
Aggressivita
Aggressività
Il comportamento aggressivo è assai frequente nel regno animale e di solito si conclude con l’ottenimento, da parte del vincitore, dell’accesso a una risorsa disponibile in quantità limitata. Esso prevede in molti casi uno scambio iniziale di segnali di esibizione da una certa distanza e prosegue, in un crescendo di intensità, prima con approcci fisici diretti, poi con il combattimento vero e proprio. I conflitti possono risolversi anche senza concludere l’intera sequenza, ma le ferite gravi, a volte mortali, non sono affatto rare. La propensione all’aggressività varia notevolmente tra gli individui di una determinata specie e molti studi cercano di spiegare questa variabilità. Generalmente l’espressione dell’aggressività negli animali dipende da fattori ormonali, neuronali, genetici e di ecologia comportamentale. I termini ‘aggressione’ e ‘aggressività’ vengono spesso utilizzati nel linguaggio comune e sono riferiti di solito a un assalto o a un attacco non provocato. Da un lato, questi vocaboli generici sono vantaggiosi poiché facilitano agli etologi il compito di spiegare le loro ricerche ai non specialisti. Tuttavia, un termine generico causa alcuni problemi se usato in un contesto scientifico, perché, sebbene sia chiaro a tutti cos’è il comportamento aggressivo, questa espressione viene spesso usata per indicare cose piuttosto differenti tra loro. In altri termini, esiste un problema di definizione di questo argomento che non è di facile risoluzione e che ha suscitato infuocate controversie. La definizione di comportamento aggressivo che viene utilizzata più comunemente in etologia è la seguente: «un comportamento con il quale un animale danneggia, realmente o potenzialmente, un altro membro della sua stessa specie» (Huntingford e Turner 1987). L’avverbio ‘potenzialmente’ serve a includere tra i comportamenti aggressivi un attacco fallito. Precisare che la vittima deve essere un conspecifico serve a escludere che il termine possa essere riferito anche a un attacco sferrato da un predatore a una preda, o a un contrattacco da parte di quest’ultima; azioni, queste, che possono entrambe danneggiare l’avversario.
Questa definizione così semplice non risolve la questione se debbano essere considerate aggressive anche azioni non intenzionali, attacchi sferrati in risposta al comportamento dell’altro (da contrapporre agli attacchi non provocati), ferite causate con le migliori intenzioni nell’interesse dell’animale che le subisce o, infine, comportamenti che abbiano come conseguenza danni psicologici anziché fisici. Si tratta certamente di problemi importanti, ma essi non possono essere adeguatamente affrontati all’interno di una definizione generale senza estendere e complicare la definizione stessa al punto tale da renderla inutile. La strategia migliore, che adotteremo anche qui, consiste nel mantenere una certa flessibilità, utilizzando il termine aggressività secondo la definizione sopra riportata per indicare una categoria generale di comportamenti, chiarendone se necessario gli aspetti specifici.
Sommario: 1. Forme di aggressività nel regno animale. 2. La variabilità intraspecifica. 3. Le cause del comportamento aggressivo. 4. Costi e benefici dell’aggressività: la teoria dei giochi. □ Bibliografia.
1. Forme di aggressività nel regno animale
Il comportamento con il quale, realmente o potenzialmente, viene danneggiato un conspecifico (cioè il comportamento aggressivo) è largamente diffuso nel regno animale a ogni livello di complessità evolutiva. Così, per esempio, i cervi volanti maschi (Lucanus cervus) si battono per ottenere l’accesso alle femmine, utilizzando le proprie mandibole ipersviluppate per afferrare e rovesciare l’avversario; oppure gli Iguanidi maschi (Uta stansburiana) si disputano i territori riproduttivi; i galli domestici (Gallus domesticus) e altri uccelli si battono per stabilire il proprio status sociale; infine le zebre maschio si lanciano l’una contro l’altra colpendosi con gli zoccoli per aggiudicarsi le femmine. La maggior parte dei combattimenti tra animali coinvolge due individui, sebbene a volte il conflitto possa interessare gruppi più numerosi, come nel caso delle formiche che occupano colonie adiacenti e che si contendono i territori alimentari.
Le posture e gli atti che caratterizzano il comportamento aggressivo sono i più diversi, ma è possibile delineare alcune caratteristiche comuni. In primo luogo, di solito gli animali dispongono di un repertorio di segnali che precedono lo scontro fisico diretto. Il comportamento aggressivo comprende tanto segnali di esibizione emessi a una certa distanza dall’avversario, quanto azioni che prevedono il contatto fisico, ma sono innocue, oppure atti decisamente suscettibili di danneggiare l’avversario, quali il mordere e lo sferrare calci. Di solito (ma non sempre) si assiste a un graduale crescendo d’intensità, con i segnali di esibizione a distanza emessi tendenzialmente all’inizio, seguiti dal contatto fisico diretto e, infine, dal combattimento vero e proprio. Per risolvere un conflitto possono essere sufficienti i segnali di esibizione e le interazioni innocue, anche se le ferite gravi e le uccisioni sono eventi tutt’altro che rari.
Un aspetto fondamentale delle interazioni aggressive tra animali è che esse sono in genere provocate dal bisogno di una determinata risorsa, vitale per entrambi i contendenti, il cui accesso però è permesso esclusivamente o preferenzialmente al vincitore dello scontro. L’accesso alla risorsa contesa può essere immediato, come quando gli animali si disputano il cibo o un partner sessuale, ma può anche essere dilazionato nel tempo: per esempio, spesso gli animali lottano per ottenere il controllo di un’area in cui è molto probabile trovare risorse diverse (cibo, partner sessuali, riparo, ecc.). Il combattimento porta allora allo stabilirsi di una territorialità, ovvero del controllo, più o meno assoluto, da parte di un individuo su una porzione di ambiente vitale. Altre volte il premio per un incontro aggressivo è la dominanza o la supremazia sociale sui rivali, cosa che in futuro significherà avere un accesso preferenziale alle risorse, come avviene per esempio nel caso dei galli domestici che si battono per l’accoppiamento.
2. La variabilità intraspecifica
Gli animali appartenenti a una data specie combattono in un modo unico e caratteristico, cioè specie-specifico, ma i singoli individui possono differire notevolmente nella propensione alla lotta. Un esempio di ciò è il comportamento del maschio dello spinarello (Gasterosteus aculeatus) il quale, durante il periodo riproduttivo, attacca i potenziali rivali che entrano nel suo territorio se l’area ventrale del loro corpo è colorata di rosso. La frequenza di attacchi per minuto mostra risultati variabili: alcuni soggetti di fatto ignorano l’intruso, altri invece lo mordono con una frequenza anche di due volte al secondo! Tale variabilità può essere manipolata mediante incroci selettivi. La variabilità intraspecifica nel comportamento aggressivo può essere ancora più estrema. Per esempio, alcuni maschi di una specie strettamente imparentata con lo spinarello (lo spinarello a dieci spine, Pygosteus pungitius) non combattono affatto: essi si insinuano di soppiatto nel territorio di un altro maschio, entrano nel nido in cui questo ha indotto la sua femmina a deporre le uova e le fertilizzano prima del rivale. Questo comportamento rappresenta un caso, all’interno di una stessa popolazione di conspecifici, di strategia alternativa per la competizione in vista delle risorse ed è diffuso in molte altre specie: per esempio, tra alcune rane e rospi, i cui maschi privi di territorio intercettano e si accoppiano con le femmine che si dirigono verso i maschi territoriali che le hanno attirate con i loro richiami.
La variabilità nella competizione aggressiva per le risorse da parte dei membri di una data specie, oltre a rappresentare un fenomeno interessante di per sé, è anche un utilissimo strumento di ricerca. Studiando la variabilità riscontrata nella frequenza di attacco dello spinarello, si è visto che essa può essere in parte ascritta alle diverse condizioni fisiche di ciascun individuo: i maschi con scarse riserve energetiche si dimostrano combattenti assai poco entusiasti. Un’altra parte della variabilità può invece essere spiegata considerando il regime di predazione prevalente nei diversi habitat dei Pesci: gli spinarelli che vivono in aree prive di predatori sono più aggressivi di quelli che vivono in siti in cui questi abbondano, poiché in quest’ultimo caso il combattimento è più rischioso. Le differenze comportamentali che si riscontrano tra Pesci provenienti da diversi siti sono probabilmente basate anche su differenze genetiche, come suggerito da alcuni esperimenti di selezione nei quali sono state progressivamente create e differenziate due diverse linee, all’interno di ciascuna delle quali veniva fatto riprodurre esclusivamente il maschio meno aggressivo o, viceversa, quello più aggressivo. I risultati di tali esperimenti dimostrano notevoli differenze nella propensione all’aggressività (rispettivamente ridotta e accresciuta) delle diverse linee ottenute. Ciò significa che, almeno in parte, la variabilità naturale nel comportamento aggressivo dello spinarello maschio è dovuta a fattori ereditari.
Le cause del comportamento aggressivo
Il controllo ormonale
In molte ricerche si è visto che, nelle specie in cui i maschi competono direttamente per l’accesso alle femmine durante il periodo riproduttivo, l’ormone testosterone (un androgeno, prodotto tipicamente dai maschi dei Vertebrati) stimola il comportamento aggressivo, sebbene tale effetto sia meno pronunciato laddove l’organizzazione sociale della specie considerata è più complessa. Oltre a esercitare un’influenza sull’animale adulto, il testosterone può agire nel corso dello sviluppo, come dimostrato dal fatto che, almeno nella classe dei Mammiferi, l’esposizione a questo ormone in una fase ontogenetica precoce è in grado di sensibilizzare l’animale alla stimolazione esercitata dallo stesso una volta raggiunta l’età adulta.
Il gruppo di specie delle lucertole si è dimostrato molto utile per studiare le basi fisiologiche del comportamento aggressivo, poiché molte specie sono sufficientemente grandi e robuste da poter essere manipolate sperimentalmente sia in laboratorio sia sul campo. Inoltre, il loro comportamento è allo stesso tempo abbastanza semplice da poter essere registrato con facilità e tuttavia sufficientemente complesso da risultare interessante. Alcuni punti sono stati stabiliti con certezza per molte specie: (a) l’aumentata propensione all’aggressività osservabile nelle lucertole maschio all’approssimarsi della stagione riproduttiva, così come la colorazione brillante che il loro corpo assume in questo periodo, sono fenomeni sicuramente favoriti dall’incremento dei livelli di androgeni; (b) la castrazione di maschi in piena fase riproduttiva ne riduce drasticamente l’aggressività; (c) la somministrazione di androgeni è in grado di contrastare questo effetto; (d) gli androgeni sono concentrati in aree cerebrali notoriamente coinvolte nel controllo del comportamento aggressivo.
Come i maschi di spinarello, anche quelli di diverse specie di lucertole si disputano le femmine utilizzando una di due o più possibili strategie. Così si possono trovare, all’interno di una stessa popolazione, tanto maschi che mostrano un comportamento aggressivo e che difendono i propri territori, quanto maschi nomadi che non mostrano un comportamento aggressivo e che riescono ad accoppiarsi intrufolandosi di soppiatto. Questi differenti comportamenti sono spesso associati a diverse colorazioni corporee. Per esempio in Urosaurus ornatus i maschi arancione-blu (così chiamati perché presentano una sacca golare o ‘ventola della gola’ di colore arancione, al centro della quale spicca una brillante macchia blu) mostrano uno spiccato comportamento aggressivo e difendono i propri territori, mentre i maschi che hanno la gola interamente arancione non possiedono un territorio fisso e sono relativamente poco aggressivi. Questi due tipi di maschi non differiscono nei livelli di androgeni circolanti e un’alterazione di questi livelli indotta sperimentalmente nei soggetti adulti durante il periodo riproduttivo non causa alcuna trasformazione nella colorazione della gola. Tuttavia, si ottengono risultati molto diversi se si manipolano i livelli ormonali nel corso dello sviluppo.
Se per esempio alcuni soggetti appena usciti dall’uovo vengono castrati e altri, invece, sono forniti di un dispositivo che rilascia testosterone, il risultato è che la maggior parte dei maschi castrati precocemente diventa del tipo non aggressivo di colore arancione, mentre quelli esposti a più alti livelli di testosterone si sviluppano per lo più in individui aggressivi, con colorazione arancione-blu. Inoltre si è visto che, per effetto del trattamento con testosterone aggiuntivo, si abbassa significativamente la percentuale di lucertole trattate con forti dosi di testosterone che riesce a sopravvivere oltre i 90 giorni di età: un risultato, questo, da mettere probabilmente in relazione con il fatto che uno stile di vita aggressivo implica generalmente costi considerevoli. Un’interazione di questo tipo, tra gli effetti irreversibili di differenti ormoni durante lo sviluppo e gli effetti reversibili che gli ormoni hanno negli adulti, si osserva in molte delle specie animali utilizzate negli studi di endocrinologia del comportamento aggressivo.
Il controllo neuronale
Il controllo neuronale del comportamento aggressivo nei Vertebrati è molto complesso, perché basato su un sistema di strutture cerebrali che lavorano interagendo tra loro e che presentano in molti casi un’alta densità di recettori per gli androgeni e gli estrogeni. Ci limiteremo a descrivere un singolo esempio che illustrerà come l’esistenza di strategie alternative possa essere utilizzata per identificare i substrati nervosi implicati nella regolazione dei diversi aspetti del comportamento aggressivo e per caratterizzare la loro dipendenza dagli androgeni. Porichthys notatus è una specie di Pesci in cui, durante il periodo riproduttivo, la maggior parte dei maschi (del tipo 1) costruisce il nido tra le fenditure della zona intertidale e lo difende dagli intrusi. Una caratteristica importante del comportamento di questi maschi è rappresentata dalle vocalizzazioni, emesse da gruppi di muscoli specializzati che fanno vibrare la vescica natatoria: gli ‘hum’ (ronzii) vengono emessi per attirare le femmine, i ‘grunt’ (brontolii) durante i combattimenti territoriali contro gli intrusi.
Circa il 10% dei maschi in fase riproduttiva (del tipo 2) è invece composto da individui più piccoli, che possiedono gonadi relativamente grandi e che non difendono alcun nido né emettono vocalizzazioni essi si accoppiano insinuandosi di soppiatto nei nidi dei maschi del tipo 1. Studi anatomici e registrazioni elettrofisiologiche condotte sui muscoli che producono le vocalizzazioni, hanno evidenziato importanti differenze tra gli apparati fonatori dei due tipi di maschi, portando alla scoperta del meccanismo che spiega le loro eclatanti differenze comportamentali. Così, nei maschi del tipo 1 i muscoli che fanno vibrare la vescica natatoria risultano notevolmente ingrossati e dotati di fibre muscolari più numerose e più grandi (caratterizzate da una maggiore densità di mitocondri) rispetto ai maschi del tipo 2. I neuroni motori che innervano questi muscoli, e che partono da specifici nuclei del cervello posteriore (i quali sono anch’essi molto più estesi), sono fino a tre volte più grandi nei maschi del tipo 1, che possiedono giunzioni neuromuscolari più elaborate. La frequenza di attività dei circuiti centrali di produzione del suono è maggiore del 20% nei maschi del tipo 1 rispetto ai maschi del tipo 2.
L’apparato di produzione del suono dei maschi di tipo 1 inizia a svilupparsi quando gli individui sono ancora immaturi e le fibre muscolari aumentano rapidamente in numero e grandezza, a quanto pare sotto l’influsso del livello di androgeni circolanti e in particolare dell’11-chetotestosterone, un ormone che risulta molto potente nel promuovere lo sviluppo delle caratteristiche sessuali secondarie nei Pesci. I maschi del tipo 2 hanno in circolo alti livelli di testosterone, ma non producono affatto 11-chetotestosterone e l’architettura delle loro strutture muscolari non risente dei livelli di androgeni. Dunque, l’evidente divergenza comportamentale tra i maschi del tipo l e quelli del tipo 2 riflette importanti differenze nella grandezza, nella struttura e nella fisiologia del sistema neuromuscolare responsabile della produzione del suono, differenze che si stabiliscono a partire dalle fasi precoci dello sviluppo sotto l’influsso degli androgeni circolanti.
Fattori genetici
Una gran quantità di studi ha mostrato come il comportamento aggressivo si sviluppi attraverso una complessa interazione tra fattori ereditari ed effetti legati all’esperienza. Una rassegna dell’articolata letteratura esistente sull’argomento è contenuta nei lavori di Felicity A. Huntingford e Angela K. Turner (1987) e di John Archer (1987). I topi selvatici e i loro discendenti addomesticati (Mus spp.) sono stati largamente impiegati per questo tipo di ricerche, in parte perché mostrano un comportamento aggressivo ben sviluppato, in parte perché è più facile impiegarli nei complessi programmi di allevamento necessari per separare l’effetto dei fattori genetici sullo sviluppo dell’aggressività da quello dei fattori ambientali. Le popolazioni di topi alternano periodi in cui sono organizzate in gruppi familiari stabili e riproduttivamente isolati tra loro a periodi di migrazione e colonizzazione di nuove aree. Durante le fasi di stabilità, i maschi adulti si contendono i territori in cui risiedono diverse femmine mature. Si descriverà qui uno dei programmi di ricerca più importanti e rappresentativi tra quelli condotti sulla genetica del comportamento aggressivo nel topo. Se, esaminando i maschi adulti di una popolazione di topi selvatici, si misura la rapidità con la quale essi attaccano un individuo che si introduce nel loro territorio, è possibile suddividerli in due gruppi, il primo composto da soggetti che attaccano rapidamente e il secondo da topi più lenti ad aggredire. Si ritiene che gli animali che attaccano rapidamente abbiano maggiore successo degli altri quando si tratta di difendere un territorio durante un periodo di stabilità, mentre i soggetti più lenti si dimostrano più abili nel conquistare un territorio in seguito alla migrazione e alla colonizzazione di nuove aree.
Per studiare l’influenza genetica su questo comportamento, è possibile estendere per numerose generazioni successive di animali un programma di allevamento selettivo di due diverse linee, caratterizzate una dalla rapidità e una dalla lentezza nel comportamento di attacco. La selezione artificiale può avere un effetto chiaro e marcato, che rappresenta una prova importante del fatto che una parte significativa della variabilità riscontrata nel comportamento aggressivo della popolazione selvatica originaria dipende da differenze genetiche a carico di specifiche porzioni del cromosoma Y. È interessante ricordare che queste differenze sono associate a molte altre diversità comportamentali: nel complesso, i topi più lenti nell’attaccare mostrano generalmente, rispetto ai più veloci, una maggiore flessibilità nell’affrontare i mutamenti dell’ambiente. La fierezza con la quale un determinato maschio lotta costituisce dunque solamente una componente della strategia, di origine ereditaria, che esso utilizza per affrontare il suo ambiente sociale.
La misurazione dei livelli di testosterone plasmatico nei diversi ceppi di topi appena descritti ha mostrato che la latenza all’attacco non è correlata ai livelli di testosterone circolante nei maschi adulti, e dunque è chiaro che le differenze comportamentali non sono il risultato di differenze nella produzione di androgeni indotte dal cromosoma Y in età adulta. Se i topi appartenenti alle due linee parentali e i loro reciproci ibridi vengono castrati al cinquantesimo giorno di età (subito dopo la pubertà) e si inietta loro una dose standard di testosterone, le differenze di latenza tra i diversi ceppi persistono. Ciò indica che le differenze comportamentali originarie dipendono da una maggiore sensibilità, negli animali con una latenza all’attacco bassa, alla stimolazione esercitata dagli androgeni circolanti durante lo sviluppo ontogenetico. Sembra infatti che una diversità nella quantità di androgeni circolanti durante la pubertà, indotta da elementi contenuti nella porzione appaiata del cromosoma Y, produca una variazione nella sensibilità agli androgeni circolanti, che a sua volta causa un aumento nella rapidità di attacco quando è raggiunta l’età adulta.
Costi e benefici dell’aggressività: la teoria dei giochi
Molto è stato scritto recentemente sul significato adattativo dell’aggressività. Si tratta di capire come e quanto gli animali traggano beneficio, in termini di fitness, dal comportarsi in modo aggressivo durante gli scontri tra conspecifici. A questo scopo, si è rivelato promettente l’approccio economico dell’ecologia comportamentale, basato su modelli matematici che calcolano i costi e i benefici delle diverse opzioni comportamentali e ne deducono quale sia quello che massimizza il guadagno netto in termini di fitness. In particolare, una grande influenza è stata esercitata dalla teoria dei giochi, che analizza la correlazione tra costi e benefici del comportamento aggressivo e la frequenza con cui questo è messo in atto. Questo approccio implica innanzitutto la definizione delle diverse opzioni comportamentali a disposizione di un’ipotetica popolazione animale.
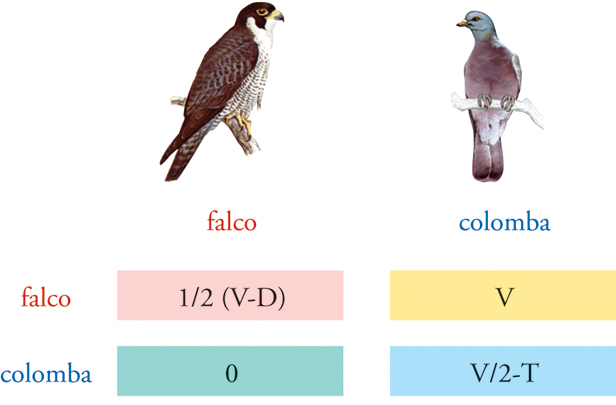
Per esempio, nel famoso modello falco-colomba, che è stato elaborato al fine di comprendere perché i conflitti vengano spesso risolti mediante i soli segnali di esibizione anche nei casi in cui il vincitore abbia buone probabilità di accaparrarsi una risorsa di notevole valore, le due opzioni o strategie consistono nell’utilizzo dei soli segnali di esibizione (colombe) oppure nell’utilizzo del solo scontro fisico diretto (falchi). Le ipotesi su come i conflitti vengano risolti e sui costi e i benefici delle diverse strategie sono formalizzate matematicamente mediante la matrice dei tornaconti (fig. 4), che definisce il guadagno netto derivante dalla combinazione delle strategie seguite da un individuo con le strategie seguite dal suo rivale. Quando il falco combatte contro un altro falco ha le stesse possibilità di vincere o rimanere ferito dal suo avversario (fig. 4, box rosa, dove V sta per il valore della vittoria e D per il costo di un ferimento grave); se combatte contro la colomba il falco vince sempre (box giallo) perché l’avversario si arrende sempre. Quando la colomba combatte contro il falco non vince mai ma neanche rimane ferita (box verde); se combatte con un’altra colomba vince nella metà dei casi ed entrambi gli avversari perdono tempo (box celeste, dove T sta per costo dei segnali di esibizione in termini di tempo).
In altre parole, a partire dalla frequenza relativa delle diverse strategie e dai tornaconti netti, in questo modello è possibile calcolare i guadagni e le perdite in termini di fitness cui andrebbero incontro gli individui nel caso in cui adottassero ciascuna strategia. In particolare, è spesso possibile identificare tra le strategie quella (o quelle) che, se fosse seguita dalla maggior parte degli individui appartenenti all’ipotetica popolazione, non potrebbe essere soppiantata da nessun’altra (naturalmente tra quelle definite). Queste strategie sono definite ‘evolutivamente stabili’ e corrispondono al modo in cui dovrebbero comportarsi gli animali reali, qualora il modello descriva correttamente le conseguenze adattative degli incontri aggressivi.
Il modello falco-colomba prevede per esempio che, se lo scontro fisico diretto comporta costi elevati, la strategia del falco (cioè ingaggiare subito un intenso combattimento) non è evolutivamente stabile e potrebbe essere sostituita dalla strategia della colomba. Questo perché, se ipotizziamo una bassa frequenza di aggressioni, le colombe, sebbene non vincano mai, non pagano mai costi notevoli a causa delle ferite, mentre i falchi rimangono gravemente feriti nel 50% dei casi. Il modello mostra anche che, se i benefici della vittoria non superano i costi dovuti alle ferite, la strategia evolutivamente stabile sarebbe costituita da una combinazione di falchi e colombe, in cui le due strategie finirebbero per essere equivalenti in termini di fitness. Per poter mettere ulteriormente a fuoco il problema delle conseguenze dello scontro fisico diretto in termini di fitness, nel modello falco-colomba si assume che i due rivali, pur differendo nella tattica di combattimento utilizzata, siano per il resto del tutto identici. Naturalmente è molto improbabile che nella realtà possa esistere una simile circostanza, e al contrario i due rivali si differenziano solitamente per molti aspetti, tra cui le dimensioni corporee (che influenzano pesantemente l’esito dei combattimenti) e l’importanza che la risorsa contesa ha per ciascuno di loro. Sono stati condotti molti studi, sia di natura teorica sia di natura empirica, sull’influenza che le capacità di lotta e il valore della risorsa dovrebbero esercitare e di fatto esercitano sul modo di combattere degli animali.
Un noto modello che descrive le capacità di lotta, il gioco di valutazione sequenziale (sequential assessment game), definisce i combattimenti come interazioni il cui scopo è quello di raccogliere, pagando il minor costo possibile, informazioni sulle capacità di lotta dell’avversario modulate dal valore della risorsa contesa; il modello descrive cioè quel che avviene durante un incontro aggressivo come se si trattasse di un processo di campionamento statistico tra due rivali che inizialmente ignorano completamente le rispettive capacità di lotta ma che, a ogni scambio comportamentale, acquisiscono informazioni sempre più precise l’uno sull’altro. L’individuo meno abile (che di solito è anche il più piccolo) desiste quando viene raggiunta una combinazione di soglia tra la stima che esso fa dello svantaggio che gli deriva dalle diverse dimensioni corporee e l’accuratezza di questa stima. Questo modello prevede, tra le altre cose, che gli scontri tra individui assortiti omogeneamente, durante i quali le rispettive capacità sono più difficili da valutare, si trasformeranno più facilmente in combattimenti intensi. Tale predizione è confermata empiricamente dall’osservazione del comportamento dei Pesci Ciclidi (Nannacara anomala) maschi: essa suggerisce che per quanto riguarda questa specie l’acquisizione a basso costo di informazioni sulle capacità di lotta dell’avversario costituisce un’importante funzione delle interazioni aggressive.
Per quel che riguarda il valore della risorsa contesa, ci si aspetta che quanto più esso è elevato, tanto più lunghi e intensi siano i combattimenti e che, quando gli avversari differiscono per i benefici che l’acquisizione della risorsa apporterebbe loro in termini di fitness, l’individuo che ha più da guadagnare lotti più intensamente e vinca. Questa predizione è supportata, per esempio, dal fatto che la durata delle lotte tra i maschi adulti del granchio della rena (Necora puber) aumenta se nell’acqua viene aggiunto – per simulare la presenza di una risorsa di valore – l’odore di una femmina sessualmente ricettiva. Inoltre, bisogna sottolineare che la maggior durata dei combattimenti in presenza dell’odore della femmina è dovuta soprattutto al fatto che in questo caso i granchi più piccoli desistono più tardi dalla lotta rispetto alla situazione di controllo. Di conseguenza i granchi più grandi non hanno più una maggiore probabilità di vincere rispetto a essi. Da ciò si può dedurre che una femmina costituisca una risorsa di maggior valore per gli individui più piccoli (forse perché in condizioni naturali essi hanno poche speranze di giungere di nuovo così vicino alla meta) e che questi sono dunque disposti a pagare notevoli costi prima di arrendersi
Bibliografia
Archer 1987: Archer, John, The behavioural biology of aggression, Cambridge, Cambridge University Press, 1987.
Bakker 1986: Bakker, Theo C.M., Aggressiveness in sticklebacks (Gasterosteus aculeatus L.): a behaviour-genetic study, “Behaviour”, 98, 1986, pp. 1-144.
Bass 1992: Bass, Andrew H., Dimorphic male brains and alternative reproductive tactics in a vocalising fish, “Trends in neurosciences”, 15, 1992, pp. 139-145.
Becker 1992: Behavioral endocrinology, edited by Jill Becker, Marc S. Breedlove and David Crews, Cambridge (Mass.)-London, MIT Press, 1992.
Benus 1988: Benus, R.F., Aggression and coping, Groningen, Diss. Rijksuniversiteit, 1988.
Benus 1991: Benus, R.F. e altri, Heritable variation for aggression as a reflection of individual coping strategies, “Experientia”, 47, 1991, pp. 1008-1019.
Christiansen, Jobling 1990: Christiansen, Jørgen S. - Jobling, Malcolm, The behaviour and the relationship between food intake and growth of juvenile Arctic charr, Salvelinus alpinus L., subjected to sustained exercise, “Canadian journal of zoology”, 68, 1990, pp. 2185-2191.
De Ruiter 1992: De Ruiter, Anne J.H. e altri, Differential testosterone secretory capacity of the testes of aggressive and non-aggressive house mouse during ontogeny, “Aggressive behaviour”, 18, 1992, pp. 149-157.
Enquist 1990: Enquist, Magnus e altri, A test of the sequential assessment game: fighting in the cichlid fish Nannacara anomala, “Animal behaviour”, 40, 1990, pp. 1-14.
Hews 1994: Hews, Diane K. - Knapp, Rosemary - Moore, Michael C., Early exposure to androgens affects adult expression of alternative male types in tree lizards, “Hormones and behaviour”, 28, 1994, pp. 96-115.
Huntingford 1982: Huntingford, Felicity A., Do inter- and intra-specific aggression vary in relation to predation pressure in sticklebacks?, “Animal behaviour”, 30, 1982, pp. 909-916.
Huntingford, Turner 1987: Huntingford, Felicity A. - Turner, Angela K., Animal conflict, London, Chapman and Hall, 1987.
Maynard Smith 1974: Maynard Smith, John, The theory of games and the evolution of animal conflicts, “Journal of theoretical biology”, 47, 1974, pp. 209-221.
Maynard Smith 1982: Maynard Smith, John, Evolution and the theory of games, Cambridge, Cambridge University Press, 1982.
Morris 1958: Morris, Desmond, The reproductive behaviour of the ten-spined stickleback (Pygosteus pungitius), Leiden, Brill, 1958.
Onions 1973: The shorter Oxford English Dictionary, edited by Charles T. Onions, Oxford, Clarendon, 1973.
Sinervo, Lively 1996: Sinervo, Barry - Lively, Curtis M., The rock-paper-scissor game and the evolution of alternative mating strategies, “Nature”, 380, 1996, pp. 240-243.
Smith 1994: Smith, Ian P. e altri, Strategic decisions during agonistic behaviour in the velvet swimming crab, Necora puber (L), “Animal behaviour”, 47, 1994, pp. 885-894.
Turnbull 1996: Turnbull, James F. - Richards, Randolph H. - Robertson, Darrell A., Gross, histological and scanning electron microscopic appearance of dorsal fin rot in farmed Atlantic salmon, Salmo salar L., “Journal of fish diseases”, 19, 1996, pp. 415-427.
Van Oortmerssen, Bakker 1981: Van Oortmerssen, Geert A. - Bakker, Theo C.M., Artificial selection for short and long attack latencies in wild Mus musculus domesticus, “Behavior genetics”, 2, 1981, pp. 115-126.
Van Oortmerssen, Sluyter 1994: van Oortmerssen, Geert A. - Sluyter, Frans, Studies on wild house mice. V. Aggression in lines selected for attack latency and their Y-chromosomal congenics, “Behavior genetics”, 24, 1994, pp. 73-78.