
Adattamento
Adattamento
di Giorgio Morpurgo e Richard Ch. Lewontin
ADATTAMENTO
Adattamento fisiologico di Giorgio Morpurgo
sommario: 1. Introduzione. 2. L'adattamento fisiologico nei Procarioti. 3. Fenomeni di adattamento negli Eucarioti inferiori. 4. L'adattamento fisiologico in organismi pluricellulari. 5. L'adattamento fisiologico alla temperatura: generalità. 6. L'adattamento fisiologico alla temperatura nelle piante. 7. L'adattamento fisiologico alla temperatura nei Vertebrati. 8. Adattamento fisiologico e sistema nervoso. □ Bibliografia.
1. Introduzione
Con l'espressione ‛adattamento fisiologico' si intende generalmente la capacità propria di ogni organismo vivente di reagire alle variabili condizioni ambientali in modo da adattare il suo fenotipo alle nuove condizioni. Si usa anche distinguere l'adattamento fisiologico dall'adattamento genetico: quest'ultimo, infatti, sarebbe il risultato di cambiamenti ereditari che con il trascorrere delle generazioni adeguano l'organismo all'ambiente in cui vive, mentre il primo - nella comune accezione - comporterebbe soltanto variazioni non ereditarie, limitate quindi al singolo individuo e non ai suoi discendenti. In realtà il problema è assai più complesso e non è possibile separare completamente i due concetti: la capacità dell'organismo di reagire ad un determinato stimolo dipende prima di tutto dai sistemi genetici dell'individuo stesso, sistemi che permettono, in relazione alle variazioni ambientali, l'oscillazione del carattere entro un certo ambito. Nello stesso organismo l'ambito di oscillazione può essere estremamente differente per i diversi caratteri: così in un particolare batterio la sintesi di alcuni enzimi, chiamati ‛enzimi adattativi', può variare di migliaia di volte in dipendenza di fattori ambientali, mentre altri enzimi vengono sintetizzati con velocità più o meno costante malgrado il variare delle condizioni ambientali. In tutti e due i casi è però bene ricordare, se si vuole arrivare ad una spiegazione non superficiale dei fenomeni di adattamento fisiologico, che la possibilità o meno di reagire allo stimolo esterno dipende dalla costituzione genetica della cellula, costituzione che è il risultato dell'adattamento genetico evolutivo. Studiare a fondo i fenomeni di adattamento fisiologico vuol dire perciò non accontentarsi della descrizione della variazione del carattere in risposta allo stimolo, ma capire come il sistema genetico consenta l'esistenza della variazione nell'ambito che osserviamo.
Non è ovviamente possibile nello spazio di un articolo dare una descrizione, sia pure sommaria, di tutti gli innumerevoli fenomeni di adattamento noti nelle diverse forme viventi. Cercheremo invece di dare qualche esempio che ne illustri i meccanismi fondamentali finora noti ed i problemi che restano da risolvere. Nel far ciò cercheremo di avere una visione il più possibile evolutiva, dato che diverso è il comportamento fisiologico ai vari gradini della scala evolutiva, tanto che si può certamente parlare di una evoluzione dei sistemi di adattamento fisiologico.
2. L'adattamento fisiologico nei Procarioti
Gli organismi con piano strutturale più semplice che conosciamo, e, verosimilmente, che esistono oggi sulla Terra, sono i Batteri o più in generale i Procarioti. Con quest'ultimo termine si comprendono i Batteri e le Alghe azzurre, che hanno sul piano cellulare una struttura fondamentale comune e ben differenziata da quella degli organismi cellulari o Eucarioti. Le differenze sono talmente importanti che è lecito affermare che uno dei passi fondamentali dell'evoluzione è stato proprio il passaggio dalla struttura cellulare di tipo procariotico alla struttura cellulare di tipo eucariotico. Conviene qui riassumere brevemente tali differenze, perché ad esse sono certamente collegati importantissimi cambiamenti nei sistemi di adattamento.
Le caratteristiche fondamentali dei due gruppi sono elencate nella tab. I. Le differenze ivi indicate sono estremamente nette: questo vuol dire che al primo tipo di struttura appartengono tutti gli organismi classificati fra i Procarioti ed al secondo tutti gli organismi classificati fra gli Eucarioti, e che a tutt'oggi non si conoscono organismi con caratteristiche intermedie fra i due gruppi. Ciò significa che da un punto di vista cellulare un lievito è molto più vicino ad un uomo che ad un altro microrganismo di tipo batterico.

La differenza fondamentale che condiziona tutte le altre è probabilmente la penultima: l'intera struttura del Batterio è condizionata dalla necessità di poter adeguare velocemente gran parte dei suoi sistemi sintetici alle variazioni ambientali. Di fatto vediamo che la maggior parte dei sistemi di sintesi degli enzimi può variare nei Batteri in maniera incredibile. Per esempio il batterio Escherichia coli, comune ospite del nostro intestino, è normalmente incapace di scindere il lattosio nei suoi componenti; tuttavia, dopo un breve periodo di tempo di contatto con il lattosio o altra sostanza che abbia un legame β-galattosidico, le cellule di coli iniziano la sintesi dell'enzima β-galattosidasi, capace di scindere il lattosio, con una tale intensità che l'enzima può raggiungere una concentrazione pari al 3% di tutte le proteine cellulari. Un altro esempio ben noto è la produzione, da parte di alcuni batteri, di penicillinasi, un enzima capace di distruggere la penicillina. Questo tipo di sintesi enzimatica prende il nome di sintesi adattativa in quanto l'enzima si forma solo come risposta ad uno stimolo ambientale. Apparentemente opposto è il sistema di sintesi reprimibile: un batterio come il coli è capace di sintetizzare tutti i metaboliti essenziali a partire da sali minerali e da glucosio come fonte di energia. Quando si usa un terreno culturale di questo tipo è quindi necessario che nelle cellule esista una catena sintetica per ogni metabolita essenziale, catena biosintetica che deve funzionare con una velocità adatta al ritmo riproduttivo della cellula: infatti una velocità di sintesi eccessiva porterebbe ad uno spreco, una velocità troppo bassa costituirebbe un fattore limitante della crescita. Se poi il prodotto finito, il metabolita, si trova già preformato nel terreno culturale, è inutile sintetizzarlo, e porta ad un risparmio di energia interrompere completamente la sintesi degli enzimi deputati alla costruzione del metabolita. Ciò è esattamente quanto succede: la cellula batterica ha di norma un sistema che le permette di regolare in maniera coordinata la sintesi degli enzimi interessati ad una determinata via metabolica in dipendenza della concentrazione intracellulare del metabolita. La base per la interpretazione del funzionamento di questo sistema di controllo fu posta da una ricerca di M. Demerec (v., 1964), il quale osservò in un batterio del genere Salmonella che i geni interessati ad una stessa catena biosintetica si trovano localizzati l'uno vicino all'altro. L'osservazione di Demerec è stata ben presto estesa ad altre specie batteriche. Il merito principale nello stabilire i principi operativi del sistema di controllo della sintesi enzimatica spetta alle ricerche di F. Jacob e J. Monod (v., 1961), che hanno definito in maniera generalmente valida per i Batteri il funzionamento sia dei sistemi reprimibili, sia di quelli inducibili.
Questi sistemi consistono: a) di una serie di geni ‛strutturali', cioè di geni che contengono l'informazione necessaria a costruire, a dare la struttura primaria agli enzimi; b) di un gene chiamato ‛operatore', adiacente ai geni strutturali; recentemente si è visto (v. Beckwith, 1967) che in realtà il gene operatore è diviso in due unità funzionali, il promotore che è il punto di attacco dell'RNA-polimerasi e l'operatore propriamente detto che ha per così dire le funzioni di un interruttore; c) di un gene che produce una sostanza chiamata ‛repressore', più tardi identificata come una proteina, la quale può avere affinità per l'operatore. Quando il repressore è legato all'operatore i geni strutturali non vengono trascritti dall'RNA-polimerasi in RNA messaggero e non si ha quindi sintesi enzimatica. Se però il repressore trova nel citoplasma molecole dell'induttore (lattosio nel caso della galattosidasi, penicillina nel caso della penicillinasi) la specifica molecola proteica che costituisce il repressore cambia la sua struttura, perde l'affinità per l'operatore e si sblocca la sintesi dell'RNA messaggero e degli enzimi.
Il contrario avviene nel caso di una sintesi reprimibile, per esempio nel caso del sistema biosintetico dell'istidina studiato a fondo da B. Ames (v. Ames e altri, 1967). In questo caso il repressore non ha di norma affinità per l'operatore e la sintesi degli enzimi procede regolarmente; se però la concentrazione dell'istidina nel citoplasma supera un certo livello, il repressore si lega all'amminoacido, cambia la sua struttura e acquista un'affinità per l'operatore bloccando in tal modo la sintesi degli enzimi. Questo schema è stato formulato da Jacob e Monod nel 1961 essenzialmente sulla base del comportamento fisiologico di mutanti che presentano difetti nella regolazione, e lo schema è rimasto fondamentalmente valido. Naturalmente si è trovato successivamente che spesso la situazione è un po' più complicata di quella descritta e che esistono delle eccezioni alla regola della repressione o attivazione coordinata; tuttavia nelle sue linee generali lo schema è certamente valido per la quasi totalità dei sistemi enzimatici dei Batteri. Come abbiamo già detto, negli ultimi anni (v. Gilbert e Muller-Hill, 1966) è stato anche isolato e parzialmente purificato uno dei repressori, che è risultato essere una proteina le cui proprietà corrispondono esattamente a quelle previste dalla teoria.
Questo sistema è integrato da altri due che ne costituiscono il necessario complemento. Anche se non si ha sintesi di RNA messaggero, la sintesi degli enzimi non dovrebbe cessare immediatamente, perché dovrebbe continuare la traduzione dell'RNA messaggero preformato. La cellula batterica ha risolto anche questo problema, perché la cosiddetta vita media (si intende per vita media il tempo in cui la concentrazione della molecola si riduce a metà) del messaggero è soltanto di pochi minuti: anche questo sistema è quindi adattato per interrompere la sintesi degli enzimi in un tempo brevissimo, eliminando la possibilità che gli enzimi continuino a venir prodotti attraverso la lettura di messaggeri preformati.
Infine esiste ancora un sistema che permette alla cellula di regolare la sintesi del prodotto finale delle catene biosintetiche in una maniera pressoché istantanea, portando ad una variazione dell'attività del primo enzima della catena biosintetica.
Questo sistema è chiamato ‛sistema di regolazione allosterica', o anche ‛sistema di retroinibizione', e fu descritto per la prima volta da Monod, Changeux e Jacob (v., 1963). In una normale catena biosintetica (facciamo di nuovo l'esempio dell'istidina) il primo enzima della catena, in questo caso la fosforibosil-ATP-pirofosforilasi, ha un sito diverso dal sito attivo, chiamato ‛sito allosterico', che è capace di legare il prodotto finale della catena biosintetica. L'enzima legato all'istidina subisce un cambiamento conformazionale da cui consegue la perdita dell'attività enzimatica. Il mancato funzionamento del primo enzima produce l'assenza di substrato per il secondo enzima della catena ed in tal modo l'attività biosintetica dell'intera catena viene a cessare. È evidente che in questa maniera la concentrazione del prodotto finale, regolando ad ogni momento l'attività catalitica della catena biosintetica, regola con estrema rapidità, adeguandola alle necessità, la sua stessa produzione. In altri casi più complessi esistono regolatori allosterici che inibiscono l'attività dell'enzima, come nell'esempio che abbiamo fatto, ed altri regolatori che al contrario ne aumentano l'attività.
Dobbiamo ancora ricordare che a questi sistemi di regolazione ormai classici negli ultimi anni se ne è aggiunto un altro, anche questo certamente di enorme importanza, e cioè un sistema che si basa sulla concentrazione di AMP ciclico nella cellula. Per esempio la produzione di galattosidasi da parte del coli dipende dalle concentrazioni di AMP ciclico e di una proteina che si lega all'AMP ciclico da una parte e dall'altra al DNA in un locus vicino all'operatore. Così anche la scelta fra ciclo lisogeno e ciclo virulento da parte di un fago temperato dipende dalla concentrazione intracellulare di AMP ciclico. In questi due casi l'effetto regolatore si esercita al livello della trascrizione. In innumerevoli altri casi sia nei Procarioti sia negli Eucarioti superiori sembra che l'AMP ciclico svolga una funzione regolatrice che opera al momento della traduzione. È perciò impossibile al momento attuale riunire in un unico schema le innumerevoli osservazioni che riguardano l'attività di questo composto (v. Jost e Rickenberg, 1971).
Se adesso consideriamo la cellula batterica da un punto di vista generale, possiamo fare le seguenti considerazioni: un normale batterio come il coli possiede circa 2-3.000 geni e di questi certamente una gran parte sono implicati nella codificazione degli enzimi delle normali vie metaboliche. Tutte queste vie sono dotate di sistemi rapidi ed efficienti per adattare sia la sintesi sia l'attività degli enzimi alle necessità metaboliche, e quindi il livello della maggior parte delle funzioni metaboliche di un batterio può oscillare di migliaia di volte fra il valore iniziale ed il valore massimo dopo induzione, in relazione alle fluttuazioni esterne. Tranne poche eccezioni lo sviluppo delle funzioni metaboliche di un batterio non segue un programma prefissato, ma è al contrario qualcosa di molto fluido ed adattabile. Nonostante questa sia la condizione generale, è però interessante notare che nei bacilli esiste una forma di adattamento che, in certe condizioni, comporta lo scatenamento di un programma interno prefissato. Si tratta di un adattamento fisiologico alle cattive condizioni ambientali, ma l'adattamento non consiste solamente nel reprimere o attivare una sintesi enzimatica, bensì nel promuovere un programma preordinato che trasforma completamente la cellula in una forma di resistenza o spora che permette al batterio di resistere per un tempo indefinito in stato di quiescenza, anche in condizioni di temperatura esterna molto sfavorevoli. Questo caso è assai interessante perché costituisce nei Batteri forse l'unico esempio di adattamento fisiologico preordinato e assomiglia quindi, almeno formalmente, al differenziamento degli organismi superiori.
È anche interessante notare che, per quanto riguarda la temperatura, non esiste fondamentalmente, nell'ambito dei Procarioti, alcuna altra forma di adattamento fisiologico. Si conoscono batteri che vivono e si moltiplicano alle temperature più diverse fra o e 101 °C (v. Brock, 1970). Tuttavia ogni specie ha la sua temperatura ottimale ed un ambito non modificabile entro cui può vivere e moltiplicarsi, ambito che normalmente è di circa 20-30 °C. La temperatura ottimale, cioè quella che assicura la massima velocità di crescita, è generalmente molto vicina al limite superiore oltre il quale anche un piccolo aumento è letale. Variando la temperatura il batterio non si adatta, ma semplicemente varia il suo ritmo di accrescimento. Nelle specie batteriche quindi esiste un notevolissimo grado di adattamento genetico alla temperatura, tanto che le diverse specie batteriche possono colonizzare praticamente ogni ambiente fra la temperatura di congelamento e quella di ebollizione dell'acqua (oltrepassando, sia pure di poco, questi limiti in ambienti particolari) mentre l'adattamento fisiologico è scarso o nullo. Ciò non deve sorprendere perché la messa in opera di meccanismi di adattamento fisiologico alla temperatura (a parte l'entrare in stato quiescente) comporta la risoluzione di problemi complicatissimi. Infatti, data la relazione fra attività enzimatica e temperatura, sarebbe necessario un adeguamento dell'attività di ogni enzima al variare della temperatura, il che comporterebbe probabilmente una complicazione eccessiva per cellule relativamente semplici come quelle procariotiche.
3. Fenomeni di adattamento negli Eucarioti inferiori
Istintivamente si è portati a pensare che un lievito o una muffa siano più simili ad un batterio che alla cellula di un organismo superiore, come per esempio un mammifero. Come abbiamo già avuto modo di notare (v. tab. I) è invece esattamente il contrario. Sia da un punto di vista strutturale, sia da un punto di vista fisiologico un lievito è un organismo ormai lontanissimo da un batterio. Negli ultimi anni si sono moltiplicati gli studi per cercare di capire come negli Eucarioti inferiori siano regolati i processi di sintesi adattativa degli enzimi. Per il momento non si è riusciti a costruire un quadro chiaro, ma chiare sono soltanto le conclusioni negative che possono essere riassunte nella maniera seguente.
1. I fenomeni di adattamento della sintesi degli enzimi sono generalmente molto più ridotti di quanto non lo siano nei Batteri; un enzima adattativo in un lievito o in una muffa può oscillare rispetto al valore basale di un fattore dieci o venti, e cioè l'ampiezza dell'oscillazione è sempre enormemente inferiore a quella che si osserva nei Batteri.
2. Viene conservato inalterato (e inalterato resta in tutto il resto della scala evolutiva) il sistema di regolazione allosterica dell'attività degli enzimi, il quale viene quindi ad acquistare una importanza relativa molto maggiore.
3. Non è ancora del tutto chiaro in alcun caso come funzioni il sistema di regolazione della sintesi degli enzimi. È comunque certo che, tranne rare eccezioni, i geni non stanno più raggruppati in unità funzionali, gli operoni, per cui non è più possibile un sistema di sintesi coordinata del tipo di quello batterico. Vedremo più avanti, in questo articolo, che probabilmente la scomparsa del sistema di sintesi coordinata è legata strettamente alla comparsa dei fenomeni differenziativi.
Almeno nel lievito sembra inoltre (v. Tauro e Halvorson, 1966) che ai normali sistemi di adattamento se ne sovrapponga un altro che dipende in qualche modo dal ciclo riproduttivo della cellula: Halvorson ha infatti visto, in studi condotti su popolazioni sincrone di lievito, che la produzione, sia questa costitutiva o adattativa, degli enzimi avviene solamente ad un determinato momento del ciclo cellulare, momento che è differente per i vari geni e che dipende dalla posizione del gene sul cromosoma.
Esiste però una importantissima eccezione al punto 1; esistono cioè dei sistemi di enzimi che nelle cellule degli Eucarioti si comportano, se si considera l'entità dei fenomeni di adattamento, come gli enzimi dei Batteri. Si tratta dei sistemi di enzimi legati alla respirazione, come i citocromi, e degli enzimi del ciclo di Krebs, enzimi che sono del tutto assenti nelle cellule di lievito coltivate in anaerobiosi totale. Tutti questi enzimi sono rapidamente sintetizzati in seguito ad una semplice incubazione in un tampone glucosato ed aerato: in capo a due o tre ore la concentrazione degli enzimi arriva al suo massimo. Il processo di adattamento respiratorio si può studiare chiaramente soltanto nel lievito, perché questo è l'unico eucariote che possa crescere perfettamente in condizioni sia aerobiche che anaerobiche. Data la costanza della struttura e della funzione dei mitocondri in tutte le cellule eucariotiche, è tuttavia probabile che ciò che è valido per il lievito sia generalmente valido per tutti i gradini della scala evolutiva. Recentemente, del resto, si è potuto vedere che la citocromossidasi viene indotta dall'ossigeno (v. Nakami e Pious, 1967) in cellule di Mammiferi in coltura, e un simile dato rinforza queste conclusioni.
Ciò che rende interessante questo fenomeno è il fatto che gli unici enzimi fortemente adattativi sono quelli legati al mitocondrio (anche se i geni che codificano enzimi hanno probabilmente una localizzazione nucleare), organulo che sotto vari aspetti strutturali e funzionali ricorda un batterio: di tipo batterico è il sistema di sintesi proteica proprio del mitocondrio, e di tipo batterico è anche la struttura del cromosoma mitocondriale. Questa osservazione ci può portare a formulare l'ipotesi che nell'evoluzione si sia sempre conservato, all'interno della cellula eucariotica, un doppio sistema genetico e di sintesi proteica per poter consentire alle funzioni che devono poter oscillare entro ampi limiti di rimanere funzionalmente legate alla struttura di tipo batterico del mitocondrio. Lo stesso discorso si può ripetere a proposito del cloroplasto nella cellula vegetale. Anche nelle cellule vegetali infatti la maggior parte degli enzimi sono costitutivi: come nel caso precedente vediamo però che gli enzimi legati al processo fotosintetico sono rapidamente adattabili in risposta alla luce. Come il mitocondrio, il cloroplasto ricorda molto la sua presunta origine batterica sia per la struttura del sistema di sintesi proteica sia per la struttura del DNA.
Considerando le cose da un punto di vista evolutivo, potremmo quindi concludere che il progresso maggiore che si è avuto nel passaggio dalla cellula di tipo procariotico a quella di tipo eucariotico è consistito nel cambiare i sistemi di regolazione della sintesi degli enzimi in modo da renderli meno sensibili alle fluttuazioni esterne. In tal modo la cellula eucariotica ha acquisito una maggior costanza del suo ambiente interno e la possibilità di svolgere un programma prefissato.
Tuttavia le funzioni che era necessario continuassero ad oscillare in dipendenza delle variazioni ambientali sono rimaste legate ad organuli cellulari che hanno conservato la struttura e la capacità di adattamento tipiche del batterio. Non è comunque chiaro a prima vista perché la cellula abbia rinunciato a conservare - o, se vogliamo mettere il problema in termini non finalistici, perché sia risultato selettivamente poco vantaggioso conservare - il sistema ad operoni e il sistema di controllo coordinato. Non è chiaro perché la cellula abbia trovato utile disperdere i geni con funzioni in parti differenti del genoma. Non è possibile dare una risposta sicura a questa domanda; una risposta plausibile potrebbe essere la seguente: l'Eucariote, raffrontato al Procariote, dà una maggiore importanza, come abbiamo visto, allo sviluppo di un programma interno rispetto al diretto adattamento alle condizioni esterne. Sviluppo di un programma interno vuol dire che geni diversi devono venire attivati a momenti sequenziali e differenti. Per esempio, nell'uomo, ad un determinato momento dello sviluppo è attivo il gene che determina la struttura della catena dell'emoglobina fetale, mentre in uno stadio più tardivo è attivo il gene che determina la struttura della catena β tipica dell'emoglobina adulta. Lo stesso succede certamente per numerose attività enzimatiche. Se esaminiamo adesso la struttura primaria delle due catene γ e β, vediamo che fra loro esistono fortissime somiglianze strutturali tanto che possiamo essere sicuri che i due geni, che codificano rispettivamente la catena γ e β, derivano l'uno dall'altro per duplicazione. Certamente per duplicazione di un gene ancestrale comune derivano anche i geni che codificano la catena α, la catena della mioglobina e la catena ε tipica dell'emoglobina embrionale. Se i geni rispettivi fossero rimasti contigui in un sistema operonico, necessariamente sarebbero stati soggetti allo stesso tipo di regolazione: tutti sarebbero entrati contemporaneamente in funzione e contemporaneamente sarebbero ritornati allo stato quiescente, il che non è evidentemente compatibile con un tipo di sviluppo che richiede un'attivazione sequenziale nel tempo. Di qui la necessità, con il sorgere di un programma di sviluppo, di spezzare i raggruppamenti operonici ed il sistema di controllo coordinato disperdendo i geni in diverse parti del genoma. È chiaro che non si tratta che di un'ipotesi, la quale però ha il vantaggio di fare una previsione soggetta a verifica sperimentale, perché prevede che l'ordine dei geni in un organismo eucariotico sia in qualche modo legato al momento in cui il gene deve entrare in attività.
4. L'adattamento fisiologico in organismi pluricellulari
I problemi dell'adattamento fisiologico diventano immediatamente molto più complessi non appena nella scala evolutiva si passa da strutture unicellulari con scarso differenziamento a organismi multicellulari complessi. Fermo restando che la base di ogni fenomeno va ricercata alla fine in cambiamenti a livello molecolare, noi vediamo adesso che negli organismi superiori non ci si può limitare a cercare la risposta adattativa all'ambiente come cambiamento di una o più attività enzimatiche.
L'adattamento fisiologico comporta spesso, oltre a variazioni in una particolare molecola, variazioni in interi organi o sistemi. Ogni fenomeno va pertanto studiato e descritto a due livelli differenti: vanno cioè descritte le variazioni nell'individuo indotte da un determinato stimolo e, se possibile, queste variazioni in ogni organo o sistema interessato vanno studiate a livello molecolare. Un compito già difficile diviene quindi enormemente più complesso. Inoltre nel cercare di dare interpretazioni a livello molecolare siamo ostacolati dal fatto che, come abbiamo visto, i meccanismi di regolazione della sintesi enzimatica non sono per niente chiari neanche negli Eucarioti inferiori. In quelli superiori compaiono due ulteriori complicazioni. Anzitutto, sia nel regno animale sia in quello vegetale compaiono delle molecole chiamate ‛ormoni' che esercitano effetti fondamentali sui più diversi organi, effetti che sappiamo derivare in alcuni casi da variazioni nella sintesi degli enzimi al livello dell'organo interessato: per esempio i corticosteroidi inducono la sintesi di tirosina transaminasi nel fegato e anche in cellule di epatoma in cultura, un ormone vegetale chiamato ‛gibberellina' induce nel pericarpo del seme la sintesi di amilasi, ecc.; ora, poiché molte delle variazioni indotte dall'ambiente sono mediate da variazioni nel livello degli ormoni, sarebbe necessario sapere come questi agiscono, ma anche in questo campo le nostre conoscenze sono molto scarse e soltanto ora cominciano, in qualche raro caso, ad avvicinarci alla comprensione del loro meccanismo di azione.
Negli animali, poi, le cose si complicano ulteriormente per la presenza del sistema nervoso, che esercita un'azione importantissima come mediatore dei nostri fenomeni adattativi. Bisognerebbe quindi avere un'idea chiara del funzionamento del sistema nervoso, idea che siamo ben lontani dall'avere, e poi capire come questo possa servire da mediatore. È chiaro quindi che anche nei casi più studiati non possiamo avere che un quadro frammentario dell'intero fenomeno.
Per cercare di dare un'idea della complessità del problema e di come questo vada affrontato (nell'evidente impossibilità nella quale ci troviamo di parlare di tutti gli infiniti fenomeni di adattamento nelle innumerevoli forme viventi), esporremo quanto si sa di un caso relativamente ben studiato, e cioè dei fenomeni di adattamento alla temperatura. Questo caso è stato scelto non soltanto perché gli studi sull'argomento sono relativamente numerosi, ma anche perché la temperatura è certamente una delle variabili più importanti nella vita di un organismo. Anche così delimitato il problema è ancora di gran lunga troppo vasto per poter essere affrontato in questa sede. Né d'altra parte è questo il nostro proposito, perché non si intende qui elencare i fatti finora noti, ma si tenta di mettere a fuoco i problemi principali.
5. L'adattamento fisiologico alla temperatura: generalità
Uno dei fattori fondamentali che condizionano la vita di tutti gli organismi viventi è certamente la temperatura ambientale. Tranne ambienti estremi, dove la temperatura può arrivare a circa 100 °C come nelle sorgenti calde e a −80 °C come in Antartide, la temperatura può oscillare fra −50 °C e +50 °C, ma naturalmente non sono molti gli organismi esposti, nell'arco della loro vita, ad oscillazioni di temperatura di questa ampiezza. Tuttavia oscillazioni di temperatura fra la minima invernale e la massima estiva di circa 60 °C non sono eccezionali, neppure nei climi temperati, per gli organismi terrestri. L'oscillazione è generalmente inferiore, ma può facilmente raggiungere i 30-40 °C per gli organismi viventi in acqua dolce. Si tratta sempre di oscillazioni molto forti cui l'organismo deve reagire in qualche maniera. Reagire, adattarsi, ha però qui due possibili significati: può significare semplicemente sopravvivere in condizioni di temperatura sfavorevoli, e può voler dire adattare il proprio metabolismo corporeo in modo da renderlo totalmente o parzialmente indipendente dalla temperatura esterna. In tutti e due i casi sono necessari molto spesso dei processi attivi di adattamento fisiologico che cercheremo adesso di descrivere con qualche dettaglio. Sarà comunque utile prima di tutto vedere quali sono le difficoltà cui va incontro un organismo in seguito alla variazione della temperatura.
Al di sotto del punto di congelamento dell'acqua il problema fondamentale per gli organismi che non regolino la propria temperatura corporea - e sono la grande maggioranza - è come impedire la formazione di ghiaccio intracellulare e quindi come resistere al processo di congelamento e scongelamento, che come è noto è un ottimo sistema per ottenere la rottura delle pareti cellulari e quindi la morte.
Al di sopra di una certa temperatura, che può essere di 50-60 °C, a seconda delle specie, si ha il problema, certamente non meno grave, di impedire la denaturazione irreversibile delle proteine cui conseguirebbe inevitabilmente la morte. Alle temperature intermedie la difficoltà fondamentale, da cui derivano tutte le altre, consiste nel fatto che con il diminuire della temperatura diminuisce l'attività degli enzimi, dimezzandosi generalmente (ma non sempre) per una diminuzione di dieci gradi. Ciò comporta ovviamente un rallentamento di tutte le attività, rallentamento che è stato messo in evidenza in maniera assai brillante in alcune ricerche condotte sulle formiche. Si è infatti visto che la velocità con cui camminano le formiche è proporzionale alla velocità di scissione dell'ATP e quindi all'attività ATP-asica che diminuisce con la temperatura. Presa quindi una determinata specie di formiche, misurando la velocità con cui cammina si può sapere la temperatura esterna e l'attività dell'enzima. Viceversa, conoscendo la velocità di scissione dell'ATP, si conosce di nuovo la temperatura esterna e la velocità di traslazione dell'insetto (v. Levy e altri, 1959).
Non tutti gli organismi possono fisiologicamente reagire alle variazioni di temperatura esterna: sono numerosissimi quelli che possono vivere soltanto in un certo ambito di temperatura, più o meno vasto, ed in questo ambito semplicemente reagiscono rallentando o aumentando la loro attività. Così si comportano per esempio i Batteri. Esistono specie, come abbiamo detto, che sono adattate a vivere a temperature diversissime, fra 0 e 100 °C, ma l'ambito di temperatura in cui possono moltiplicarsi è rigidamente prefissato e non può essere modificato, il che vuol dire che non hanno possibilità di adattamento fisiologico. In altri gruppi di organismi queste possibilità sono molto sviluppate ed esamineremo adesso alcuni di questi casi.
6. L'adattamento fisiologico alla temperatura nelle piante
L'adattamento alla temperatura nelle piante è stato relativamente molto meno studiato che negli animali. Esistono e sono stati descritti anche nel regno vegetale notevoli fenomeni adattativi; ne riporteremo qui un paio di esempi per dare un'idea dei problemi che si pongono alla pianta.
Uno dei problemi che le piante devono normalmente affrontare nel loro ciclo di vita è come resistere nell'inverno dei paesi freddi e temperati a temperature al di sotto del punto di congelamento. Di norma la pianta prima del sopraggiungere dell'inverno entra in uno stato di quiescenza che si manifesta nella maggior parte delle piante arboree dei nostri climi con la perdita delle foglie. Questo periodo di preparazione si rende evidente prima che sopravvengano le condizioni sfavorevoli: la quiescenza della pianta non è quindi la conseguenza dell'insorgere delle condizioni sfavorevoli, ma è uno stato che anticipa queste condizioni e prepara la pianta contro di esse. L'inizio della quiescenza e la caduta delle foglie in molte piante sono regolati dalla lunghezza del giorno, che diminuisce in autunno, e quindi dalla quantità della luce che la pianta riceve. Questo fenomeno è stato chiamato ‛fotoperiodismo' ed avviene verosimilmente attraverso variazioni nelle condizioni ormonali della pianta. Lo stimolo che serve a porre fine allo stato di quiescenza è un trattamento freddo, la cui lunghezza varia a seconda delle piante da qualche settimana a parecchi mesi, a temperature prossime o inferiori al punto di congelamento. Si tratta evidentemente di fenomeni di adattamento fisiologico (ma geneticamente determinati) che stabiliscono una correlazione fra il ciclo delle stagioni e l'attività della pianta, in modo che questa si esplichi solo nel periodo favorevole per la crescita.
Durante il periodo di quiescenza la temperatura può scendere anche di parecchie diecine di gradi sotto 0 °C, onde si è cercato di comprendere come la pianta possa sopravvivere a temperature inferiori al punto di congelamento, e quindi alla formazione di cristalli di ghiaccio nel suo interno. Questi ultimi si possono formare in due modi: se il raffreddamento è molto rapido, si formano all'interno della cellula e del vacuolo, e si realizza così un processo di congelamento, sempre letale; se invece la temperatura scende lentamente, come normalmente avviene in natura, la formazione di ghiaccio avviene negli spazi intercellulari a spese dell'acqua che trasuda dalle cellule adiacenti, e il ghiaccio si forma in grandi ammassi lenticolari fra strati di cellule disidratate e contratte.
Al momento dello scongelamento la pianta resterà viva o morirà a seconda dello stato fisiologico in cui si trovava al momento del congelamento. Studi accurati (v. Siminovič e altri, 1967) condotti su Robinia pseudoacacia hanno dimostrato che le cellule della corteccia prelevate in estate non tollerano un raffreddamento fino −10 °C, mentre le cellule invernali possono resistere al congelamento e successivo scongelamento fino alla temperatura dell'aria liquida. L'aumento di resistenza è molto brusco, ma è per ora difficile dire se ciò sia rigidamente determinato come ciclo circannuale o sia invece dipendente dalla caduta autunnale della temperatura o legato all'accorciarsi delle giornate. Ancor più difficile è comprendere quali siano i metodi con cui la pianta ottiene questo risultato. In effetti parallelamente al prodursi dell'aumento di resistenza al freddo si verificano cambiamenti metabolici fondamentali praticamente in tutti i sistemi cellulari, per cui è al momento impossibile concludere quale sia il meccanismo responsabile dell'aumentata resistenza. Mentre in questo caso è difficile dire se l'accresciuta resistenza al freddo sia indotta dall'abbassamento di temperatura o dipenda dal ciclo fisiologico, si è invece potuto accertare che le piante (sono stati fatti esperimenti su fagioli, tabacco, cocomero, girasoli, ecc.) possono aumentare molto rapidamente la propria resistenza a bruschi aumenti di temperatura. Gli esperimenti sono stati condotti (v. Yarwood, 1967) prendendo una coppia di foglie gemelle e sottoponendo una delle due per un periodo di tempo molto breve, dell'ordine della diecina di secondi, ad una temperatura elevata. Alcune ore dopo le due foglie venivano nuovamente trattate con una temperatura eccessivamente elevata per il tipo di pianta in esame. Di regola si aveva un'aumentata sopravvivenza della foglia sottoposta a trattamento in confronto a quella non pretrattata.
Anche in questo caso si dimostra l'esistenza di meccanismi rapidi ed efficienti per resistere a variazioni brusche di temperatura, ma non si ha la minima idea di come la pianta possa così rapidamente variare il livello della sua resistenza al calore.
7. L'adattamento fisiologico alla temperatura nei Vertebrati
In questo capitolo esamineremo i problemi di adattamento alla temperatura nei Vertebrati; la scelta di questo gruppo non è determinata dal fatto che i problemi siano fondamentalmente diversi da quelli esistenti in altri gruppi di organismi, ma dal fatto che nei Vertebrati gli studi sono relativamente più avanzati.
Teoricamente di fronte a forti variazioni della temperatura ambientale all'animale si presentano quattro possibilità.
1. Regolare la temperatura del corpo in modo da tenerla pressoché costante. Questo sistema è stato adottato dai due gruppi di Vertebrati più evoluti, i Mammiferi e gli Uccelli, ma ha il difetto di portare ad un enorme dispendio di energia. Una semplice considerazione evolutiva ci porta da questo fatto a concludere che se due gruppi indipendenti hanno scelto la dispendiosa via di termostatare il proprio corpo, è chiaro che gli altri sistemi sono necessariamente poco funzionali. A parte Uccelli e Mammiferi, una limitata possibilità di termostatazione può esistere anche in alcuni poichilotermi: per esempio nei rospi la temperatura corporea rimane più bassa di quella ambientale grazie alla continua evaporazione dalla pelle. Più raro è il caso di un poichiloterma che possa, sia pure eccezionalmente, elevare la propria temperatura corporea; forse l'unico caso è quello rappresentato dal pitone durante la cova delle uova. Questo rettile, infatti, attraverso la continua contrattura muscolare, riesce ad elevare la sua temperatura anche di dieci gradi al di sopra dell'ambiente. Casi del genere sono conosciuti anche in alcuni Insetti. Ovviamente nei poichilotermi che vivono nell'acqua anche questo scarso adattamento è impossibile a causa della ovvia assenza di evaporazione e della forte dispersione di calore. La temperatura interna di un poichiloterma acquatico è quindi sempre uguale a quella del mezzo.
2. Entrare in uno stato di vita latente (ibernazione o estivazione) quando la temperatura varia oltre i limiti tollerabili per l'organismo. Lo stato di ibernazione (più raramente di estivazione) è presente nella gran maggioranza dei Vertebrati poichilotermi terrestri dei climi temperati, mentre è praticamente assente nei Vertebrati acquatici con forse qualche rara eccezione. Ibernano anche numerosi Mammiferi ed alcuni Uccelli. Vedremo più avanti che lo stato di ibernazione, almeno nei Mammiferi, non è un semplice stato di torpore determinato da condizioni di temperatura sfavorevoli, ma necessita invece di complessi addattamenti fisiologici.
3. Cercare di mantenere relativamente costante la temperatura del corpo attraverso un continuo cambiamento di ambiente alla ricerca di quello più soleggiato o più ombroso in modo da rendere minime le variazioni.
4. Variare l'attività degli enzimi ed in generale delle proteine con l'attività fisiologica, modificando sia l'attività specifica sia la velocità massima delle reazioni e l'affinità per il substrato, in maniera da mantenere relativamente costante la loro attività in un certo ambito di temperatura. A variazioni al livello degli enzimi si possono affiancare modificazioni al livello dei tessuti o degli organi.
È evidente che alcuni o anche tutti questi meccanismi di adattamento possono essere contemporaneamente presenti. È anche possibile, e teoricamente probabile, che siano tutti assenti in casi particolari. Per esempio un pesce abissale che vive ad una temperatura costante intorno a 2-4 °C non ha alcuna necessità di avere un meccanismo fisiologico sviluppato di adattamento alla temperatura.
Cominciamo con l'esaminare quest'ultima possibilità, che è anche una delle più interessanti, e consideriamo sia come si può vedere teoricamente il problema, sia che cosa in pratica si è trovato. È però bene premettere che tuttora, data la enorme complessità dei fenomeni in gioco, si è ben lungi dall'avere un'idea completa di ciò che avviene e di come avviene.
La prima difficoltà che si trova ad affrontare immediatamente un vertebrato poichilotermo al variare della temperatura è una difficoltà respiratoria. Questa deriva da tre ordini di fatti.
1. Se l'animale è acquatico, dal fatto che la solubità dell'ossigeno nell'acqua diminuisce fortemente con l'aumentare della temperatura.
2. Dal fatto che il trasportatore di ossigeno, l'emoglobina, varia in linea generale la sua affinità per l'ossigeno in ragione inversa al variare della temperatura, nel senso che un aumento di questa porta ad una diminuzione dell'affinità per l'ossigeno. È chiaro che una variazione troppo forte nell'affinità, se non è in qualche modo compensata, è incompatibile con la vita: infatti un suo aumento porta ad una facile ossigenazione del sangue al livello dei polmoni o delle branchie, ma porta contemporaneamente all'impossibilità di liberare l'ossigeno al livello dei tessuti, perché, anche a basse pressioni parziali del gas, l'emoglobina riniane completamente saturata; al contrario, un aumento della temperatura con conseguente diminuzione dell'affinità porta ad un'inefficiente saturazione del sangue al livello dell'organo respiratorio e quindi ad analoghe difficoltà.
3. Infine, le difficoltà di ossigenazione divengono ancora maggiori quando aumenta la richiesta di ossigeno conseguentemente all'accresciuto metabolismo determinato dall'aumento di temperatura. In effetti nel pesce rosso (Carassius auratus), che pure può adattarsi e vivere per un tempo indefinito fra 0 e 40 °C, un brusco aumento di temperatura porta a chiari sintomi di asfissia. La prima reazione infatti è quella di aumentare il ritmo respiratorio nei limiti del possibile, e cioè la circolazione d'acqua al livello delle branchie, e di cercare alla superficie acqua più ossigenata. Pesci come il carassio che non hanno speciali adattamenti per poter respirare ossigeno gassoso possono tuttavia, in queste condizioni, giungere al tentativo di prendere delle boccate d'aria per aumentare l'ossigenazione. Ad ogni modo se il cambiamento non è troppo brusco il pesce sopravvive, ed in capo a pochi giorni i sintomi asfittici scompaiono. Il problema si pone allora in questi termini: cosa succede al livello del trasportatore di ossigeno per poter fronteggiare cambiamenti respiratori di questa entità?
Il tipo e l'entità dell'adattamento variano certamente a seconda delle abitudini ecologiche dell'animale. Gli animali ibernanti hanno verosimilmente scarsissima capacità di adattare la curva che misura l'affinità dell'emoglobina per l'ossigeno: anzi si sa che il tipo di curve è tale da ridurre al minimo la cessione di ossigeno ai tessuti alle basse temperature il che è probabilmente vantaggioso per l'animale provocando un abbassamento del metabolismo nel periodo di letargo (v. Morpurgo e Leggio, 1968). Necessariamente diverso deve essere il comportamento dell'emoglobina in un animale che sia capace di vita attiva in un vasto ambito di temperature.
Studi condotti su tritoni (Triton cristatus) e carassi hanno dimostrato in effetti (v. Morpurgo e altri, Negative Bohr effect..., 1970) che durante l'adattamento varia profondamente l'affinità per l'ossigeno alle varie temperature e ai vari pH. Se consideriamo in maniera più approfondita la curva di dissociazione dell'Hb vediamo che per la respirazione uno dei fattori essenziali è dato dalla posizione della curva in relazione al pH del sangue o meglio alla tensione di CO2. L'affinità dell'emoglobina per l'ossigeno varia infatti (effetto Bohr) al variare del pH (v. Antonini e Brunori, 1971), e in tutti i Vertebrati - con l'unica eccezione degli Urodeli -, nell'ambito di pH fisiologico, l'affinità diminuisce con il diminuire di questo. Questo fatto ha per la respirazione una importanza enorme, perché, al livello dei tessuti il pH del sangue, a causa della liberazione di CO2, è più basso e quindi la diminuita affinità per il trasportatore facilita la cessione di O2 ai tessuti. L'organismo avrebbe quindi un gran vantaggio a regolare le curve di dissociazione in maniera tale da consentire sempre, in seguito alla variazione del pH e della temperatura, la massima ossigenazione al livello dei polmoni o delle branchie e la massima cessione di O2 ai tessuti. È appunto quanto accade: nel pesce rosso, per esempio, l'acclimatazione per un mese a 4 e 37 °C porta ad un profondo spostamento della curva di dissociazione dell'emoglobina e dell'effetto Bohr, con un significato chiaramente adattativo. Si ha infatti, con l'aumentare della temperatura, un aumento dell'effetto Bohr e contemporaneamente uno spostamento verso i pH più acidi. Negli animali a sangue freddo, in effetti, il pH del sangue non è costante e si sposta verso i valori acidi durante l'adattamento alla temperatura alta, forse per l'aumentata liberazione di CO2. Lo spostamento che si è trovato nelle curve di dissociazione ha quindi un chiaro significato fisiologico.
Si pone adesso il problema fondamentale di sapere come possa cambiare nell'organismo l'affinità dell'emoglobina per l'ossigeno. Teoricamente esistono due possibilità: a) sostituire la molecola dell'emoglobina con un'altra a struttura primaria differente, il che comporterebbe l'esistenza e l'attivazione di un gene; b) cambiare la curva di dissociazione attraverso interazioni di tipo allosterico.
Negli animali a sangue freddo e più in generale nei normali fenomeni adattativi dei Vertebrati adulti è risultata vera la seconda ipotesi. La posizione della curva in relazione alla temperatura e al pH è determinata dalla presenza di cofattori la cui concentrazione intracellulare può variare rapidamente a seconda delle condizioni ambientali. Per il momento si sa che questa funzione di tipo allosterico è determinata nei Vertebrati dai fosfati organici (2-3 difosfoglicerato nei Mammiferi, inosinesafosfato nelle tartarughe e negli Uccelli, ATP in Pesci e Rettili; v. Benesch e altri, 1968) ma è molto probabile che esistano anche altri cofattori non ancora scoperti. Naturalmente una regolazione di questo tipo comporta l'esistenza di un sistema di regolazione della sintesi dei fosfati organici, sistema che deve essere direttamente sensibile o alla concentrazione dell'ossigeno, o alla percentuale di emoglobina ridotta all'interno del globulo rosso.
È però interessante notare che in un particolare caso, in cui è necessario un profondo cambiamento fisiologico, avviene esattamente il contrario, vale a dire il necessario spostamento nella curva di dissociazione si realizza attraverso un cambiamento nella molecola dell'emoglobina e quindi attraverso l'attivazione di un nuovo gene e la conseguente inattivazione di un altro: questo cambiamento si riscontra in forme molto diverse tra loro, come gli Anfibi e i Mammiferi, in corrispondenza del passaggio dalla vita embrionale a quella adulta, e quindi del momento della metamorfosi, negli Anfibi, e della nascita, nei Mammiferi. In tutti e due i casi si ha la sintesi ex novo di un nuovo tipo di emoglobina. Nell'uomo infatti l'emoglobina adulta è formata da un tetramero composto da due catene α e due β mentre l'emoglobina fetale è formata da due catene α e due γ. Ci si può chiedere perché in questo caso l'organismo ricorra alla sintesi di una nuova proteina mentre nel normale adattamento fisiologico si ricorre alla sintesi di un regolatore allosterico. Probabilmente la scelta è di tipo puramente economico: il passaggio dalla vita embrionale a quella adulta avviene una sola volta e avviene sempre, e pertanto può essere conveniente pianificarlo nel genoma dell'organismo. Il cambiamento necessario per l'adattamento fisiologico dell'organismo è invece saltuario e imprevedibile e un procedimento che si basi sulla sintesi e distruzione rapida di proteine sarebbe impossibile negli Eucarioti, dove i processi di controllo della sintesi proteica sono, come abbiamo visto, molto ridotti.
È anche interessante notare a questo proposito che l'emoglobina fetale e quella adulta purificate hanno la stessa affinità per l'O2, ma che le due emoglobine differiscono nella loro affinità per i fosfati organici. Al momento di iniziare la vita adulta, e cioè al momento in cui l'organismo da un ambiente costante entra in uno variabile, l'emoglobina fetale viene sostituita con un'altra capace di reagire alle variazioni ambientali in maniera adattativa.
Nei Mammiferi e negli Uccelli fenomeni del genere di quelli che abbiamo descritto sono certamente molto più limitati, perché questi due gruppi nell'evoluzione hanno raggiunto una costanza quasi assoluta della temperatura e del pH del sangue. Pur avendo importanza più limitata, il sistema di adattamento è - almeno nei Mammiferi - ancora in funzione, e permette un limitato adeguamento della curva alle necessità nei casi in cui si abbia deficit di ossigeno, per esempio in alta montagna o in caso di malattie cardiache. L'adattamento dell'uomo alla vita alle grandi altezze è stato abbastanza ben studiato nel corso di spedizioni alpinistiche. Ne facciamo qui soltanto un breve accenno, perché esso segue nelle sue linee essenziali le modalità di adattamento ai cambiamenti di temperatura negli animali poichilotermi.
Per quanto ancora molto rimanga da comprendere, alcuni fenomeni essenziali sono abbastanza chiari. La prima reazione consiste ovviamente nell'aumentare il ritmo respiratorio per cercare di compensare il deficit di ossigeno. Successivamente intervengono importanti modificazioni al livello del sangue: fondamentalmente aumenta il valore dell'ematocrito e conseguentemente dell'emoglobina e diminuisce il rapporto fra plasma e cellule circolanti. L'aumento delle cellule circolanti è a sua volta determinato da un aumento nel livello di eritropoietina, un ormone che stimola la proliferazione delle cellule della serie rossa. Inoltre, con un meccanismo analogo a quello che abbiamo descritto nei Pesci, a causa di un aumento nella concentrazione di 2-3 difosfoglicerato diminuisce l'affinità per l'ossigeno dell'emoglobina facilitando il rilascio del gas ai tessuti. Forse è anche possibile nell'uomo un aumento adattativo nell'effetto Bohr (v. Morpurgo e altri, Higher Bohr effect..., 1970). Inoltre il soggiorno prolungato a grande altezza provoca un aumento nel volume alveolare totale e, per facilitare il rilascio dell'ossigeno ai tessuti, aumenta anche la capillarizzazione al livello dei muscoli. Nell'uomo quindi, anche se su scala più limitata, in seguito ad ipossia si verificano gli stessi fenomeni che si producono negli animali a sangue freddo in seguito a un cambiamento di temperatura. Molto interessanti sono probabilmente anche i cambiamenti che si verificano nei Mammiferi ibernanti durante la fase di letargo, ma per il momento non si hanno dati al riguardo.
Sarebbe per esempio interessante sapere se in questi Mammiferi, come avviene nel caso dei poichilotermi ibernanti, l'effetto Bohr si annulla alle basse temperature per rendere più difficile il rilascio dell'ossigeno ai tessuti ed abbassare il metabolismo. È difficile fare delle previsioni al riguardo, perché nei Mammiferi il letargo è un fenomeno certamente molto più complesso di quanto non lo sia negli animali poichilotermi. Nei Mammiferi infatti la capacità di termoregolare nello stato di ibernazione non è affatto perduta, ma semplicemente il centro nervoso che controlla la temperatura del corpo è spostato su valori più bassi; la temperatura del corpo non scende sotto i 4 °C anche se quella esterna si abbassa notevolmente al di sotto di questo valore.
Come si vede, per quanto i dati siano ancora molto lacunosi, si ha ormai un'idea di quanto può avvenire, in seguito al cambiamento di temperatura, al livello respiratorio. Ma col cambiare della temperatura le difficoltà di un animale non si limitano certamente a quelle respiratorie. Gli stessi problemi che si hanno per l'emoglobina si ripetono per ogni enzima: è infatti ben noto che l'attività di questi varia con la temperatura, riducendosi in generale alla metà per una diminuzione di 10 °C. È chiaro che anche in questo caso deve necessariamente esistere qualche sistema di adattamento, ma gli studi al riguardo, tecnicamente molto più difficili di quanto non siano stati quelli condotti sull'emoglobina, sono solo all'inizio. Vediamo comunque anche qui quali sono le possibilità teoriche, possibilità che elencheremo di seguito e che non sono spesso mutualmente esclusive.
1. Regolazione o meglio adattamento dell'attività enzimatica attraverso meccanismi di regolazione allosterica che modifichino la velocità massima dell'enzima e l'affinità dell'enzima per il substrato (Km) o tutti e due questi parametri. Le modificazioni dell'attività enzimatica potrebbero anche non essere di natura allosterica, ma mediate da un cambianiento di pH intracellulare che, come abbiamo visto, nei poichilotermi non è fisso.
2. Adattamento attraverso una modificazione del numero di molecole di enzima presenti: questo cambiamento potrebbe avvenire o variando la concentrazione intracellulare degli enzimi, o più semplicemente modificando le dimensioni o il numero di cellule di un particolare organo.
3. Sostituzione di una particolare specie enzimatica con un'altra specie, meglio adattata alle nuove condizioni, per attivazione di un gene quiescente. In casi particolari, e cioè quando gli enzimi sono eteropolimerici, potrebbe variare la percentuale dei due diversi monomeri nell'enzima ed in questo caso non vi sarebbe bisogno di attivare un nuovo gene.
Solo da pochi anni si è cercato di comprendere che cosa realmente accada. Soprattutto per merito di P. W. Hochachka si è visto che esiste certamente qualche meccanismo di adattamento al livello enzimatico, ma si è ancora ben lungi dal poter formulare delle conclusioni generalmente valide. Nel pesce rosso (v. Hochachka, 1965) sembra che per quanto riguarda la latticodeidrogenasi sia valida l'ultima ipotesi. La latticodeidrogenasi può essere formata da non meno di cinque differenti monomeri. La variazione percentuale dei vari monomeri in seno ad una determinata molecola ne modifica le proprietà elettroforetiche e l'attività. Nel carassio la composizione e l'intensità delle diverse bande elettroforetiche varia a seconda dell'adattamento alla temperatura e si può pensare che questa variazione abbia un significato adattativo.
Nella trota, Baldwin e Hochachka (v., 1970) hanno trovato che l'acclimatazione a 2 e 17 °C porta alla formazione di forme elettroforeticamente distinguibili di acetilcolinesterasi. Pesci acclimatati ad una temperatura intermedia presentano contemporaneamente le due forme di enzimi. La velocità massima delle diverse forme di enzima in relazione alla temperatura non sembra cambiata mentre varia il Km che presenta un minimo alla temperatura di adattamento. L'aumento di affinità dell'enzima per il substrato, tuttavia, pur migliorando la situazione dell'animale non sembra che possa spiegare compiutamente i fenomeni di adattamento. Dati dello stesso genere, per quanto meno chiari, sono stati ottenuti, sempre da Hochachka, anche sulla isocitrico deidrogenasi di trota. Per quanto interessanti questi dati non permettono per ora di rispondere ad uno dei quesiti fondamentali che ci eravamo posti. La comparsa di forme elettroforeticamente diverse di un enzima in seguito ad adattamento non ci permette infatti di dire se si tratta di un enzima ‛nuovo' con struttura primaria diversa da quella precedente, che deriva dall'attivazione di un nuovo gene, o se si tratta di una diversa forma conformazionale dello stesso enzima. Ma, ripetiamo, gli studi in questo campo sono ancora ad uno stato embrionale e non permettono conclusioni generalmente valide.
Infine si deve fare una considerazione assai importante: quando si parla di adattamento si usa molto spesso il termine per indicare che l'animale sopravvive ed è attivo nelle nuove condizioni migliorando in qualche modo col tempo il suo stato. Manca però uno studio completo che ci dica quanto si è adattato, quali sono state le sue variazioni metaboliche e funzionali, quali le variazioni in peso e composizione degli organi (o nel sangue le variazioni nel numero delle cellule circolanti), quali le variazioni di un certo numero di enzimi, quali infine le variazioni nelle concentrazioni dei loro substrati all'interno delle cellule. Solo quando tutti questi dati saranno disponibili in almeno una specie animale - e si tratta di una impresa ardua - sarà possibile tracciare un quadro completo dei fenomeni di adattamento alla temperatura e vedere quale sia il peso di un determinato fenomeno nel quadro complessivo. È mia impressione comunque, sulla base di considerazioni evolutive e sulla base degli scarsi dati a disposizione (v. tab. II), che in nessun caso un animale a sangue freddo abbia trovato il modo di adattarsi in maniera rapida ed efficiente alle variazioni di temperatura. Altrimenti non avrebbe avuto evolutivamente senso l'affermarsi della termostatazione indipendentemente nei Mammiferi e negli Uccelli, processo che rende costanti le funzioni dell'organismo solo a prezzo di un dispendio energetico enorme.

8. Adattamento fisiologico e sistema nervoso
In uno sguardo evolutivo, sia pure sommario, ai fenomeni adattativi non si può tralasciare di parlare dell'intelligenza. Si è visto finora che l'evoluzione, anche se in maniera non sempre perfettamente lineare, ha teso in linea generale a costruire organismi in cui le variazioni dovute ad un programma interno hanno sempre maggior importanza rispetto alle variazioni strettamente dipendenti dall'ambiente. Naturalmente rimane sempre una certa possibilità di variazione fisiologica, ma si ha l'impressione che questa vada in media diminuendo nei gruppi più evoluti, anche se esistono notevoli eccezioni a questa regola. Per esempio un'eccezione di estrema importanza, di cui però non possiamo parlare qui, data la complessità del problema (v. immunologia e immunopatologia), è data dal funzionamento del sistema immunitario dei Vertebrati che presenta un ambito di adattamento fisiologico che apparentemente non ha limiti.
In uno di questi gruppi più evoluti, e cioè nei Vertebrati e massimamente nell'uomo, esistono però un organo ed una funzione la cui risposta è strettamente dipendente dall'ambiente in maniera adattativa: si parla del cervello e quindi dell'intelligenza. Non voglio qui parlare dei possibili meccanismi fisiologici da cui può dipendere il funzionamento del cervello e quindi lo sviluppo dell'intelligenza e della memoria, ma solo far notare che l'intelligenza, nella serie evolutiva che ha portato all'uomo, è un fattore estremamente influenzabile dall'ambiente. Quanto alla memoria il concetto stesso di questa non è scindibile, separabile da quello dell'ambiente e delle sue variazioni. È comunque innegabile che in un uomo il livello di intelligenza (comunque lo si voglia definire) dipende in maniera fondamentale, oltre che da fattori genetici, dall'educazione e quindi dalle esperienze acquisite. L'assenza di educazione, caso che si è saltuariamente verificato nei cosiddetti ‛bambini selvaggi' - cioè in quei bambini che sono stati da piccoli rapiti da animali selvaggi e da questi allevati -, può determinare uno stato di idiozia pressoché totale anche in soggetti presumibilmente normali alla nascita. Sembra anche che alcune cognizioni, come per esempio l'apprendimento del linguaggio, non solo siano strettamente dipendenti dall'educazione, ma debbano anche essere assimilate prima di una certa età.
Senza arrivare a casi così estremi è ormai chiaro e comunemente accettato che il livello di intelligenza, sia nel singolo individuo, sia in intere popolazioni, è influenzato in maniera determinante dal tipo di educazione, intendendo con tale termine l'insieme di esperienze cui l'individuo è sottoposto nell'arco della sua vita. Ciò è valido per gli animali superiori come i Mammiferi, molti dei quali infatti insegnano alla prole attraverso il ‛gioco' il comportamento giusto per una determinata circostanza, ma è certamente nell'uomo che tutto questo è massimamente evidente. Avendo allora l'intelligenza un valore adattativo, si può forse dire che il più vistoso fenomeno di adattamento nella specie umana è dato proprio dallo sviluppo dell'intelligenza in seguito ad appropriati stimoli ambientali. Questo fatto ha certamente delle implicazioni interessanti anche per quanto riguarda gli altri fenomeni di adattamento della stessa specie umana. Se consideriamo per esempio la distribuzione attuale dell'uomo sul nostro pianeta, notiamo che esso è, senza alcun dubbio, l'animale più ubiquitario: non esiste praticamente ambiente che sia così inospitale da rendere impossibile l'insediamento umano. Così l'uomo da tempo vive sia in regioni completamente e permanentemente coperte dai ghiacci sia nei climi equatoriali. Non si può dire che viva nelle regioni desertiche per la semplice ragione che quando l'uomo vi si stabilisce queste cessano di essere desertiche e vi è quindi ‟la contraddizion che nol consente". Questa dispersione dell'uomo è dovuta solo in misura molto secondaria ad una differenziazione genetica fra le varie razze che permetta all'uomo di vivere in climi così diversi. Per quanto riguarda la resistenza al freddo moltissimi animali posseggono sistemi di adattamento fisiologico molto migliori di quelli dell'uomo: l'uomo sotto tutte le latitudini è per sua natura praticamente nudo mentre gli animali sono coperti di una pelliccia il cui spessore è per giunta variabile a seconda delle variazioni di temperatura: così il gatto - tanto per fare un esempio a tutti familiare - ha una pelliccia variabilissima nel suo spessore a seconda della temperatura esterna.
Nulla di simile accade nell'uomo che riesce a sopravvivere solo modificando l'ambiente stesso, costruendosi rifugi, accendendo il fuoco, coprendosi di vestiti ecc. Chiaramente, quindi, per l'uomo, le sue capacità di modificare l'ambiente sono risultate tanto vantaggiose da rendere in molti casi superflui i normali meccanismi di adattamento fisiologico, che sono, di conseguenza, in parte andati perduti nel corso dell'evoluzione. E d'altro lato non si tratta di meccanismi limitati all'uomo: nelle lucertole, ad esempio, il principale meccanismo di adattamento alla temperatura consiste semplicemente nel cercare continuamente le aree con la temperatura ottimale, è quindi un meccanismo dipendente dalla ricerca attiva e cioè dall'intelligenza. Questo tipo di adattamento è enormemente più sviluppato nell'uomo il quale ha in questo modo imparato a supplire a molte delle sue carenze fisiologiche. Per la verità il problema è assai più complesso; l'adattamento attraverso l'intelligenza è infatti al tempo stesso causa ed effetto delle carenze di adattamento fisiologico: è molto probabile che in futuro le capacità adattative della specie umana non dipendenti dall'intelligenza andranno ancora diminuendo non avendo più per la specie un valore selettivo positivo. Possiamo quindi concludere che nella linea evolutiva che ha portato alla formazione dei Vertebrati e quindi dell'uomo il sistema di gran lunga più fisiologicamente adattabile (nel senso di dare una risposta adattativa agli stimoli ambientali) è quello nervoso, il cui tipo di sviluppo determina in parte il livello di adattabilità anche degli altri sistemi.
Per finire vogliamo tuttavia far notare che quanto è successo nei Vertebrati e soprattutto nell'uomo non costituisce necessariamente l'unico sviluppo possibile in gruppi in cui le capacità nervose siano molto importanti. Negli Insetti nel corso dell'evoluzione è accaduto infatti esattamente l'opposto: l'attività nervosa è determinata in maniera molto più rigida attraverso complicatissimi istinti scarsamente modificabili dall'esperienza e certamente non determinati da questa. L'Insetto quindi nascendo ‛sa', senza che alcuno glielo abbia insegnato, ciò che deve fare anche se si tratta di operazioni estremamente complicate. Per esempio una particolare vespa (Sphegidae) sa alla nascita quale bruco deve pungere e dove iniettargli il suo veleno perché l'animale non muoia, ma rimanga paralizzato in modo da poter servire come terreno di crescita non deperibile per le sue uova deposte all'interno del bruco stesso. Si tratta di un istinto fra i più semplici dei moltissimi descritti negli Insetti che in nulla dipende dall'esperienza, perché la vespa non ha mai conosciuto sua madre né altre vespe che compiano la stessa operazione. Sembra quindi che negli Insetti l'ambito di variazione fisiologica, per quanto riguarda le caratteristiche del sistema nervoso, sia molto piccolo e si sia quindi raggiunta una fissità molto maggiore (almeno per quanto riguarda questo carattere, ma forse anche per altri) di quanto non si sia verificato in altri gruppi. Sebbene sia pericoloso fare affermazioni troppo drastiche, si ha l'impressione che in questo gruppo l'importanza dell'adattamento fisiologico tenda a divenire sempre minore: l'organismo è scarsamente plastico nei riguardi dell'ambiente e l'adattamento avviene allora fondamentalmente a livello della popolazione per selezione ed adattamento genetico.
bibliografia
Ames, B., Goldberger, R., Hartman, P., Martin, R., Roth, J., The histidine operon in regulation of nucleic acid and protein biosynthesis, Amsterdam 1967.
Antonini, E., Brunori, M., Hemoglobin and myoglobin in their reactions with ligands, Amsterdam 1971.
Baldwin, J., Hochachka, P. W., Functional significance of isoenzymes in thermal acclimatization, in ‟Biochemical journal", 1970, CXIV, pp. 883-887.
Beckwith, J. R., Regulation of the lac operon, in ‟Science", 1967, CLVI, pp. 597-604.
Benesch, E., Benesch, R. E., Yu, C. I., Reciprocal binding of oxygen and diphosphoglycerate by human hemoglobin, in ‟Proceedings of the National Academy of Sciences", 1968, LIX, pp. 526-532.
Brock, T. D., Biology of microorganisms, Englewood Cliffs, N. J., 1970.
Demerec, M., Clustering of functionally related genes in Salmonella typhimurium, in ‟Proceedings of the National Academy of Sciences", 1964, LI, pp. 1057-1060.
Ekberg, D. R., Respiration in tissues of goldfish adapted to high and low temperatures, in ‟Biological bulletin", 1958, CXIV, pp. 308-316.
Gilbert, W., Muller-Hill, B., Isolation of the lac repressor, in ‟Proceedings of the National Academy of Sciences", 1966, LVI, pp. 1891-1898.
Hochachka, P. W., Isoenzymes in metabolic adaptation of a poikilotherm: subunit relationships in lactic dehydrogenases of goldfish, in ‟Archives of biochemistry and biophysics", 1965, CXI, pp. 96-103.
Jacob, F., Monod, J., On the regulation of gene activity, in ‟Coldspring Harbor symposia on quantitative biology", 1961, XXVI, pp. 193-211.
Jost, J. P., Rickenberg, H. V., Cyclic AMP, in ‟Annual review of biochemistry", 1971, XL, pp. 741-774.
Kanungo, S. M., Prosser, C. M. L., Physiological and biochemical adaptation of goldfish to cold and warm temperatures. I. Standard and active oxygen consumptions of cold and warm-acclimated goldfish at various temperatures, in ‟Journal of cellular and comparative physiology", 1959, LIV, pp. 259-263.
Levy, H. M., Sharon, N., Koshland, D. E., Purified muscle proteins and the walking rate of ants, in ‟Proceedings of the National Academy of Sciences", 1959, XLV, pp. 785-791.
Monod, J., Changeux, J. P., Jacob, F., Allosteric proteins and cellular control systems, in ‟Journal of molecular biology", 1963, VI, pp. 306-329.
Morpurgo, G., Battaglia, P. A., Leggio, T., Negative Bohr effect in newt haemolysates and its regulation, in ‟Nature", 1970, CCXXV, pp. 76-77.
Morpurgo, G., Bernini, L., Battaglia, P. A., Paolucci, A. M., Modiano, G., Higher Bohr effect in Indian natives of Peruvian highlands as compared with Europeans, in ‟Nature", 1970, CCXXVII, pp. 387-388.
Morpurgo, G., Leggio, T., Dissociation curves of toad haemoglobin and a hypothesis on the cause of hibernation, in ‟Nature", 1968, CCXIX, pp. 493-494.
Nakami, H., Pious, D. A., Regulation of cytochrome oxydase in human cells, in ‟Nature", 1967, CCXVI, pp. 1087-1090.
Siminovič, D., Rheaume, B., Sachar, R., Seasonal increase in protoplasm and metabolic capacity in tree cells during adaptation to freezing, in Molecular mechanisms of temperature adaptation (a cura di L. Prosser), Washington 1967, pp. 3-40.
Tauro, P., Halvorson, H. O., Effect of gene position on the timing of enzyme synthesis in synchronous cultures of yeast, in ‟Journal of bacteriology", 1966, XCII, p. 652-661.
Yarwood, C. E., Adaptation of plant and plant pathogen to heat, in Molecular mechanisms of temperature adaptation (a cura di L. Prosser), Washington 1967, pp. 75-89.
Adattamento genetico di Richard Ch. Lewontin
sommario: 1. Introduzione. 2. Adattamento come stato. 3. I caratteri come adattamento. 4. La natura come progettatrice. 5. Selezione naturale e adattamento relativo. 6. L'adattamento nella genetica di popolazioni. 7. Adattamento ed estinzione. 8. Altri criteri di adattamento di popolazioni. 9. Adattabilità. □ Bibliografia.
1. Introduzione
Il concetto di adattamento è uno dei più problematici e confusi tra tutti quelli usati in biologia, e questa confusione ha origine in parte dal fatto che l'idea di adattamento è stata ereditata dalla biologia moderna da una lunghissima tradizione di pensiero sociale, economico e religioso. La nozione che tutti gli organismi, incluso l'uomo, sono costituiti meravigliosamente, perfettamente adatti al mondo, è molto antica. Il senso di meraviglia per lo squisito adattamento tra gli organismi e il loro ambiente, e particolarmente il progetto in apparenza perfetto di vari organi del corpo destinati a svolgere funzioni particolari, è stato a lungo considerato come la dimostrazione evidente della esistenza di un progettatore supremo. ‟I cieli narrano la gloria di Dio e il firmamento mostra l'opera delle sue mani", dice il Salmo biblico. Darwin era consapevole che la teoria della selezione naturale, per poter essere accettata, avrebbe dovuto lottare contro il formarsi di ‟organi di estrema perfezione e complicazione". ‟[...] Supporre che l'occhio, con tutti i suoi inimitabili congegni per mettere a fuoco distanze differenti, per lasciar passare differenti quantità di luce, e per correggere le aberrazioni sferiche e cromatiche possa essersi formato attraverso la selezione naturale, sembra, lo ammetto francamente, assurdo a tutti i livelli" (v. Darwin, 1859, p. 186).
Pur potendo spiegare l'origine delle specie, la teoria di Darwin non sarebbe stata soddisfacente se non avesse anche affrontato il problema dell'adattamento. ‟Considerando l'‛origine delle specie', è senza dubbio concepibile che un naturalista [...] sia potuto giungere alla conclusione che ogni specie [...] fosse derivata, come varietà, da altre specie. Tuttavia, tale conclusione, anche se ben fondata, non sarebbe soddisfacente fino a che non fosse possibile dimostrare come le innumerevoli specie che abitano questo mondo siano state modificate, così da ottenere quella perfezione di struttura e di adattamento reciproco che molto giustamente suscita la nostra ammirazione" (v. Darwin, 1859, p. 3). Così, per Darwin, l'adattamento rappresentava il problema centrale nella teoria dell'evoluzione e, in un certo senso, più essenziale di quello dell'origine delle specie. La teoria di Darwin della selezione naturale proponeva che le specie avessero avuto origine dal processo di adattamento.
2. Adattamento come stato
Mentre sembrava ovvio agli studiosi di storia naturale come anche ai teologi, ai darwiniani come agli anti-darwiniani che gli organismi fossero strettamente adattati al mondo e gli uni agli altri, serie difficoltà si presentarono ad un attento esame del concetto. In primo luogo, l'idea di adattamento implica sempre che vi sia una forma, intesa come condizione effettiva o come ideale preesistente, in virtù della quale un oggetto possa essere inserito in un tutto armonico: senza un ideale preesistente del genere non ci può essere ‛adattamento'. Così, una chiave che non entra in una serratura può esservi adattata con un'accurata limatura, o un apparecchio elettrico fabbricato per funzionare a un determinato voltaggio può essere reso idoneo mediante un trasformatore a funzionare ad un altro voltaggio. Per i prodotti del lavoro umano non è difficile stabilire la condizione preesistente alla quale un oggetto deve essere adattato. Similmente, quando si riteneva che il mondo naturale fosse la creazione preordinata di un artefice supremo, appariva ovvio che i pesci fossero stati fatti per nuotare nel mare e gli uccelli per volare nell'aria: cioè, che l'universo fisico fosse stato creato per primo e fosse stato successivamente popolato con organismi che fossero in armonia con esso.
Nella biologia moderna questo aspetto riappare nel concetto di ‛nicchia ecologica' come modo di prefabbricare un posto adatto alla vita, che può essere ‛occupato' o no da una specie esistente. Secondo questo punto di vista l'evoluzione è un processo che produce nuove specie adatte a nicchie ecologiche non occupate precedentemente. La selezione naturale è considerata un agente che adatta le nuove specie alla nicchia preesistente, come la chiave viene successivamente adattata alla serratura. Una difficoltà di questo approccio è che gli organismi in ogni momento della loro storia evolutiva devono occupare ‛qualche' nicchia ecologica, in modo che in ogni momento la specie sia adattata a qualche ambiente: se così non fosse, si estinguerebbe. Così, l'evoluzione potrebbe essere un processo che cambia questo adattamento di una specie da una nicchia prestabilita ad un'altra, o che scinde una specie, che era adattata a una nicchia, in numerose specie, ognuna con il suo nuovo adattamento; ma in nessun senso essa può essere considerata in grado di adattare una forma precedentemente non adattata. Un diverso modo di concepire la nicchia ecologica è quello per cui essa può essere definita in maniera appropriata solo da un organismo che la occupa, cosi che il concetto di ‛nicchia non occupata' è un controsenso. Secondo questo punto di vista, i modi in cui la mente umana può suddividere l'universo sono infinitamente numerosi ed è un puro pregiudizio scegliere alcuni di questi come nicchie ecologiche reali alle quali l'evoluzione possa adattare o meno alcune specie. Questa definizione esistenziale di nicchia ecologica rende privo di significato il concetto di adattamento dal momento che non c'è niente a cui adattare. Le specie cominciano ad esistere, cambiano, si estinguono: è tutto. Niente della loro storia è chiarito dalla nozione del loro stato di adattamento.
3. I caratteri come adattamento
Quando si passa dalla questione dell'adattamento come modo di essere delle specie al concetto di adattamenti specifici come meccanismi particolari, sorgono degli altri problemi. Ogni struttura particolare prenderà parte a molti processi differenti, i quali non possono essere considerati tutti come funzioni adattative. Così, il becco forte e aguzzo del pappagallo è generalmente considerato come un adattamento per la nutrizione, benché incidentalmente esso possa essere usato per infliggere dei morsi dolorosi. Neppure l'uso regolare di una struttura è sufficiente per capire la sua funzione adattativa. Le pinne frontali della tartaruga verde, Chelonia mydas, sono usate per far muovere l'animale sopra la sabbia asciutta verso un posto adatto per deporre le uova, ma nessuno che abbia assistito a questo laborioso spettacolo potrà considerare le pinne come adattamenti per la locomozione sopra un terreno asciutto. Nel caso di forme conosciute solo attraverso i fossili, ci sono delle reali ambiguità. La larga ‛pinna' dorsale del Pelicosauro era un adattamento per la regolazione del calore o per la manifestazione di aggressività o per l'attrazione sessuale? In realtà, il possesso della pinna potrebbe aver dato origine a tutte e tre le conseguenze per il Pelicosauro.
Alcune strutture o schemi di comportamento vengono talvolta considerati come adattamenti per particolari funzioni, se la sopravvivenza dell'organismo o della specie dipende dalla loro esistenza. I polmoni sono un adattamento per la respirazione nei grandi animali terrestri poiché, presumibilmente, nessun grande Mammifero terrestre avrebbe potuto sopravvivere senza di essi, e la causa della morte sarebbe stata il soffocamento. Ciò nonostante, un simile criterio è realmente troppo rigoroso, e molti adattamenti non supererebbero questo test.
In primo luogo non è affatto sicuro che la mancanza della maggior parte degli adattamenti sarebbe stata fatale. Le pinne sono un adattamento al nuoto nell'accezione corrente del termine, eppure i serpenti di mare e le anguille se la cavano benissimo senza di esse. Come ha fatto notare G. C. Williams, la pelliccia bianca dell'orso polare è un vantaggio per potersi nascondere nella neve, ma non si può affermare che la specie non sarebbe sopravvissuta se il suo mantello fosse stato scuro.
In secondo luogo, la presenza o l'assenza di un comportamento o di una struttura non sono gli unici stati possibili. Darwin scelse l'occhio dei Vertebrati come un esempio di ‟organi di estrema perfezione", eppure ammise che recettori luminosi meno complessi erano altrettanto adattabili per i loro possessori. In realtà la adattabilità di ogni forma intermedia di recettori luminosi in tutto il regno animale era un'asserzione cruciale per Darwin dato che, senza questa, non avrebbe potuto asserire che l'evoluzione dell'occhio dei Vertebrati era avvenuta con piccoli cambiamenti in tempi successivi. Il paradosso della posizione di Darwin era che, mentre ogni stadio dello sviluppo dell'occhio dei Vertebrati era un sufficiente adattamento per l'animale, durante l'evoluzione ci doveva essere stato anche un orientamento verso la ‛perfezione' dell'occhio. La soluzione darwiniana del paradosso si trova nel meccanismo della selezione naturale che operava per mezzo di differenze nell'adattamento relativo tra forme competitive.
4. La natura come progettatrice
In base al modo in cui Darwin usava questo concetto, e in base al modo in cui viene considerato dai moderni anatomisti, un adattamento non è una causa necessaria né sufficiente della selezione naturale. Quello che si origina nell'evoluzione può essere solo l'effetto indiretto di qualche adattamento primario, oppure un adattamento può non essere incorporato in una specie perché i suoi effetti collaterali potrebbero essere svantaggiosi. Dal momento che la conseguenza del processo evoluzionistico non definisce, di per se stessa, un adattamento, torniamo nuovamente alla natura a priori del concetto. Un carattere viene considerato un adattamento se un'analisi razionale mostra che esso è, in un certo senso, la soluzione. di un dato problema funzionale; più precisamente, si considera che un carattere sia più adatto di un altro se risponde maggiormente alle richieste di una macchina progettata per una particolare funzione. Così si dice che la forma delle spugne è un adattamento per la nutrizione, perché eleva al massimo grado il volume di acqua che può essere filtrata da una spugna di una data dimensione per ottenere microplancton (v. Leigh, 1971, cap. 2). Se la forma attuale delle spugne risultasse differente da quella ottimale, ma a questa molto vicina, sarebbe ancora considerata come un adattamento imperfetto; non è chiaro di quanto la forma delle spugne avrebbe dovuto essere lontana dall'adattabilità perché gli anatomo-ingegneri cercassero un'altra spiegazione per il loro aspetto. Inoltre, il successo di un tale tipo di analisi dipende dalle componenti dell'intera attività vitale dell'organismo prese come ‛problemi' da risolvere. Una spugna può essere la forma migliore per nutrirsi caricandosi di acqua, mentre diventa ben poco adatta a resistere ai predatori vaganti nell'ambiente. Come risultato, un anatomista risoluto può quasi sempre produrre una prova convincente che un carattere è un adattamento, specialmente dal momento che la conoscenza di tutti gli aspetti della storia della vita è di solito incompleta.
Il notevole successo di una simile spiegazione funzionale ha portato qualche evoluzionista ad ammettere che tutti i caratteri sono degli adattamenti, persino quelli per i quali non è stata ancora ideata una storia plausibile, o quelli che non si sono realmente evoluti. Persino uno scettico come Williams, reso estremamente cauto nell'applicazione di questo concetto dalla sua penetrante analisi dell'adattamento, fa questa ipotesi: ‟Trovo che l'assenza di viviparità tra gli Uccelli sia quanto mai misteriosa. L'enorme tasso di mortalità tra le uova in molte specie [...] suggerisce che un uccello potrebbe trarre un grande beneficio dalla viviparità, anche se la fecondità fosse molto ridotta in modo che il peso dei feti non interferisse seriamente con il volo. [...] Forse, gli Uccelli sono privi di un qualche importante preadattamento, che i primi Mammiferi possedevano, per superare gli ostacoli immunologici alla viviparità" (v. Williams, 1966, p. 170).
Questo argomento può essere considerato come modello adattativo: isola la viviparità come soluzione al problema della mortalità delle uova, adatta il meccanismo proposto in modo da tener conto di un effetto collaterale indesiderabile, l'interferenza con il volo, e infine suggerisce che la mancata evoluzione di questo carattere risieda nella intrinseca inflessibilità della fisiologia degli Uccelli. Ragionando su queste basi, diventano ovvie le radici intellettuali degli argomenti sull'adattamento.
Questa nozione è stata introdotta nel pensiero biologico derivandola dal concetto predarwiniano e preevoluzionistico di disegno e creazione speciale. È l'unica traccia della storia teistica delle idee sull'universo in uno schema per il resto meccanicistico e materialistico. C'è in questo caso un sorprendente parallelismo con le origini dell'idea della selezione naturale. Darwin iniziò il suo lavoro sull'origine delle specie con una discussione sulle variazioni dovute all'addomesticamento, e sul modo in cui la selezione artificiale degli allevatori dava come risultato la stabilizzazione di razze distinte. Per analogia, Darwin voleva stabilire che la variazione tra gli individui nelle popolazioni naturali poteva portare alla formazione di specie distinte, e per spiegare ciò egli postulava la natura come selettore, per quanto cieco e meccanico. Il vero termine ‛selezione naturale' porta con sé la sua storia intellettuale; lo stesso succede per ‛adattamento'. Mentre gli evoluzionisti moderni non credono in un Divino Ingegnere che risolve i problemi che egli stesso pone, l'organizzazione dei problemi evoluzionistici come problemi di adattamento naturale è l'eredità diretta di tale passato teologico. Mentre non ci sono problemi tecnici di dinamica dell'evoluzione e di formazione della specie che richiedano l'introduzione dell'idea di adattamento, la maggior parte degli evoluzionisti guarda all'adattamento come al problema centrale e non superficiale degli studi evoluzionistici (v. Stern, 1970).
5. Selezione naturale e adattamento relativo
La teoria darwiniana dell'evoluzione operata dalla selezione naturale si basa su tre proposizioni: a) c'è variabilità nella morfologia, fisiologia e comportamento tra gli individui nell'ambito di una specie (principio della variabilità); b) la variabilità è in parte ereditabile così che il figlio somigli più ai genitori che alla media della popolazione (principio dell'ereditarietà); c) forme differenti lasciano un numero diverso di figli nelle generazioni immediatamente successive o in quelle più lontane (principio della selezione naturale). Da queste tre proposizioni deriva che la specie evolverà in modo che le forme con un potenziale riproduttivo minore vengano eventualmente sostituite da quelle che hanno maggiori probabilità di sopravvivenza e di riproduzione. Come è stato detto prima, questi tre principi non dicono niente riguardo all'adattamento. Essi asseriscono soltanto che, se in una popolazione esiste una variabilità ereditaria e se, in media, alcune varianti danno una discendenza maggiore delle altre, la popolazione evolverà. Il concetto di adattamento appare solo in una quarta proposizione che non è essenziale per la dimostrazione di una evoluzione operata dalla selezione naturale. Questa quarta proposizione, la ‛lotta per l'esistenza', era intesa da Darwin come quella che forniva la causa fondamentale della selezione naturale. Rifacendosi a Malthus, Darwin asserì che tutte le specie producono una progenie più numerosa di quella che è in grado di sopravvivere, a causa delle limitate risorse di cibo e di spazio. Questi organismi combatteranno, allora, gli uni contro gli altri, per le risorse non sufficienti, e la vittoria in questa lotta andrà ordinariamente a quella forma la cui morfologia e la cui fisiologia saranno meccanicamente più adatte alla lotta. Il corridore più veloce, l'individuo con il metabolismo più efficiente, il più abile, il più forte dei due antagonisti riuscirà ad attingere maggiormente alle limitate risorse, oppure riuscirà ad avere una progenie maggiore, e verrà così selezionato.
Darwin allargava il concetto di lotta al di là della sola competizione tra organismi per delle risorse limitate. ‟Devo premettere che uso il termine ‛lotta per l'esistenza' in senso lato e metaforico [...]. Si può dire veramente che due cani, in un periodo di carestia, lottano l'uno contro l'altro per ottenere cibo e vita. Ma di una pianta al limite del deserto si dice che lotta per la vita contro la siccità [...]" (v. Darwin, 1859, p. 62). Estendendo la nozione di lotta fino ad includere la dipendenza dai fattori fisici dell'ambiente, Darwin ha liberato la teoria della selezione naturale dalla dipendenza dal meccanismo della sovrappopolazione, dato che la lotta di una pianta per avere l'acqua al limite di un deserto avverrebbe anche in assenza di altri individui. In ambedue i casi la vittoria nella lotta appartiene alla forma che meglio si è adattata, nel senso che la sua morfologia, la sua fisiologia e il suo comportamento danno come risultato diretto un livello di riproduttività più alto. Tutte le forme possono essere perfettamente adattate dal momento che una specie consistente in una certa forma ha potuto sopravvivere e anche accrescersi numericamente. L'adattamento relativamente migliore di una forma rispetto a un'altra è evidente solo quando ambedue esistono nella stessa popolazione di organismi competitivi. Così ciascuna forma è stabile dal punto di vista dell'evoluzione fino a che non è messa alla prova da una meglio adattata. In tal modo tutte le forme ‛meno perfette' di ricettori luminosi possono persistere in specie diverse, anche se vi è un movimento di evoluzione verso il ‛più perfetto' occhio dei Vertebrati.
È stato qualche volta asserito che il principio della selezione naturale è tautologico, perché afferma che la forma che più si è adattata sopravvive, e definisce quindi la forma che più si è adattata come quella che sopravvive meglio. Ma qui si confonde il principio della selezione naturale con quello della lotta per la sopravvivenza. Quest'ultima è considerata dalla teoria darwiniana come la causa meccanica della prima. In termini pratici questo significa che il maggior adattamento di una forma rispetto ad un'altra, è, almeno in principio, osservabile e misurabile indipendentemente da ogni misura di efficienza riproduttiva. Così, asserendo che una forma di organo fotorecettore è più adatta di un'altra, dovrebbe essere possibile, con la sola conoscenza della struttura e fisiologia dell'organo e della natura dell'ambiente in cui vive l'organismo, predire come queste possano esercitare la loro influenza sulla capacità riproduttiva degli organismi che ne sono provvisti. Cioè, il concetto di adattamento è distinto dal concetto di selezione naturale, perché è un concetto a priori, e insistendo su una meccanica predizione a priori si evita il circolo vizioso: ‛quello che è adattato sopravvive meglio e quello che sopravvive meglio è adattato'.
L'uso del concetto di adattamento per prevedere cambiamenti evoluzionistici richiede una conoscenza molto più completa di quella che oggi non si abbia della fisiologia e dello sviluppo di un organismo, e delle relazioni tra organismo e ambiente. È facile dire, per esempio, semplicemente partendo da principi meccanici elementari, che artigli lunghi renderebbero più adatto l'animale a scavare gallerie di quanto non farebbero delle unghie corte. Asserire, però, che i lunghi artigli sono selezionati per questo, è molto più problematico. Per prima cosa dobbiamo sapere se il fatto di scavare gallerie aumenta la provvista di cibo dell'animale, se ne diminuisce le probabilità di soccombere ai predatori, se cambia la sua posizione competitiva per l'accoppiamento, e così via. Cioè, data la attuale nicchia ecologica di un organismo, può un cambiamento, che è senz'altro un migliore adattamento ad alcuni modi di vita, portare ad una grande perdita di adattamento al suo attuale modo di vita? Se l'animale in questione fosse una talpa, potremmo predire che dei lunghi artigli sarebbero per essa un vantaggio; ma un coniglio, benché viva in tane sotterranee, perderebbe in mobilità più di quanto guadagnerebbe con un adattamento come scavatore. Mentre basta osservare per vedere che la talpa possiede lunghi artigli ed il coniglio no, prevedere la direzione della selezione naturale sulla base della lunghezza degli artigli non è facile. Una simile predizione richiederebbe una conoscenza completa dei fattori limitanti la crescita e la sopravvivenza nelle popolazioni delle talpe e dei conigli. Inoltre, bisogna prendere in considerazione le conseguenze metaboliche e dello sviluppo a seguito di un cambiamento di struttura.
Mentre un ingegnere idrodinamico può essere in grado di disegnare una pinna di forma ottimale per il nuoto di un pesce, cioè la forma di pinna più adatta, dei cambiamenti nella forma della pinna richiedono cambiamenti nella muscolatura, nell'innervazione, nella circolazione, nella regolazione dei processi di sviluppo e nei livelli metabolici. Le pinne sono aggredibili da predatori e da parassiti, possono servire come segnali di riconoscimento della specie e come organi che mostrino l'aggressività. Per prevedere se il cambiamento nella forma della pinna - prescindendo da quanto sia adattativo - verrà favorito dalla selezione naturale, sarebbe necessario avere una conoscenza della fisiologia e dello sviluppo degli organismi molto più vasta di quella disponibile oggi per ogni specie. E neanche serve predire cambiamenti adattativi, ‛rimanendo tutte le altre cose uguali', dato che sono proprio i molteplici effetti della stessa struttura cambiata che debbono essere presi in considerazione. Un cambiamento nella struttura della pinna non darà come unico effetto un cambiamento nella velocità di movimento nell'acqua.
L'uso dell'adattamento come spiegazione della selezione naturale non è invalidato dalla mancanza di una perfetta prevedibilità. Tutte le spiegazioni causali sono di questo tipo. Un dato fenomeno sarà la causa di un effetto solo quando tutte le condizioni correlate saranno soddisfatte. Si rimane sempre nell'ambito del normale significato di causalità dicendo che un attacco di cuore è stato la causa della morte di una persona, anche se gli attacchi di cuore non sono tutti invariabilmente mortali. Il risultato è soprattutto quello di attribuire una causa convincente ad un effetto osservato. È stato notato che una struttura complessa, l'occhio dei Vertebrati, si è evoluta. Gli evoluzionisti che ragionano in termini adattativi vorrebbero sostenere che l'occhio dei Vertebrati rende capace il suo possessore di vedere meglio, che una vista migliore porta ad una raccolta di cibo più fruttuosa, alla possibilità di evitare pericoli e predatori, e di trovare un compagno, in conclusione che tutte queste proprietà darebbero come risultato una maggiore produzione di prole rispetto alla media, e che è per questo che l'occhio dei Vertebrati si è sviluppato. Inoltre è perfettamente coerente con l'idea di adattamento parlare di adattamenti che non sono mai avvenuti. Così una valida domanda evoluzionistica è: ‟perché i leoni non possiedono un adattamento per attirare la preda?"; oppure ‟perché l'antilope non ha sviluppato un adattamento per combattere contro i leoni?"
Come il giudizio a priori della natura adattativa di un cambiamento potenziale non porta necessariamente a predire la direzione della selezione naturale, così l'osservazione che la selezione naturale ha trasformato un carattere non è una guida infallibile per le conseguenze adattative di esso. C'è stata nell'evoluzione umana una tendenza generale verso una diminuzione dello spessore delle ossa craniche e della lunghezza della mascella. L'entità di queste tendenze ci assicura che esse sono il risultato della selezione naturale, più che di un processo casuale e disordinato, eppure nessuno potrà sostenere che questi cambiamenti sono degli adattamenti primari. Piuttosto, si suppone che essi rappresentino gli effetti collaterali di cambiamenti di sviluppo nel cervello e nella muscolatura della testa e del collo, associati con le abitudini alimentari e con la posizione completamente eretta.
C'è una forma di adattamento generalizzato di cui la riduzione di spessore delle ossa craniche nell'uomo può essere un esempio. Il cranio spesso nell'antenato dell'uomo era quasi sicuramente un adattamento alla pesante mascella e alla muscolatura del collo degli ominidi primitivi. Quando la muscolatura incominciò a diventare più leggera, la selezione naturale per il mantenimento di una struttura ossea pesante si ridusse. Perché le ossa diventarono allora più leggere? Una supposizione ragionevole è che l'energia di sviluppo e metabolica richiesta per formare e mantenere un osso pesante era un costo di una certa importanza, così che la riduzione di spessore era un adattamento all'efficienza metabolica. In generale, la riduzione o la perdita di strutture che non sono più degli adattamenti diretti possono essere degli adattamenti ad una efficienza metabolica globale.
C'è una notevole possibilità che gran parte della selezione naturale, forse la maggior parte, non sia in nessun modo legata agli adattamenti in senso darwiniano. Studi recenti sulla variabilità degli enzimi e delle proteine in diverse specie hanno consentito di rilevare una enorme quantità di polimorfismi genetici (v. Lewontin e Hubby, 1966; v. Harris, 1970; v. Lewontin, 1973, cap. 3). Un certo numero di questi polimorfismi mostra una forte dipendenza geografica o ecologica nella frequenza delle varianti, spesso associata con una marcata dipendenza dalla temperatura, e gli esperimenti di laboratorio danno delle curve di attività in rapporto alla temperatura per le diverse varianti enzimatiche che concordano con le variazioni naturali. Le varianti enzimatiche che sono più comuni nelle regioni più fredde sono più attive alle temperature più basse, mentre quelle delle regioni calde sono più attive alle temperature più alte, oppure hanno una maggiore stabilità al calore. Una particolare variante enzimatica che è la più frequente in una popolazione è in relazione diretta con le proprietà molecolari di stabilità, di affinità per il substrato e di velocità di reazione. In base alle ricerche effettuate non è ancora chiaro quale relazione ci sia tra i processi morfogenetici e particolari varianti alternative di proteine negli organismi superiori, però è del tutto certo che una relazione esiste. Così, l'evoluzione di particolari strutture può essere più influenzata dalla stabilità al calore o dall'affinità per il substrato degli enzimi o delle altre proteine concernenti i processi morfogenetici, che da ogni altra proprietà delle strutture stesse. Un esempio è il famoso caso del melanismo industriale di Biston betularia, e di altre farfalle (v. Ford, 1964). Il grande aumento dell'inquinamento atmosferico dovuto alle industrie in Inghilterra sin dalla metà del XIX secolo è stato accompagnato da un rapido incremento delle forme melaniche di numerose specie di farfalle nelle aree industriali. I licheni grigio-verdi che crescono sui tronchi degli alberi sono particolarmente sensibili all'inquinamento dell'aria dovuto al carbone e sono praticamente scomparsi dalle regioni industriali dell'Inghilterra e, come risultato, le farfalle a colori chiari che si mimetizzavano riposandosi sui licheni, sono ora visibili agli uccelli predatori, mentre le forme melaniche sono invisibili sui tronchi scuri. È stato ampiamente dimostrato che gli uccelli predatori catturano in realtà più farfalle visibili che invisibili, e in tal modo è confermata la natura adattativa del colore. Ad una più attenta osservazione, comunque, risulta che anche le larve delle forme melaniche hanno un vantaggio selettivo nelle aree inquinate, benché le stesse larve non siano melaniche. È ancora un problema aperto sapere fino a che grado l'evoluzione del melanismo industriale è stato il risultato di un diretto adattamento all'attività predatoria e in quale misura è la conseguenza indiretta di differenze fisiologiche nella direzione biosintetica della produzione di melanina.
La probabilità che alcuni cambiamenti morfologici e di comportamento si evolvano come conseguenza secondaria di altre proprietà di sviluppo, fisiologiche o molecolari, porta anche al fenomeno del preadattamento. Caratteri che sono chiaramente degli adattamenti in senso funzionale, possono essersi evoluti originariamente come effetti indiretti della selezione naturale, diventando adattamenti solo dopo, quando la specie incontra un nuovo ambiente. E, cosa ancora più probabile, un adattamento ad un certo modo di vita può risultare casualmente un preadattamento per un altro. Un esempio classico è l'evoluzione degli ossicini dell'orecchio medio dei Mammiferi dagli elementi dell'articolazione della mascella nei Rettili. Come l'osso dentale della mascella inferiore e lo squamoso di quella superiore dei Rettili simili ai Mammiferi hanno acquisito la funzione di articolazione, così l'angolare, l'articolare e il quadrato si sono trasformati nel timpanico, martello e incudine dell'orecchio dei Mammiferi.
6. L'adattamento nella genetica di popolazioni
In contrasto con gli anatomisti, che studiano l'adattamento dal punto di vista di tecnici progettisti, la genetica evoluzionistica lo considera come un risultato a posteriori della selezione naturale. L'‛adattabilità', l'‛idoneità biologica', o il ‛valore adattativo' sono misure numeriche correlate con la natalità e la mortalità, e con il tasso di crescita e le probabilità di sopravvivenza di popolazioni e specie, e presentano solo una incerta e generalmente non verificata relazione con i particolari adattamenti di forma e funzione che compaiono nelle enunciazioni della morfologia e fisiologia comparate. Si supponga che vi siano due alleli ad un locus in un organismo diploide, e che i tre genotipi AA, Aa, e aa abbiano differenti valori di fecondità e probabilità di sopravvivenza. Siano WAA, WAa e Waa rispettivamente i numeri di uova fecondate di ciascun genotipo della generazione n + 1, rispetto alle uova fecondate di ciascun genotipo nella generazione n. Questi rapporti di riproducibilità netta per genotipo sono chiamati valori adattativi o idoneità biologiche dei genotipi. Sia inoltre W̅̅ l'adattamento medio, o valore adattativo medio della popolazione, così calcolato
W̅̅ =WAA fAA+WAa fAa+Waa faa
in cui fAA, fAa e faa sono le frequenze relative dei tre genotipi nella popolazione. Quindi, come conseguenza delle leggi della genetica mendeliana (v. anche genetica) e dei diversi valori di adattamento dei tre genotipi, la frequenza relativa dei due alleli cambierà nella popolazione secondo la legge (v. Wright, 1931)

dove Δq è la variazione della frequenza di uno degli alleli, ad esempio A, in una generazione. L'importanza di questa equazione sta nel fatto che il segno di Δq è uguale a quello di d ln W̅̅ /dq, il rapporto cioè fra il valore di idoneità biologica media e la frequenza allelica. La frequenza allelica, quindi, varierà in maniera da far aumentare il valore di W̅̅ . Questa legge, che non può essere rigorosamente esatta quando le idoneità biologiche dei tre genotipi sono funzioni delle loro frequenze, stabilisce una tendenza generale per un processo evolutivo: che cioè la selezione naturale fa aumentare l'adattamento medio di una popolazione. Alla medesima conclusione si perviene da una diversa prospettiva. La variazione totale nel valore di adattamento fra i tre genotipi nella popolazione può essere suddivisa in un effetto medio dovuto alla sostituzione di un allele A con un allele a (componente lineare) ed in una deviazione dovuta alla dominanza (componente quadratica). Se VlnW è la variazione del logaritmo dell'idoneità biologica associata all'effetto additivo, allora si avrà (v. Fisher, 1930)
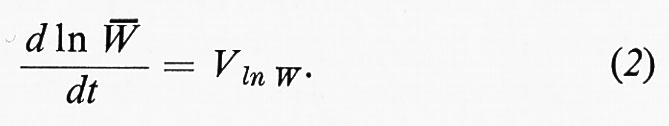
Questo ‛teorema fondamentale della selezione naturale' stabilisce che il valore del cambiamento dell'idoneità biologica media è uguale alla variazione genetica dell'adattamento (in scala logaritmica) ed è quindi necessariamente positivo. Inoltre, l'idoneità biologica media aumenta per effetto della selezione naturale. La scoperta di Wright e Fisher di queste due equazioni fornì un notevole contributo al principio che l'evoluzione fa aumentare l'adattamento. Vi sono tuttavia molti problemi impliciti in questa generalizzazione. In primo luogo, le equazioni (1) e (2) non sono del tutto corrette in alcuni casi di selezione in cui gli adattamenti genotipici non sono costanti, ed in alcuni casi in cui molti geni differenti sono sottoposti a selezione ed interagiscono fra loro nella determinazione dell'idoneità biologica. Questa obiezione è tuttavia meno grave, paragonata al problema dell'interpretazione del significato biologico di W̅̅ , l'adattamento medio. Sebbene i valori di W siano espressi come numero assoluto di uova di ciascun genotipo per uovo fecondato nella precedente generazione, le espressioni (1) e (2) mantengono la loro validità anche se i valori di adattamento vengono valutati rispetto ad uno dei genotipi, ad esempio AA, a cui si assegna un valore di idoneità biologica 1,0. D'altra parte, le espressioni (1) e (2) possono non essere esatte per i valori assoluti di idoneità biologica, poiché il valore assoluto del numero totale di uova fecondate prodotte da una popolazione per uovo fecondato nella precedente generazione è determinato in ultima analisi dalla capacità che esse hanno di superare la selezione ambientale. Per una popolazione che non si espande in ogni generazione - e la maggior parte delle popolazioni, mediamente rispetto a lunghi periodi, non si espande né diminuisce di numero - il valore assoluto di W̅̅ non può aumentare e deve essere sempre prossimo all'unità. Così, benché l'idoneità biologica media relativa di una popolazione possa aumentare per cambiamenti nella frequenza allelica, il valore di idoneità biologica media assoluta deve essere sempre vicino all'unità, se la popolazione non si sta estinguendo o espandendo su tutta la terra. Si consideri una popolazione stazionaria, formata unicamente da individui AA. Il loro valore di idoneità biologica assoluta sarà WAA=1,0, poiché le dimensioni della popolazione sono stazionarie. Si supponga, ora, che insorga una mutazione a che raddoppi la fecondità dei suoi portatori, sia omozigoti aa sia eterozigoti Aa. Se l'unico effetto dell'allele a è quello di cambiare la quantità di uova deposte, ma non l'efficienza di utilizzazione delle risorse, allora le dimensioni della popolazione totale non possono aumentare, e la deposizione di uova extra da parte di individui Aa e aa comporterà un maggior numero di morti degli embrioni e degli adulti, cosicché il valore di W̅̅ rimane 1. Se sono rari gli individui Aa e aa, i loro valori di idoneità biologica assoluta saranno vicini a 2 ed il valore di idoneità biologica assoluta degli individui AA sarà vicino ad 1, ma quando il genotipo più fertile aumenta la sua frequenza, i loro valori di idoneità biologica assoluta diminuiranno verso 1, mentre il valore di idoneità biologica assoluta di AA scende verso 0,5 in maniera tale che sia sempre W̅̅ =1. Proporzionalmente, i valori di idoneità biologica dei genotipi sono sempre 1,1 e 0,5 ed il valore di idoneità biologica media relativa aumenta lentamente da 0,5 a 1,0 a mano a mano che il nuovo gene si diffonde.
Se il nuovo mutante aumenta l'efficacia di utilizzazione delle risorse, allora cominceranno a crescere le dimensioni della popolazione, cosicché il valore assoluto W̅̅ sarà maggiore dell'unità. Quando aumenta la frequenza del nuovo genotipo, aumenterà anche W̅̅ , ma, ad una frequenza intermedia del nuovo tipo, la crescita della popolazione comincerà a diminuire allorché la popolazione si avvicina alla sua nuova capacità di adattamento, cosicché il valore assoluto di W̅̅ diminuirà nuovamente e tenderà all'unità. Alla fine del processo di sostituzione genica, la popolazione può avere raddoppiato le sue dimensioni, ma il valore assoluto di W̅̅ è nuovamente uguale ad uno, poiché la popolazione non è più in espansione.
In tal modo, che un genotipo fissato dalla selezione naturale influisca o meno sull'efficienza di utilizzazione delle risorse, il risultato finale del processo evolutivo è un valore di idoneità biologica medio assoluto identico a quello che si aveva prima che iniziasse il processo selettivo. L'aumento dell'idoneità biologica prevista dalle equazioni (1) e (2) è solamente un aumento dell'idoneità biologica relativa. Mentre il genotipo più prolifico aumenterà la sua frequenza relativa nella popolazione, non vi è nessuna legge generale che ci permetta di prevedere le conseguenze per la popolazione nel suo insieme. La ‛lotta per l'esistenza' porta alla sostituzione di un tipo con un altro, ma l'importanza di questa sostituzione non è misurabile in termini di riproduttività media della popolazione nel suo insieme.
7. Adattamento ed estinzione
Il fatto che il valore assoluto di W̅̅ sia uguale all'unità prima e dopo la selezione non significa che non accadrà nulla che influirà sul futuro della specie. Tutte le specie, quando si trovano in condizioni ambientali estremamente sfavorevoli, sono soggette ad estinzione. In effetti, più del 99,9% di tutte le specie esistite si sono estinte. È stato suggerito da J. M. Thoday che si potrebbe misurare l'idoneità biologica di una popolazione o di una specie dalla probabilità che essa non sia ancora estinta in un lontano tempo futuro.
Non vi è dubbio che i cambiamenti nelle proprietà demografiche, che si sono avuti nel processo selettivo, influiranno sulla probabilità di estinzione, ma non è vero che questa probabilità venga sempre o anche generalmente ridotta dalla selezione. Nel caso di una mutazione che aumenta la fertilità senza far variare l'efficacia di utilizzazione delle risorse, le dimensioni della popolazione saranno ridotte da variazioni climatiche estreme che fanno diminuire la schiusa delle uova o la fecondità e longevità degli adulti in maniera minore di quanto non avvenisse prima del cambiamento evolutivo. D'altra parte, vi sarà una maggiore sensibilità ad una scarsezza di cibo da parte degli individui più giovani, poiché più grande sarà il numero d'individui ai primi stadi dell'esistenza in competizione per una fonte di sussistenza presente in piccola quantità. In questo ultimo caso, l'aumento della fecondità potrebbe portare in effetti alla estinzione, poiché pochissimi piccoli potrebbero sopravvivere nella lotta per le risorse insufficienti. Il succedersi di ambienti così radicalmente diversi fra loro sarebbe pertanto fatale per le specie. Nel caso di una mutazione che generasse un aumento nell'efficacia dell'utilizzazione di risorse, ma non un cambiamento nella fecondità, la nuova popolazione prodotta dall'evoluzione sarebbe più sicura di prima rispetto a fluttuazioni che rendessero le risorse insufficienti, ma la sua maggiore densità la renderebbe più suscettibile a malattie epidemiche e più appetibile per i predatori, il cui istinto di caccia ben si adatta alla frequenza della preda. In generale, la probabilità di morte di un individuo per predazione, infestazione parassitaria e malattie infettive aumenta in funzione della densità.
Non si deve ritenere che la quantità di difficoltà ambientali, in cui una specie generalmente si viene a trovare, possa causare in qualche maniera l'accumulo di quei cambiamenti selettivi che diminuiscono la probabilità di estinzione. Anche se si è spesso verificata una grave diminuzione di cibo per gli individui più giovani, il meccanismo della selezione all'interno di una popolazione potrà sempre causare un aumento di frequenza di un gene che accresca la fecondità, anche se quest'aumento la renderà una popolazione più facilmente estinguibile per l'affollamento dei piccoli. Non possiamo mettere in evidenza, nel processo della selezione naturale fra singoli individui, alcun meccanismo che possa far aumentare la longevità di popolazioni o specie in generale, benché ciò possa avvenire ad opera di cambiamenti evolutivi particolari. L'unico meccanismo che potrebbe far diminuire la probabilità di estinzione di gruppi è la selezione fra gruppi o selezione interdemica. Per esempio, in una specie che sia soggetta spesso ad una forte diminuzione di cibo per i piccoli, una popolazione con alta fecondità si estinguerà, in media, prima di una con bassa fecondità, cosicché per un processo storico di estinzione e ricolonizzazione degli habitat da parte delle popolazioni ancora esistenti, l'intera specie può infine risultare caratterizzata da una bassa fecondità. In questa maniera, dei geni che dovrebbero diffondersi per selezione naturale all'interno di una popolazione possono essere eliminati dalla specie per selezione fra popolazioni. Si potrebbe parlare allora di un adattamento di popolazione o di specie contrario alle leggi della selezione naturale.
Numerosi ‛adattamenti' di specie sono stati attribuiti alla selezione interdemica, poiché non sembrano spiegabili come prodotto logico della selezione naturale che agisce su singoli individui. Ciò è apparso evidente per diverse interazioni sociali, nelle quali singoli individui si sacrificano per il gruppo, come i nuclei di allarme formati dagli uccelli o le cure parentali, dato che si presume sia di diretto svantaggio per un individuo richiamare su di sé l'attenzione del predatore o cedere parte del suo cibo ad un altro individuo. Con la selezione fra gruppi vengono anche spiegati cambiamenti nel sistema genetico, come ad esempio variazioni del ritmo di ricombinazione, poiché è stato osservato che il vantaggio di una variazione di questo tipo è quello di aumentare al massimo la probabilità di produrre un nuovo genotipo favorevole in un nuovo ambiente, cosicché la popolazione possa sopravvivere, benché non vi sia assolutamente alcun vantaggio per l'individuo che produce i gameti ricombinati. È stato anche ipotizzato che la selezione fra gruppi ridurrà il tasso di sfruttamento delle risorse da parte di una popolazione, in modo che essa si mantenga su di un valore numerico ‛ottimale'. Un'accurata analisi di questi ed altri casi di adattamento di gruppo compiuta da Williams ha dimostrato che essi non possono reggere ad un attento esame critico. Risultò che la maggior parte erano normali casi di selezione naturale tra singoli individui, quando ci si ricordò che la teoria genetica della selezione naturale prevede l'aumento dei genotipi basato sulla riproduzione differenziale dei geni. Così, benché un genitore possa sacrificare parte della propria idoneità biologica per far aumentare quella della sua prole, il suo genotipo non ha perso in idoneità biologica, poiché la prole porta i geni dei propri genitori. In generale, un individuo può sacrificare una qualunque parte della propria idoneità biologica a favore di un altro individuo, purché sia inferiore alla sua correlazione genetica con questo, e riuscire ad avere nonostante ciò un aumento netto nella frequenza del suo genotipo. È anche possibile dimostrare che la frazione ricombinante evolverà, di fatto, in una popolazione col meccanismo normale della selezione naturale. D'altra parte, l'evoluzione dei meccanismi di regolazione di una popolazione che mantengano le dimensioni entro valori ‛ottimali', sembra essere un'illusione piuttosto che un fenomeno da dover spiegare. Le popolazioni hanno dei valori medi e fluttuano per quel che riguarda il numero da anno ad anno. Non vi è alcun metodo oggettivo per determinare se una popolazione sarebbe in qualche maniera senz'altro favorita se il suo valore numerico medio fosse inferiore e dovesse subire minori fluttuazioni. Fa parte del pregiudizio comune alla scienza occidentale del XX secolo credere che mancanza di fluttuazione e valori stabili significhino qualcosa di positivo.
Vi sono alcuni casi ben documentati di estinzione di popolazioni che hanno comportato un adattamento. Uno di questi, esemplificativo di una classe di fenomeni, è l'evoluzione del Virus Mixoma che infetta i conigli in Australia. Dopo l'introduzione di questo virus per controllare la diffusione dei conigli, si era notato che esso evolveva verso uno stato temperato, non patogeno. La spiegazione è che un virus patogeno causerà la morte del coniglio e quindi la morte dell'intera popolazione virale all'interno del coniglio. Poiché la malattia viene diffusa dalle zanzare, che pungono solo conigli viventi, le popolazioni di virus che contengono alcune particelle letali non riusciranno a colonizzare altri conigli, mentre quelle popolazioni che sono completamente non-letali riusciranno a diffondersi.
Il caso del coniglio e del Virus Mixoma illustra un problema teorico fondamentale, indicando l'estinzione come una delle forze principali che permettono l'adattamento. Ogni coniglio è un deme per il virus, ed in Australia vi sono milioni di questi demi. La mortalità dei conigli è molto alta, come è alta la diffusione dell'infezione da parte delle zanzare; così pure la velocità di estinzione e ricolonizzazione dei demi è molto alta. In tali circostanze, la selezione interdemica può agire con una forza paragonabile a quella della selezione naturale fra singoli individui, il cui indice è controllato dai valori di natalità e di mortalità degli individui. Per la maggior parte delle popolazioni di quasi tutte le specie, tuttavia, le probabilità di estinzione e ricolonizzazione di intere popolazioni per unità di tempo è molto inferiore ai valori di natalità e mortalità di singoli individui all'interno delle popolazioni. In queste circostanze, la selezione interdemica è molto più debole della selezione all'interno delle popolazioni, cosicché ci si aspetta di trovare le specie caratterizzate dai geni favoriti dalle leggi normali della selezione naturale (v. Lewontin, 1970).
Un'importante prova del fatto che l'evoluzione non agisce come una forza per l'adattamento nel senso di far diminuire la percentuale di estinzione, deriva dall'esame oggettivo delle probabilità di estinzione nei diversi gruppi tassonomici. È ben noto che le probabilità di estinzione variano considerevolmente da un gruppo tassonomico ad un altro e da un'epoca storica ad un'altra (v. Simpson, 1944). Se, tuttavia, si calcola dai reperti fossili la probabilità di estinzione di un gruppo per unità di tempo in un'era geologica, risulta che essa è indipendente dall'età di tale gruppo (v. Van Valen, 1973). Cioè, un genere di Brachiopodi articolati, che è presente da 10 milioni di anni, ha le medesime probabilità di sopravvivere per altri 10 milioni di anni di un genere che è già vissuto per 80 milioni di anni. I generi che sono vissuti a lungo sono semplicemente quelli che, per un caso, non si sono ancora estinti, ma non vi è nulla nel loro equilibrio naturale che li ha predisposti alla longevità. Essi non hanno delle capacità di adattamento superiori alla media.
8. Altri criteri di adattamento di popolazioni
Il valore assoluto dell'idoneità biologica media, W̅̅ , e la probabilità di estinzione di una popolazione non sembrano misure soddisfacenti dei cambiamenti nell'adattamento che avvengono per selezione naturale. In un tentativo di quantificare gli effetti della selezione naturale, sono state utilizzate molte altre misure demografiche. Le due più comuni derivano dal semplice modello della crescita di una popolazione. Se assumiamo che un habitat è stato colonizzato da poco da una specie, la crescita di numero è dapprima rapida, ma poi, quando è stata esaurita la risorsa limitante, la popolazione si livella completamente ad una certa densità di saturazione. La descrizione matematica più semplice di una crescita di popolazione di questo tipo è la classica equazione di Lotka-Volterra
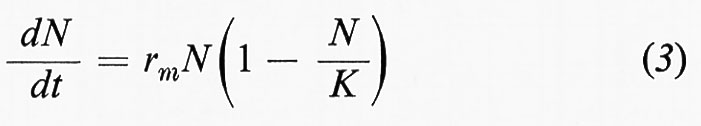
dove N è il valore numerico della popolazione, rm è il tasso intrinseco di accrescimento, cioè il tasso di crescita della popolazione durante la prima fase in cui la densità è assai bassa, e K è la capacità di carico, il numero di saturazione che la popolazione può infine raggiungere (v. Andrewartha e Birch, 1954). È ragionevole pensare che o il tasso iniziale di crescita, rm, senza limiti, o il valore numerico totale sopportabile dall'ambiente, K, possano aumentare per adattamento se la nozione di adattamento ha qualche valore. Sono stati eseguiti un certo numero di esperimenti con popolazioni sia polimorfiche sia monomorfiche di Drosophila nel tentativo di mostrare una maggiore idoneità biologica delle popolazioni polimorfiche, poiché queste permangono in un equilibrio genetico stabile sotto la pressione della selezione naturale. Sia K che rm sembrano essere superiori per popolazioni polimorfiche che si trovino in condizioni di stress, ma in condizioni ambientali migliori vi erano piccole differenze nel valore di rm (per una rassegna v. Dobzhansky, 1968). F. J. Ayala ha trovato che l'introduzione di nuove mutazioni in una popolazione mediante irradiazione porta, dopo molte generazioni, ad un valore di K superiore, benché inizialmente le dimensioni delle singole popolazioni fossero inferiori a quelle dei controlli: effetto, questo, dovuto chiaramente alle mutazioni letali e semiletali indotte dall'irradiazione, ma eliminate alla fine. Vi sono finora dati insufficienti per poter dire se l'aumento di K o rm sia un fenomeno generale, risultato della selezione naturale.
9. Adattabilità
Nel mondo naturale, l'ambiente in cui gli organismi vivono varia notevolmente nel tempo, cosicché varia anche l'idoneità biologica dei vari genotipi. Per un certo periodo alcuni genotipi possono aumentare la loro frequenza per le loro migliori possibilità di sopravvivenza in un determinato ambiente, ma se poi l'ambiente viene alterato in maniera drastica, allora alcuni genotipi potranno difficilmente sopravvivere e il valore medio d'idoneità biologica assoluta può scendere al di sotto dell'unità, cosicché diminuirà anche il numero degli individui. Nello stesso tempo, cominceranno ad aumentare nella popolazione dei genotipi più adatti, ma non è assolutamente certo che il valore adattativo assoluto medio raggiunga nuovamente l'unità prima che la popolazione sia estinta. Un aspetto caratteristico della velocità di cambiamento della frequenza genica, come è mostrato nell'equazione simile alla (1)
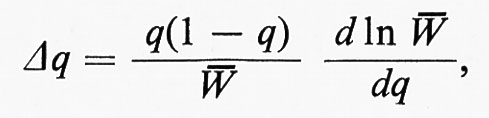
è quello di dipendere non solo dall'inclinazione della funzione dell'idoneità biologica, che può essere considerata come un valore di forza, ma anche dal prodotto q (1−q), una specie di valore d'inerzia. Se un allele è quasi fissato nella popolazione (q o 1−q prossimi allo zero), il cambiamento di frequenza genica sarà assai piccolo a prescindere da quanto forte sia la selezione. Si supponga che un ambiente sia rimasto in condizioni piuttosto uniformi per un lungo periodo, cosicché la selezione abbia portato uno degli alleli ad una frequenza assai prossima allo zero; se successivamente viene invertita la direzione di selezione, la popolazione reagirà molto lentamente, perché q (1−q) è piccolo. Per lungo tempo la composizione genotipica della popolazione rifletterà il lungo periodo precedente di selezione uniforme, e non rifletterà la situazione attuale. Questo fenomeno della popolazione, risultando fuori fase rispetto all'ambiente, può avere serie conseguenze per la sua sopravvivenza e sarà soprattutto evidente in ambienti con lunghi cicli cronologici.
L'effetto può essere ridotto, paradossalmente, diminuendo la rispondenza generale della frequenza genica alla selezione, cosicché anche dopo lunghi periodi di condizioni ambientali uniformi la composizione genetica della popolazione non sarà stata ‛selezionata' in un particolare genotipo che può poi essere quasi letale, quando cambiano le condizioni ambientali.
Il problema di quale sia il sistema genetico ottimale per conservare valori di idoneità biologica assoluta media in condizioni ambientali fluttuanti, il problema dell' ‛adattabilità', è stato studiato teoricamente da R. Levins (v., 1968), ma esso deve essere ancora in gran parte indagato e analizzato.
Pertanto, il problema reale degli studi evolutivi non è se la selezione naturale porti o meno all'adattamento, ma quali sono le condizioni di adattabilità che fanno ritardare l'estinzione di popolazioni e specie al variare delle condizioni ambientali.
bibliografia
Andrewartha, H. C., Birch, L. C., The distribution and abundance of animals, Chicago 1954.
Ayala, F. J., Evolution of fitness. III. Improvement of fitness in irradiated populations of Drosophila serrata, in ‟Proceedings of the National Academy of Sciences", 1967, LVIII, pp. 1919-1923.
Darwin, Ch., On the origin of species, London 1859.
Dobzhansky, Th., On some fundamental concepts of Darwinian biology, in ‟Evolutionary biology", 1968, II, pp. 1-34.
Fisher, R. A., The genetical theory of natural selection, Oxford 1930.
Ford, E. B., Ecological genetics, London 1964.
Harris, H., The principles of human biochemical genetics, Amsterdam 1970.
Leigh, E., Adaptation and diversity, New York 1971.
Levins, R., Evolution in changing environments, Princeton 1968.
Lewontin, R. Ch., The units of selection, in ‟Annual review of systematics and ecology", 1970, I, pp. 1-18.
Lewontin, R. Ch., The genetical basis of evolutionary change, New York 1973.
Lewontin, R. Ch., Hubby, J. L., A molecular approach to the study of genic heterozygosity in natural populations. II. Amount of variation and degree of heterozygosity in natural populations of Drosophila pseudoobscura, in ‟Genetics", 1966, LIV, pp. 595-609.
Simpson, G. G., Tempo and mode in evolution, New York 1944.
Stern, J. T. Jr., The meaning of ‛adaptation' and its relation to the phenomenon of natural selection, in ‟Evolutionary biology", 1970, IV, pp. 39-66.
Thoday, J. M., Components of fitness, in ‟Symposium of the Society for Experimental Biology", 1953, VII, pp. 96-113.
Van Valen, L., A new evolutionary law, in ‟Evolutionary theory", 1973, I, pp. 1-30.
Williams, G. C., Adaptation and natural selection, Princeton 1966.
Wright, S., Evolution in Mendelian populations, in ‟Genetics", 1931, XVI, pp. 97-159.