aspettarci la scomparsa di circa 25.000 specie ogni anno. Lomborg però contesta queste affermazioni sulla base di tre elementi. La prima osservazione è che il rapporto tra specie e territorio è stato ricavato confrontando specie che vivono su isole e quindi non può essere esteso automaticamente a specie viventi in habitat frammentati sulla terraferma. Secondo quanto ha riferito Gretchen Daily, biologa alla Stanford University, nel 2001 oltre la metà delle specie di uccelli originarie della Costa Rica vive in habitat rurali in gran parte soggetti a deforestazione, insieme a percentuali simili di mammiferi e farfalle. Anche se in condizioni difficili, buona parte della fauna selvatica può sopravvivere nei terreni agricoli e nelle macchie boscose residue, ma per quanto tempo ancora possa farlo, nessuno lo sa.
Se confermato, questo fenomeno aiuterebbe a spiegare la seconda critica di Lomborg, relativa al fatto che sia negli Stati Uniti orientali, sia a Porto Rico la distruzione del 98% della foresta primaria non ha causato l'estinzione dei circa tre quarti delle specie di uccelli che vi vivevano. Al contrario, quattro secoli di taglio delle foreste hanno avuto come conseguenza la scomparsa di appena quattro specie di uccelli sulle 200 esistenti negli Stati Uniti e di sette specie sulle 60 originarie di Porto Rico.
Stuart L. Pimm, della Columbia University, ha negato la validità di questa seconda osservazione, affermando che la distruzione dell'habitat agisce in maniera molto selettiva. Solo le specie endemiche, presenti esclusivamente nella zona disboscata, si estinguono, mentre quelle maggiormente diffuse si salvano. Dei 200 tipi di uccelli originari delle foreste degli Stati Uniti orientali, solo 28 non erano presenti anche in altre regioni. Inoltre, la foresta è stata abbattuta in modo molto graduale e gradualmente ha avuto anche la possibilità di ricrescere nelle zone agricole abbandonate dall'uomo. Per questa ragione, anche nel momento di minore estensione, che è stato raggiunto intorno al 1872, l'area boscosa copriva circa la metà dell'estensione della foresta originaria. La teoria specie-territorio prevede che una riduzione di habitat del 50% provochi la scomparsa del 16% delle specie endemiche: in questo caso quattro specie, cioè esattamente quante se ne sono estinte in realtà.
Anche se si ammette la validità dell'equazione specie-territorio, Lomborg ha un terzo argomento da opporre agli ambientalisti più pessimisti. Le statistiche ufficiali indicano che il tasso di deforestazione sta rallentando e che si trova attualmente ben al di sotto dell'1% annuo. Secondo una recente stima della FAO, tra il 1990 e il 2000 la copertura forestale si è ridotta nel mondo a un tasso medio annuo dello 0,2% (11,5 milioni di ettari perduti, a cui vanno sottratti 2,5 milioni di ettari di nuova crescita). Nelle regioni tropicali, tuttavia, dove vive la maggior parte delle specie rare e minacciate, questo tasso è stato dello 0,5%. Gli ecologisti hanno cercato altri mezzi per prevedere quali saranno i tassi di estinzione nel futuro. May e i suoi collaboratori hanno seguito gli spostamenti di diverse specie di vertebrati compresi nella Lista Rossa dell'IUCN per un periodo di quattro anni (due anni nel caso delle piante); successivamente hanno proiettato nei prossimi 7000 anni i dati, peraltro molto limitati, di cui disponevano, giungendo alla conclusione che il tasso di estinzione è destinato a moltiplicarsi da 15 a 55 volte nei prossimi tre secoli. Georgina M. Mace, della Zoological Society di Londra, è giunta a una conclusione molto simile combinando tra loro modelli matematici che permettono di studiare le possibilità di sopravvivenza di alcune specie ben conosciute in funzione del tempo. Nigel E. Stork, un entomologo del Natural History Museum di Londra, ha osservato che nel Regno Unito un volatile ha una probabilità di estinzione 10 volte maggiore di quella di un insetto. Estrapolando questo rapporto al resto del mondo, ha previsto l'estinzione entro il 2300 di un numero di specie di insetti compreso tra 100.000 e 500.000. Lomborg giudica più attendibile quest'ultimo modello, in base al quale conclude che il tasso di estinzione per tutti gli animali rimarrà al di sotto dello 0,208% per decennio e probabilmente sarà inferiore allo 0,7% per i prossimi 50 anni.
Formulare proiezioni di così lunga e vasta portata su una base tanto esigua e incerta di dati richiede una notevole dose di coraggio per qualunque scienziato, soprattutto se si tiene conto del fatto che probabilmente i dati sulle specie a rischio di cui disponiamo sono molto influenzati dai metodi di raccolta, di indagine e di classificazione utilizzati dai tassonomisti. Su circa 5000 sistematici operanti in tutto il mondo, le percentuali di quanti si dedicano allo studio dei vertebrati, delle piante e degli invertebrati sono più o meno le stesse; tuttavia le piante costituiscono non più del 7% del totale delle specie esistenti sul nostro pianeta e i vertebrati meno del 2%.
Esistono comunque buone ragioni per temere che, anche se l'estinzione di massa non è ancora iniziata, il collasso sia in ogni caso imminente. Kevin Higgins, dell'Università dell'Oregon, ha sviluppato recentemente un modello al computer che permette di seguire l'evoluzione di una popolazione di organismi artificiali, simulandone i tassi di mutazione genetica, il comportamento riproduttivo e le interazioni ecologiche. Higgins ha potuto riscontrare che nelle popolazioni di piccole dimensioni le mutazioni tendono a essere così esigue da sfuggire alle maglie della selezione naturale, con una drastica riduzione della capacità di sopravvivenza. Con il restringimento degli habitat e la diminuzione delle popolazioni (al ritmo forse di 16 milioni di individui all'anno, secondo i calcoli di Daily), l'accumulo delle mutazioni genetiche potrebbe rivelarsi una specie di 'bomba a orologeria' genetica, l'equivalente di un'estinzione di massa sotterranea già in atto. Ma dimostrare che questa bomba è già innescata non è facile.
Riuscire a provare se le complesse interdipendenze negli ecosistemi rendano le specie più resistenti o più vulnerabili alla distruzione degli habitat è un problema che divide i biologi. Che cosa accadrà alle piante del genere Ficus, il più diffuso ai tropici, se andrà perduta l'unica varietà di vespe parassite che impollina ogni anno ognuna delle sue 900 specie? O quale sarà la sorte del 79% degli alberi della foresta pluviale di Samoa, se i cacciatori uccideranno tutte le volpi volanti dalle quali dipende la loro sopravvivenza? La preoccupazione con cui molti ambientalisti guardano al futuro riflette in parte la loro convinzione che, una volta distrutta la pietra angolare, tutto l'edificio di un ecosistema sia destinato a crollare.
Non tutti però sono disposti ad accettare questa metafora. Diversi studi recenti hanno mostrato l'esistenza di una certa ridondanza negli ecosistemi. In teoria, l'evoluzione dovrebbe rendere le specie più resilienti (cioè capaci di ritornare al loro stato iniziale dopo essere state sottoposte a un'alterazione), mettendole in condizione di affrontare almeno in parte anche i danni ambientali prodotti dall'attività umana.
Se non lo fa la selezione naturale, potrebbe farlo quella artificiale: questo almeno è quanto sembrano indicare le ricerche di Thomas M. Brooks, della Conservation International, di Andrew Balmford, dell'Università di Sheffield, e di altri studiosi. Brooks e Balmford hanno messo a confronto i tassi di estinzione delle specie vegetali recentemente registrati in quattro regioni molto simili tra loro dal punto di vista ecologico e hanno scoperto che nell'area antropizzata da più lungo tempo e più inquinata, quella del Mediterraneo, si registrava il tasso di estinzione più basso (1,3 specie su mille nell'età moderna). I tassi di estinzione erano più elevati in California (4,0) e in Sudafrica (3,0) e raggiungevano il massimo in Australia occidentale (6,6). La soluzione di questo apparente paradosso deve essere ricercata nel fatto che le specie che non possono coesistere con l'attività umana tendono a scomparire subito dopo la colonizzazione di un territorio, mentre quelle che sopravvivono a questo primo impatto sono meglio 'attrezzate' per respingere gli attacchi a cui le sottoponiamo. Le estinzioni indotte dall'uomo tenderebbero, secondo questa teoria, a diminuire progressivamente con il passare del tempo.
Se questa ipotesi fosse confermata, vi sarebbero diverse implicazioni. Infatti, alcune migliaia di anni fa i nostri antenati potrebbero aver sterminato in Europa, in Asia e nelle altre regioni in cui l'uomo si è insediato da più tempo un numero di specie molto maggiore di quello per cui ci preoccupiamo attualmente. D'altro canto, potremmo avere più tempo del previsto per evitare le possibili catastrofi nelle regioni in cui gli esseri umani fanno già parte dell'ecosistema, e meno tempo per impedirle nelle poche aree rimaste ecologicamente intatte.
La salvaguardia della biodiversità
Mentre imparano ad affrontare la complessa questione della perdita della biodiversità, i biologi sono alla ricerca di nuovi argomenti in grado di convincere i leader politici ed economici, che controllano l'uso del territorio, dell'importanza della sua salvaguardia. L'approccio tradizionale puntava alla protezione dell'habitat di alcuni animali 'simbolo' (come tigri, elefanti e panda), nella speranza che anche le altre specie ne potessero trarre beneficio. Tuttavia questo approccio, sostengono May e altri studiosi, ha il difetto di sacrificare molte specie preziose dal punto di vista scientifico, come, per es., il tuatara, un grande rettile simile a un iguana che vive solo su alcune isolette al largo della Nuova Zelanda. Le uniche due specie di tuatara superstiti formano un gruppo che si è diramato dal tronco principale dell'albero evolutivo dei rettili da talmente tanto tempo da costituire un genere, un ordine e quasi una sottoclasse a sé stanti.
Un secondo gruppo di ecologisti sostiene invece la necessità di passare dalla difesa dei prodotti dell'evoluzione a quella dell'evoluzione stessa. Secondo David Woodruff, dell'Università della California a San Diego, questo approccio permetterebbe di ottenere la soluzione più efficace ed economicamente vantaggiosa del problema generale della conservazione della natura.
Vi sono ancora alcune vaste aree in cui è la selezione naturale a determinare quali specie debbano sopravvivere e quali soccombere. In regioni come la Guiana, in America Meridionale, vive un numero di specie di gran lunga superiore a quello di alcuni dei cosiddetti hot spots (cioè aree della Terra dove la biodiversità è particolarmente elevata), tradizionalmente difesi dalle organizzazioni per la conservazione della natura. Inoltre, mentre questi ultimi sono già stati profondamente trasformati dall'uomo, gli ecosistemi delle regioni vergini sono ancora più o meno intatti.
La salvaguardia di queste aree richiederebbe l'acquisto di porzioni di territorio sufficientemente ampie da permettere lo spostamento latitudinale di interi ecosistemi in risposta ai mutamenti climatici. Sarebbe inoltre necessario proibire qualsiasi sfruttamento del territorio da parte dell'uomo. Benché ambiziosa, questa soluzione potrebbe rivelarsi praticabile: infatti nelle aree naturali non ancora sviluppate i terreni sono relativamente a buon mercato e il numero dei potenziali acquirenti è cresciuto negli ultimi tempi in misura esponenziale. Con l'aiuto di alcuni magnati, per esempio, l'associazione Conservation International è riuscita a battere le industrie del legname, aggiudicandosi l'acquisto di ampie zone di foresta nel Suriname e in Guiana. Con un investimento di 100.000 dollari, questa organizzazione ha ottenuto la conservazione di un'area più vasta del Lussemburgo (2382 km2). Anche Nature Conservancy si è assicurata recentemente il possesso di un appezzamento di foresta pluviale vasto all'incirca come il Libano (10.117 km2), messo in vendita per appena un milione e mezzo di dollari. Il Perù ha infine rilasciato a un gruppo ambientalista la prima 'concessione per la conservazione' - in pratica una licenza rinnovabile per il diritto di non sviluppare il territorio - riguardante 130.000 ettari di foresta. Lo stesso paese si è dichiarato disposto a rilasciare concessioni analoghe su 60 milioni di ettari di foresta demaniale e sono in atto alcuni tentativi di raggiungere accordi di questo genere anche in Guatemala e nel Camerun. Tutto sommato, forse è ancora presto per disperarsi.
repertorio
Il concetto di specie
La biodiversità è la varietà di specie, attuali e potenziali, con la quale si esprime la vita nei diversi ecosistemi. Il concetto di specie ha subito notevoli modifiche con il progredire delle conoscenze. La formulazione più antica, riconducibile al platonismo, è quella di specie tipologica: le specie, distinte e immutabili, erano dotate di caratteristiche, specialmente morfologiche, da rapportare a un modello originario (o archetipo) creato dalla divinità. Secondo questa concezione era possibile definire una specie scegliendo un esemplare tipo come termine di riferimento e confronto con altri eventuali soggetti da identificare. Questo criterio morfologico viene ancora seguito perché, in molti casi, è l'unico applicabile e permette, inoltre, la classificazione delle specie fossili. Tuttavia, numerosi fenomeni biologici, come la variabilità stagionale o legata all'età, il dimorfismo sessuale, il polimorfismo genetico e l'esistenza di specie sorelle, non possono essere interpretati riferendosi al concetto tipologico. L'affermarsi delle concezioni evoluzionistiche ha proposto dunque le specie come entità storiche, suscettibili di continui cambiamenti durante il corso del tempo. Da questi presupposti deriva il concetto di specie biologica che, modificato e riscritto più volte, può essere così enunciato: una specie è una comunità di popolazioni, riproduttivamente isolata dalle altre, che in natura occupa una nicchia specifica. Anche il concetto di specie biologica è stato criticato, sia perché non si può applicare agli organismi che si riproducono solo asessualmente, sia perché non considera le dimensioni spazio-temporali della specie stessa. Intorno agli anni Quaranta, proprio con lo scopo di integrare con la dimensione tempo il concetto di specie biologica, è stato quindi introdotto il concetto di specie evolutiva, con il quale si intende una singola linea evolutiva di popolazioni derivanti da uno stesso antenato, che rimangono distinte da altre simili linee e hanno tendenze evolutive e destino storico propri. Questo concetto può ovviamente essere riferito a organismi che si riproducono sia sessualmente sia asessualmente; una linea evolutiva potrà essere individuata come specie per tutto il tempo in cui permarrà una continuità delle caratteristiche diagnostiche, mentre modifiche rapide di queste caratteristiche segneranno i limiti di specie diverse nel tempo evolutivo. L'affermarsi della sistematica filogenetica o cladistica, infine, ha comportato l'introduzione del concetto filogenetico di specie, secondo il quale la specie è indicata come un insieme indivisibile di organismi, che comprende tutti i discendenti da un fenomeno di cladogenesi (per il quale da una specie o un gruppo più elevato iniziale si originano diverse specie o gruppi che evolvono poi indipendentemente) ed è diagnosticamente distinguibile da altri gruppi. Le specie animali che sono state descritte fino a oggi sono all'incirca 1.800.000, mentre quelle vegetali ammontano a circa 500.000; Linneo nel 18° secolo ne descrisse 4263 animali e circa 10.000 vegetali.
repertorio
La teoria dell'evoluzione
Quasi tutti gli antichi biologi, fino a Linneo, non dubitavano che le specie animali e vegetali fossero state create con atti distinti e si fossero perpetuate nei secoli con le caratteristiche originarie: tale concezione (teoria del fissismo) era soprattutto legata alla dottrina teologica della creazione diretta di tutti gli esseri. La teoria dell'evoluzione afferma invece che sulla Terra sono apparsi dapprima esseri molto semplici, da cui, nel corso dei tempi, sono poi derivati quelli più complessi, i quali sono quindi i naturali discendenti dei primi. Le specie non sarebbero fisse e immutabili ma in via di continua trasformazione (evoluzionismo o trasformismo). Molti sono stati, fin da tempi antichi, i precursori delle teorie evoluzionistiche. Il primo che diede loro un'organica struttura fu Jean-Baptiste Lamarck (1744-1829), ma la sua voce fu sopraffatta da creazionisti e fissisti, e in particolare dalla grande autorità di Georges Cuvier (1769-1832). A Charles Darwin (1809-1882) spetta il merito di avere esposto una teoria coerente e logica. Quando fu introdotta nel mondo scientifico, la sua teoria dell'evoluzione suscitò enorme interesse e conobbe subito un entusiastico successo, pur contrastato da vive opposizioni; inoltre esercitò una profonda influenza, tuttora operante, sullo sviluppo della biologia e di altre scienze. L'ipotesi darwiniana sulla graduale trasformazione delle specie animali e vegetali muove da due osservazioni comuni: nell'ambito di una stessa specie gli organismi mostrano una variabilità biologica, cioè differiscono tra loro per caratteri morfologici, fisiologici, comportamentali ecc.; inoltre in ogni specie il numero degli individui che nascono è molto maggiore del numero che può essere mantenuto in vita dalle possibilità offerte dall'ambiente. Per Darwin ne consegue che gli individui che raggiungono l'età riproduttiva sono quelli meglio adattati alle condizioni ambientali (selezione naturale) e che le variazioni individuali che aumentano le possibilità di sopravvivenza e di riproduzione dei loro portatori sono trasmesse ereditariamente alle generazioni successive: si determinano così una lenta e graduale trasformazione della specie originaria e la formazione di specie nuove che risultano meglio adattate alle diverse condizioni ambientali in cui vivono.
Le prove
Le prove della validità della teoria dell'evoluzione sono tratte da varie discipline. La paleontologia dimostra che in epoche geologiche passate vissero animali e piante diversi dagli attuali. I creazionisti tentarono di spiegare questo fatto incontrovertibile con l'ipotesi dei cataclismi (Cuvier e Alcides d'Orbigny) che avrebbero sconvolto alcune regioni della Terra in diverse epoche geologiche, determinando l'estinzione di molte specie di piante e di animali; tali regioni sarebbero state in seguito nuovamente popolate da specie diverse provenienti da regioni limitrofe. I dati della paleontologia, complessivamente considerati, parlano invece in favore di una graduale evoluzione da forme meno complesse a forme più complesse, anche se non sempre si riesce a rintracciare e a disporre in esatta serie i documenti fossili, a volte frammentari. In molti casi tuttavia si sono ricostruite serie di discendenze che appaiono altamente probabili e significative. La documentazione paleontologica si è arricchita nel tempo di molti importanti reperti. I più antichi finora conosciuti, di organismi a struttura cellulare simili ai cianobatteri, sono databili a circa 3,8 miliardi di anni fa.
L'anatomia comparata dimostra che fra gli organismi oggi viventi esistono più o meno strette affinità di struttura, facilmente interpretabili come indice di più o meno prossima parentela. Soprattutto colpiscono sia l'unità del piano di struttura nell'ambito dei vari tipi animali (per es., vertebrati, molluschi, artropodi), sia il fatto che le stesse parti fondamentali possano, opportunamente modificate, costituire organi diversi quali, per es., la mano dell'uomo, il piede anteriore di un cane, la pinna della balena e l'ala del pipistrello, in cui si ritrovano sempre le stesse ossa. Anche della presenza di organi rudimentali (come, per es., l'occhio della talpa, il polmone sinistro dei serpenti), destituiti della loro funzione, può essere data un'interpretazione in senso evoluzionistico.
L'embriologia conferma, integra ed estende i dati dell'anatomia comparata. Molti organi, che esistono negli embrioni, non si trovano nell'adulto, oppure sono così profondamente trasformati da rendere difficile stabilirne le omologie. Così, per es., nei feti di balena si trovano gli abbozzi dei denti, che scompaiono nell'adulto; nel feto umano si trovano come ossa distinte gli intermascellari, che nell'adulto si fondono con i mascellari; negli embrioni di rettili, uccelli e mammiferi si formano le fessure branchiali, simili a quelle che nei pesci e negli anfibi sono funzionali, mentre nelle classi citate si trasformano, nel corso dello sviluppo, per dare origine a organi diversi. Su fatti di questo genere Ernst Haeckel, verso la fine del 19° secolo, fondò la cosiddetta legge biogenetica fondamentale, la quale afferma che l'ontogenesi (sviluppo embriologico) è la ricapitolazione della filogenesi (storia evolutiva). Sebbene la validità di tale legge appaia oggi più limitata di quanto non si pensasse ai tempi di Haeckel, è certo che alcune affinità e omologie si possono stabilire soltanto in base a ricerche embriologiche. In alcuni casi, specialmente là dove il parassitismo ha profondamente modificato la struttura del corpo, lo studio dello sviluppo e delle forme larvali rivela affinità che nell'adulto sarebbe impossibile scoprire (per es., i crostacei parassiti del gruppo Rizocefali si sono rivelati come crostacei solo il giorno in cui se ne sono conosciute le larve).
La distribuzione geografica degli animali e delle piante presenta singolarità (per es., l'assenza di mammiferi nella Nuova Zelanda, dei placentati nell'Australia, di orsi bianchi nell'Antartide e di pinguini nell'Artide; la presenza dei marsupiali soltanto in Australia e, con pochissime specie, in America) che si possono abbastanza bene interpretare in termini evoluzionistici. Infine la sistematica zoologica e botanica, che si basa sulle affinità di struttura fra gli esseri viventi, offre argomenti di prima importanza a favore della evoluzione. In conclusione, si può affermare che la teoria dell'evoluzione, se pure non dimostrata da prove assolutamente irrefutabili, è suffragata da un gran numero di indizi, alcuni dei quali di grande valore, che, nel loro insieme, costituiscono un'imponente documentazione.
Le teorie
Ammettendo dunque l'evoluzione come evento storico che si sarebbe svolto nei millenni, occorre vedere quali possano essere le cause che l'hanno determinata. Le principali teorie che hanno cercato di rispondere a tale quesito sono, in ordine cronologico: lamarckismo, darwinismo, mutazionismo e ologenesi.
Secondo il lamarckismo, elaborato da Lamarck, la continuità della natura, dimostrata dai resti di organismi fossili appartenuti a epoche remote, è il frutto di un lungo processo storico e le affinità percepibili fra i fossili e le specie viventi testimoniano lontani rapporti di discendenza. I primi microrganismi, formati per generazione spontanea, avrebbero prodotto tutti gli altri viventi, sempre più complessi, trasformandosi successivamente. Questa teoria fa dipendere l'evoluzione dai mutamenti ambientali, secondo precisi nessi di causa-effetto. L'ambiente provoca negli organismi nuovi bisogni che li costringono a contrarre nuove abitudini, le quali, a loro volta, impongono di esercitare determinati organi con una frequenza e un'intensità diverse dalle precedenti; ne deriva quindi la prima legge dell'evoluzionismo lamarckiano (detta dell'uso e disuso), secondo cui l'aumentato e costante uso di un organo lo rafforza, mentre il suo disuso lo indebolisce fino a poterne provocare l'atrofizzazione e quindi la scomparsa. Dalla prima legge deriva la seconda (detta dell'ereditarietà dei caratteri acquisiti), secondo la quale le modificazioni subite per questa via vengono trasmesse ai discendenti: questi col tempo potrebbero allontanarsi dagli antenati fino a costituire una nuova specie distinta.
Il darwinismo è la denominazione delle teorie elaborate da Darwin per interpretare l'evoluzione degli organismi attraverso il meccanismo di selezione naturale. La diffusione di una specie deriva dalla diretta interazione tra gli individui e l'ambiente, la quale determina la riproduzione selettiva dei soli individui idonei. Secondo Darwin, il campo di variabilità in cui agisce il meccanismo di selezione può essere determinato sia da cause interne, di natura genetica, sia da cause esterne, di natura ambientale. Il neodarwinismo, con a capo August Weismann (1834-1914), ha poi negato recisamente che i fattori ambientali possano determinare variazioni ereditarie, non essendo stata dimostrata l'eredità dei caratteri acquisiti per azione dell'ambiente, mentre l'importanza della selezione è stata sperimentalmente riconosciuta e le variazioni ereditarie su cui essa agisce sono state identificate con le mutazioni geniche.
Il mutazionismo è la teoria dell'evoluzione formulata da Hugo de Vries (1848-1935), secondo la quale ogni specie animale o vegetale, in un determinato periodo della sua esistenza, presenta la formazione di numerose razze nuove in seguito alla comparsa di un gran numero di mutazioni. La selezione naturale decide poi quale di queste razze può rimanere in vita, nei diversi ambienti. L'evoluzione avverrebbe quindi per salti bruschi, anziché in modo continuo, come pensava Darwin. In seguito, fu dimostrato che molte delle variazioni studiate da de Vries nella pianta Oenothera lamarckiana erano da interpretarsi come combinazioni di geni preesistenti, che le mutazioni esistono, ma spesso sono rappresentate da variazioni di minima entità, che il processo di formazione di mutazioni è costante e continuo in ogni specie. L'analisi delle mutazioni, quale è stata eseguita dalla genetica moderna, confermando l'importanza delle mutazioni per l'evoluzione e negando la limitazione del periodo mutativo e il salto di notevole ampiezza dal punto di vista fenotipico, ha ricondotto il mutazionismo nell'ambito del darwinismo e ha riconosciuto nelle mutazioni la sorgente della variabilità ereditaria, su cui la selezione naturale agisce in modo continuo e graduale. L'ologenesi, infine, è la teoria evoluzionistica elaborata da Daniele Rosa (1857-1944), secondo cui l'evoluzione degli organismi sarebbe avvenuta per processi interni e preordinati, a partire dagli organismi più semplici; il processo di formazione di nuove specie sarebbe inoltre riconducibile a una ramificazione dicotomica di una specie preesistente.
I meccanismi dell'evoluzione
La scoperta dei meccanismi che sono alla base dell'evoluzione è dovuta per la massima parte ai grandi progressi compiuti dalla genetica nel 20° secolo; questa scienza ha il merito di aver dimostrato inconfutabilmente l'erroneità della teoria lamarckiana dell'ereditarietà dei caratteri acquisiti. I processi evolutivi finora ben dimostrati sono: la mutazione, la selezione, la deriva genetica e l'isolamento geografico. Essi determinano sostanzialmente una variazione delle frequenze alleliche nelle popolazioni. Lo studio della variabilità ereditaria e della sua modulazione nel tempo e nello spazio è stato effettuato da G. Hardy e W. Weinberg (1908) i quali, indipendentemente, arrivarono a scoprire quella che oggi è nota come legge dell'equilibrio di Hardy-Weinberg. L'importanza di questa legge fu però riconosciuta solo dopo il 1920 e d'allora in poi si sviluppò la trattazione matematica del problema dell'evoluzione, soprattutto per opera di R.A. Fisher, J.B.S. Haldane e S. Wright. Le mutazioni. Le mutazioni operano a tre livelli: genico, producendo nuovi alleli; cromosomico, producendo nuove strutture cromosomiche; genomico, dando luogo a cambiamenti del numero dei cromosomi. Le mutazioni rappresentano la base dell'evoluzione: le mutazioni geniche possono produrre effetti sull'insieme dei caratteri visibili di un organismo (fenotipo); avvengono spontaneamente con frequenza molto bassa (circa 1/100.000 o 1/1.000.000 per generazione); la loro frequenza può essere aumentata da agenti mutageni, come agenti ionizzanti o alcune sostanze chimiche. Comunque le mutazioni che si verificano in un determinato ambiente sono casuali, cioè non si verificano con maggiore frequenza le mutazioni che consentono un migliore adattamento degli individui all'ambiente stesso. Le mutazioni cromosomiche non producono modificazioni delle caratteristiche morfologiche ma controllano caratteristiche funzionali; inoltre rivestono un ruolo importante nel determinare la sterilità degli ibridi negli incroci interspecifici. Le mutazioni genomiche, molto diffuse nelle piante e rare negli animali, causano variazioni dell'assetto cromosomico che, invece di essere diploide (2n, con una doppia serie di cromosomi), come di norma, può essere triploide (3n), tetraploide (4n) ecc.; possono produrre anche variazioni che interessano una sola o poche coppie di cromosomi (aneuploidia).
La selezione naturale. - L'orientamento dell'evoluzione è il risultato dei fenomeni di selezione naturale, i quali agiscono sulla variabilità genetica, fornita dalle mutazioni casuali, attraverso la riproduzione differenziale; infatti i portatori di combinazioni geniche favorite dalla selezione contribuiscono con un maggior numero di discendenti alla generazione successiva. L'esistenza e l'azione della selezione naturale è suffragata da numerose prove sperimentali, date da esperimenti di laboratorio e da osservazioni in natura. Per es., ceppi di batteri incapaci di sopravvivere in presenza di un certo antibiotico divengono improvvisamente resistenti a esso e si moltiplicano, nonostante la presenza dell'antibiotico, grazie alla comparsa per mutazione di individui resistenti. Questa mutazione, preesistente nel genotipo dei batteri alla comparsa dell'antibiotico, risulta neutra o addirittura sfavorevole in ambienti privi di antibiotici, ma è invece favorita dalla selezione in presenza di antibiotici; infatti solo gli individui portatori di tale mutazione sopravvivono e si riproducono, mentre tutti gli altri muoiono: ciò spiega perché alcuni antibiotici rivelatisi molto efficaci al momento della loro comparsa perdono in breve tempo la capacità di agire su molti germi patogeni. Analoghi ai fenomeni di resistenza dei batteri agli antibiotici sono quelli di resistenza agli insetticidi da parte di molti artropodi; anche gli insetticidi hanno quindi agito come forze selettive modificando in breve tempo il patrimonio ereditario di numerose popolazioni.
Deriva genetica. - Si hanno fenomeni di deriva quando la frequenza di uno o più alleli (o addirittura la presenza o l'assenza di essi) in una popolazione è determinata unicamente dal caso, e ciò avviene quando più alleli hanno valore selettivo simile e quando una popolazione viene drasticamente ridotta di numero (per es., in una popolazione di zanzare che abita una zona trattata con insetticidi, la popolazione che si formerà dopo il trattamento porterà solo gli assetti cromosomici delle zanzare sopravvissute); gli effetti della deriva sono tanto più accentuati quanto minore è il numero degli individui della popolazione. Il ruolo della deriva genetica nell'evoluzione biologica è un argomento ancora oggi controverso.
Isolamento geografico. - L'isolamento geografico riveste un ruolo fondamentale nell'evoluzione e in particolare nel processo di formazione delle specie, o speciazione, che ne è il punto cruciale. Esso, impedendo il flusso genico e quindi il rimescolamento genetico tra popolazioni adiacenti, permette alle popolazioni isolate di adattarsi indipendentemente ai diversi ambienti, ed è quindi la causa primaria del loro differenziamento in razze, semispecie e specie distinte. Così, se una singola specie è distribuita uniformemente su un certo territorio e in seguito, sia per effetto delle diverse pressioni selettive che si realizzano nelle varie parti dell'areale, sia per la presenza di una barriera geografica (catena montuosa, tratto di mare o più semplicemente un territorio inabitabile per la specie), si stabiliscono differenze genetiche tra gruppi di popolazioni a mano a mano isolati, questi si evolveranno indipendentemente gli uni dagli altri dando origine a razze geografiche o a specie distinte. Se la barriera viene a cadere e i gruppi di popolazioni ritornano in contatto, possono o ibridarsi o risultare riproduttivamente isolati, e in tal caso il processo di speciazione è compiuto. È da notare che l'isolamento riproduttivo non viene necessariamente raggiunto nel corso dell'isolamento geografico, perché, finché le popolazioni rimangono isolate, nessuna forza selettiva agisce in favore dell'insorgenza di barriere riproduttive; è quando le popolazioni tornano in contatto che l'isolamento riproduttivo può essere favorito dalla selezione naturale, instaurarsi e rafforzarsi con meccanismi di varia natura (genetica, ecologica, etologica ecc.). Se infatti la prole derivata da due popolazioni precedentemente isolate ha un grado di adattamento (fitness) inferiore a quello della prole derivata da incroci intrapopolazionali, la selezione naturale favorirà le mutazioni e le combinazioni geniche che rendono più probabile l'accoppiamento tra individui della stessa popolazione.
Formazione delle specie per poliploidia. - Nelle piante, ove è largamente diffusa la riproduzione vegetativa, la formazione delle specie attraverso l'ibridazione di due entità diverse e successiva poliploidizzazione assume un ruolo importante. Infatti, anche se gli ibridi tra due specie sono sterili, a causa di disturbi nella formazione dei gameti, essi possono ugualmente propagarsi per riproduzione vegetativa. Uno dei più noti e importanti organismi poliploidi è il frumento da pane (Triticum aestivum) con 42 cromosomi, che si è originato in seguito alla produzione di un ibrido tra un frumento coltivato, con 28 cromosomi, e una graminacea selvatica dello stesso genere, con 14 cromosomi. Fenomeni simili non sono conosciuti negli animali e ciò è probabilmente in rapporto alla quasi generale assenza in essi della riproduzione vegetativa; sono invece frequenti casi di animali in cui l'ibridazione e la poliploidia hanno sostenuto un ruolo importante nell'evoluzione (platelminti, anellidi, crostacei, insetti, alcuni vertebrati); in questi casi però vi è presenza anche di ermafroditismo o di partenogenesi.
Teoria sintetica dell'evoluzione
Le conoscenze dei meccanismi operanti nell'evoluzione biologica sono riassunte nella cosiddetta teoria sintetica (o sintesi moderna): in tutti gli organismi esiste una certa variabilità, alimentata da una parte dal costante insorgere di mutazioni e dall'altra dalla ricombinazione genica che avviene a ogni generazione in seguito alla riproduzione sessuale. Su questa variabilità genetica, che è puramente casuale e non orientata in alcuna direzione, agisce come fattore direttivo principale la selezione naturale; quando il numero degli individui della popolazione è molto basso, entra in gioco la deriva genetica. Le popolazioni di una specie divengono così geneticamente diverse nelle varie parti dell'areale; il processo di adattamento di queste popolazioni ai rispettivi ambienti è favorito dall'isolamento geografico che impedisce il flusso genico. Si formano così le razze geografiche, alcune delle quali possono divenire specie distinte in seguito all'insorgere di barriere di isolamento riproduttivo, che è il punto focale della speciazione. Le varie specie costituiscono ciascuna un complesso isolato di geni e si evolvono indipendentemente l'una dall'altra, dando origine, nel corso delle ere geologiche, alle categorie sistematiche superiori. La teoria sintetica fornisce così un'interpretazione, verificabile anche sperimentalmente, della cosiddetta microevoluzione e cioè del differenziamento di razze, semispecie e specie da progenitori comuni. Molti biologi ritengono che i meccanismi evolutivi proposti dalla teoria sintetica possano spiegare anche, se proiettati alla scala dei tempi geologici, l'origine delle categorie sistematiche superiori alla specie, come generi, famiglie, ordini, classi e phyla (la cosiddetta macroevoluzione). Altri biologi ritengono invece necessario, per spiegare la macroevoluzione, ricorrere a diversi meccanismi ipotetici come, per es., particolari mutazioni, dette mutazioni sistemiche, che avrebbero effetti molto più cospicui di quelle la cui esistenza è stata dimostrata dalla genetica. Non è stata peraltro fornita alcuna prova convincente dell'esistenza delle mutazioni sistemiche. Le recenti tecniche di biologia molecolare (ibridazione DNA-DNA e DNA-RNA, polimorfismi enzimatici) hanno apportato un notevole contributo agli studi microevolutivi, permettendo di stabilire a livello genetico, nell'ambito di vari gruppi di organismi, le relazioni filogenetiche che intercorrono tra le varie entità: popolazioni, razze, sottospecie, specie gemelle ecc. Negli anni Settanta e Ottanta, nuove acquisizioni hanno messo in crisi la teoria sintetica classica, tanto da far invocare la necessità di una nuova sintesi. In particolare, gli studi di biologia molecolare hanno lanciato due sfide dirette alla teoria sintetica. Una di queste è l'ipotesi che lo sviluppo di variazioni nel DNA sia stimolato da un tipo di determinismo molecolare, e non dal puro caso. L'altra è la teoria della neutralità, contrapposta all'ipotesi precedente e in base alla quale il caso controlla non solo la comparsa iniziale delle variazioni genetiche, ma anche la loro successiva affermazione in una popolazione. Un differente tipo di sfida è venuto invece dalla paleontologia ed è noto come teoria degli equilibri intermittenti (o punteggiati). Basata su nuove interpretazioni dei resti fossili, questa sostiene che l'evoluzione procede non a passo continuo, ma irregolarmente, a balzi.
repertorio
Le estinzioni nella storia della vita sulla Terra
La scomparsa di specie animali o vegetali è un fenomeno che si è prodotto molte volte nel corso della storia della Terra, come ci attestano i resti fossili di numerose specie (o di più vasti gruppi sistematici) oggi non più esistenti. L'estinzione può essere dovuta a varie cause, anche concomitanti: eventi geologici, cambiamenti climatici, variazioni del chimismo delle acque, epidemie, competizione con altre specie ecc. Tra i numerosi gruppi animali scomparsi in ere geologiche passate si ricordano i Tetracoralli, i Tabulati (celenterati) e le Trilobiti, estinti alla fine del Paleozoico; le Ammoniti, le Belemniti e le Rudiste, scomparse, insieme a molti gruppi di rettili (Saurischi, Ornitischi, Pterosauri, Ittiosauri, Sauropterigi), alla fine del Mesozoico. In geologia stratigrafica, l'estinzione delle specie, e quindi il ritrovamento dei loro resti fossili, è un criterio di fondamentale importanza ai fini della determinazione dell'età di un corpo sedimentario. Un'estinzione di massa è una crisi biologica che continua per un intervallo di tempo definito (1-1,5 milioni di anni) su scala mondiale. Recenti studi hanno messo in evidenza che si sono verificate almeno cinque estinzioni di massa nel passato geologico della Terra. La prima è avvenuta alla fine dell'Ordoviciano (438 milioni di anni fa) e ha comportato la scomparsa di circa l'85% delle specie marine esistenti; la successiva si è verificata nel tardo Devoniano (circa 370 milioni di anni fa) e si calcola che si siano estinte l'83% delle specie marine; la terza, quella principale, è avvenuta alla fine del Permiano (250 milioni di anni fa), con la scomparsa del 96% delle specie di invertebrati marini allora viventi; la quarta è avvenuta alla fine del Triassico (210 milioni di anni fa), con l'80% di specie marine estinte, e la quinta alla fine del Cretaceo (67 milioni di anni fa), con l'estinzione del 76% di specie marine. Le cause presunte di queste estinzioni sono tuttora oggetto di indagine da parte degli scienziati. Numerose ipotesi sono state avanzate: dall'impatto sulla Terra di meteoriti e asteroidi all'intensa attività vulcanica, fenomeni che avrebbero potuto alterare la composizione dell'atmosfera, causando surriscaldamento dell'aria e forti fluttuazioni del livello dei mari. L'ipotesi di una periodicità delle crisi biologiche (ogni 28-30 milioni di anni circa), calcolata su basi statistiche e forse legata a movimenti orbitari periodici della Terra in seno al sistema solare o alla sua galassia, è anch'essa assai dibattuta. Anomalie riscontrate su scala mondiale nella concentrazione di platino e iridio nei sedimenti in concomitanza delle estinzioni di massa sembrano avvalorare un'origine extraterrestre (meteoriti, comete), senza tuttavia far escludere cause endogene, come per es. il vulcanismo.
Dell'estinzione, avvenuta in epoca storica, di numerose specie è in gran parte responsabile l'uomo, sia attraverso lo sterminio diretto (caccia, distruzione delle uova, ricerca di pellicce, penne e piume ecc.), sia indirettamente attraverso le modificazioni che apporta all'ambiente (costruzione di aree industriali o di abitazione, introduzione di nuove specie in comunità già in equilibrio dal punto di vista ecologico, inquinamento ecc.). Negli ultimi 500 anni l'uomo ha portato all'estinzione più di 1000 specie; tra quelle più note si possono ricordare: l'alca (Pinguinus impennis), uccello inetto al volo che abitava le isole dell'Atlantico settentrionale e che si estinse nel 1844 a causa di una caccia sfrenata per il suo pregiato piumaggio; la colomba migratrice (Ectopistes migratorius), abbondantissima fino a due secoli fa, il cui ultimo esemplare morì negli anni Venti; il moa (Dinornis maximus), grosso uccello corridore della Nuova Zelanda scomparso intorno al 1770; il dodo (Raphus cucullatus), columbiforme della grossezza di un tacchino, inetto al volo, che si estinse nell'isola Maurizio nel 1693; la ritina (Hydrodamalis stelleri), grosso sirenide marino, scomparso verso la metà del 19° secolo; il quagga (Equus quagga), il cui ultimo esemplare morì nel giardino zoologico di Amsterdam nel 1883; l'uro (Bos primigenius), estintosi nel 17° secolo nel territorio della Vistola in Masovia ecc. Quasi estinto è il bisonte europeo (Bison bonasus): se ne allevano alcuni capi in taluni parchi. Anche il bisonte americano (Bison bison) è stato molto prossimo all'estinzione e viene accuratamente protetto. In Italia, in particolare, si sono estinti agli inizi del 20° secolo l'avvoltoio degli agnelli, l'aquila di mare, il falco pescatore e sono in via di estinzione numerosissimi uccelli, tra cui alcune specie endemiche italiane. La stessa situazione, in proporzioni maggiori, si verifica per gli invertebrati, per i quali anche forme di collezionismo indiscriminato, non estranee a notevoli interessi economici, hanno contribuito e contribuiscono alla scomparsa di specie (farfalle, coleotteri, carabidi) particolarmente ricercate dai collezionisti. Per ovviare al pericolo di lasciare scomparire piante o animali, sono stati istituiti parchi nazionali e riserve e sono state fondate società per la protezione della natura. Contemporaneamente, si è cercato da tempo di stabilire dei criteri che permettano di definire in modo migliore lo status dei taxa in pericolo. L'IUCN (International union for conservation of nature and natural resources, oggi denominato World conservation union) ha così stabilito quattro categorie: specie minacciate, vulnerabili, rare, indeterminate. Poiché ci si è resi conto che l'attribuzione di una specie, animale o vegetale, a uno dei gruppi summenzionati era troppo legata a giudizi, a volte soggettivi, degli specialisti, è iniziato un progetto internazionale, sempre a cura dell'IUCN, che ha lo scopo di individuare nuove categorie, utilizzando metodi di calcolo probabilistico che dovrebbero permettere di stabilire le possibilità di sopravvivenza della specie nel tempo.
repertorio
La lista Rossa
La Lista Rossa delle specie a rischio di estinzione, a cura dell'IUCN, è la più prestigiosa stima sullo stato della biodiversità nel mondo. La lista, che viene aggiornata anno per anno ma è ripubblicata ogni quattro anni, non solo classifica gli animali in base al rischio di estinzione, ma fornisce anche indicazioni sul trend delle popolazioni, le minacce principali, gli habitat ecc. Il grado di rischio che ogni specie corre è classificato in sette categorie (v. tabb. 1 e 2): 1) estinto (ES); 2) estinto in natura (EN); 3) criticamente minacciato (CR); 4) minacciato (MI); 5) vulnerabile (VU); 6) a rischio minore (RM); 7) dati insufficienti (DI). Se una specie rientra nella categoria 3, 4 o 5, viene definita minacciata.
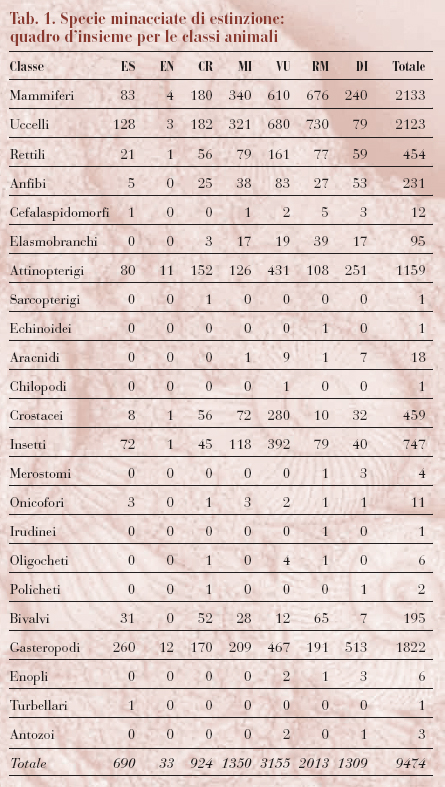
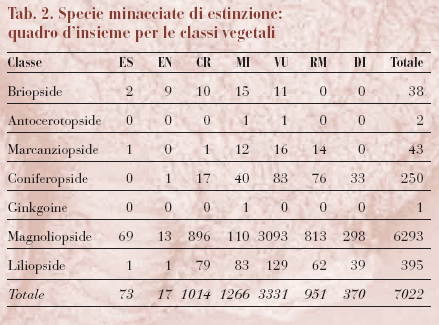
Nella lista pubblicata nel 2000 sono incluse 18.276 tra specie e sottospecie; in totale, 11.046 specie, tra animali e vegetali, sono minacciate, affrontando un alto rischio di estinzione nel breve termine. In particolare, risultano a rischio il 24% delle specie di mammiferi, il 12% di uccelli, il 25% di rettili, il 20% di anfibi e il 30% di pesci, soprattutto d'acqua dolce. Il numero totale di specie animali minacciate è passato da 5205 a 5435 negli ultimi quattro anni e i paesi con il maggior numero di specie a rischio sono Indonesia, India, Brasile e Cina, mentre le specie vegetali diminuiscono rapidamente in America Latina, in Africa orientale e occidentale e nel Sud-Est asiatico. In generale, la percentuale totale di mammiferi e uccelli a rischio non è cambiata molto dalla compilazione della precedente Lista Rossa, mentre è aumentato il numero delle specie che dalla categoria a rischio minore sono passate a quelle a più alto rischio.