
Autoimmunità
Autoimmunità
La funzione del sistema immunitario è quella di individuare le macromolecole estranee all'organismo e di attivare le difese idonee a eliminare sia queste, sia gli 'invasori' che le producono (virus, batteri, metazoi), come risulta chiaramente dall'osservazione che gli individui nei quali il sistema immunitario non è sufficientemente sviluppato, oppure è stato inattivato da farmaci, radiazioni o virus (per es., HIV), soccombono a molteplici infezioni dovute ad agenti diversi. Questa funzione di difesa richiede che il sistema immunitario sia in grado di distinguere le macromolecole proprie da quelle estranee. È questo un problema di non facile soluzione, specialmente nel caso delle proteine, perché non esistono proprietà chimiche o di carattere generale in base alle quali il sistema immunitario possa operare questa distinzione. Pertanto, è necessario che il sistema immunitario, durante lo sviluppo e la maturazione delle sue cellule (i linfociti), 'impari' a riconoscere e a convivere con le macromolecole (gli antigeni) proprie, conservando e ampliando nella sua memoria la capacità di reagire contro gli antigeni estranei. La base di questo processo di apprendimento, che dura per tutta la vita, risiede nella selezione clonale dei linfociti.
I recettori per gli antigeni, sia nei linfociti T sia nei linfociti B, sono controllati da geni che durante la maturazione di queste cellule subiscono processi di riarrangiamento nei quali il caso svolge un ruolo fondamentale. Vengono così prodotti cloni diversi di linfociti, ciascuno con un differente recettore. Il ruolo del caso nella generazione della diversità clonale dei linfociti è essenziale per la produzione di una popolazione di cellule capaci di reagire, in linea di principio, contro antigeni diversi e non prevedibili a priori, quali sono quelli dei potenziali agenti infettanti presenti all'esterno dell'organismo. Tuttavia, proprio l'elemento casuale prospetta due grandi problemi: il primo consiste nella generazione di cellule con recettori completamente inutili; il secondo invece nella produzione di cellule con recettori dannosi, in quanto capaci di reagire efficacemente contro gli antigeni propri dell'organismo. Ovviamente, il sistema immunitario delle cellule mature deve comprendere il minimo numero possibile di cloni inutili e deve eliminare o poter controllare i cloni potenzialmente dannosi. Il rispetto di questa regola fondamentale si ottiene attraverso processi di selezione positiva e di selezione negativa, che operano durante la maturazione dei linfociti negli organi linfoidi centrali e che, mediante l'eliminazione della grande maggioranza delle cellule prodotte a quel livello, provvedono a disseminare negli organi linfatici periferici, là dove si svolgerà la funzione protettiva del sistema immunitario, i cloni di cellule potenzialmente utili e non potenzialmente dannose.
La selezione clonale dei linfociti
I linfociti T: selezione positiva e selezione negativa; la soppressione
Consideriamo i processi di selezione positiva e negativa dei linfociti T e B, poiché la loro comprensione è necessaria per chiarire sia la funzione normale del sistema immunitario, sia le sue deviazioni patologiche che conducono alle malattie autoimmuni. I linfociti T sono forniti di recettori clonali che riconoscono frammenti di antigeni proteici inseriti in una 'tasca' presente nella struttura delle molecole di istocompatibilità. Le strutture molecolari con le quali i recettori dei linfociti T reagiscono sono quindi costituite da un complesso, formato da un peptide e dalla molecola di istocompatibilità (MHC, Major histocompatibility complex) che lo presenta. Nel timo, dove avviene la maturazione e la selezione dei linfociti T, normalmente non vi sono antigeni proteici estranei all'organismo; pertanto la selezione dei cloni T, sia quella positiva sia quella negativa, è operata dal contatto dei recettori esclusivamente con complessi di peptidi derivati da proteine 'proprie' e da molecole MHC proprie.
Siamo dunque di fronte al problema del come e del perché le stesse strutture molecolari possano determinare sia la selezione positiva sia la selezione negativa dei linfociti T, e di fronte all'apparente paradosso di due risultati opposti conseguenti al contatto dei recettori clonali con gli stessi complessi di peptidi di derivazione endogena, presentati dalle proprie molecole MHC. Su questo argomento, di importanza centrale per tutta l'immunologia e in particolare per il capitolo dell'autoimmunità, si è detto e scritto moltissimo, ma a tutt'oggi non possiamo asserire di essere giunti a un completo chiarimento del problema; tuttavia, particolarmente in seguito a recenti esperimenti condotti in vitro su colture di frammenti di timo, cominciamo a comprenderne gli aspetti fondamentali. Anzitutto sono stati accertati due fatti: (a) i linfociti, che durante il processo di maturazione nel timo esprimono sulla loro membrana recettori che non reagiscono contro i complessi MHC-peptidi presenti sulle cellule della matrice linfoepiteliale timica, non evolvono ulteriormente, ma sono eliminati mediante l'attivazione di un programma di autodistruzione; (b) al contrario, i linfociti il cui recettore reagisce con il complesso MHC-peptidi presentato loro dalle cellule della matrice timica evolvono fino alla maturazione e continuano a proliferare.
Questa è la base della 'selezione positiva', mediante la quale sopravvivono solo linfociti con recettori capaci di riconoscere peptidi presentati dalle molecole MHC proprie dell'organismo stesso. Il significato funzionale di questo aspetto della selezione positiva è facilmente intuibile, in quanto la funzione ultima dei linfociti T è quella di riconoscere antigeni (peptidi) presentati dalle molecole MHC proprie dell'organismo e sarebbero inutili quei cloni con recettori non idonei a reagire contro un complesso molecolare del quale le molecole MHC proprie sono parte essenziale. Meno evidente è la funzione, nella selezione positiva, della reattività dei recettori nei confronti di peptidi derivati da proteine endogene, presenti e presentati nel timo. Questo aspetto della selezione positiva sembra, a prima vista, destinato a produrre una popolazione di linfociti T autoreattivi, potenzialmente capaci di generare ogni sorta di fenomeni autoimmunitari. In realtà, la propensione autoreattiva della selezione positiva è limitata dalla selezione negativa. Possiamo tuttavia domandarci, in linea teorica, se non vi sia un profondo motivo biologico per la selezione positiva di cloni linfocitari con recettori reattivi contro molecole MHC proprie che presentano peptidi endogeni. Questo motivo può essere individuato nella somiglianza generale di peptidi potenzialmente derivati dalla frammentazione di antigeni batterici e, soprattutto, virali con i peptidi prodotti dalla frammentazione di proteine endogene.
Perciò la selezione positiva nel timo evita la sopravvivenza di tutta una serie di cloni il cui recettore, originato da eventi in parte casuali a livello del DNA cellulare, ha una struttura non idonea a reagire contro i complessi MHC-peptidi che potrebbero essere prodotti nel mondo esterno da potenziali agenti infettanti. In altri termini, la selezione positiva nel suo complesso rinuncia a generare una popolazione di linfociti onnipotente, ma largamente sovradimensionata rispetto allo spazio reale del sistema immunitario, in favore di una popolazione, pur sempre molto ampia, già orientata verso una possibile utilizzazione pratica della propria specificità. A questo punto, è necessario un processo capace di eliminare, già a livello del timo, almeno quell'aliquota di cloni che hanno superato il vaglio della selezione positiva, ma che, essendo provvisti di recettori dotati di un'alta affinità per i complessi MHC-peptidi presenti nel timo stesso, sarebbero sicuramente capaci, una volta maturi e trasferiti nel sistema linfatico periferico, di generare reazioni autoimmunitarie. Questo processo è la 'selezione negativa'.

Nel loro complesso, dunque, le azioni selettive operanti nel timo eliminano sia i cloni che non reagiscono contro i complessi MHC-peptidi presentati dalle cellule stromali, sia i cloni che reagiscono contro gli stessi complessi con alta affinità (fig. 2). La popolazione di cellule che matura e lascia il timo per colonizzare gli organi linfatici periferici è quindi quella che reagisce con bassa affinità contro i complessi MHC-peptidi propri ed è incapace di produrre spontaneamente reazioni immunitarie nelle varie sedi dell'organismo. Tuttavia, questa popolazione include cloni in grado di reagire con alta affinità contro complessi formati da molecole MHC proprie e peptidi derivati da antigeni estranei, e quindi capaci di una efficace reazione immunitaria di difesa contro agenti infettanti. Questa capacità è legata alla potenziale omologia, più o meno stretta, fra i peptidi derivati dall'elaborazione (processing) delle proteine proprie e quelli derivati dalle proteine batteriche o virali: in altri termini, un clone selezionato sulla base di recettori che reagiscono in modo approssimativo contro i complessi MHC-peptidi endogeni ha una certa probabilità di possedere un recettore che reagisce bene quando è confrontato con una molecola MHC propria, la cui tasca sia occupata da un peptide derivato, per esempio, dall'elaborazione di una proteina virale.
Un'altra proprietà del sistema, ancorché probabilmente non legata direttamente a motivi funzionali, è la alloreattività, cioè la capacità di un'aliquota relativamente alta di linfociti T (attorno al 3%) di reagire energicamente nei confronti di cellule che esprimono molecole MHC allogeniche, cioè codificate da geni alleli presenti nella stessa specie. Questa proprietà ‒ che è stata alla base della scoperta e della definizione degli antigeni di istocompatibilità ‒ si spiega tenendo conto della possibilità che cloni relativamente numerosi di linfociti T, selezionati nel timo sulla base dell'espressione di recettori di bassa affinità per complessi MHC-peptidi propri, siano anche cross-reattivi, con affinità relativamente alta, contro complessi formati da molecole MHC simili (ma non identiche) alle proprie e peptidi uguali a quelli elaborati nel timo dell'individuo nel quale i linfociti sono maturati.
Così congegnata, la selezione negativa nel timo può agevolmente controllare la produzione di cloni di linfociti autoreattivi verso le proteine solubili prodotte dall'organismo stesso ‒ sempre che queste possano raggiungere il timo in concentrazione sufficiente per essere internalizzate, frammentate e presentate sulle molecole MHC di classe II delle idonee cellule ivi presenti ‒ e verso tutte le proteine non solubili sintetizzate nel citoplasma delle cellule stromali del timo, i cui peptidi sono presentati in quella sede dalle molecole MHC di classe I. In questo modo, tutte le proteine presenti nell'insieme delle cellule dell'organismo ‒ gli enzimi che partecipano alle funzioni metaboliche generali e le proteine strutturali comuni a tutte le cellule, cioè i prodotti dei cosiddetti 'geni di mantenimento' (household) ‒ sono protette dalla selezione negativa nel timo (la cd. 'tolleranza immunitaria centrale'). Di conseguenza, queste proteine raramente divengono autoantigeni, salvo in quelle malattie nelle quali appunto la tolleranza centrale è deficitaria per varie ragioni, come nel lupus eritematoso sistemico e nei suoi modelli sperimentali. Un interessante esempio di tolleranza centrale è dato da un esperimento nel corso del quale in un embrione di pollo è stato trapiantato un timo di quaglia; successivamente, nell'embrione è stato innestato l'abbozzo di un'ala di quaglia, che si è normalmente sviluppata.
Tuttavia, si pone il problema di come la tolleranza centrale possa determinare l'eliminazione di cloni T potenzialmente autoreattivi nei confronti di antigeni di differenziazione espressi da cellule di organi periferici. Questi antigeni, specialmente se non solubili ed espressi nelle cellule periferiche in bassa quantità, hanno scarse possibilità di raggiungere il timo e determinare in quella sede la selezione negativa dei cloni T necessaria per stabilire la tolleranza centrale. L'ipotesi che nel timo, in cellule speciali, possa aver luogo la sintesi ectopica di molte delle proteine proprie degli organi periferici e che questo possa dar luogo alla tolleranza centrale nei confronti degli antigeni di differenziazione, fu avanzata nel 1989 da R. Linsk, Max Gottesman e Benvenuto Pernis. Più recentemente è stato dimostrato che, nell'uomo, una sindrome autoimmunitaria multipla, caratterizzata da una poliendocrinopatia, candidiasi e distrofia ectodermica (APECED), è legata a mutazioni nel gene di un controllore di trascrizione detto AIRE (Autoimmuneregulator), la cui funzione è precisamente quella di indurre nelle cellule epiteliali del midollo timico la sintesi di alcune centinaia di proteine di differenziazione degli organi periferici.
Uno studio completo della situazione immunologica in topi transgenici, nei quali il gene omologo (aire) era stato modificato in modo da produrre una proteina inattiva, ha portato a risultati estremamente interessanti. Nel timo dei topi aire-deficienti mancavano i messaggeri corrispondenti ad alcune centinaia di geni che erano invece attivi nel timo dei topi normali. Questi messaggeri corrispondevano in gran parte a proteine di differenziazione dei tessuti periferici. Si confermava così che, sotto l'influenza del fattore aire, il timo normale sintetizza una serie di proteine corrispondenti a una 'ombra' della periferia. Questa ombra è deficitaria se il fattore aire non funziona e, osservazione fondamentale, in questi topi compaiono multiple reazioni autoimmunitarie evidenziate da infiltrazioni linfocitarie in diversi organi e molteplici autoanticorpi. Queste manifestazioni autoimmuni sono evidentemente la conseguenza di una deficienza della tolleranza centrale conseguente alla mancata espressione nel timo di geni di differenziazione degli organi periferici. L'autoimmunità che ne consegue non è però immediata ma è progressiva nei primi mesi di vita del topo, e inoltre varia da individuo a individuo con interessamento di organi diversi e di differenti strutture nello stesso organo.
Questi ultimi fatti, a mio avviso, sono interpretabili facilmente sulla base dell'esistenza di un altro livello di tolleranza oltre la tolleranza centrale, e cioè la tolleranza periferica. L'esistenza di meccanismi di tolleranza periferica è stata dimostrata nel modo migliore con esperimenti condotti in topi transgenici, nei quali è stato introdotto un gene esterno unito a un segmento regolatore capace di determinarne l'espressione in un dato organo: sono stati prodotti, per esempio, topi che esprimevano una molecola MHC di classe II o di classe I allogenica, propria cioè di un altro ceppo di topi, esclusivamente nelle cellule insulari o in quelle acinari del pancreas, a seconda che il gene per la molecola MHC estranea fosse collegato con il segmento regolatore per l'insulina o per l'elastasi. Con lo stesso meccanismo di introduzione di geni estranei nella linea germinale, sono state prodotte linee di topi che esprimevano a livello delle cellule insulari del pancreas una classe di molecole MHC, le molecole I-E, inesistenti nell'organismo di quel particolare ceppo di topi, oppure addirittura una proteina virale (l'antigene T del virus SV40), ovviamente del tutto estranea a quell'organismo. In questi topi, in linea di massima, non si è determinata una reazione immunitaria di cellule T dirette contro il tessuto pancreatico e neppure sono state dimostrate in vitro cellule T capaci di reagire contro gli antigeni transgenici; in altri termini, in questi topi è stata dimostrata una tolleranza ad antigeni che sicuramente non erano espressi nel timo, cioè una tolleranza periferica.
La tolleranza periferica differisce tuttavia dalla tolleranza centrale per il meccanismo della sua genesi: i cloni T autoreattivi non sono distrutti dai meccanismi della tolleranza periferica, come accade invece nel caso dei cloni sottoposti alla selezione negativa nel timo, ma sono semplicemente inattivati. Questo meccanismo di inattivazione, detto 'anergia', permette la sopravvivenza dei cloni di linfociti T interessati, ma li rende incapaci di reagire nei confronti degli antigeni anche se conservano i recettori idonei a tale scopo. Il meccanismo mediante il quale determinati antigeni espressi dai tessuti periferici possono inattivare i cloni T di specificità corrispondente, maturati nel timo e disseminati nel sistema periferico, non è completamente noto; probabilmente è legato al modo col quale questi antigeni sono presentati e alla mancanza, nelle cellule presentanti, di molecole necessarie per la piena attivazione dei linfociti T. Sono state compiute importanti osservazioni che nella determinazione della tolleranza periferica chiamano in causa lo stato di attivazione delle cellule che hanno precisamente la funzione di presentare gli antigeni alle cellule T, e cioè le cellule dendritiche.
Un'interessante ipotesi chiama in causa, nella tolleranza periferica, la molecola CTLA-4. Questa molecola è omologa del co-recettore CD28 e come questo reagisce con alta affinità con le molecole B-7 che si trovano sulle cellule che presentano l'antigene; la sua funzione fondamentale è quella di regolare l'omeostasi dei linfociti T dopo la stimolazione antigenica. Infatti, la molecola CTLA-4 è espressa, a livelli crescenti, nei linfociti attivati e in ultima analisi blocca la continuazione della risposta dei linfociti T, sia interferendo col segnale positivo dato dal CD28, sia provvedendo direttamente a fornire un segnale negativo. In topi nei quali il gene per la molecola CTLA-4 sia stato inattivato, le risposte agli antigeni non hanno termine e gli animali muoiono in poche settimane in seguito a un'invasione generalizzata di linfociti proliferanti. È possibile, peraltro, che la molecola CTLA-4 intervenga nel bloccare sul nascere le risposte dei linfociti T contro autoantigeni, in quanto questi spesso vengono presentati da cellule con limitata espressione di B-7, condizione, questa, che favorisce le molecole CTLA-4 rispetto alle molecole CD28.
Poiché i cloni anergici non proliferano, ma neppure scompaiono, si capisce come la tolleranza periferica sia meno stabile della centrale. Infatti, nei topi transgenici che esprimono nel pancreas l'antigene virale T SV40, per esempio, la tolleranza periferica è imperfetta in quegli animali nei quali l'antigene viene espresso più tardivamente durante lo sviluppo. In buona sostanza, la tolleranza periferica è una garanzia solo parziale contro l'autoimmunità e molte fra le più frequenti malattie autoimmuni, come, per esempio, quelle della tiroide, sono probabilmente dovute al venir meno, a un certo punto della vita, di alcuni processi di tolleranza periferica. È interessante notare che alcuni polimorfismi delle molecole CTLA-4 sono stati associati con l'immunità antitiroide della malattia di Graves. Nella tolleranza periferica sono stati segnalati, in alcuni casi particolari, meccanismi di delezione clonale delle cellule T: per esempio, in topi transgenici nei quali l'antigene introdotto (la molecola allogenica H-2Kb) era sotto il controllo del promotore della metallotioneina ed era espresso nel fegato. In questi casi, la delezione delle cellule autoreattive avviene probabilmente mediante l'attivazione del sistema Fas/Fas L, come nel caso del lupus eritematoso sistemico dei topi MLR-MLR.
Quando la tolleranza fallisce e dopo che la reazione autoimmunitaria è iniziata, il sistema immunitario ha un'ultima possibilità per evitare o limitare il danno che ne può derivare, attivando l'immunosoppressione. Tuttavia, questa non va considerata come un processo destinato esclusivamente al controllo dell'autoimmunità, ma più in generale come un meccanismo di autoregolazione del sistema immunitario, capace di limitare le reazioni eccessive o troppo prolungate; molti casi di immunosoppressione sono stati infatti osservati non soltanto in malattie autoimmuni, ma anche in malattie infettive croniche, come la lebbra, nelle quali una reazione immunitaria diretta contro gli antigeni del microrganismo può indirettamente danneggiare e provocare mutilazioni nell'individuo infettato. L'immunosoppressione è operata da linfociti T, specialmente della sottopopolazione CD25 CD4, alla quale è stato dato il nome di 'cellule regolatrici', i quali agiscono eliminando o rendendo anergici altri linfociti T impegnati in una determinata reazione immunitaria.
L'immunosoppressione è specifica, ma le basi molecolari di questa specificità non sono chiare. Vi sono due possibilità: o le cellule T soppressive reagiscono con l'antigene e liberano linfochine (IL-4, IL-10, TGF-β) che sopprimono l'attivazione da parte dello stesso antigene delle altre cellule T; oppure le cellule soppressive riconoscono direttamente il recettore clonale delle cellule T che sono bersaglio della soppressione ed esercitano su queste un'azione citotossica o anergizzante. Perché le cellule T soppressive possano riconoscere il recettore clonale delle cellule T bersaglio, è necessario che queste operino una elaborazione del loro recettore e la presentazione di peptidi corrispondenti sulle loro molecole MHC di classe I, secondo le regole generali per la presentazione di antigeni capaci di essere riconosciuti da parte di altre cellule T: in questo caso, quindi, le cellule T soppressive riconoscono i peptidi derivati dai recettori clonali delle cellule T che sono bersaglio della soppressione. Esperimenti di immunizzazione con peptidi derivati dai recettori T confermano questo meccanismo in alcune condizioni, comprese alcune malattie autoimmuni sperimentali. Concludiamo sottolineando l'importanza centrale delle cellule T per l'autoimmunità: infatti, anche l'autoimmunità da cellule B, cioè da autoanticorpi, richiede nella maggioranza dei casi la rottura della tolleranza delle cellule T che sono necessarie come coadiuvanti dei linfociti B nella produzione di anticorpi contro gli antigeni proteici, ivi compresi gli autoantigeni.
I linfociti B: selezione positiva e selezione negativa
I linfociti B, come i linfociti T, producono i loro recettori clonali per gli antigeni (le immunoglobuline di membrana), sotto il controllo di geni che subiscono processi di rielaborazione somatica secondo regole che includono elementi casuali. Pertanto, come i linfociti T, costituiscono una popolazione che deve essere sottoposta a processi di selezione, sia positiva sia negativa. Le modalità e le sedi di questa selezione sono però molto diverse da quelle dei linfociti T, essenzialmente in rapporto alle seguenti fondamentali differenze funzionali tra linfociti B e T: (a) i recettori dei linfociti B reagiscono contro l'antigene direttamente, senza necessità di preparazione dell'antigene e di presentazione dei suoi frammenti da parte delle molecole MHC; (b) i geni per i recettori delle cellule B subiscono processi di rielaborazione non solo nell'organo centrale dove queste cellule maturano (il midollo osseo nei Mammiferi), ma anche in particolari sedi dei tessuti linfatici periferici (i centri germinativi dei follicoli linfatici). La stimolazione dei linfociti B non è operata direttamente dagli antigeni proteici, ma richiede l'intervento di particolari linfociti T con specificità per lo stesso antigene; i linfociti B devono preparare frammenti dell'antigene stesso e presentarli alla cellula T cooperante, mediante le molecole MHC di classe II delle quali sono provvisti.
Consideriamo pertanto la selezione dei linfociti B a tre diversi livelli: nel midollo osseo, nei centri germinativi dei follicoli linfatici e nella periferia dei tessuti linfatici. Nel midollo osseo le cellule B non richiedono, come i linfociti T nel timo, una selezione positiva sulla base dei loro recettori completi (una funzione in tal senso di un recettore incompleto, il recettore pre-B, ha tutt'altro significato). Invece, una forma di selezione negativa è possibile, come è stato dimostrato in topi transgenici nei quali era stata indotta la coesistenza di un antigene MHC di classe I e di una maggioranza di linfociti B esprimenti nella loro membrana un recettore immunoglobulinico (prodotto sotto il controllo del transgene) diretto contro lo stesso antigene: in questi esperimenti, la maturazione delle cellule B autoreattive si arresta. Il destino di queste cellule può essere diverso: o la morte per attivazione di un programma di autodistruzione, chiamato 'apoptosi' (un termine di derivazione greca che indica la caduta di un frutto maturo dall'albero), o il recupero mediante l'espressione di una catena immunoglobulinica leggera diversa da quella impiegata nel primitivo recettore autoreattivo. Il midollo osseo produce così una popolazione di linfociti B largamente purgata dalle cellule autoreattive, ma provvista di recettori capaci di reagire con antigeni estranei all'organismo, generalmente con bassa affinità.
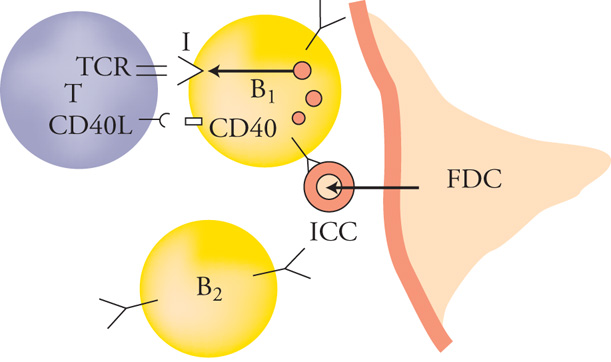
La trasformazione di queste cellule in elementi con recettori ad alta affinità ‒ quindi capaci di produrre in ultima analisi anticorpi efficienti ‒ avviene nei centri germinativi dei follicoli linfatici sotto la selezione positiva degli antigeni estranei all'organismo. In questa sede, per l'attivazione di particolari enzimi, il DNA che controlla la parte variabile delle catene pesanti e leggere delle immunoglobuline subisce rapidi processi di mutazione che modificano la capacità della cellula di reagire con l'antigene esterno concentrato nel centro germinativo. Poiché nei centri germinativi l'antigene è presente sotto forma di complessi antigene-anticorpo depositati sulla superficie delle cellule dendritiche (non linfoidi) del centro stesso, si determinano le condizioni per una rapida selezione positiva delle cellule B con recettori per l'antigene ad affinità superiore a quella della media degli anticorpi sino allora prodotti, probabilmente secondo un meccanismo simile a quello schematizzato nella fig. 3.
Naturalmente il processo di ipermutazione può generare, nel centro germinativo, anche cellule B con recettori autoreattivi, ma queste cellule non ricevono il necessario aiuto dalle cellule T presenti in sede e vanno incontro a un processo di autodistruzione per picnosi (apoptosi), del quale vi è ampia evidenza microscopica in tutti i centri germinativi attivi. In questo modo, a livello dei centri germinativi ha luogo anche un'estesa selezione negativa delle cellule B autoreattive, per effetto della mancata cooperazione con le cellule T specifiche per l'antigene estraneo. Infine, nei tessuti periferici l'incontro di cellule B mature con un antigene proteico per il quale hanno un'alta affinità può risultare, in assenza di aiuto T-specifico, nella anergia dei cloni di cellule B, con un processo di tolleranza periferica non molto dissimile dall'analogo esito in anergia al quale possono andare incontro le cellule T. In definitiva, sia le cellule B sia le cellule T sono sottoposte a controlli a diversi livelli, che normalmente riescono a impedire l'autoimmunità. Tali controlli, tuttavia, non possono essere così stringenti da orientare il sistema immunitario verso lo stato opposto, cioè l'immunodeficienza, e alcune maglie deboli della tolleranza immunitaria persistono e possono amplificarsi col processo di invecchiamento del sistema immunitario. Questi punti deboli possono in definitiva aprire la strada a processi autoimmunitari di varia natura, che hanno come bersaglio antigeni diversi dell'organismo, provocando la comparsa di differenti malattie autoimmuni.
L'autoimmunità
Caratteristiche generali delle malattie autoimmuni
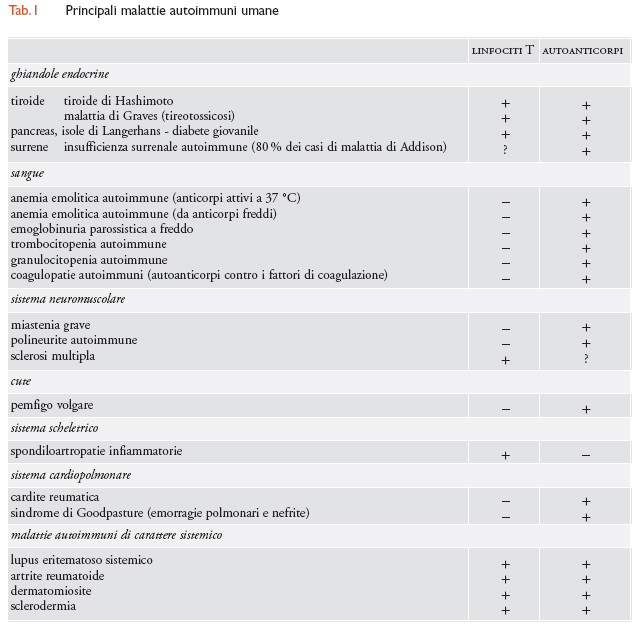
Nell'uomo sono state osservate più di quaranta diverse malattie autoimmuni; alcune di queste si sviluppano spontaneamente anche negli animali, mentre altre possono esservi indotte mediante procedimenti di immunizzazione. La frequenza delle malattie autoimmuni nella specie umana è alta, interessando quote dal 5 al 7% della popolazione, con variazioni geografiche per certe malattie. Alcune di queste, come il diabete di tipo I e la sclerosi multipla, sono allo stesso tempo frequenti e seriamente invalidanti, e costituiscono quindi un grave problema di salute pubblica. Nella tab. 1 è riportato un elenco delle principali malattie autoimmuni umane, suddivise in base ai sistemi che interessano e con l'indicazione del ruolo dei linfociti T autoreattivi e/o dei linfociti B (autoanticorpi) nella determinazione del danno. Alcune caratteristiche generali delle malattie autoimmuni meritano attenzione. Anzitutto la loro frequente associazione, più o meno stretta, con determinati alleli dei geni che controllano le molecole di istocompatibilità (MHC). Questa è una caratteristica delle malattie autoimmuni evidentemente legata alla necessità, per i linfociti T autoreattivi, di riconoscere i determinanti degli autoantigeni proteici presentati nella tasca delle molecole MHC: alcuni alleli MHC formano complessi con i peptidi derivati dagli autoantigeni, mentre altri non lo fanno o formano complessi più labili.
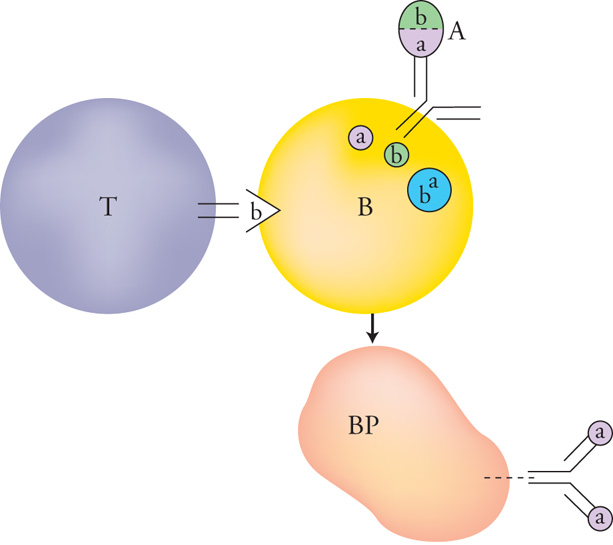
Sfortunatamente non sono ancora stati individuati, nella maggior parte dei casi, i determinanti, o epitopi, implicati nelle diverse risposte autoimmuni umane. Esistono probabilmente numerosi epitopi per ogni malattia, e ciò spiega perché singoli alleli MHC siano in prevalenza, ma mai in modo obbligatorio, associati con singole malattie, anche se in misura diversa (il cd. 'rischio relativo'). Il caso più evidente di associazione MHC-malattia è quello dell'allele HLA-B27 di classe I con le spondiloartropatie infiammatorie; in questo caso il rischio relativo supera il 90%. Evidentemente, nelle spondiloartriti sono in gioco linfociti T autoreattivi della sottopopolazione CD8+, quelli cioè che reagiscono con peptidi presentati dalla classe I delle molecole MHC. Tuttavia, anche in questo caso gli epitopi (peptidi) degli autoantigeni non sono stati ancora identificati con sicurezza. Un'altra caratteristica comune a molte delle malattie autoimmuni, specialmente quelle organo-specifiche, è quella di presentarsi dopo un episodio infettivo batterico o virale: possiamo pensare che ciò avvenga per la rottura parziale della tolleranza periferica determinata da una somiglianza (mimicry) fra gli antigeni dell'agente infettante e quelli propri dell'organismo infettato. Un caso specifico è quello relativo alla produzione di autoanticorpi da parte di cellule B, che ricevono un aiuto improprio dai linfociti T CD4+ mediante una molecola, o un complesso, che possiede allo stesso tempo sia determinanti capaci di interagire con le immunoglobuline potenzialmente autoreattive dei linfociti B, sia determinanti estranei all'organismo, per i quali non vi è tolleranza nella popolazione di linfociti T (fig. 4).
Meccanismi del danno tissutale
Infine, un accenno ai meccanismi del danno tissutale causato dalle reazioni autoimmuni, non differenti, in linea di principio, dai meccanismi mediante i quali il sistema immunitario danneggia e quindi elimina gli agenti invasori. I linfociti T CD4+ determinano, laddove incontrano gli autoantigeni, una reazione infiammatoria di ipersensibilità ritardata (DTH, Delayed-type hypersensibility), nella quale il danno ai tessuti interessati è legato alla liberazione di una serie di citochine e all'attivazione dei macrofagi. I linfociti citotossici CD8+ uccidono direttamente le cellule bersaglio, esattamente come agirebbero, per esempio, nei confronti di cellule infettate da un virus. Gli autoanticorpi provocano danni per vie diverse: possono determinare, per esempio, la lisi dei globuli rossi mediante l'attivazione del complemento, oppure possono agglutinare i trombociti e determinarne il sequestro nel sistema reticoloendoteliale.
Gli autoanticorpi diretti contro recettori cellulari possono agire bloccando la funzione di questi recettori, oppure, al contrario, possono provocare un danno stimolando (funzione agonista) i recettori stessi. Nell'autoimmunità antitiroide, per esempio, autoanticorpi diretti contro il recettore dell'ormone stimolante la tiroide (TSH) possono causare un ipotiroidismo, come nella malattia di Hashimoto; oppure anticorpi un poco diversi, diretti contro lo stesso recettore, possono determinare una stimolazione della funzione tiroidea, come nella malattia di Graves. Inoltre, gli autoanticorpi coesistono solitamente con un'elevata quantità di autoantigeni. Se l'affinità della reazione è sufficiente, si verificano le condizioni ideali per la produzione di abbondanti immunocomplessi, i quali, da soli o mediante la fissazione del complemento, sono causa di una rilevante patologia, specialmente a livello dei vasi, dei reni e delle articolazioni. Malattie autoimmuni che sono insorte spontaneamente in alcuni animali da laboratorio, o che possono essere indotte in questi mediante appropriata immunizzazione, forniscono elementi essenziali per la comprensione delle corrispondenti malattie umane e dell'autoimmunità in generale.
Bibliografia
Alam 1996: Alam, S. Mohammed e altri, T-cell-receptor affinity and thymocyte positive selection, ‟Nature", 331, 1996, pp. 616-620.
Anderson 2002: Anderson, Mark S. e altri, Projection of an immunological self shadow within the thymus by the aire protein, "Science", 298, 2002, pp. 1395-1401.
Ashton-Rickardt, Tonegawa 1994: Ashton-Rickardt, Philip G. - Tonegawa, Susumu, A differential-avidity model for T-cell selection, ‟Immunology today", 15, 1994, pp. 362-366.
Blackman 1990: Blackman, Marcia - Kappler, John - Marrack, Phillipa, The role of the T cell receptor in positive and negative selection of developing T cells, ‟Science", 248, 1990, pp. 1335-1341.
Boehmer 1994: von Boehmer, Harald, Positive selection of lymphocytes, ‟Cell", 76, 1994, pp. 219-228.
Chen 2006: Chen, Min e altri, Dendritic cell apoptosis in the maintenance of immune tolerance, "Science", 311, 2006, pp. 1160-1164.
Cohen, Young 1991: Cohen, Irun R. - Young, Douglas B., Autoimmunity, microbial immunity and the immunological homunculus, ‟Immunology today", 12, 1991, pp. 105-110.
Koopman, Pals 1992: Koopman, Gerrit - Pals, Steven T., Cellular interactions in the germinal center: role of adhesion receptors and significance for the pathogenesis of AIDS and malignant lymphoma, ‟Immunological reviews", 126, 1992, pp. 21-45.
Kuchroo 1995: Kuchroo, Vijay K. e altri, B7-1 and B7-2 costimulatory molecules activate differentially the Th1/Th2 developmental pathways: application to autoimmune disease therapy, ‟Cell", 80, 1995, pp. 707-718.
Laufer 1996: Laufer, Terri M. e altri, Unopposed positive selection and autoreactivity in mice expressing class II MHC only on thymic cortex, ‟Nature", 383, 1996, pp. 81-85.
Linsk 1989: Linsk, Richard - Gottesman, Max - Pernis, Benvenuto, Are tissues a patch quilt of ectopic gene expression?, "Science", 246, 1989, pp. 261.
Nossal 1994: Nossal, Gustav J.V., Negative selection of lymphocytes, ‟Cell", 76, 1994, pp. 229-239.
Schwartz, Datta 1989: Schwartz, Robert S. - Datta, Syamal K., Autoimmunity and autoimmune diseases, in: Fundamental immunology, 2. ed., edited by William E. Paul, New York, Raven, 1989, pp. 820-866.
Sercarz, Krzych 1991: Sercarz, Eli - Krzych, Ursula, The distinctive specificity of antigen-specific suppressor T cells, ‟Immunology today", 12, 1991, pp. 111-118.
Sinha 1990: Sinha, Aminesh A. - Lopez, M. Teresa - McDevitt, Hugh O., Autoimmune diseases: the failure of self tolerance, ‟Science", 248, 1990, pp. 1380-1388.
Waterhouse 1995: Waterhouse, Paul e altri, Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4, ‟Science", 270, 1995, pp. 985-988.