
Apprendimento canoro
Apprendimento canoro
Gli uccelli comunicano fra loro mediante un repertorio di vocalizzazioni. I richiami sono di solito brevi e monosillabici, i canti sono invece pattern sonori più estesi. In quasi la metà delle 9000 specie di uccelli il canto è appreso, in particolare negli oscini, la cui caratteristica è di avere organi complessi e uno specifico circuito del prosencefalo che consente l'apprendimento. Gli ormoni androgeni sono molto importanti per lo sviluppo del canto, e il circuito cerebrale a essi associato mostra spesso un dimorfismo sessuale. A differenza dei richiami, i canti degli oscini non si sviluppano normalmente se i giovani maschi non hanno la possibilità di ascoltare i canti degli adulti. L'importanza dell'apprendimento è dimostrata dalla presenza di dialetti canori locali paragonabili a quelli umani. La produzione del canto è guidata da tracce acustiche in memoria contenute nel cervello che codificano le caratteristiche specie-specifiche e consentono il processo di apprendimento. L'imitazione dei suoni è rara in natura e gli uccelli, se hanno la possibilità di scelta, preferiscono imparare il canto dei conspecifici.
Introduzione
Tutti gli uccelli possiedono un repertorio di vocalizzazioni per comunicare fra loro informazioni su pericoli, cibo, sesso, spostamenti in gruppo e per molti altri scopi. Nell'ambito di questo repertorio, costituito da una dozzina di segnali o poco più, si può operare una distinzione fra i richiami, in genere brevi e spesso monosillabici, e il canto, che è un pattem di vocalizzazione più prolungato, talora rauco, spesso tonale e melodico, non di rado fonte di piacere per l'ascoltatore umano (Catchpole e Slater, 1995). Secondo questa definizione, praticamente tutti gli uccelli possiedono qualcosa che possiamo chiamare un canto, sia che si tratti del canto di un gallo sia che si tratti della melodia di un usignolo. In quasi la metà delle 9000 specie di uccelli note il canto è appreso. Gran parte delle specie che apprendono il canto appartiene ai passeriformi, uno dei 27 ordini in cui i tassonomi classificano gli uccelli. Sono esclusi i caradriformi, i galliformi, i picchi, i cuculi, i colombi, i rapaci diurni e notturni e tutti gli uccelli marini e di acqua dolce. Gli unici gruppi che hanno i requisiti necessari per apprendere il canto sono i pappagalli, i colibrì e, in particolare, il grande sottordine dei passeriformi noto come oscini, caratterizzato dalla complessità degli organi vocali e da peculiari circuiti neuronali del prosencefalo, evolutisi espressamente per l'apprendimento del canto. l pappagalli, e forse i colibrì, sembrano aver ottenuto indipendentemente la stessa capacità con circuiti nervosi equivalenti, ma neuroanatomicamente distinti (Striedter, 1994).
Il canto è in genere, ma non sempre, una prerogativa del sesso maschile dipendente dagli ormoni androgeni (Brenowitz et al., 1985; Brenowitz e Amold, 1989). Nelle specie che apprendono il canto il circuito neuronale interessato è spesso sessualmente dimorfo ed è ormai disponibile un'estesa letteratura sul grado e sul significato funzionale di questo dimorfismo e sulle basi del suo sviluppo (Nottebohm e Amold, 1976; Amold et al., 1986; Amold, 1990; Devoogd, 1991; Brenowitz e Kroodsma, 1996). Il maggiore sviluppo del sistema deputato al canto nel cervello del maschio implica chiaramente una maggiore propensione all' apprendimento canoro; la correlazione tra la dimensione dei nuclei del sistema vocale, la presenza del canto e l'ampiezza del repertorio vocale si estende non solo al confronto tra i sessi, ma anche a quello tra specie, sotto specie e addirittura individui (Nottebohm et al., 1981; Canady et al., 1984; Devoogd et al., 1993).
A differenza dei richiami, che solo in qualche caso sono appresi e sono tipicamente innati, i canti degli oscini si sviluppano sempre in maniera anormale se un maschio giovane è allevato senza poter ascoltare gli adulti. Una conseguenza comune della dipendenza dall'apprendimento è la presenza di dialetti locali dei canti che spesso variano su scala geografica in maniera simile a quanto si osserva per i dialetti del linguaggio umano (Baker e Cunningham, 1985). Il loro grado di differenza dipende da fattori quali i pattern di dispersione dal luogo natio e il periodo in cui avviene l'apprendimento canoro. Quando i dialetti sono ben definiti la regola generale sembra essere che i canti, come il dialetto proprio di un dato individuo, sono particolarmente efficaci come segnali vocali sia per i maschi territoriali sia per le femmine alla ricerca di un partner (Baker et al., 1981; 1987; Baker, 1983; Balaban, 1988). Il dialetto di un uccello può corrispondere o meno a quello prevalente nell' area dove è nato.
Si possono scoprire molte analogie tra l'apprendimento del canto negli uccelli e l'acquisizione del linguaggio nell'uomo. Non c'è primate non umano i cui pattern vocali siano trasmessi culturalmente di generazione in generazione. A parte i cetacei, anch'essi in grado di apprendere repertori vocali, gli uccelli costituiscono il più valido modello animale per studiare le basi comportamentali, ormonali e neuronali della plasticità vocale (Konishi, 1985; Marler, 1991; Nottebohm, 1993).
Meccanismi cerebrali specifici per l'apprendimento del canto
A partire dalla metà degli anni Settanta, quando F. Nottebohm, T.M. Stokes e C.M. Leonard (1976) scoprirono il sistema deputato al canto nel cervello del canarino, sono stati compiuti notevoli progressi nella comprensione dei meccanismi neuronali specializzati che mediano la plasticità vocale degli uccelli. Tuttavia, molte questioni importanti rimangono ancora irrisolte.
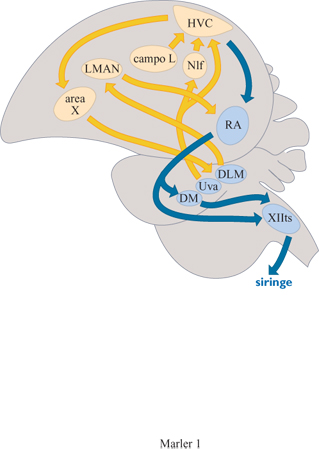
Vi sono due requisiti funzionali principali per il controllo della produzione di suoni. In primo luogo, deve costituirsi un' appropriata configurazione degli organi per la produzione di suoni, in particolare la siringe e il tratto vocale postsiringe. In secondo luogo, deve essere generato un pattern di flusso d'aria appropriato per far vibrare tali organi. Questi sistemi di controllo sono necessari sia se il canto è appreso sia se è innato. Come si poteva prevedere, parte del circuito responsabile è relativamente conservativo, situato nel mesencefalo e nel midollo allungato (Wild, 1994). Gli ulteriori requisiti per l'apprendimento canoro sono forniti da due importanti circuiti nel prosencefalo (fig. 1), uno specializzato per sostenere il processo effettivo di apprendimento, la cosiddetta via del prosencefalo anteriore e l'altro, la via motoria, deputato al controllo della produzione dei suoni appresi (Nottebohm, 1993). Mentre un'alterazione della via motoria causa sempre la distorsione e persino l'eliminazione dei canti appresi, la via del prosencefalo anteriore sembra essere ridondante, eccetto che durante lo sviluppo effettivo del canto (Bottjer et al., 1984). Come per i canti innati, la via motoria per i canti appresi deve aver accesso al tratto vocale e alla muscolatura della siringe, localizzata in una parte della via motoria, e anche al sistema respiratorio, a livello del nucleo robusto dell'archistriato (Vicario, 1991). I requisiti del circuito della via motoria sono complicati dall' esistenza di due vibratori semiindipendenti situati nella siringe degli uccelli canori, che richiedono una coordinazione precisa essendo entrambi coinvolti nella produzione di canti appresi (Greenewalt, 1968; Suthers, 1990).
L'udito degli uccelli è stato studiato in dettaglio in relazione al comportamento vocale (Dooling, 1982). Molte parti del sistema vocale rispondono alla stimolazione sonora oltre ad adempiere alle loro funzioni motorie. Stranamente, uno degli stimoli più forti, specialmente nel centro vocale superiore dei maschi adulti, ma anche altrove, è il canto cristallizzato dell 'uccello stesso che lo distingue individualmente (Margoliash, 1986). Questa scoperta ha portato all'ipotesi plausibile che i maschi adulti di alcune specie utilizzino una retro azione uditiva del loro proprio canto come metro per calibrare la potenza dei canti di altri individui quando li ascoltano, specialmente se si tratta di vicini conspecifici e di estranei.
La sintonizzazione del sistema del canto di un dato maschio sul proprio canto individuale, comprese tutte le sue variazioni personali, emerge solo quando il canto cristallizza (Doupe, 1993; Volman, 1993). Purtroppo è stata prestata poca attenzione alla sensibilità di risposta a stimoli canori del cervello di individui giovani prima della produzione del canto plastico e cristallizzato. l pattern di re attività uditiva del cervello di un uccello canoro durante la prima fase di memorizzazione dell' apprendimento sono praticamente sconosciuti (Whaling et al., 1995). Le analisi comportamentali effettuate durante questa età dimostrano una reattività innata a molte caratteristiche del canto, specialmente a quelle degli uccelli della stessa specie (Nelson e Marler, 1993; Whaling et al., 1997). Le caratteristiche del processo di memorizzazione del canto sono compatibili con il coinvolgimento di un set di predisposizioni acustiche innate che ciascuna specie ha a disposizione, e che fungono da guida per il processo di apprendimento canoro selettivo. l meccanismi neuronali responsabili sono ancora in attesa di indagini neurofisiologiche e finché questi non saranno chiariti non potremo avere la piena comprensione delle basi fisiologiche dell'apprendimento canoro. Attualmente non siamo in grado di spiegare come un giovane maschio sia capace di memorizzare i canti, così come non sappiamo dove vengano immagazzinati nel cervello per guidare poi la produzione canora.
Un'importante caratteristica degli uccelli è il continuo sviluppo di nuovi neuroni in molte parti del cervello, incluso il sistema deputato al canto, sia nell'infanzia sia in fase adulta (Goldman e Nottebohm, 1983). Sebbene non sia chiara l'importanza della neurogenesi nell'apprendimento canoro, vi sono molti indizi suggestivi tra cui la dimostrazione del fatto che, in una sorta di tour de force tecnico, alcuni nuovi neuroni sono effettivamente inseriti nel sistema del canto e si attivano lì (Paton e Nottebohm, 1984; Alvarez-Buylla et al., 1990). Si potrebbe ipotizzare che il ricambio neuronale nella via motoria prepari la strada per la plasticità del canto che alcuni uccelli, come il canarino (Nottebohm e Nottebohm, 1978), mostrano da adulti. Come accennato, un'altra sede deputata ad attività legate alla produzione del canto si trova nei circuiti uditivi del prosencefalo e riguarda l'elaborazione percettiva degli stimoli vocali. Uno di questi, il neo striato caudale mediale, è stato identificato come il sito dell'attivazione genica in risposta a stimolazione da parte di individui della stessa specie (Mello et al., 1992; Mello e Clayton, 1994) e come sede di quelle modificazioni che dipendono dall'esperienza nella reattività elettro fisiologica agli stimoli canori (Vicario e Simpson, 1995). Anche circuiti esterni al sistema del canto potrebbero essere coinvolti nell'apprendimento canoro più di quanto si pensi.
Periodi sensibili per l'apprendimento
Nello sviluppo del canto, come nell'ontogenesi di altri tipi di comportamenti, la rapidità e la precisione con cui l'apprendimento si manifesta variano durante lo sviluppo da una fase all'altra del ciclo vitale. Tutti gli uccelli canori sembrano condividere una sequenza simile di fasi nel processo di apprendimento. In primo luogo, vi è la fase di acquisizione, spesso ristretta a un breve periodo, quando l'uccello ascolta i canti e ne custodisce una parte nella memoria. Dopo l'immagazzinamento, per un periodo che varia da giorni a mesi a seconda della specie, i canti sono richiamati dalla memoria e cominciano le imitazioni. Le rese sono relativamente fedeli al modello originale in certe specie, ma in altre se ne discostano piuttosto radicalmente e in esse l'inventiva vocale svolge un ruolo significativo.
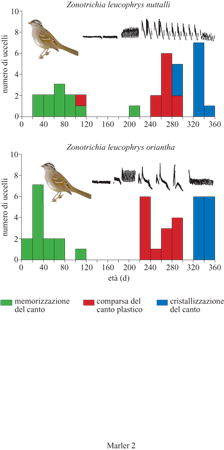
La separazione nel tempo tra l'acquisizione, o fase sensoria, e la produzione, o fase sensomotoria, che termina con la produzione del canto cristallizzato dell'adulto è molto importante e significativa. In molti uccelli la sequenza si verifica solo una volta, spesso nelle prime fasi dell'adolescenza. In questi casi i pattern canori dell' adulto rimangono in seguito fissati e la loro produzione cresce e cala alternativamente dopo ciascuna stagione riproduttiva. In altre specie la sequenza ricorre ripetutamente durante la fase adulta, cosicché i pattern possono continuare a cambiare. Anche specie filogeneticamente affini come i canarini e i passeri possono mostrare differenze sotto questo aspetto. Molte specie di passeri presentano un periodo sensibile per l'acquisizione del canto che inizia a circa due settimane di età, poco dopo aver lasciato il nido, e termina da sei a otto settimane dopo (Nelson et al., 1995). La durata precisa varia anche in differenti popolazioni della stessa specie (fig. 2). I periodi sensibili per l'apprendimento non sono fissi, ma mutano entro certi limiti e dipendono da fattori come l'intensità della stimolazione canora, la possibilità di accesso a un 'tutore' vivo e interattivo (Baptista e Petrinovich, 1986) e fattori fisiologici quali gli stati ormonali che variano con la stagione. Il periodo di acquisizione può talora protrarsi in assenza dello stimolo (Kroodsma e Pickert, 1980), come se la fine della fase sensoria fosse ritardata fmché non fosse stata provata un'adeguata stimolazione. Se i giovani nascono tardi nella stagione e l'attività canora è già terminata per quell'anno, il periodo sensibile può estendersi in alcune specie fino alla primavera successiva; questo ritardo non si osserva in tutti i casi.
Ontogenesi del canto appreso

Al contrario di quanto avviene nelle specie che apprendono il canto, quelle che non lo apprendono sviluppano un canto normale se gli animali sono allevati in isolamento e anche nel caso che siano stati resi sordi (Kroodsma, 1984; Kroodsma e Konishi, 1991). Ciò vale per un colombo maschio, un gallo e anche per quei passeriformi non oscini come i pigliamosche del nuovo mondo (fig. 3). In tutti questi casi, quando un giovane uccello inizia a cantare, i suoi primi sforzi sono chiaramente identificabili come versioni immature di quello che alla fine sarà il canto normale dell'adulto. I primi tentativi di canto possono essere rumorosi e frammentati, ma vi è sempre una chiara progressione lineare nello sviluppo dei canti innati.
Nelle specie che apprendono il loro canto, la progressione è piuttosto differente nel corso dello sviluppo. Essa inizia con un unico pattern vocale detto sottocanto, un gorgheggio sommesso e variabile che può iniziare a manifestarsi già a tre o quattro settimane di età. Questa fase ricorda lo stadio precoce del mormorio (babbling) che si osserva durante lo sviluppo del linguaggio umano (Locke, 1993). Tipicamente non sembra somigliare al canto maturo, sebbene conosciamo la sua struttura e funzione meno di ogni altro aspetto dello sviluppo del canto. Non vi sono elementi di imitazione nel sottocanto. Nelle specie che apprendono il canto si osserva un tipo di metamorfosi vocale tra sottocanto e stadi successivi dello sviluppo canoro. La struttura amorfa e l'organizzazione rumorosa dello spettro del sottocanto suggeriscono che esso può avere un ruolo nell'acquisizione delle capacità motorie generali del canto, per far fronte alla complessità della siringe degli oscini e anche nell'affmare la capacità di guidare la voce con l'orecchio, un requisito indispensabile per l'imitazione vocale (Thorpe, 1961; Nottebohm, 1972; Marler e Peters, 1982b). La durata del periodo del sottocanto varia molto anche nella stessa specie e dipende dalle storie individuali. In uccelli sordi e socialmente isolati può durare settimane, ma un individuo cresciuto accanto a un adeguato 'tutore' è in grado di progredire rapidamente dal sottocanto alla fase successiva, il canto plastico, in pochi giorni.

Nel canto plastico compaiono per la prima volta i segni della struttura del canto maturo. Emergono e iniziano a stabilizzarsi pattern canori precedentemente memorizzati, dapprima gradualmente, poi rapidamente, fino a ottenere il canto cristallizzato. A questo punto il giovane maschio è pronto a intraprendere pienamente la stagione del canto maturo (fig. 4).
Sovrapproduzione, scarto e apprendimento basato sull'azione
L'apprendimento canoro è stato tradizionalmente considerato come basato sulla memoria. l canti sono acquisiti e memorizzati e in seguito utilizzati per guidare lo sviluppo motorio tramite retroazione uditiva, senza alcuna pratica al momento della memorizzazione. Un altro tipo di plasticità vocale, che coinvolge solo la memoria operativa e svolge un ruolo chiave nella pratica motoria, si osserva durante la fase del canto plastico.
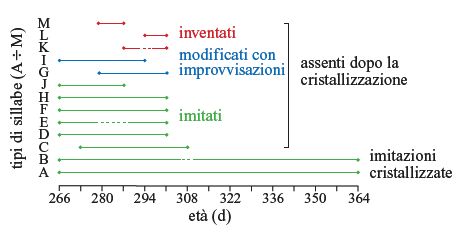
Molte specie che apprendono il canto manifestano una sovrapproduzione durante il canto plastico e generano più di quanto necessiterebbero per il canto maturo, scartando poi l'eccesso di canti plastici nel momento in cui si avvicina il periodo della 'cristallizzazione' (fig. 5).
Alcuni suoni in eccesso sono rnventati, ma molti sono imitati e la ricchezza della prima esperienza canora costituisce un fattore rilevante nel determinare la quantità di repertorio prodotto in eccesso (Nelson et al., 1996). Tuttavia, la sovrapproduzione e lo scarto si verificano anche in uccelli con tutore e allevati in isolamento individuale in laboratorio (Marler e Peters, 1982a). In tal caso la decisione su quali parti scartare sembra essere casuale, mentre in natura è mediata dall'esperienza sociale.
Nei passeri la fase di scarto (attrition), nella quale i suoni in eccesso sono scartati dal repertorio, coincide col momento in cui i giovani maschi stabiliscono il loro primo territorio. Non appena si instaura la competizione con gli adulti, questi canti del giovane uccello, che corrispondono, anche se solo approssimativamente, alle tipologie sonore di rivali più anziani, sono i più efficaci nello stimolare controcanti e forse anche il comportamento di difesa del territorio (Beecher, 1996). l giovani maschi tendono a conservare e a cristallizzare quei canti plastici che sono le imitazioni più efficaci di quelli dei loro rivali e a scartare il resto.
Gli effetti delle interazioni tra maschi, sia reali sia simulate, sulla modulazione del canto in seguito a scarti sono stati descritti in diverse specie di passeri (Nelson, 1992; Nelson e Marler, 1994). In un'altra specie, il molotro nero, sono le risposte delle femmine a modellare le decisioni del maschio su quali elementi del canto conservare (West e King, 1988). Il processo mediante il quale un repertorio di svariati pattem motori è rielaborato selettivamente tramite retro azione sociale (Marler e Nelson, 1993), è stato definito apprendimento basato sull'azione (action based learning). Esistono analogie con lo sviluppo di altri tipi di comportamenti motori come il gioco. Processi di rielaborazione selettiva dipendenti dall'uso potrebbero essere ampiamente diffusi nello sviluppo di molti tipi di attività comportamentali. Le attività scartate dal repertorio potrebbero esse stesse possedere un significato funzionale. Per esempio, i canti eseguiti in gioventù e poi scartati possono fornire una banca di memoria potenziale per il futuro, forse consultata per valutare i canti di altri uccelli. Nelle specie che mantengono la plasticità vocale fino all'età adulta, i suoni scartati potrebbero essere riutilizzati come base per produrre nuovi canti più tardi nel corso della vita.
L'apprendimento basato sull' azione e quello basato sulla memoria hanno un decorso temporale diverso nel ciclo della vita. Il primo presenta la proprietà di riapparire quando, per qualunque ragione, un animale proceda attraverso una fase di pattern d'azione mutevole, seguita da rielaborazione selettiva delle varianti dipendente da uso. È necessario un meccanismo per il rinforzo selettivo di certe varianti motorie rispetto ad altre. Diversamente dall'apprendimento basato sulla memoria, non sono richiesti meccanismi cerebrali altamente specializzati come il sistema del canto degli oscini. Di conseguenza, l'apprendimento basato sull'azione presenta verosimilmente un'ampia distribuzione filogenetica. Per esempio, sebbene i primati non umani appaiano incapaci di apprendimento vocale basato sulla memoria, essi mostrano un limitato grado di plasticità vocale, presumibilmente mediato dall'apprendimento basato sull'azione.
Effetti dell'isolamento e della sordità

Indipendentemente dall'opportunità di apprendere, negli oscini certi aspetti del normale comportamento canoro della specie si sviluppano sempre. Per esempio, quando i passeri sono allevati in isolamento, la struttura delle note e la qualità tonale dei loro canti è aberrante, ma alcuni aspetti della sintassi del canto di base si sviluppano normalmente (fig. 6). Certe caratteristiche del canto della specie vengono prodotte anche se sono assenti nei canti dei 'tutori' che un maschio ha avuto l'esperienza di ascoltare, sotto l'importante condizione che le capacità uditive siano intatte. I pattern di canto di un maschio oscino allevato in isolamento e di un maschio reso sordo precocemente e allevato in isolamento sono piuttosto diversi. Tipicamente i canti di un maschio sordo sono altamente degradati e variabili, mancando di gran parte delle poche caratteristiche normali che un maschio udente è in grado di sviluppare in isolamento. I canti dei maschi resi sordi precocemente non sono del tutto amorfi. Ciò è vero anche quando siano state prese le dovute precauzioni per essere sicuri che essi non abbiano cantato prima di essere stati resi sordi (Nottebohm, 1968). Questa condizione mancava in alcuni degli studi condotti in passato. L'emergenza di alcuni elementi di sintassi canora specie-specifica in maschi resi sordi precocemente nella vita, fa ipotizzare l'esistenza di programmi motori innati nello sviluppo del canto (Marler e Sherman, 1983). Il pattern canoro fortemente anomalo caratteristico degli uccelli sordi si sviluppa sia se un maschio diviene sordo prima di avere un 'tutore', sia se è reso sordo dopo, ma prima che il canto si sia sviluppato (Konishi, 1965). Non sembrano esistere circuiti cerebrali interni in grado di far sì che i canti memorizzati siano direttamente disponibili per guidare lo sviluppo vocale. Per trasformare un canto memorizzato in un canto effettivo, l'uccello deve essere in grado di ascoltare la propria voce. Potremmo supporre che alcuni aspetti della produzione del canto siano guidati da stampi uditivi per il canto, situati nel cervello, con caratteristiche che variano da specie a specie (Konishi, 1965; Marler e Sherman, 1983). I neurobiologi devono tuttavia dimostrare dove sono situati questi stampi e come funzionano (Whaling et al., 1997).
Come gli ornitologi, in natura gli uccelli si basano in gran parte sul canto per l'identificazione specifica. Ciascun uccello ha il suo peculiare modo di cantare che è riconosciuto dagli altri membri della specie. D'altra parte i canti degli uccelli resi sordi precocemente sono chiaramente, sebbene non proprio del tutto, mancanti di caratteristiche specie-specifiche e sono spesso indistinguibili tra specie diverse (v. figura 6). I canti dei maschi allevati in isolamento, ma con capacità uditive intatte, si trovano più o meno a metà strada. Conservano alcuni elementi specie-specifici e ne perdono altri.
Quando canti registrati vengono fatti ascoltare a maschi selvatici territoriali o a femmine indotte in estro con trattamenti ormonali, gli uccelli esperti di ambo i sessi rispondono fortemente al canto nonnale della propria specie, ma non si curano affatto dei canti degli animali resi sordi come segnali sociali (Searcy et al., 1985; Searcye Marler, 1987). ln maniera simile, i canti di altre specie non stimolano alcuna delle intense risposte aggressive dei maschi territoriali o il corteggiamento di femmine trattate con estradiolo, che il canto di un conspecifico è invece in grado di evocare. Tuttavia, i canti di individui che sono cresciuti in isolamento sociale conservano un livello minimo di efficacia. Così l'efficacia del canto di un uccello come segnale sociale è incrementata durante lo sviluppo sia dall'apprendimento sia dall'intervento ontogenetico di stampi uditivi innati che codificano certe caratteristiche specie-specifiche del canto normale.
Preferenze nell'apprendimento
Un tempo i teorici dell' apprendimento credevano che, in condizioni appropriate, qualsiasi stimolo potesse essere appreso e che le differenze nell'intensità dello stimolo, sebbene presenti, fossero abbastanza irrilevanti da essere ignorate nella formulazione delle teorie sui meccanismi di apprendimento. Oggi sappiamo che le predisposizioni innate giocano un ruolo cruciale nella capacità di adattamento dei processi naturali di apprendimento e che l'apprendimento canoro degli uccelli offre molti esempi di questo principio. Nonostante gli uccelli siano capaci di apprendere una moltitudine di suoni differenti, compresi quelli di molte altre specie (fig. 7), l'imitazione di altre specie è relativamente rara in natura (Kroodsma e Baylis, 1982). A eccezione delle specie imitatrici, che costituiscono circa il 15% degli uccelli canori, il fatto di evitare di considerare i canti delle altre specie come modelli di apprendimento ha un valore adattativo.
Quando i maschi di due specie come il passero cantore e il passero delle paludi sono portati in laboratorio ed esposti a registrazioni identiche che comprendono i canti di entrambe, ciascuna mostra una chiara preferenza per il proprio, anche se non lo ha mai ascoltato prima (Marler e Peters, 1988; 1989). Questa preferenza non deriva dall'incapacità di produrre il canto dell'altra specie. Un canto estraneo che un maschio rifiuta quando gli si concede di scegliere può divenire accettabile se costituisce l'unica opzione (v. figura 7), specialmente quando è presentato in una situazione altamente interattiva con un 'tutore' vivo in una gabbia vicina (Baptista e Petrinovich, 1986). Tuttavia, quando gli si concede di scegliere, il maschio mostra tipicamente una preferenza per il canto dell'individuo appartenente alla stessa specie.
Il ruolo della conoscenza innata nello sviluppo del canto
La visione del cervello come una tabula rasa, una lavagna bianca, su cui per troppo tempo si è basato il pensiero dei teorici dell'apprendimento, è chiaramente assurda. Per qualunque tipo di compito, il cervello porta con sé un set di predisposizioni neuronali, ciascuna con la sua propria storia evolutiva e culturale, su come procedere nel modo migliore per risolvere un dato numero di problemi di apprendimento. Alcune di queste predisposizioni sono generiche e panspecifiche, altre sono altamente singolari e anche specie-specifiche, come nel caso dei processi innati che conducono il bambino a sviluppare il linguaggio parlato. Questo è anche il caso degli uccelli canori.
Il giovane uccello inizia il processo di sviluppo del canto armato di predisposizioni innate, sia sensorie che motorie, su come procedere nel modo migliore. Alcune predisposizioni sono generiche, come la tendenza molto comune a emettere suoni tonali, evidente anche quando gli uccelli tentano imitazioni di suoni che non siano in origine strettamente tonali (Nowicki et al., 1992; Peters e Nowicki, 1996). Altre predisposizioni sono specie-specifiche. Quando i passeri imparano a cantare preferiscono il canto di un conspecifico. In tal modo essi non agiscono sulla base di un singolo stimolo segnale etologico, ma di una serie di caratteristiche canore. Giovani uccelli naifs, involati da poco dal nido, sono in grado di distinguere gli stimoli canori che percepiscono più forti, come rivelato dalla loro tendenza a emettere più segnali di richiesta di cure parentali in risposta al play-back del canto (Nelson e Marler, 1993). La reattività innata verso una serie di attributi del canto dei conspecifici è stata dimostrata facendo ascoltare a uccelli giovani alcuni canti modificati sperimentalmente (Whaling et al., 1995). Risulta chiaro dal loro comportamento che gli uccelli giovani possiedono una notevole preconoscenza del canto della propria specie, prima ancora di cominciare il processo di apprendimento del canto. L'evidenza sperimentale indica che essi sanno assai più di quanto si potrebbe ipotizzare sulla base dei canti prodotti da un maschio quando è allevato in isolamento sociale. Essi si comportano come se l'esperienza uditiva che si aggiunge alla loro conoscenza innata fosse richiesta prima che la conoscenza abbia avuto un effetto sullo sviluppo del canto (Marler, 1984).
L'apprendimento canoro può essere considerato come un processo di validazione tramite l'uso di un sottoinsieme di conoscenze innate estratte da un repertorio più esteso codificato nel cervello, che detta le regole per il comportamento canoro di ciascuna specie (Marler e Nelson, 1992). Questo repertorio innato è condiviso da tutti i membri della specie. Le regole universali della specie codificate sono sufficientemente flessibili da fornire opportunità quasi infinite per generare comportamenti individuali differenti a seconda dell'esperienza personale, e tuttavia abbastanza rigide da limitare lo sviluppo di eccessive divergenze tra individui.
Vi sono analogie con le regole della composizione musicale ordinata posseduta da ogni cultura umana e contenenti la potenzialità per generare un infinito numero di differenti melodie (Jackendoff, 1994). Un altro esempio è dato dalle basi genetiche del sistema immunitario, in cui il complesso principale di istocompatibilità procura alle li a sufficienza per un vasto numero di possibili combinazioni. L'imposizione di limiti è importante perché assicura che, per quanto individualistico possa essere il comportamento canoro di un uccello, esso sia ancora sufficientemente conforme a certe regole universali della specie che sembrano minimizzare gli ostacoli alla comunicazione con altri individui.
Oltre alle predisposizioni che sono evidenti nella fase sensoria della memorizzazione del canto, ve ne sono altre che si manifestano durante lo sviluppo motorio. L'emergere di alcune basi specie-specifiche nella sintassi del canto di maschi resi sordi precocemente è già stata evidenziata (Marler e Sherman, 1983). Un uccello indotto con un inganno ad apprendere canti di individui di altre specie potrebbe scartarli solo negli ultimi stadi del canto plastico quando si manifesta la struttura specie-specifica (Marler e Peters, 1989). J. Podos (1996) ha esposto giovani passeri a canti modificati con un numero di sillabe anormalmente alto. Essi imitavano i canti, ma li riorganizzavano in pattern inventati che si conformavano di più all'organizzazione temporale del canto normale della propria specie. Di nuovo si osserva una grande flessibilità in queste predisposizioni motorie, ma nonostante i canti con organizzazione temporale anomala possano essere insegnati, sussiste una tendenza di base a seguire le norme specie-specifiche.
Le differenze individuali e di popolazione che caratterizzano i canti appresi degli uccelli sono assolutamente diffuse, tuttavia esse raramente confondono i birdwatchers esperti nell' identificare le specie responsabili. Considerata la loro fitness genetica a rischio, è molto improbabile che gli uccelli stessi siano meno bravi dei birdwatchers.
Altri aspetti del processo di apprendimento canoro
Alcune delle caratteristiche distintive dell'apprendimento canoro ricorrono in altri casi di plasticità durante lo sviluppo. Non è raro che sistemi comportamentali e neuronali in fase di sviluppo mostrino fasi sensibili in cui la potenzialità per la plasticità è insolitamente elevata. Il completamento di uno stadio è spesso un presupposto necessario per intraprendere lo stadio successivo (Rauschecker e Marler, 1987; Pinker, 1994).
A differenza di altre forme di apprendimento ampiamente studiate, nell' apprendimento del canto non è richiesto un rinforzo esterno, anche se lo stimolo sociale potrebbe accrescere il processo di acquisizione. Affinché il canto venga appreso è sufficiente che l'uccello sia esposto a un set appropriato di stimoli nel momento opportuno dello sviluppo. Nei momenti cruciali del periodo sensibile l'apprendimento può avvenire con grande rapidità. Talora sono sufficienti poche esposizioni al canto della propria specie. Nei passeri una certa acquisizione avviene a partire da trenta ripetizioni del canto di un conspecifico. Per i merli sono necessarie da quindici a venti presentazioni in un singolo giorno.
Un virtuoso del canto è l'usignolo, in grado di apprendere con precisione una sequenza di tipi di canto dopo venti presentazioni (ma a certi maschi ne bastano anche cinque). Il virtuosismo degli usignoli è tale che essi possono apprendere sequenze contenenti fino a sessanta canti diversi e riprodurle nell'ordine originale. Tuttavia, il sequenziamento è conservato solo in sottoinsiemi che vanno da tre a sette canti. Negli studi condotti sugli usignoli si osserva che dopo aver prodotto tale sequenza i soggetti sperimentali cambiano sottoinsieme, iniziando in un punto differente. Questi uccelli si comportano come se dividessero una sequenza di molti canti in sottoinsiemi più semplici, al massimo di sette canti, utilizzando per memorizzare le sequenze lunghe una strategia che ricorda quella utilizzata dagli uomini per memorizzare le sequenze di parole (Hultsch e Todt, 1989; 1992). Questa è una delle molte peculiarità degli uccelli che ricordano certe caratteristiche del comportamento vocale umano.
Sviluppo del canto come processo creativo
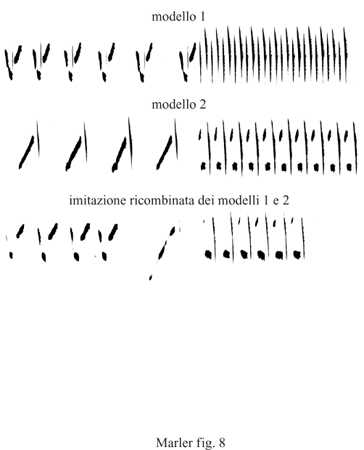
Forse la capacità più misteriosa e affascinante degli uccelli canori è quella di inventare suoni nuovi. Essi raggiungono questo scopo in molti modi differenti. Alcuni uccelli cominciano con l'imitare le componenti dei canti che hanno ascoltato e poi, a mano a mano che procedono durante la fase del canto plastico, le ricombinano per creare nuove sequenze (Marler e Peters, 1987). In questo modo le parti possono essere scambiate all'interno di un canto o tra differenti canti e anche tra canti che sono stati acquisiti a molti mesi di distanza (fig. 8). Piuttosto che rimanere fedeli alla struttura dei suoni imitati, alcuni uccelli manifestano l'irresistibile tentazione di improvvisare, specialmente durante la fase del canto plastico, cosicché i canti inizialmente imitati diventano tanto diversi da non essere riconoscibili (Marler et al., 1972).
Non di rado gli uccelli inventano nuovi pattern sonori che essi stessi non hanno mai ascoltato. Ciò avviene frequentemente in uccelli allevati in isolamento, col risultato che, sebbene i canti di tali uccelli siano per certi aspetti insolitamente semplici, essi sono anche estremamente variati. L'atto in sé di produrre suoni nuovi e con pattern distintivi sembrerebbe costituire un rinforzo, a testimonianza della forte e profondamente radicata predisposizione all'inventiva vocale di molti uccelli canori.
La tendenza a inventare è più forte in certe specie che in altre. Certi uccelli restano fedeli in maniera piuttosto rigida al loro 'tutore' aggiungendo componenti individuali appena sufficienti a personalizzare il loro canto e niente più. L'aderenza ai loro modelli li rende particolarmente adatti per ricerche di laboratorio sull'apprendimento canoro. Forse per questo motivo gli uccelli canori più fantasiosi tendono a essere poco rappresentati nella lista dei soggetti preferiti per studi scientifici. Gli enormi repertori individuali che i maschi di alcuni uccelli possiedono, talora con centinaia di tipologie canore assemblate tramite la ripetizione dei processi di imitazione, segmentazione, ricombinazione, improvvisazione e invenzione, li rendono quasi impossibili da studiare per scopi scientifici. Ma per chiunque sia interessato alla teoria estetica, gli uccelli con vasti repertori che offrono all'ascoltatore umano un così grande piacere, come i mimi e i loro affini, l'uccello lira australiano e certi tordi, sono insuperabili come soggetti per l'ascolto. Gli argomenti utilitaristici sono solo un aspetto del fascino che il canto degli uccelli suscita in noi. Se volessimo cercare modelli animali dell' impulso creativo il canto degli uccelli sarebbe uno dei pochi che si avvicina, seppur vagamente, alle tipologie umane.
Conclusioni
A mano a mano che il numero delle specie studiate aumenta ci rendiamo conto che ciascuna di esse ha raggiunto le proprie soluzioni specifiche ai problemi dello sviluppo del canto. Gli uccelli canori partecipano a un sistema di comunicazione sociale che si basa su segnali appresi da altri. Alcune specie apprendono prontamente soltanto dalla stimolazione canora e spesso sono necessari pochi stimoli, come se il cervello fosse predisposto a essere reattivo soprattutto verso il canto della propria specie. Altre specie impiegano più tempo per apprendere e necessitano di una stimolazione sociale.
Nonostante posseggano estese conoscenze innate sui canti della propria specie e una forte tendenza a preferirli quando si offre loro una scelta, in speciali circostanze gli uccelli possono essere indotti ad apprendere canti di altri. Certe specie intraprendono naturalmente il processo di mimetismo tra specie e creano un canto specie-specifico imponendo pattern temporali distintivi alle imitazioni quando esse sono esibite. Alcuni uccelli sono fedeli al canto che imitano e altri usano le imitazioni come base per l'improvvisazione, conservando certe caratteristiche del 'tutore' e cambiandone altre.
La diversità di questi pattern di sviluppo comportamentale è resa possibile in quanto il cervello di ciascuna specie di uccello determina un set distinto di proprietà e predisposizioni per imparare a cantare. La sfida per i neurobiologi è comprendere non solo i meccanismi cerebrali che sono alla base delle capacità generali di apprendimento di tutti gli uccelli canori, ma anche quelli più elusivi e particolari con cui una data specie aggiunge il suo peculiare tocco al processo di sviluppo di un canto per renderlo adatto al proprio particolare contesto sociale, ecologico e filogenetico.
Bibliografia citata
ALVAREZ-BUYLLA, A., KIRN, J.R., NOTTEBOHM, F. (1990) Birth of projection neurons in adult avian brain may be related to perceptual or motor leaming. Science, 249, 1444-1446.
ARNOLD, A.P. (1990) The passerine bird song system as a model in neuroendocrine research. J. Exp. Zool. Suppl., 4, 22-30.
ARNOLD, A.P., BOTTJER, S.W., BRENOWITZ, E.A., NORDEEN, E.J., NORDEEN, K.W. (1986) Sexual dimorphisms in the neural vocal control system in song birds: ontogeny and phylogeny. Brain Behav. Evol., 28, 22-31.
BAKER, M.C. (1983) The behavioral response of female Nuttall's white-crowned sparrows to male song of natal and alien dialects. Behav. Ecol. Sociobiol., 12, 309-314.
BAKER, M.C., BJERKE, T.K., LAMPE, H., ESPMARK, Y.O. (1987) Sexual response of female yellowhammers to differences in regional song dialects and repertoire sizes. Anim. Behav., 35, 395-401.
BAKER, M.C., CUNNINGHAM, M.A. (1985) The biology of birdsong dialects. Behav. Brain Sci., 8, 85-133.
BAKER, M.C., SPITLER-NABORS, K.J., BRADLEY, D.C. (1981) Early experience determines song dialect responsiveness of female sparrows. Science, 214, 819-821.
BALABAN, E. (1988) Bird song syntax: learned intraspecific variation is meaningful. Proc. Natl. Acad. Sci. USA, 85, 3657-3660.
BAPTISTA, L.P., PETRINOVICH, L. (1986) Song development in the white-crowned sparrow: social factors and sex differences. Anim. Behav., 34, 1359-1371.
BEECHER, M.D. (1996) Birdsong leaming in the laboratory and field. In Ecology and evolution of acoustic communication in birds, a c. di Kroodsma D.E., Miller E.H., Ithaca, New York, Comstock PubI. Assoc. (Comell University Press).
BOTTJER, S.W., MIESNER, E., ARNOLD, A.P. (1984) Forebrain lesions disrupt development but not maintenance of song in passerine birds. Science, 224, 901-903.
BRENOWITZ, E.A., ARNOLD, A.P. (1989) Accumulation of estrogen in a vocal control brain region of a duetting song bird. Brain Res., 480, 119-125.
BRENOWITZ, E.A., ARNOLD, A.P., LEVIN, R. (1985) Neural correlates of female song in tropical duetting birds. Brain Res., 343, 104-112.
BRENOWITZ, E., KROODSMA, D.E. (1996) The neuroethology of birdsong. In Ecology and evolution of acoustic communication in birds, a c. di KroodsmaD.E., Miller E.H., Ithaca, ComstockPubl.
CANADY, R.A., KROODSMA, D.E., NOTTEBOHM, F. (1984) Population differences in complexity of a leamed skill are correlated with the brain space involved. Proc. Natl. Acad. Sci. USA, 81, 6232-6234.
CATCHPOLE, C.K., SLATER, P.J.B. (1995) Bird song: biological themes and variations. Cambridge- New York, Cambridge University Press.
DEVOOGD, T.J. (1991) Endocrine modulation of the development and adult function of the avian song system. Psychoneuroendocrinology, 16, 41-66.
DEVOOGD, T.J., KREBS, J.R., HEALY, S.D., PURVIS, A. (1993) Relations between song repertoire size and the volume of brain nuclei related to song: comparative evolutionary analyses amongst oscine birds. Proc. R. Soc. Lond. B Biol. Sci., 254, 75-82.
DOOLING, R.J. (1982) Auditory perception in birds. In Acoustic communication in birds, a c. di Kroodsma D.E., Miller E.H., voI. I, New York, Academic Press, pp. 95-130.
DOUPE, A.J. (1993) A neural circuit specialized for vocalleaming. Curr. Opin. Neurobiol., 3, 104-111.
GOLDMAN, S.A., NOTTEBOHM, F. (1983) Neuronal production, migration, and differentiation in the vocal control nucleus of the adult female canary brain. Proc. Natl. Acad. Sci. USA, 80, 2390-2394.
GREENEWALT, C.H. (1968) Bird song: acoustics and physiology. Washington, Smithsonian Institution Press.
HULTSCH, H., TODT, D. (1989) Memorization and reproduction of songs in nightingales: evidence for package formation. J. Comp. Physiol. A, 165, 197-203.
HULTSCH, H., TODT, D. (1992) The serial order effect in the song acquisition of birds: relevance of exposure frequency to song models. Anim. Behav., 44, 590-592.
JACKENDOFF, R. (1994) Patterns in the mind: language and human nature. New York, Basic Books.
KONISHI, M. (1965) The role of auditory feedback in the control of vocalization in the white-crowned sparrow. Z. Tierpsychol., 22, 770-783.
KONISHI, M. (1985) Birdsong: from behavior to neuron. Annu. Rev. Neurosci., 8,125-170.
KROODSMA, D.E. (1984) Songs of the alder flycatcher (Empidonax alnorum) and willow flycatcher (Empidonax traillii) are innate. Auk, 101, 13-24.
KROODSMA, D.E., BAYLIS, J.R. (1982) A world survey of evidence for vocallearuing in birds. In Acoustic communication in birds, a c. di Kroodsma D.E., Miller E.H., voI. I, New York, Academic Press, pp. 311-337.
KROODSMA, D.E., KONISHI, M. (1991) A suboscine bird (Eastem phoebe, Sayornis phoebe) develops normal song without auditory feedback. Anim. Behav., 42, 477-487.
KROODSMA, D.E., PICKERT, R. (1980) Enviroumentally dependent sensitive periods for avian vocal learuing. Nature, 288, 477-479.
LOCKE, J.L. (1993) The path to spoken language. Cambridge, Mass., Harvard University Press.
MARGOLIASH, D. (1986) Preference for autogenous song by auditory neurons in a song system nucleus of the white-crowned sparrow. J. Neurosci., 6, 1643-1661.
MARLER, P. (1984) Song learning: innate species differences in the learning process. In The biology of learning, a c. di Marler P., Terrace H.S., Berlin, Springer Verlag.
MARLER, P. (1991) Song learning behavior: the interface with neuroethology. Trends Neurosci., 14, 199-206.
MARLER, P., MUNDINGER, P., W ASER, M.S., LUTJEN, A. (1972) Effects of acoustical stimulation and deprivation on song development in red-winged blackbirds (Agelaius phoeniceus). Anim. Behav., 20, 586-606.
MARLER, P., NELSON, D. (1992) Neuroselection and song leaming in birds: species nniversals in a culturally transmitted behavior. In Seminars in the neurosciences, a c. di Marler P., voI. 4, London, Sannders Scientific Publications, pp. 415-423.
MARLER, P., NELSON, D.A. (1993) Action-based leaming: a new form of developmental plasticity in bird song. Neth. J. of Zool., 43, 91-103.
MARLER, P., PETERS, S. (1982a) Developmental overproduction and selective attrition: new processes in the epigenesis of bird song. Dev. Psychobiol., 15, 369-378.
MARLER, P., PETERS, S. (1982b) Subsong and plastic song: their role in the vocal learning process. In Acoustic communication in birds, a c. di Kroodsma D.E., Miller E.H., voI. 2, New York, Academic Press, pp. 25-50.
MARLER, P., PETERS, S. (1987) A sensitive period for song acquisition in the song sparrow, Melospiza melodia, a case of agelimited learning. Ethology, 76, 89-100.
MARLER, P., PETERS, S. (1988) The role of song phonology and syntax in vocal learning preferences in the song sparrow, Melospiza melodia. Ethology, 77, 125-149.
MARLER, P., PETERS, S. (1989) Species differences in auditory responsiveness in early vocal leaming. In The comparative psychology of audition: perceiving complex sounds, a c. di Dooling R.J., Hulse S.H., Hillsdale, New York, Lawrence Erlbaum Associates, pp. 243-273.
MARLER, P., SHERMAN, V. (1983) Song structure without auditory feedback: emendations of the auditory template hypothesis. J. Neurosci., 3, 517-531.
MELLO, C.V., CLAYTON, D.F. (1994) Song-induced ZENK gene expression in auditory pathways of songbird brain and its relation to the song control system. J. Neurosci., 14, 6652-6666.
MELLO, C.V., VICARIO, D.S., CLAYTON, D.P. (1992) Song presentation induces gene expression in the songbird's forebrain. Proc. Natl. Acad. Sci. USA, 89, 6818-6822.
NELSON, D.A. (1992) Song overproduction and selective attrition lead to song sharing in the field sparrow (Spizella pusilla). Behav. Ecol, Sociobiol., 30, 415-424.
NELSON, D.A., MARLER, P. (1993) Innate recognition of song in white-crowned sparrows: a role in selective vocal learning? Anim. Behav., 46, 806-808.
NELSON, D.A., MARLER, P. (1994) Selection-based learuing in bird song development. Proc. Natl. Acad. Sci. USA, 91, 10498-10501.
NELSON, D.A., MARLER, P., PALLERONI, A. (1995) A comparative approach to vocallearning: intra-specific variation in the learning processo Anim. Behav., 50, 83-97.
NELSON, D.A., WHALING, C., MARLER, P. (1996) The capacity for song memorization varies in populations of the same species. Anim. Behav., 52, 379-387.
NOTTEBOHM, F. (1968) Auditory experience and song development in the chaffinch, Fringilla coelebs. Ibis, 110, 549-568.
NOTTEBOHM, P. (1972) Neural lateralization of vocal control in a passerine bird II. Subsong, calls and a theory of learning. J. Exp. Zool., 179, 35-49.
NOTTEBOHM, F. (1993) The search for neural mechanisms that define the sensitive period for song learning in birds. Neth. J. Zool., 43, 193-234.
NOTTEBOHM, P., ARNOLD, A.P. (1976) Sexual dimorphism in vocal control areas ofthe songbird brain. Science, 194, 211-213.
NOTTEBOHM, F., KASPARIAN, S., PANDAZIS, S. (1981) Brain space for a learned task. Brain Res., 213(1), 99-109. NOTTEBOHM, F., NOTTEBOHM, M.E. (1978) Relationship between song repertoire and age in the canary Serinus canaria. Z. Tierpsychol., 46, 298-305.
NOTTEBOHM, F., STOKES, T.M., LEONARD, C.M. (1976) CentraI control of song in the canary Serinus canaria. J. Comp. Neurol., 165, 457-486.
NOWICKI, S., MARLER, P., MAYNARD, A., PETERS, S. (1992) Is the tonaI quality of birdsong leamed? Evidence from song sparrows. Ethology, 90, 225-235.
PATON, G.A., NOTTEBOHM, F. (1984) Neurons generated in the adult brain are recruited into functional circuits. Science, 225, 1046-1048.
PETERS, S., NOWICKI, S. (1996) Development of tonaI quality in birdsong: further evidence from song sparrows. Ethology, 102, 323-335.
PINKER, S. (1994) The Language Instinct. New York, W. Morrow and Comp.
PODOS, J. (1996) Motor constraints on vocal development in a songbird. Anim. Behav., 51, 1061-1070.
RAUSCHECKER, J.P., MARLER, P., a C. di (1987) Imprinting and Cortical Plasticity. New York, John Wiley & Sons.
SEARCY, W.A., MARLER, P. (1987) Response of sparrows to songs of deaf and isolation-reared males: further evidence for innate auditory templates. Dev. Psychobiol., 20, 509-519.
SEARCY, W.A., MARLER, P., PETERS, S.S. (1985) Song of isolationreared sparrows function in communication, but are significantly less effective than leamed songs. Behav. Ecol. Sociobiol., 17, 223-229.
STRIEDTER, G.P. (1994) The vocal control pathways in budgerigars differ from those in songbirds. J. Comp. Neurol., 343, 35-56.
SUTHERS, R.A. (1990) Contributions to birdsong from the left and right sides of the intact syrinx. Nature, 347, 473-477.
THORPE, W.H. (1961) Bird Song. Cambridge, Mass., Cambridge University Press.
VICARIO, D.S. (1991) Neural mechanisms of song production in songbirds. Curr. Opin. Neurobiol., l, 595-600.
VICARIO, D.S., SIMPSON, H.B. (1995) Electrical stimulation in forebrain nuclei elicits leamed vocal pattems in songbirds. J. Neurophysiol., 73(6), 2602-2607.
VOLMAN, S. (1993) Development of neural selectivity for birdsong during vocal learning. J. Neurosci., 13, 4737-4747.
WEST, M.J., KING, A.P. (1988) Female visual displays affect the development of male song in the cowbird. Nature, 334, 244-246.
WHALING, C.S., SOLIS, M.M, CARRILLO, G., SOHA, J.A., MARLER, P., DOUPE, A.J. (1995) Song learning in sparrows: from behavior to brain. Soc. for Neurosci., 21(2), 958.
WHALING, C.S., SOLIS, M.M, DOUPE, A.J., SOHA, J.A., MARLER, P. (1997) Acoustic and neural bases for innate recognition of song. Proc. Natl. Acad. Sci. USA, 94, 12694-12698.
WILD, J.M. (1994) The auditory-vocal-respiratory axis in birds. Brain Behav. Evol., 44, 192-209.